

Carotenoids in Skin Photoaging: Unveiling Protective Effects, Molecular Insights, and Safety and Bioavailability Frontiers

 , ,
, ,
Abstract
1. Introduction
2. Skin Photoaging
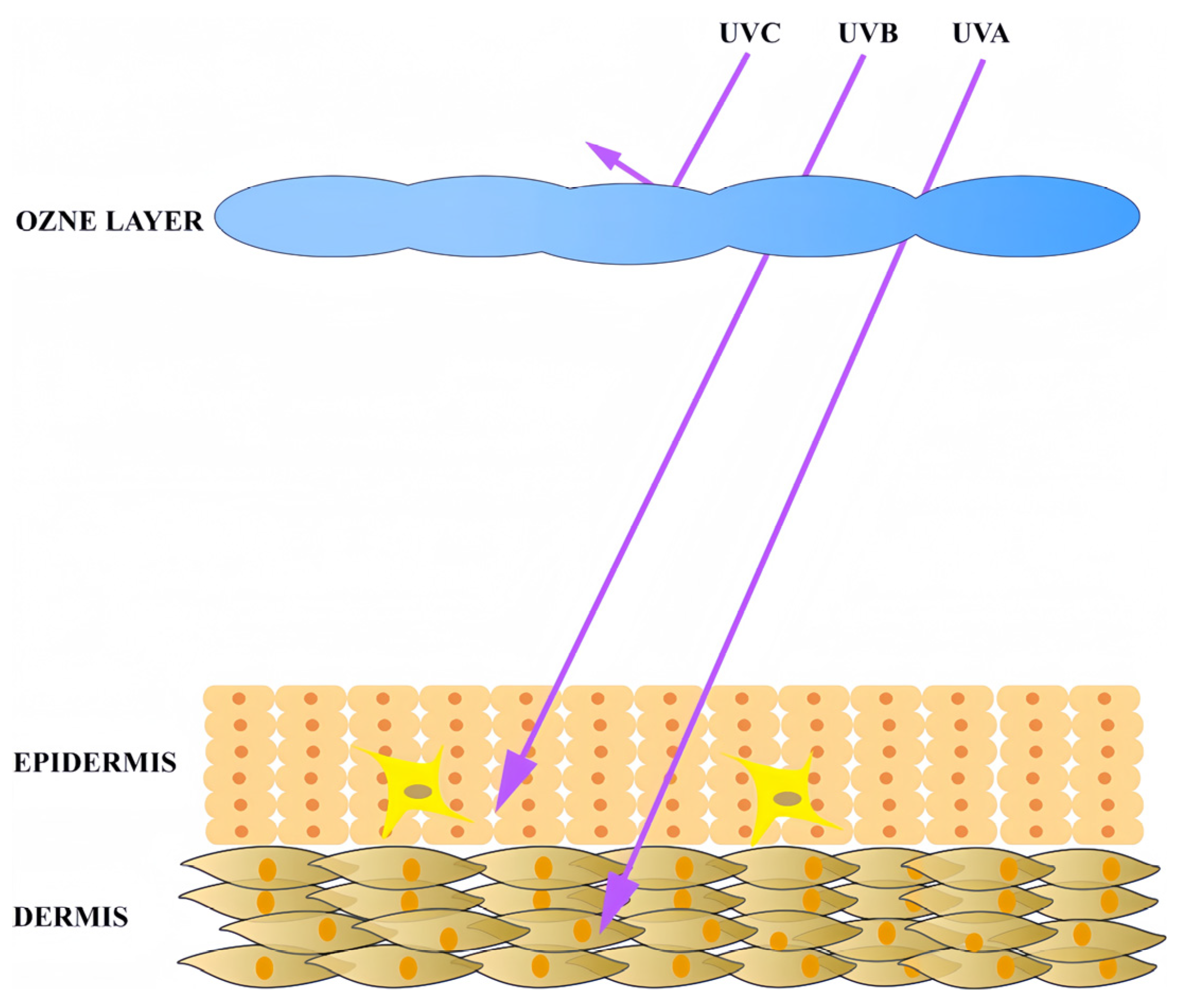
2.1. Ultraviolet Radiation as the Primary Trigger of Skin Photoaging
2.2. Skin Photoaging and Oxidative Stress
2.2.1. Bio-Macromolecule Damage by Oxidative Stress
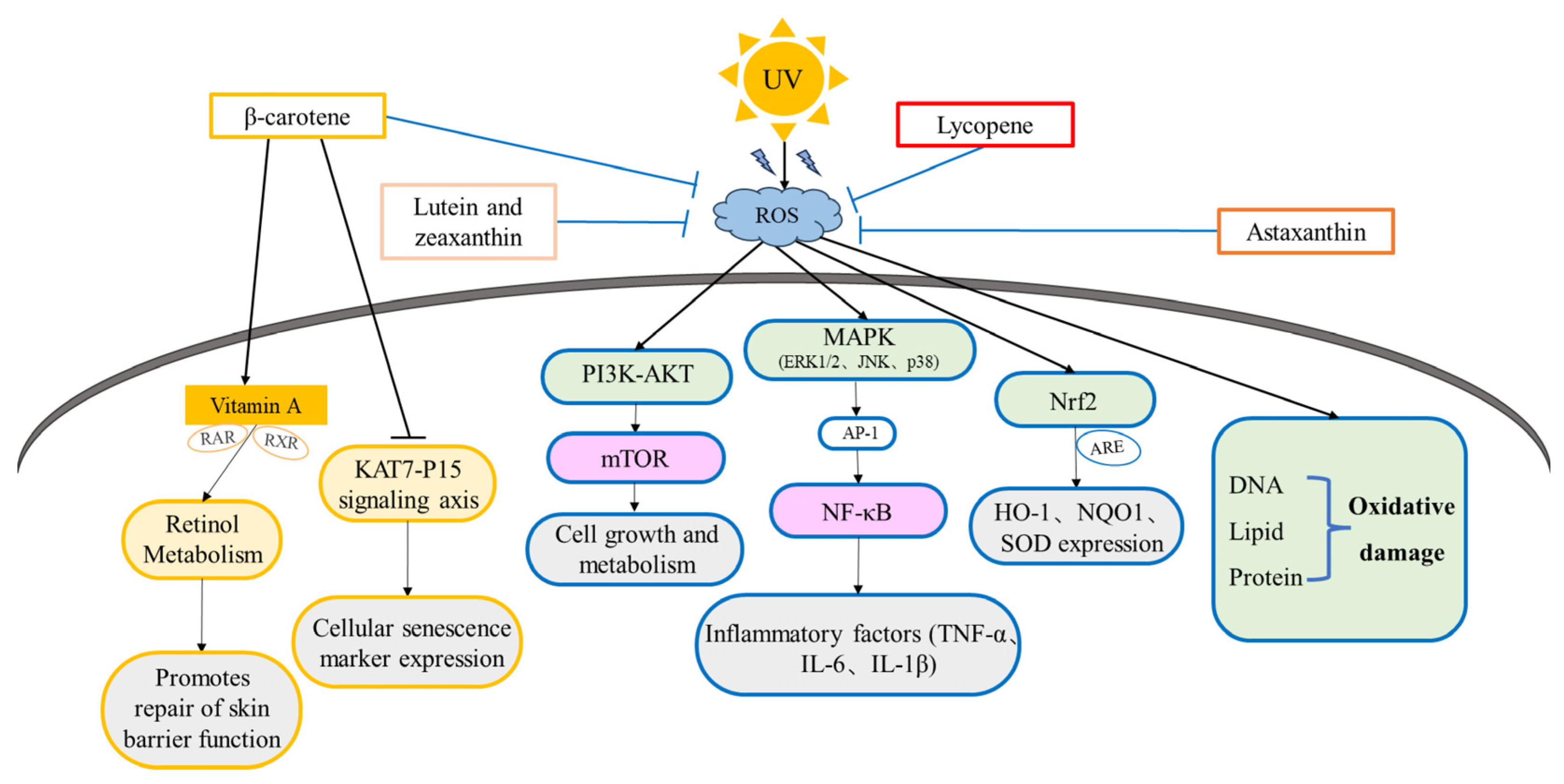
2.2.2. Signal Pathways Activated by Oxidative Stress
3. The Anti-Photoaging Effect and Mechanism of Carotenoids
3.1. The Anti-Photoaging Effect and Mechanism of C40 Carotenoids
3.1.1. The Anti-Photoaging Effects and Mechanism of β-Carotene
3.1.2. The Anti-Photoaging Effects and Mechanism of Lycopene
3.1.3. The Anti-Photoaging Effects and Mechanism of Astaxanthin
3.1.4. The Anti-Photoaging Effects and Mechanism of Lutein and Zeaxanthin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Antioxidant Capacity | In Vitro Experiment | In Vivo Experiment | Signal Pathways | References |
|---|---|---|---|---|---|
| β-carotene | Singlet oxygen (all-E-isomer-rich, IC50 0.38 μg/mL; Z-isomer-rich IC50 = 0.95 μg/mL) | Human skin fibroblasts (HSFs), human229 neonatal skin fibroblasts (NB1RGB), and B16274 mouse melanoma cells: enhancing hyaluronic acid production, promoting proliferation, anti-elastase activity, anti-melanogenic activity and anti-tyrosinase activity, inhibition of type I collagen production, and inhibition of melanin precursor darkening | - | - | [130] |
| - | Keratinocyte: inhibition of UVA-induced ECM degradation and enhancement of UVA-induced expression of tanning-related protease-activated receptor 2, promotes cell differentiation | - | - | [117,118] | |
| - | Mesenchymal stem cells (MSCs): reducing the expression of cellular senescence markers (e.g., SA-β-gal, p21, p53), enhancing cellular antioxidant capacity, and reducing oxidative stress-induced cell damage | C57 mice: improving the aging state of many tissues and organs, reducing expression of inflammatory factors | KAT7-P15 | [116] | |
| - | Human mammary cancer cells (MCF-7) and human hepatocellular carcinoma cells (HepG 2): activating ARE, inducing the expression of antioxidant enzymes and phase II detoxification enzymes, and enhancing cellular antioxidant and detoxification capacity | - | Nrf2/ARE | [113] | |
| - | Rat Small Intestine Crypt Epithelial Cells (IEC): Down-regulation of caspase-3, Bax levels and LC3II/I ratio, and up-regulation of Bcl-2 and p62 levels were used to reduce autophagy and inhibit apoptosis | - | PI3K/AKT/mTOR | [114] | |
| - | - | Mice: decreasing malondialdehyde, TNF-α and IL-6 levels, and increasing glutathione peroxidase and superoxide dismutase levels | NF-κB/MAPK/Nrf2 | [115] | |
| - | - | Healthy female subjects: improving facial wrinkles and elasticity, increases collagen type I mRNA levels, reduces UV-induced DNA damage | - | [105] | |
| - | - | Hairless mice: inhibiting MMP-9 expression and reducing skin wrinkles and sagging | - | [107] | |
| - | - | 11 male and 11 female subjects: protecting human skin from UVA and UVB-induced erythema, reducing serum lipid peroxidation | - | [106] | |
| Lycopene | Singlet oxygen (all-E-isomer-rich IC50 0.26 μg/mL, Z-isomer-rich IC50 1.06 μg/mL) | HSF: Enhancing hyaluronic acid production, promoting proliferation | Anti-elastase activity, anti-melanogenic activity, and anti-tyrosinase activity, inhibition of melanin precursor darkening | - | [130] |
| - | - | Human oral intake: reducing erythematous reaction | - | [131] | |
| - | Chinese hamster ovary cell (M146L cell): reduction in oxidative stress and apoptosis, upregulation of antioxidant and anti-apoptotic proteins, downregulation of pro-apoptotic proteins | - | PI3K/Akt/Nrf2 | [139] | |
| - | Macrophages: inhibiting LPS-induced IκB phosphorylation, IκB degradation, and NF-κB translocation, blocking phosphorylation of ERK1/2 and p38 MAP kinase | - | MAPK/NF-κB | [140] | |
| - | Primary mouse neurons cell: enhancing cell viability, restoring mitochondrial membrane potential, and reducing ROS production | - | PI3K/Akt | [138] | |
| - | HSF: decreasing the content of ROS, β-galactosidases, and AGEs and increases mitochondrial membrane potential | - | - | [128] | |
| - | Human neuroblastoma cells (SH-SY5Y): blocking neuro-inflammation and apoptosis | - | - | [129] | |
| - | HSF: Combined with vitamin E, enhancing the inhibition of MMP-1 expression after UVA radiation | - | - | [137] | |
| - | - | 60 female subjects: improving skin elasticity, firmness, brightness, tone, and fine lines and wrinkles | - | [132] | |
| - | - | 5 male and 5 female subjects: reduction in markers of inflammatory oxidative damage (e.g., malondialdehyde, protein carbonyls, etc.) and low-density lipoprotein peroxidase protein levels | - | [133] | |
| - | - | 33 healthy male volunteers aged 20 to 30 years old: enhancing skin hydration and elasticity, reducing erythema, melanin, and sebum levels | - | [134] | |
| - | - | 20 volunteers between 40 and 50 years of age: reducing the number of wrinkles and roughness | - | [135] | |
| Astaxanthin | ABTS(IC50 7.7 μg/mL); DPPH(IC50 17.5 μg/mL) | - | - | - | [161] |
| ABTS(IC50 17.56 μg/mL); DPPH(IC50 50.93 μg/mL) | - | - | - | [162] | |
| - | HSF: inhibiting cellular damage caused by free radicals and reducing UVA radiation-induced elevation of IL6 expression | - | - | [151] | |
| - | Mouse photoreceptor cells (661W): reducing ROS production and attenuating apoptosis | - | PI3K/Akt/Nrf2 | [153] | |
| - | - | Male hairless mice: reducing UV-induced collagen degradation and elastic fiber damage, reducing the expression of MMPs (e.g., MMP-1, MMP-3, MMP-9) | Nrf2/NF-κB/MAPK | [154] | |
| - | HaCaT keratinocytes: inhibition of cell apoptosis by reducing INOS and COX-2 | - | - | [152] | |
| - | - | Between 30 and 56 years of age, 21 women and 2 men: improving skin elasticity and hydration | - | [149] | |
| - | - | 30 healthy female subjects: improving wrinkles, elasticity, transepidermal water loss, moisture content, and sebum oil levels | - | [150] | |
| Lutein and zeaxanthin | - | Human RPE cell: directly quenching ROS and facilitating glutathione synthesis | - | - | [156] |
| - | Rat tracheal epithelial cells: reduction in UVA radiation-induced DNA damage | - | - | [157] | |
| - | - | Human subjects: improving skin hydration, elasticity, and photoprotective activity | - | [158] | |
| - | - | Hairless mice: decreasing UVB-induced epidermal hyper-proliferation and acute inflammation in hairless mice | - | [159] | |
| - | - | Mice: reducing wrinkles and dryness | - | [160] |
3.2. The Potential of Novel Carotenoids on Anti-Photoaging
3.2.1. C30 Carotenoids
3.2.2. C50 Carotenoids
4. Safety and Bioavailability of Carotenoids
4.1. Oral Administration: Safety Considerations
4.2. Topical Application: Efficacy and Safety Profile
5. Conclusions and Outlooks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Millar, S.E. Revitalizing Aging Skin through Diet. Cell 2018, 175, 1461–1463. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Duan, E. Fighting against Skin Aging: The Way from Bench to Bedside. Cell Transplant. 2018, 27, 729. [Google Scholar] [CrossRef] [PubMed]
- Tsatsou, F.; Trakatelli, M.; Patsatsi, A.; Kalokasidis, K.; Sotiriadis, D. Extrinsic Aging. Dermato-Endocrinology 2012, 4, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.H.; Chien, A.L. Photoaging: A Review of Current Literature. Curr. Dermatol. Rep. 2020, 9, 22–29. [Google Scholar] [CrossRef]
- Kammeyer, A.; Luiten, R.M. Oxidation Events and Skin Aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Amaro-Ortiz, A.; Yan, B.; D’Orazio, J. Ultraviolet Radiation, Aging and the Skin: Prevention of Damage by Topical cAMP Manipulation. Molecules 2014, 19, 6202–6219. [Google Scholar] [CrossRef]
- Gendrisch, F.; Esser, P.R.; Schempp, C.M.; Wölfle, U. Luteolin as a Modulator of Skin Aging and Inflammation. BioFactors 2021, 47, 170–180. [Google Scholar] [CrossRef]
- Daré, R.G.; Nakamura, C.V.; Ximenes, V.F.; Lautenschlager, S.O.S. Tannic Acid, a Promising Anti-Photoaging Agent: Evidences of Its Antioxidant and Anti-Wrinkle Potentials, and Its Ability to Prevent Photodamage and MMP-1 Expression in L929 Fibroblasts Exposed to UVB. Free. Radic. Biol. Med. 2020, 160, 342–355. [Google Scholar] [CrossRef]
- Shin, S.; Cho, S.H.; Park, D.; Jung, E. Anti-skin Aging Properties of Protocatechuic Acid in Vitro and in Vivo. J. Cosmet. Dermatol. 2020, 19, 977–984. [Google Scholar] [CrossRef]
- Yutani, R.; Kikuchi, T.; Teraoka, R.; Kitagawa, S. Efficient Delivery and Distribution in Skin of Chlorogenic Acid and Resveratrol Induced by Microemulsion Using Sucrose Laurate. Chem. Pharm. Bull. 2014, 62, 274–280. [Google Scholar] [CrossRef]
- Galano, A.; Álvarez-Diduk, R.; Ramírez-Silva, M.T.; Alarcón-Ángeles, G.; Rojas-Hernández, A. Role of the Reacting Free Radicals on the Antioxidant Mechanism of Curcumin. Chem. Phys. 2009, 363, 13–23. [Google Scholar] [CrossRef]
- Ciardullo, G.; Orlando, C.; Russo, N.; Marchese, E.; Galano, A.; Marino, T.; Prejanò, M. On the Dual Role of (+)-Catechin as Primary Antioxidant and Inhibitor of Viral Proteases. Comput. Biol. Med. 2024, 180, 108953. [Google Scholar] [CrossRef] [PubMed]
- Grivard, A.; Goubet, I.; Duarte Filho, L.M.D.S.; Thiéry, V.; Chevalier, S.; de Oliveira-Junior, R.G.; El Aouad, N.; Guedes da Silva Almeida, J.R.; Sitarek, P.; Quintans-Junior, L.J.; et al. Archaea Carotenoids: Natural Pigments with Unexplored Innovative Potential. Mar. Drugs 2022, 20, 524. [Google Scholar] [CrossRef]
- Dias, M.G.; Olmedilla-Alonso, B.; Hornero-Méndez, D.; Mercadante, A.Z.; Osorio, C.; Vargas-Murga, L.; Meléndez-Martínez, A.J. Comprehensive Database of Carotenoid Contents in Ibero-American Foods. A Valuable Tool in the Context of Functional Foods and the Establishment of Recommended Intakes of Bioactives. J. Agric. Food Chem. 2018, 66, 5055–5107. [Google Scholar] [CrossRef]
- Lu, W.; Shi, Y.; Wang, R.; Su, D.; Tang, M.; Liu, Y.; Li, Z. Antioxidant Activity and Healthy Benefits of Natural Pigments in Fruits: A Review. Int. J. Mol. Sci. 2021, 22, 4945. [Google Scholar] [CrossRef]
- Khan, U.M.; Sevindik, M.; Zarrabi, A.; Nami, M.; Ozdemir, B.; Kaplan, D.N.; Selamoglu, Z.; Hasan, M.; Kumar, M.; Alshehri, M.M.; et al. Lycopene: Food Sources, Biological Activities, and Human Health Benefits. Oxidative Med. Cell. Longev. 2021, 2021, 2713511. [Google Scholar] [CrossRef] [PubMed]
- Rossi, N.; Grosso, C.; Delerue-Matos, C. Shrimp Waste Upcycling: Unveiling the Potential of Polysaccharides, Proteins, Carotenoids, and Fatty Acids with Emphasis on Extraction Techniques and Bioactive Properties. Mar. Drugs 2024, 22, 153. [Google Scholar] [CrossRef]
- Gallego, R.; Valdés, A.; Suárez-Montenegro, Z.J.; Sánchez-Martínez, J.D.; Cifuentes, A.; Ibáñez, E.; Herrero, M. Anti-Inflammatory and Neuroprotective Evaluation of Diverse Microalgae Extracts Enriched in Carotenoids. Algal Res. 2022, 67, 102830. [Google Scholar] [CrossRef]
- Bolhassani, A.; Khavari, A.; Bathaie, S.Z. Saffron and Natural Carotenoids: Biochemical Activities and Anti-Tumor Effects. Biochim. Et Biophys. Acta (Bba)-Rev. Cancer 2014, 1845, 20–30. [Google Scholar] [CrossRef]
- Koklesova, L.; Liskova, A.; Samec, M.; Zhai, K.; Abotaleb, M.; Ashrafizadeh, M.; Brockmueller, A.; Shakibaei, M.; Biringer, K.; Bugos, O.; et al. Carotenoids in Cancer Metastasis—Status Quo and Outlook. Biomolecules 2020, 10, 1653. [Google Scholar] [CrossRef]
- Satomi, Y. Antitumor and Cancer-Preventative Function of Fucoxanthin: A Marine Carotenoid. Anticancer. Res. 2017, 37, 1557–1562. [Google Scholar] [CrossRef]
- Hirakida, H.; Nakamura, S.; Inagaki, S.; Tsuji, S.; Hayashi, M.; Shimazawa, M.; Hara, H. Anti-Diabetic Effects of Astaxanthin-Rich Extract Derived from Paracoccus carotinifaciens on Pancreatic β Cells. J. Funct. Foods 2022, 97, 105252. [Google Scholar] [CrossRef]
- Zununi Vahed, S.; Zuluaga Tamayo, M.; Rodriguez-Ruiz, V.; Thibaudeau, O.; Aboulhassanzadeh, S.; Abdolalizadeh, J.; Meddahi-Pellé, A.; Gueguen, V.; Barzegari, A.; Pavon-Djavid, G. Functional Mechanisms of Dietary Crocin Protection in Cardiovascular Models under Oxidative Stress. Pharmaceutics 2024, 16, 840. [Google Scholar] [CrossRef]
- Maoka, T. Recent Progress in Structural Studies of Carotenoids in Animals and Plants. Arch. Biochem. Biophys. 2009, 483, 191–195. [Google Scholar] [CrossRef]
- Naik, A.A.; Gadgoli, C.H.; Naik, A.B. Formulation Containing Phytosomes of Carotenoids from Nyctanthes Arbor-Tristis and Tagetes Patula Protect D-Galactose Induced Skin Aging in Mice. Clin. Complement. Med. Pharmacol. 2023, 3, 100070. [Google Scholar] [CrossRef]
- Imokawa, G. The Xanthophyll Carotenoid Astaxanthin Has Distinct Biological Effects to Prevent the Photoaging of the Skin Even by Its Postirradiation Treatment. Photochem. Photobiol. 2019, 95, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G. Intracellular Signaling Mechanisms Involved in the Biological Effects of the Xanthophyll Carotenoid Astaxanthin to Prevent the Photo-Aging of the Skin in a Reactive Oxygen Species Depletion-Independent Manner: The Key Role of Mitogen and Stress-Activated Protein Kinase 1. Photochem. Photobiol. 2019, 95, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet Radiation-Induced Skin Aging: The Role of DNA Damage and Oxidative Stress in Epidermal Stem Cell Damage Mediated Skin Aging. Stem Cells Int. 2016, 2016, 7370642. [Google Scholar] [CrossRef]
- Amer, R.I.; Ezzat, S.M.; Aborehab, N.M.; Ragab, M.F.; Mohamed, D.; Hashad, A.; Attia, D.; Salama, M.M.; El Bishbishy, M.H. Downregulation of MMP1 Expression Mediates the Anti-Aging Activity of Citrus sinensis Peel Extract Nanoformulation in UV Induced Photoaging in Mice. Biomed. Pharmacother. 2021, 138, 111537. [Google Scholar] [CrossRef]
- Gromkowska-Kępka, K.J.; Puścion-Jakubik, A.; Markiewicz-Żukowska, R.; Socha, K. The Impact of Ultraviolet Radiation on Skin Photoaging—Review of in Vitro Studies. J. Cosmet. Dermatol. 2021, 20, 3427–3431. [Google Scholar] [CrossRef]
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New Insights in Photoaging, UVA Induced Damage and Skin Types. Exp. Dermatol. 2014, 23, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Guo, K.; Chen, Y.; Zhao, J.; Jing, R.; Wang, L.; Li, X.; Hu, Z.; Xu, N.; Li, X. Keratinocyte Growth Factor 2 Ameliorates UVB-Induced Skin Damage via Activating the AhR/Nrf2 Signaling Pathway. Front. Pharmacol. 2021, 12, 655281. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, R.; Kanimozhi, G.; Prasad, N.R.; Agilan, B.; Ganesan, M.; Srithar, G. Alpha Pinene Modulates UVA-Induced Oxidative Stress, DNA Damage and Apoptosis in Human Skin Epidermal Keratinocytes. Life Sci. 2018, 212, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Q.; Travers, J.B.; Kemp, M.G. Roles of UVA Radiation and DNA Damage Responses in Melanoma Pathogenesis. Environ. Mol. Mutagen. 2018, 59, 438–460. [Google Scholar] [CrossRef]
- Shin, J.-W.; Kwon, S.-H.; Choi, J.-Y.; Na, J.-I.; Huh, C.-H.; Choi, H.-R.; Park, K.-C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef]
- Lai-Cheong, J.E.; McGrath, J.A. Structure and Function of Skin, Hair and Nails. Medicine 2021, 49, 337–342. [Google Scholar] [CrossRef]
- Mu, J.; Chen, H.; Ye, M.; Zhang, X.; Ma, H. Acacetin Resists UVA Photoaging by Mediating the SIRT3/ROS/MAPKs Pathway. J. Cell Mol. Med. 2022, 26, 4624–4628. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Syafitri, S.M.; Geraldine, B.A.F.D.; Hamdin, C.D.; Frediansyah, A.; Miyake, M.; Kobayashi, D.; Hazama, A.; Sunarpi, H. UVA Photoprotective Activity of Brown Macroalgae Sargassum Cristafolium. Biomedicines 2019, 7, 77. [Google Scholar] [CrossRef]
- Lohakul, J.; Chaiprasongsuk, A.; Jeayeng, S.; Saelim, M.; Muanjumpon, P.; Thanachaiphiwat, S.; Tripatara, P.; Soontrapa, K.; Lumlerdkij, N.; Akarasereenont, P.; et al. The Protective Effect of Polyherbal Formulation, Harak Formula, on UVA-Induced Photoaging of Human Dermal Fibroblasts and Mouse Skin via Promoting Nrf2-Regulated Antioxidant Defense. Front. Pharmacol. 2021, 12, 649820. [Google Scholar] [CrossRef]
- Galano, A.; Mazzone, G.; Alvarez-Diduk, R.; Marino, T.; Alvarez-Idaboy, J.R.; Russo, N. Food Antioxidants: Chemical Insights at the Molecular Level. Annu. Rev. Food Sci. Technol. 2016, 7, 335–352. [Google Scholar] [CrossRef]
- Demirci-Çekiç, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of Oxidative Stress and Antioxidant Defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, E.; Bishayee, A.; Fimognari, C. On a Beam of Light: Photoprotective Activities of the Marine Carotenoids Astaxanthin and Fucoxanthin in Suppression of Inflammation and Cancer. Mar. Drugs 2020, 18, 544. [Google Scholar] [CrossRef]
- Morales-García, B.C.; Pérez-González, A.; Galano, A. Spirochromene and Spiroindene Compounds as Antioxidants. J. Mol. Struct. 2024, 1318, 139355. [Google Scholar] [CrossRef]
- Polefka, T.G.; Meyer, T.A.; Agin, P.P.; Bianchini, R.J. Cutaneous Oxidative Stress. J. Cosmet. Dermatol. 2012, 11, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Romanhole, R.C.; Ataide, J.A.; Moriel, P.; Mazzola, P.G. Update on Ultraviolet A and B Radiation Generated by the Sun and Artificial Lamps and Their Effects on Skin. Int. J. Cosmet. Sci. 2015, 37, 366–370. [Google Scholar] [CrossRef]
- Marionnet, C.; Pierrard, C.; Golebiewski, C.; Bernerd, F. Diversity of Biological Effects Induced by Longwave UVA Rays (UVA1) in Reconstructed Skin. PLoS ONE 2014, 9, e105263. [Google Scholar] [CrossRef]
- Cadet, J.; Wagner, J.R. DNA Base Damage by Reactive Oxygen Species, Oxidizing Agents, and UV Radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Miyata, Y.; Okada, K.; Fujimoto, A.; Hata, K.-I.; Kagami, H.; Tomita, Y.; Ueda, M. The Effect of the Long-Term Cultivation on Telomere Length and Morphology of Cultured Epidermis. J. Dermatol. Sci. 2004, 34, 221–230. [Google Scholar] [CrossRef]
- Yadav, D.K.; Kumar, S.; Choi, E.-H.; Chaudhary, S.; Kim, M.-H. Molecular Dynamic Simulations of Oxidized Skin Lipid Bilayer and Permeability of Reactive Oxygen Species. Sci. Rep. 2019, 9, 4496. [Google Scholar] [CrossRef]
- Ling, H.; Lou, Y. Total Flavones from Elsholtzia Blanda Reduce Infarct Size during Acute Myocardial Ischemia by Inhibiting Myocardial Apoptosis in Rats. J. Ethnopharmacol. 2005, 101, 169–175. [Google Scholar] [CrossRef]
- Andrés Juan, C.; Pérez de Lastra, J.M.; Plou Gasca, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.A.; Vasilyeva, A.D.; Yurina, L.V.; Bychkova, A.V. Oxidation of Proteins: Is It a Programmed Process? Free Radic. Res. 2018, 52, 14–38. [Google Scholar] [CrossRef] [PubMed]
- Grune, T. Redox Regulation in Aging: Role of Protein Aggregates. Free Radic. Biol. Med. 2018, 124, 564. [Google Scholar] [CrossRef]
- Höhn, A.; König, J.; Grune, T. Protein Oxidation in Aging and the Removal of Oxidized Proteins. J. Proteom. 2013, 92, 132–159. [Google Scholar] [CrossRef]
- Jang, H.-Y.; Kim, G.-B.; Kim, J.-M.; Kang, S.Y.; Youn, H.-J.; Park, J.; Ro, S.Y.; Chung, E.-Y.; Park, K.-H.; Kim, J.-S. Fisetin Inhibits UVA-Induced Expression of MMP-1 and MMP-3 through the NOX/ROS/MAPK Pathway in Human Dermal Fibroblasts and Human Epidermal Keratinocytes. Int. J. Mol. Sci. 2023, 24, 17358. [Google Scholar] [CrossRef]
- Kciuk, M.; Gielecińska, A.; Budzinska, A.; Mojzych, M.; Kontek, R. Metastasis and MAPK Pathways. Int. J. Mol. Sci. 2022, 23, 3847. [Google Scholar] [CrossRef]
- Zhang, J.; Bowden, G.T. Activation of P38 MAP Kinase and JNK Pathways by UVA Irradiation. Photochem. Photobiol. Sci. 2012, 11, 54–61. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Y.; Wang, C. P38 MAPK in Regulating Cellular Responses to Ultraviolet Radiation. J. Biomed. Sci. 2007, 14, 303–312. [Google Scholar] [CrossRef]
- Lasa, M.; Mahtani, K.R.; Finch, A.; Brewer, G.; Saklatvala, J.; Clark, A.R. Regulation of Cyclooxygenase 2 mRNA Stability by the Mitogen-Activated Protein Kinase P38 Signaling Cascade. Mol. Cell Biol. 2000, 20, 4265–4274. [Google Scholar] [CrossRef]
- Tanveer, M.A.; Rashid, H.; Tasduq, S.A. Molecular Basis of Skin Photoaging and Therapeutic Interventions by Plant-Derived Natural Product Ingredients: A Comprehensive Review. Heliyon 2023, 9, e13580. [Google Scholar] [CrossRef]
- Yoshioka, H.; Hino, Y.; Iwata, K.; Ogawa, T.; Yoshioka, M.; Ishihama, N.; Adachi, H. Dynamics of Plant Immune MAPK Activity and ROS Signaling in Response to Invaders. Physiol. Mol. Plant Pathol. 2023, 125, 102000. [Google Scholar] [CrossRef]
- Zhang, D.D.; Lo, S.-C.; Cross, J.V.; Templeton, D.J.; Hannink, M. Keap1 Is a Redox-Regulated Substrate Adaptor Protein for a Cul3-Dependent Ubiquitin Ligase Complex. Mol. Cell Biol. 2004, 24, 10941–10953. [Google Scholar] [CrossRef]
- Wang, T.; Jian, Z.; Baskys, A.; Yang, J.; Li, J.; Guo, H.; Hei, Y.; Xian, P.; He, Z.; Li, Z.; et al. MSC-Derived Exosomes Protect against Oxidative Stress-Induced Skin Injury via Adaptive Regulation of the NRF2 Defense System. Biomaterials 2020, 257, 120264. [Google Scholar] [CrossRef]
- Wu, K.C.; Cui, J.Y.; Klaassen, C.D. Beneficial Role of Nrf2 in Regulating NADPH Generation and Consumption. Toxicol. Sci. 2011, 123, 590–600. [Google Scholar] [CrossRef]
- Tian, F.F.; Zhang, F.F.; Lai, X.D.; Wang, L.J.; Yang, L.; Wang, X.; Singh, G.; Zhong, J.L. Nrf2-Mediated Protection against UVA Radiation in Human Skin Keratinocytes. Biosci. Trends 2011, 5, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; McMahon, M.; Chowdhry, S.; Dinkova-Kostova, A.T. Cancer Chemoprevention Mechanisms Mediated through the Keap1-Nrf2 Pathway. Antioxid. Redox Signal 2010, 13, 1713–1748. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Suzuki, T.; Hiramoto, K.; Asami, S.; Naganuma, E.; Suda, H.; Iso, T.; Yamamoto, H.; Morita, M.; Baird, L.; et al. Characterizations of Three Major Cysteine Sensors of Keap1 in Stress Response. Mol. Cell Biol. 2016, 36, 271–284. [Google Scholar] [CrossRef]
- Hu, N.; An, R.; Yu, K.; Chang, Y.; Gao, W. PF4 Induces Inflammatory Response through NF-kB Signal Pathway in Rats with Intracerebral Haemorrhage. Folia Neuropathol. 2023, 61, 379–386. [Google Scholar] [CrossRef]
- Barber, K.; Mendonca, P.; Evans, J.A.; Soliman, K.F.A. Antioxidant and Anti-Inflammatory Mechanisms of Cardamonin through Nrf2 Activation and NF-kB Suppression in LPS-Activated BV-2 Microglial Cells. Int. J. Mol. Sci. 2023, 24, 10872. [Google Scholar] [CrossRef]
- Pahl, H.L. Activators and Target Genes of Rel/NF-kappaB Transcription Factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef]
- García-García, V.A.; Alameda, J.P.; Page, A.; Casanova, M.L. Role of NF-κB in Ageing and Age-Related Diseases: Lessons from Genetically Modified Mouse Models. Cells 2021, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Wen, X.; Hao, D.; Zhang, N.; He, G.; Jiang, X. NF-κB Signaling in Skin Aging. Mech. Ageing Dev. 2019, 184, 111160. [Google Scholar] [CrossRef]
- Youssef, N.S.; Elzaitony, A.S.; Baky, N.A.A. Diacerein Attenuate LPS-Induced Acute Lung Injury via Inhibiting ER Stress and Apoptosis: Impact on the Crosstalk between SphK1/S1P, TLR4/NFκB/STAT3, and NLRP3/IL-1β Signaling Pathways. Life Sci. 2022, 308, 120915. [Google Scholar] [CrossRef]
- Gong, W.; Li, J.; Chen, W.; Feng, F.; Deng, Y. Resveratrol Inhibits Lipopolysaccharide-Induced Extracellular Matrix Accumulation and Inflammation in Rat Glomerular Mesangial Cells by SphK1/S1P2/NF-κB Pathway. Diabetes Metab. Syndr. Obes. 2020, 13, 4495–4505. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.A.A.; Grougnet, R.; Nicolau, E.; Picot, L.; de Oliveira Junior, R.G. Carotenoids from Marine Microalgae as Antimelanoma Agents. Mar. Drugs 2022, 20, 618. [Google Scholar] [CrossRef]
- Aslanbay Guler, B.; Saglam-Metiner, P.; Deniz, I.; Demirel, Z.; Yesil-Celiktas, O.; Imamoglu, E. Aligned with Sustainable Development Goals: Microwave Extraction of Astaxanthin from Wet Algae and Selective Cytotoxic Effect of the Extract on Lung Cancer Cells. Prep. Biochem. Biotechnol. 2023, 53, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Matsumoto, T.; Takuwa, M.; Saeed Ebrahim Shaiku Ali, M.; Hirabashi, T.; Kondo, H.; Fujino, H. Protective Effects of Astaxanthin Supplementation against Ultraviolet-Induced Photoaging in Hairless Mice. Biomedicines 2020, 8, 18. [Google Scholar] [CrossRef]
- Komatsu, T.; Sasaki, S.; Manabe, Y.; Hirata, T.; Sugawara, T. Preventive Effect of Dietary Astaxanthin on UVA-Induced Skin Photoaging in Hairless Mice. PLoS ONE 2017, 12, e0171178. [Google Scholar] [CrossRef]
- Arslansoy, N.; Fidan, O.; Arslansoy, N.; Fidan, O. Carotenoids and Their Antioxidant Power. In The Power of Antioxidants-Unleashing Nature’s Defense Against Oxidative Stress; IntechOpen: London, UK, 2024; ISBN 978-0-85466-205-0. [Google Scholar]
- Zeb, A.; Hussain, A. Chemo-Metric Analysis of Carotenoids, Chlorophylls, and Antioxidant Activity of Trifolium Hybridum. Heliyon 2020, 6, e03195. [Google Scholar] [CrossRef]
- Chu, X.; Liu, J.; Gu, W.; Tian, L.; Tang, S.; Zhang, Z.; Jiang, L.; Xu, X. Study of the Properties of Carotenoids and Key Carotenoid Biosynthesis Genes from Deinococcus Xibeiensis R13. Biotechnol. Appl. Biochem. 2022, 69, 1459–1473. [Google Scholar] [CrossRef]
- Sahli, K.; Gomri, M.A.; Esclapez, J.; Gómez-Villegas, P.; Bonete, M.-J.; León, R.; Kharroub, K. Characterization and Biological Activities of Carotenoids Produced by Three Haloarchaeal Strains Isolated from Algerian Salt Lakes. Arch. Microbiol. 2021, 204, 6. [Google Scholar] [CrossRef] [PubMed]
- Stajčić, S.; Ćetković, G.; Čanadanović-Brunet, J.; Djilas, S.; Mandić, A.; Četojević-Simin, D. Tomato Waste: Carotenoids Content, Antioxidant and Cell Growth Activities. Food Chem. 2015, 172, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Shimidzu, N.; Goto, M.; Miki, W. Carotenoids as Singlet Oxygen Quenchers in Marine Organisms. Fish. Sci. 1996, 62, 134–137. [Google Scholar] [CrossRef]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological Properties of Carotenoids Extracted from Halobacterium Halobium Isolated from a Tunisian Solar Saltern. BMC Complement. Altern. Med. 2013, 13, 255. [Google Scholar] [CrossRef]
- Hou, J.; Cui, H.-L. In Vitro Antioxidant, Antihemolytic, and Anticancer Activity of the Carotenoids from Halophilic Archaea. Curr. Microbiol. 2018, 75, 266–271. [Google Scholar] [CrossRef]
- Hegazy, G.E.; Abu-Serie, M.M.; Abo-Elela, G.M.; Ghozlan, H.; Sabry, S.A.; Soliman, N.A.; Abdel-Fattah, Y.R. In Vitro Dual (Anticancer and Antiviral) Activity of the Carotenoids Produced by Haloalkaliphilic Archaeon Natrialba Sp. M6. Sci. Rep. 2020, 10, 5986. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, S.; Zargar, M.; Zolfaghari, M.R.; Amoozegar, M.A. Carotenoid Pigment of Halophilic Archaeon Haloarcula Sp. A15 Induces Apoptosis of Breast Cancer Cells. Cell Biochem. Funct. 2023, 41, 344–354. [Google Scholar] [CrossRef]
- Gómez-Villegas, P.; Vigara, J.; Vila, M.; Varela, J.; Barreira, L.; Léon, R. Antioxidant, Antimicrobial, and Bioactive Potential of Two New Haloarchaeal Strains Isolated from Odiel Salterns (Southwest Spain). Biology 2020, 9, 298. [Google Scholar] [CrossRef]
- Sheokand, P.; Tiwari, S.K. Characterization of Carotenoids Extracted from Haloferax Larsenii NCIM 5678 Isolated from Pachpadra Salt Lake, Rajasthan. Extremophiles 2024, 28, 33. [Google Scholar] [CrossRef]
- Fariq, A.; Yasmin, A.; Jamil, M. Production, Characterization and Antimicrobial Activities of Bio-Pigments by Aquisalibacillus Elongatus MB592, Salinicoccus Sesuvii MB597, and Halomonas Aquamarina MB598 Isolated from Khewra Salt Range, Pakistan. Extremophiles 2019, 23, 435–449. [Google Scholar] [CrossRef]
- Shahmohammadi, H.R.; Asgarani, E.; Terato, H.; Saito, T.; Ohyama, Y.; Gekko, K.; Yamamoto, O.; Ide, H. Protective Roles of Bacterioruberin and Intracellular KCl in the Resistance of Halobacterium Salinarium against DNA-Damaging Agents. J. Radiat. Res. 1998, 39, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Kurniawan, R.; Taslim, N.A.; Aman, A.M.; Syauki, A.Y.; Bukhari, A.; Bahar, B.; Mayulu, N.; Putra, D.E.; Syahputra, R.A.; Kim, B.; et al. Pharmacoinformatics and ex vivo Studies of Carotenoids from Green Algae Caulerpa Racemosa as Functional Biomolecules to Modulate Type-2 Diabetes Markers. S. Afr. J. Bot. 2025, 178, 348–359. [Google Scholar] [CrossRef]
- Mao, M.; Lei, H.; Liu, Q.; He, R.; Zuo, Z.; Zhang, N.; Zhou, C. Lycopene Inhibits Neointimal Hyperplasia through Regulating Lipid Metabolism and Suppressing Oxidative Stress. Mol. Med. Rep. 2014, 10, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Gao, R.; Song, X.; Li, X.; Zhu, J. Cardio-Protective and Anti-Atherosclerosis Effect of Crocetin on Vitamin D3 and HFD-Induced Atherosclerosis in Rats. J. Oleo Sci. 2021, 70, 1447–1459. [Google Scholar] [CrossRef]
- Liu, S.; Mohri, S.; Tsukamoto, M.; Yanai, Y.; Manabe, Y.; Sugawara, T. Preventive Effects of Dietary Fucoxanthin on Ultraviolet A Induced Photoaging in Hairless Mice. J. Sci. Food Agric. 2025, 105, 453–464. [Google Scholar] [CrossRef]
- Shanaida, M.; Mykhailenko, O.; Lysiuk, R.; Hudz, N.; Balwierz, R.; Shulhai, A.; Shapovalova, N.; Shanaida, V.; Bjørklund, G. Carotenoids for Antiaging: Nutraceutical, Pharmaceutical, and Cosmeceutical Applications. Pharmaceuticals 2025, 18, 403. [Google Scholar] [CrossRef]
- Hata, T.R.; Scholz, T.A.; Ermakov, I.V.; McClane, R.W.; Khachik, F.; Gellermann, W.; Pershing, L.K. Non-Invasive Raman Spectroscopic Detection of Carotenoids in Human Skin. J. Investig. Dermatol. 2000, 115, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J. An Overview of Carotenoids, Apocarotenoids, and Vitamin A in Agro-Food, Nutrition, Health, and Disease. Mol. Nutr. Food Res. 2019, 63, e1801045. [Google Scholar] [CrossRef]
- Novikov, V.S.; Kuzmin, V.V.; Darvin, M.E.; Lademann, J.; Sagitova, E.A.; Prokhorov, K.A.; Ustynyuk, L.Y.; Nikolaeva, G.Y. Relations between the Raman Spectra and Molecular Structure of Selected Carotenoids: DFT Study of α-Carotene, β-Carotene, γ-Carotene and Lycopene. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2022, 270, 120755. [Google Scholar] [CrossRef]
- Lin, H.-W.; Chang, T.-J.; Yang, D.-J.; Chen, Y.-C.; Wang, M.; Chang, Y.-Y. Regulation of Virus-Induced Inflammatory Response by β-Carotene in RAW264.7 Cells. Food Chem. 2012, 134, 2169–2175. [Google Scholar] [CrossRef]
- Dembińska-Kieć, A.; Malczewska-Malec, M.; Polus, A.; Kieć-Wilk, B.; Grzybowska, J.; Stachura, J.; Dyduch, G.; Pryjma, J.; Skrzeczyńska, J.; Langman, T.; et al. β-Carotene and Angiogenesis. Pure Appl. Chem. 2006, 78, 1519–1537. [Google Scholar] [CrossRef]
- Białek-Bylka, G.E.; Jazurek, B.; Dedic, R.; Hala, J.; Skrzypczak, A. Unique Spectroscopic Properties of Synthetic 15-Cis Beta-Carotene, an Important Compound in Photosynthesis, and a Medicine for Photoprotective Function. Cell Mol. Biol. Lett. 2003, 8, 689–697. [Google Scholar]
- Cui, B.; Liu, S.; Wang, Q.; Lin, X. Effect of β-Carotene on Immunity Function and Tumour Growth in Hepatocellular Carcinoma Rats. Molecules 2012, 17, 8595–8603. [Google Scholar] [CrossRef]
- Cho, S.; Lee, D.H.; Won, C.-H.; Kim, S.M.; Lee, S.; Lee, M.-J.; Chung, J.H. Differential Effects of Low-Dose and High-Dose Beta-Carotene Supplementation on the Signs of Photoaging and Type I Procollagen Gene Expression in Human Skin in Vivo. Dermatology 2010, 221, 160–171. [Google Scholar] [CrossRef]
- Lee, J.; Jiang, S.; Levine, N.; Watson, R.R. Carotenoid Supplementation Reduces Erythema in Human Skin after Simulated Solar Radiation Exposure. Proc. Soc. Exp. Biol. Med. 2000, 223, 170–174. [Google Scholar] [CrossRef]
- Minami, Y.; Kawabata, K.; Kubo, Y.; Arase, S.; Hirasaka, K.; Nikawa, T.; Bando, N.; Kawai, Y.; Terao, J. Peroxidized Cholesterol-Induced Matrix Metalloproteinase-9 Activation and Its Suppression by Dietary Beta-Carotene in Photoaging of Hairless Mouse Skin. J. Nutr. Biochem. 2009, 20, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Schagen, S.K.; Zampeli, V.A.; Makrantonaki, E.; Zouboulis, C.C. Discovering the Link between Nutrition and Skin Aging. Dermato-Endocrinology 2012, 4, 298–307. [Google Scholar] [CrossRef]
- White, A.L.; Jahnke, L.S. Contrasting Effects of UV-A and UV-B on Photosynthesis and Photoprotection of Beta-Carotene in Two Dunaliella spp. Plant Cell Physiol. 2002, 43, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Balić, A.; Mokos, M. Do We Utilize Our Knowledge of the Skin Protective Effects of Carotenoids Enough? Antioxidants 2019, 8, 259. [Google Scholar] [CrossRef]
- Camera, E.; Mastrofrancesco, A.; Fabbri, C.; Daubrawa, F.; Picardo, M.; Sies, H.; Stahl, W. Astaxanthin, Canthaxanthin and Beta-Carotene Differently Affect UVA-Induced Oxidative Damage and Expression of Oxidative Stress-Responsive Enzymes. Exp. Dermatol. 2009, 18, 222–231. [Google Scholar] [CrossRef]
- Wertz, K.; Seifert, N.; Hunziker, P.B.; Riss, G.; Wyss, A.; Hunziker, W.; Goralczyk, R. β-Carotene Interference with UVA-Induced Gene Expression by Multiple Pathways. Pure Appl. Chem. 2006, 78, 1539–1550. [Google Scholar] [CrossRef]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids Activate the Antioxidant Response Element Transcription System. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar] [CrossRef]
- Xu, G.; Ma, T.; Zhou, C.; Zhao, F.; Peng, K.; Li, B. β-Carotene Attenuates Apoptosis and Autophagy via PI3K/AKT/mTOR Signaling Pathway in Necrotizing Enterocolitis Model Cells IEC-6. Evidence-Based Complement. Altern. Med. 2022, 2022, 2502263. [Google Scholar] [CrossRef]
- Wu, T.; Xie, Y.; Wu, Z.; Li, Y.; Jiang, M.; Yu, H.; Li, X.; Wang, J.; Zhou, E.; Yang, Z. β-Carotene Protects Mice against Lipopolysaccharide and D-Galactosamine Induced Acute Liver Injury via Regulation of NF-κB, MAPK, and Nrf2 Signaling. J. Oleo Sci. 2023, 72, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.V.; Xu, W.; Li, Y.; Qin, J.; Zhou, T.; Li, D.; Xu, Y.; Cheng, X.; Xiong, Y.; Chen, Z. Anti-Aging Effect of β-Carotene through Regulating the KAT7-P15 Signaling Axis, Inflammation and Oxidative Stress Process. Cell Mol. Biol. Lett. 2022, 27, 86. [Google Scholar] [CrossRef]
- Wertz, K.; Hunziker, P.B.; Seifert, N.; Riss, G.; Neeb, M.; Steiner, G.; Hunziker, W.; Goralczyk, R. Beta-Carotene Interferes with Ultraviolet Light A-Induced Gene Expression by Multiple Pathways. J. Investig. Dermatol. 2005, 124, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Wertz, K.; Seifert, N.; Hunziker, P.B.; Riss, G.; Wyss, A.; Lankin, C.; Goralczyk, R. β-Carotene Inhibits UVA-Induced Matrix Metalloprotease 1 and 10 Expression in Keratinocytes by a Singlet Oxygen-Dependent Mechanism. Free Radic. Biol. Med. 2004, 37, 654–670. [Google Scholar] [CrossRef]
- Bayerl, C. Beta-Carotene in Dermatology: Does It Help? Acta Dermatoven APA 2008, 17, 161. [Google Scholar]
- Grabowska, M.; Wawrzyniak, D.; Rolle, K.; Chomczyński, P.; Oziewicz, S.; Jurga, S.; Barciszewski, J. Let Food Be Your Medicine: Nutraceutical Properties of Lycopene. Food Funct. 2019, 10, 3090–3102. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and Human Health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, J.; Yang, Q.; Yang, J. Metabolic Engineering Escherichia Coli for the Production of Lycopene. Molecules 2020, 25, 3136. [Google Scholar] [CrossRef]
- Arathi, B.P.; Sowmya, P.R.-R.; Vijay, K.; Baskaran, V.; Lakshminarayana, R. Metabolomics of Carotenoids: The Challenges and Prospects—A Review. Trends Food Sci. Technol. 2015, 45, 105–117. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Lee, J.-H.; Kim, N.-H.; Yeom, S.-J.; Kim, S.-W.; Oh, D.-K. Increase of Lycopene Production by Supplementing Auxiliary Carbon Sources in Metabolically Engineered Escherichia Coli. Appl. Microbiol. Biotechnol. 2011, 90, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.K.; Jeong, S.-W.; Yang, J.E.; Choi, Y.J. High-Yield Production of Lycopene from Corn Steep Liquor and Glycerol Using the Metabolically Engineered Deinococcus Radiodurans R1 Strain. J. Agric. Food Chem. 2020, 68, 5147–5153. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the Most Efficient Biological Carotenoid Singlet Oxygen Quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Huang, C.-C.; Lin, W.-T.; Hsieh, C.-C.; Huang, S.-Y.; Lin, S.-J.; Yang, S.-C. Lycopene Supplementation Attenuated Xanthine Oxidase and Myeloperoxidase Activities in Skeletal Muscle Tissues of Rats after Exhaustive Exercise. Br. J. Nutr. 2005, 94, 595–601. [Google Scholar] [CrossRef]
- Li, J.; Zeng, X.; Yang, X.; Ding, H. Lycopene Ameliorates Skin Aging by Regulating the Insulin Resistance Pathway and Activating SIRT1. Food Funct. 2022, 13, 11307–11320. [Google Scholar] [CrossRef]
- Almeer, R.; Alyami, N.M. Effect of Lycopene on TiO2 Nanoforms Induced Oxidative Stress and Neuroinflammation in SH-SY5Y Cells: An in Vitro Study. Drug Chem. Toxicol. 2025, 48, 51–61. [Google Scholar] [CrossRef]
- Honda, M. Z-Isomers of Lycopene and β-Carotene Exhibit Greater Skin-Quality Improving Action than Their All-E-Isomers. Food Chem. 2023, 421, 135954. [Google Scholar] [CrossRef]
- Stahl, W.; Heinrich, U.; Aust, O.; Tronnier, H.; Sies, H. Lycopene-Rich Products and Dietary Photoprotection. Photochem. Photobiol. Sci. 2006, 5, 238–242. [Google Scholar] [CrossRef]
- Tarshish, E.; Hermoni, K.; Sharoni, Y.; Muizzuddin, N. Effect of Lumenato Oral Supplementation on Plasma Carotenoid Levels and Improvement of Visual and Experiential Skin Attributes. J. Cosmet. Dermatol. 2022, 21, 4042–4052. [Google Scholar] [CrossRef]
- Chernyshova, M.P.; Pristenskiy, D.V.; Lozbiakova, M.V.; Chalyk, N.E.; Bandaletova, T.Y.; Petyaev, I.M. Systemic and Skin-Targeting Beneficial Effects of Lycopene-Enriched Ice Cream: A Pilot Study. J. Dairy. Sci. 2019, 102, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.; Baig, M.M.F.A.; Akhtar, N.; Chen, Y.; Xie, B.; Li, B. Topical Lycopene Emulgel Significantly Improves Biophysical Parameters of Human Skin. Eur. J. Pharm. Biopharm. 2022, 180, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Darvin, M.; Patzelt, A.; Gehse, S.; Schanzer, S.; Benderoth, C.; Sterry, W.; Lademann, J. Cutaneous Concentration of Lycopene Correlates Significantly with the Roughness of the Skin. Eur. J. Pharm. Biopharm. 2008, 69, 943–947. [Google Scholar] [CrossRef]
- Rizwan, M.; Rodriguez-Blanco, I.; Harbottle, A.; Birch-Machin, M.A.; Watson, R.E.B.; Rhodes, L.E. Tomato Paste Rich in Lycopene Protects against Cutaneous Photodamage in Humans in Vivo: A Randomized Controlled Trial. Br. J. Dermatol. 2011, 164, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Offord, E.A.; Gautier, J.-C.; Avanti, O.; Scaletta, C.; Runge, F.; Krämer, K.; Applegate, L.A. Photoprotective Potential of Lycopene, β-Carotene, Vitamin E, Vitamin C and Carnosic Acid in UVA-Irradiated Human Skin Fibroblasts. Free Radic. Biol. Med. 2002, 32, 1293–1303. [Google Scholar] [CrossRef]
- Huang, C.; Wen, C.; Yang, M.; Gan, D.; Fan, C.; Li, A.; Li, Q.; Zhao, J.; Zhu, L.; Lu, D. Lycopene Protects against T-BHP-Induced Neuronal Oxidative Damage and Apoptosis via Activation of the PI3K/Akt Pathway. Mol. Biol. Rep. 2019, 46, 3387–3397. [Google Scholar] [CrossRef]
- Fang, Y.; Ou, S.; Wu, T.; Zhou, L.; Tang, H.; Jiang, M.; Xu, J.; Guo, K. Lycopene Alleviates Oxidative Stress via the PI3K/Akt/Nrf2pathway in a Cell Model of Alzheimer’s Disease. PeerJ 2020, 8, e9308. [Google Scholar] [CrossRef]
- Feng, D.; Ling, W.-H.; Duan, R.-D. Lycopene Suppresses LPS-Induced NO and IL-6 Production by Inhibiting the Activation of ERK, p38MAPK, and NF-κB in Macrophages. Inflamm. Res. 2010, 59, 115–121. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus Astaxanthin: Applications for Human Health and Nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential Health-Promoting Effects of Astaxanthin: A High-Value Carotenoid Mostly from Microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef]
- Mutale-Joan, C.; El Arroussi, H. Biotechnological Strategies Overcoming Limitations to H. Pluvialis-Derived Astaxanthin Production and Morocco’s Potential. Crit. Rev. Food Sci. Nutr. 2025, 65, 1404–1419. [Google Scholar] [CrossRef]
- Cunha, F.F.M.D.; Tonon, A.P.; Machado, F.; Travassos, L.R.; Grazzia, N.; Possatto, J.F.; Sant’ana, A.K.C.D.; Lopes, R.D.M.; Rodrigues, T.; Miguel, D.C.; et al. Astaxanthin Induces Autophagy and Apoptosis in Murine Melanoma B16F10-Nex2 Cells and Exhibits Antitumor Activity in Vivo. J. Chemother. 2024, 36, 222–237. [Google Scholar] [CrossRef]
- Shanmugapriya, K.; Kim, H.; Kang, H.W. In Vitro Antitumor Potential of Astaxanthin Nanoemulsion against Cancer Cells via Mitochondrial Mediated Apoptosis. Int. J. Pharm. 2019, 560, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Gowd, V.; Xiao, J.; Wang, M.; Chen, F.; Cheng, K.-W. Multi-Mechanistic Antidiabetic Potential of Astaxanthin: An Update on Preclinical and Clinical Evidence. Mol. Nutr. Food Res. 2021, 65, e2100252. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, Y.; Yoshida, H.; Kondo, K. Potential Anti-Atherosclerotic Properties of Astaxanthin. Mar. Drugs 2016, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Pourbagher-Shahri, A.M.; Samarghandian, S. Anti-Inflammatory Action of Astaxanthin and Its Use in the Treatment of Various Diseases. Biomed. Pharmacother. 2022, 145, 112179. [Google Scholar] [CrossRef]
- Ito, N.; Seki, S.; Ueda, F. The Protective Role of Astaxanthin for UV-Induced Skin Deterioration in Healthy People—A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2018, 10, 817. [Google Scholar] [CrossRef]
- Tominaga, K.; Hongo, N.; Karato, M.; Yamashita, E. Cosmetic Benefits of Astaxanthin on Humans Subjects. Acta Biochim. Pol. 2012, 59, 43–47. [Google Scholar] [CrossRef]
- Suganuma, K.; Nakajima, H.; Ohtsuki, M.; Imokawa, G. Astaxanthin Attenuates the UVA-Induced up-Regulation of Matrix-Metalloproteinase-1 and Skin Fibroblast Elastase in Human Dermal Fibroblasts. J. Dermatol. Sci. 2010, 58, 136–142. [Google Scholar] [CrossRef]
- Yoshihisa, Y.; Rehman, M.U.; Shimizu, T. Astaxanthin, a Xanthophyll Carotenoid, Inhibits Ultraviolet-Induced Apoptosis in Keratinocytes. Exp. Dermatol. 2014, 23, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.-T.; Yang, C.-M.; Yang, C.-H. Astaxanthin Protects Retinal Photoreceptor Cells against High Glucose-Induced Oxidative Stress by Induction of Antioxidant Enzymes via the PI3K/Akt/Nrf2 Pathway. Antioxidants 2020, 9, 729. [Google Scholar] [CrossRef] [PubMed]
- Hama, S.; Takahashi, K.; Inai, Y.; Shiota, K.; Sakamoto, R.; Yamada, A.; Tsuchiya, H.; Kanamura, K.; Yamashita, E.; Kogure, K. Protective Effects of Topical Application of a Poorly Soluble Antioxidant Astaxanthin Liposomal Formulation on Ultraviolet-Induced Skin Damage. J. Pharm. Sci. 2012, 101, 2909–2916. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Turner, S.D.; Brautigan, D.L. Xanthophylls Lutein and Zeaxanthin Modify Gene Expression and Induce Synthesis of Hyaluronan in Keratinocyte Model of Human Skin. Biochem. Biophys. Rep. 2015, 4, 52–58. [Google Scholar] [CrossRef]
- Murillo, A.G.; Hu, S.; Fernandez, M.L. Zeaxanthin: Metabolism, Properties, and Antioxidant Protection of Eyes, Heart, Liver, and Skin. Antioxidants 2019, 8, 390. [Google Scholar] [CrossRef]
- Santocono, M.; Zurria, M.; Berrettini, M.; Fedeli, D.; Falcioni, G. Influence of Astaxanthin, Zeaxanthin and Lutein on DNA Damage and Repair in UVA-Irradiated Cells. J. Photochem. Photobiol. B Biol. 2006, 85, 205–215. [Google Scholar] [CrossRef]
- Palombo, P.; Fabrizi, G.; Ruocco, V.; Ruocco, E.; Fluhr, J.; Roberts, R.; Morganti, P. Beneficial Long-Term Effects of Combined Oral/Topical Antioxidant Treatment with the Carotenoids Lutein and Zeaxanthin on Human Skin: A Double-Blind, Placebo-Controlled Study. Skin Pharmacol. Physiol. 2007, 20, 199–210. [Google Scholar] [CrossRef]
- González, S.; Astner, S.; An, W.; Pathak, M.A.; Goukassian, D. Dietary Lutein/Zeaxanthin Decreases Ultraviolet B-Induced Epidermal Hyperproliferation and Acute Inflammation in Hairless Mice. J. Investig. Dermatol. 2003, 121, 399–405. [Google Scholar] [CrossRef]
- Roberts, R.L.; Green, J.; Lewis, B. Lutein and Zeaxanthin in Eye and Skin Health. Clin. Dermatol. 2009, 27, 195–201. [Google Scholar] [CrossRef]
- Chintong, S.; Phatvej, W.; Rerk-Am, U.; Waiprib, Y.; Klaypradit, W. In Vitro Antioxidant, Antityrosinase, and Cytotoxic Activities of Astaxanthin from Shrimp Waste. Antioxidants 2019, 8, 128. [Google Scholar] [CrossRef]
- Abd El-Ghany, M.N.; Hamdi, S.A.; Elbaz, R.M.; Aloufi, A.S.; El Sayed, R.R.; Ghonaim, G.M.; Farahat, M.G. Development of a Microbial-Assisted Process for Enhanced Astaxanthin Recovery from Crab Exoskeleton Waste. Fermentation 2023, 9, 505. [Google Scholar] [CrossRef]
- Siziya, I.N.; Hwang, C.Y.; Seo, M.-J. Antioxidant Potential and Capacity of Microorganism-Sourced C30 Carotenoids—A Review. Antioxidants 2022, 11, 1963. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Seo, D.-H.; Park, Y.-S.; Cha, I.-T.; Seo, M.-J. Isolation of Lactobacillus Plantarum Subsp. Plantarum Producing C30 Carotenoid 4,4’-Diaponeurosporene and the Assessment of Its Antioxidant Activity. J. Microbiol. Biotechnol. 2019, 29, 1925–1930. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.Y.; Cho, E.-S.; Yoon, D.J.; Seo, M.-J. Probiotic and Antioxidant Properties of C30 Carotenoid-Producing Lactiplantibacillus Plantarum Isolated from Kimchi. Food Sci. Biotechnol. 2023, 32, 543–552. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, M.S.; Lee, B.Y.; Lee, P.C. Generation of Structurally Novel Short Carotenoids and Study of Their Biological Activity. Sci. Rep. 2016, 6, 21987. [Google Scholar] [CrossRef]
- Naziri, D.; Hamidi, M.; Hassanzadeh, S.; Tarhriz, V.; Zanjani, B.M.; Nazemyieh, H.; Hejazi, M.A.; Hejazi, M.S. Analysis of Carotenoid Production by Halorubrum Sp. TBZ126; an Extremely Halophilic Archeon from Urmia Lake. Adv. Pharm. Bull. 2014, 4, 61–67. [Google Scholar] [CrossRef]
- Alvares, J.J.; Furtado, I.J. Characterization of Multicomponent Antioxidants from Haloferax Alexandrinus GUSF-1 (KF796625). 3 Biotech 2021, 11, 58. [Google Scholar] [CrossRef]
- Squillaci, G.; Parrella, R.; Carbone, V.; Minasi, P.; La Cara, F.; Morana, A. Carotenoids from the Extreme Halophilic Archaeon Haloterrigena Turkmenica: Identification and Antioxidant Activity. Extremophiles 2017, 21, 933–945. [Google Scholar] [CrossRef]
- Moopantakath, J.; Imchen, M.; Anju, V.T.; Busi, S.; Dyavaiah, M.; Martínez-Espinosa, R.M.; Kumavath, R. Bioactive Molecules from Haloarchaea: Scope and Prospects for Industrial and Therapeutic Applications. Front. Microbiol. 2023, 14, 1113540. [Google Scholar] [CrossRef]
- Flegler, A.; Lipski, A. The C50 Carotenoid Bacterioruberin Regulates Membrane Fluidity in Pink-Pigmented Arthrobacter Species. Arch. Microbiol. 2021, 204, 70. [Google Scholar] [CrossRef]
- Hwang, C.Y.; Cho, E.-S.; Kim, S.; Kim, K.; Seo, M.-J. Optimization of Bacterioruberin Production from Halorubrum Ruber and Assessment of Its Antioxidant Potential. Microb. Cell Factories 2024, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Bouhamed, S.B.H.; Chaari, M.; Baati, H.; Zouari, S.; Ammar, E. Extreme Halophilic Archaea: Halobacterium Salinarum Carotenoids Characterization and Antioxidant Properties. Heliyon 2024, 10, e36832. [Google Scholar] [CrossRef]
- Flores, N.; Hoyos, S.; Venegas, M.; Galetović, A.; Zúñiga, L.M.; Fábrega, F.; Paredes, B.; Salazar-Ardiles, C.; Vilo, C.; Ascaso, C.; et al. Haloterrigena Sp. Strain SGH1, a Bacterioruberin-Rich, Perchlorate-Tolerant Halophilic Archaeon Isolated From Halite Microbial Communities, Atacama Desert, Chile. Front. Microbiol. 2020, 11, 324. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; Gómez-Villegas, P.; de Carvalho, C.C.C.R.; Vigara, J.; Motilva, V.; León, R.; Talero, E. Up-Regulation of the Nrf2/HO-1 Antioxidant Pathway in Macrophages by an Extract from a New Halophilic Archaea Isolated in Odiel Saltworks. Antioxidants 2023, 12, 1080. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Silva, T.D.C.D.; Vidal, E.E.; de Souza, R.D.F.R.; da Cunha Schmidt, K.; Mendes, P.V.D.; da Silva Andrade, R.F.; da Silva Oliveira, F.G.; de Lucena, B.T.L.; de Oliveira, M.B.M.; dos Santos Correia, M.T.; et al. Production of Carotenoid Sarcinaxanthin by Kocuria Palustris Isolated from Northeastern Brazil Caatinga Soil and Their Antioxidant and Photoprotective Activities. Electron. J. Biotechnol. 2021, 53, 44–53. [Google Scholar] [CrossRef]
- Wang, J.; Kong, W.; Liu, M.; Wang, Y.; Zheng, Y.; Zhou, Y. Association between Dietary Carotenoids Intake and Chronic Constipation in American Men and Women Adults: A Cross-Sectional Study. BMC Public Health 2023, 23, 1597. [Google Scholar] [CrossRef]
- Bates, C.A.; Vincent, M.J.; Buerger, A.N.; Santamaria, A.B.; Maier, A.; Jack, M. Investigating the Relationship between β-Carotene Intake from Diet and Supplements, Smoking, and Lung Cancer Risk. Food Chem. Toxicol. 2024, 194, 115104. [Google Scholar] [CrossRef]
- Edwards, J.A.; Bellion, P.; Beilstein, P.; Rümbeli, R.; Schierle, J. Review of Genotoxicity and Rat Carcinogenicity Investigations with Astaxanthin. Regul. Toxicol. Pharmacol. 2016, 75, 5–19. [Google Scholar] [CrossRef]
- Ramos-Souza, C.; De Rosso, V.V. Recent Approaches for the Bioaccessibility and Bioavailability of Carotenoids. In Carotenoids: Trends and Advances; Campos Chisté, R., Helena de Aguiar Andrade, E., Santana de Oliveira, M., Eds.; Springer Nature: Cham, Switzerland, 2024; pp. 153–178. ISBN 978-3-031-75322-0. [Google Scholar]
- Bouwstra, J.A.; Ponec, M. The Skin Barrier in Healthy and Diseased State. Biochim. Biophys. Acta (BBA)-Biomembr. 2006, 1758, 2080–2095. [Google Scholar] [CrossRef]
- Liu, W.-Y.; Hsieh, Y.-S.; Ko, H.-H.; Wu, Y.-T. Formulation Approaches to Crystalline Status Modification for Carotenoids: Impacts on Dissolution, Stability, Bioavailability, and Bioactivities. Pharmaceutics 2023, 15, 485. [Google Scholar] [CrossRef]
- de Freitas Santos, P.D.; Rubio, F.T.V.; da Silva, M.P.; Pinho, L.S.; Favaro-Trindade, C.S. Microencapsulation of Carotenoid-Rich Materials: A Review. Food Res. Int. 2021, 147, 110571. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Xue, J.; Lou, X.; Abbas, S.; Guan, Y.; Feng, B.; Zhang, X.; Xia, S. Liposomes as Delivery Systems for Carotenoids: Comparative Studies of Loading Ability, Storage Stability and in Vitro Release. Food Funct. 2014, 5, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Jalali-Jivan, M.; Rostamabadi, H.; Assadpour, E.; Tomas, M.; Capanoglu, E.; Alizadeh-Sani, M.; Kharazmi, M.S.; Jafari, S.M. Recent Progresses in the Delivery of β-Carotene: From Nano/Microencapsulation to Bioaccessibility. Adv. Colloid. Interface Sci. 2022, 307, 102750. [Google Scholar] [CrossRef]
- Luo, X.; Zhou, Y.; Bai, L.; Liu, F.; Deng, Y.; McClements, D.J. Fabrication of β-Carotene Nanoemulsion-Based Delivery Systems Using Dual-Channel Microfluidization: Physical and Chemical Stability. J. Colloid. Interface Sci. 2017, 490, 328–335. [Google Scholar] [CrossRef] [PubMed]
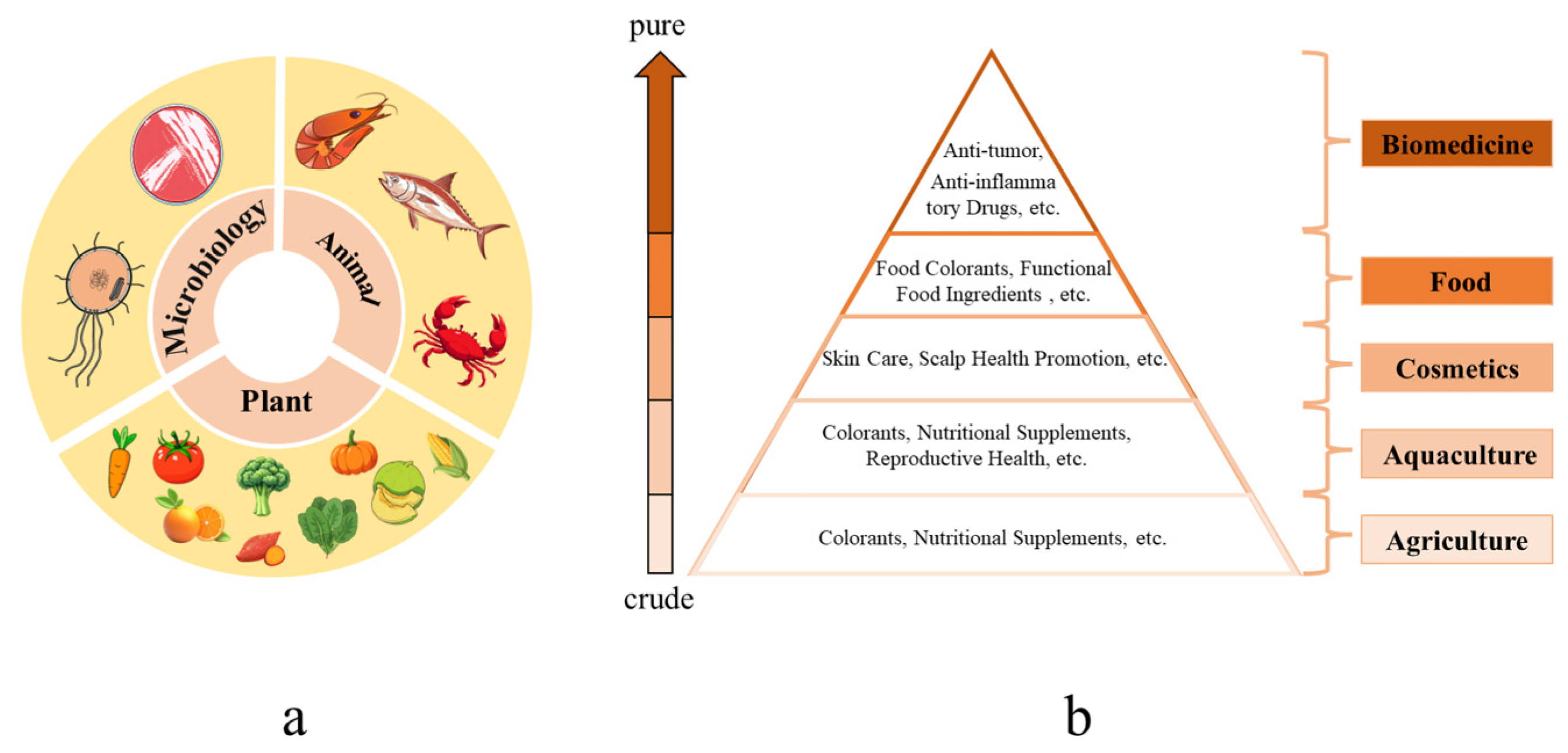


| Name | DPPH IC50 (μM) |
|---|---|
| 4,4′-Diapolycopene | 8.7 |
| 4,4′-Diaponeurosporene | 11.6 |
| 4,4′-Diaponeurosporen-4′-al | 10.2 |
| 4,4′-Diapolycopendial | 7.5 |
| 4,4′-Diaponeurosporen-4′-oic acid | 9.7 |
| 4,4′-Diapotorulene | 70.3 |
| 4,4′-Diapo-β-carotene | 77.8 |
| Name | Antioxidant Capacity | In Vitro Experiment | References |
|---|---|---|---|
| Bacterioruberin | ABTS (IC50 9.8 μg/mL), FRAP (IC50 2.1 μg/mL) | - | [172] |
| DPPH (IC50 86.67 μg/mL) | - | [173] | |
| - | Protection of cells against oxidizing DNA damaging agent | [92] | |
| - | Protection of erythrocytes from H2O2 | [86] | |
| ABTS (IC50 20.5 μg/mL), | Macrophages: reducing ROS levels and reducing levels of pro-inflammatory cytokines TNF-α and IL-6 | [175] | |
| Sarcinaxanthin | 1O2 | Displaying an SPF of 9.36 ± 0.52, exhibiting in vitro photoprotective activity | [176] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Li, C.; Su, W.; Sun, Z.; Gao, S.; Xie, W.; Zhang, B.; Sui, L. Carotenoids in Skin Photoaging: Unveiling Protective Effects, Molecular Insights, and Safety and Bioavailability Frontiers. Antioxidants 2025, 14, 577. https://doi.org/10.3390/antiox14050577
Ma Y, Li C, Su W, Sun Z, Gao S, Xie W, Zhang B, Sui L. Carotenoids in Skin Photoaging: Unveiling Protective Effects, Molecular Insights, and Safety and Bioavailability Frontiers. Antioxidants. 2025; 14(5):577. https://doi.org/10.3390/antiox14050577
Chicago/Turabian StyleMa, Yingchao, Chengxiang Li, Wanping Su, Zhongshi Sun, Shuo Gao, Wei Xie, Bo Zhang, and Liying Sui. 2025. "Carotenoids in Skin Photoaging: Unveiling Protective Effects, Molecular Insights, and Safety and Bioavailability Frontiers" Antioxidants 14, no. 5: 577. https://doi.org/10.3390/antiox14050577
APA StyleMa, Y., Li, C., Su, W., Sun, Z., Gao, S., Xie, W., Zhang, B., & Sui, L. (2025). Carotenoids in Skin Photoaging: Unveiling Protective Effects, Molecular Insights, and Safety and Bioavailability Frontiers. Antioxidants, 14(5), 577. https://doi.org/10.3390/antiox14050577