4. Discussion
In this study, incorporating GAA into the diet of the beef cattle was found to significantly boost the growth performance, a result that corroborates previous findings [
11,
17,
35]. Li et al. [
35] found that the ADG in Angus bulls significantly improved with increasing GAA supplementation of 0.3, 0.6, and 0.9 g/kg dry matter. Meanwhile, Yi et al. [
11] noted that the ADG in Angus steers improved with GAA supplementation at 0.8 g/kg and 1.6 g/kg, but they also observed that beyond a certain concentration, further increases in GAA did not lead to additional improvements in the ADG. The possibility arises that this phenomenon stems from elevated GAA supplementation triggering an increased methyl demand in the body, rendering the methyl yield from dietary breakdown inadequate to accommodate the augmented GAA provision. In this study, the IBW remained consistent across all treatment groups, while the GAM group exhibited the highest FBW, ADG, and FCE, surpassing the GAA group by 2.24%, 6.9%, and 10%, respectively. These results indicate that adding Met further improved the growth performance of the beef cattle. However, unlike the findings of Liu et al. [
17], GAA supplementation in this study did not lead to an increased feed intake. Furthermore, Li et al. [
36] reported that GAA addition significantly raised the DMI of Jinjiang bulls, which is also discordant with the findings in this study. This discrepancy might be attributed to variations in the breed and dietary composition between the studied beef cattle populations.
Rumen fermentation parameters reflect rumen health, and in this study, the rumen pH showed no significant variation between the groups, remaining stable at a healthy level. This suggests that neither the GAA nor Met negatively affected the ruminal health. Pertinent literature has revealed that GAA supplementation can lead to increased ruminal TVFA concentrations in beef cattle [
17,
35], a finding that aligns with the outcomes of our investigation. Since TVFAs originate from the degradation of nutrients by rumen microorganisms, elevated TVFAs may imply altered digestibility [
37]. Furthermore, the significantly elevated concentration of valeric acid in the rumen implies that the addition of GAA and Met potentially modulated the ruminal fermentation environment and influenced the breakdown of nutrients by rumen microbial populations. This observation was supported by the results of the apparent nutrient digestibility assessment. Our study revealed a significant increase in the NDF digestibility in the GAA and GAM groups relative to the CON group. Moreover, the DM and CP digestibilities in the GAM group exceeded those of the CON group, consistent with prior research [
11,
12,
17,
35]. The increase in nutrient digestibility also explains the significantly higher ADG in the absence of a difference in the DMI. This suggests that including GAA and Met in the diet could positively impact nutrient digestion in beef cattle.
The TP, including ALB and GLB, is an important indicator for detecting the synthetic function of the liver [
38]. ALB plays a vital role in preserving blood osmolarity and ensuring nutrient provision [
39], while GLB plays an immunological role in animals [
40]. In this study, GAA supplementation led to elevated serum concentrations of TP and GLB, with the ALB levels notably higher in the GAM group than in the CON group. These findings are consistent with those documented by Li et al. [
36]. In this study, TP, ALB, and GLB all experienced alterations within the standard concentration range, whereas ALT, AST, and ALP are pivotal markers for determining the normalcy or otherwise of liver function [
41]. The observation that these latter enzymes did not display significant disparities between the three treatment groups substantiates that the increases in the TP, ALB, and GLB were not a consequence of abnormal liver function. UREA is a product of protein metabolism [
42], which was increased in the serum of the GAM group. Collectively, these findings imply that the inclusion of GAA and Met may have moderately enhanced the liver’s protein-synthesizing function, thereby fostering improved protein metabolism within the organism. HSL, which is associated with lipolysis metabolism [
43], was significantly elevated in the sera of both the GAA and GAM groups. Similarly, FAS, which is linked to fatty acid synthesis [
44], also showed a significant increase in the serum of the GAM group. The possible reason for this phenomenon is the bidirectional regulation of energy metabolism. Specifically, the addition of GAA and Met may increase the body’s energy supply, which may stimulate adipose tissue to store fatty acids (FAS increase); in addition, GAA and Met may improve the body’s energy utilization, which increases HSL and promotes fat decomposition. Together, these observations suggest that GAA and Met supplementation may have an effect on the lipid metabolism in beef cattle. However, the serum lipid indices (TC, TG, HDL-C, and LDL-C) failed to reveal significant discrepancies between the three treatment groups, potentially due to the organism’s inherent regulatory mechanisms governing lipid metabolism [
45].
In this study, the serum GAA levels were significantly higher in the GAA group compared with the CON group, with a similar pattern observed in the GAM group. These results align with previous findings reported by Yi et al. [
11]. The results suggest that not all dietary GAA was degraded by rumen microorganisms; instead, a portion successfully entered the bloodstream, allowing it to fulfill its intended function. The lesser increase in the serum GAA concentration observed in the beef cattle from the GAM group may be attributed to the heightened utilization of GAA by the body resulting from Met supplementation. The concentrations of AGAT and GAMT, which are key rate-limiting enzymes involved in GAA synthesis and creatine metabolism [
46], were significantly higher in both the GAA and GAM groups compared with the control. This implies that the GAA supplementation stimulated creatine metabolism and augmented the demand for GAA within the animals’ bodies. In this study, no significant differences were observed in the serum creatine, creatine kinase, or ATP levels across the three groups. This could have been due to the elevated demand for creatine in the muscle tissues of the beef cattle, aligning with the higher ADG recorded. Another possible reason for this consistency is that the addition of GAA improves the body’s energy utilization, which leads to no significant increase in ATP in the blood. In addition, the animal’s own regulatory effect may also be the reason why there were no significant differences in the above three indicators [
47].
Creatine is known to possess the capacity to neutralize oxygen free radicals [
48]; hence, GAA supplementation may exert an indirect antioxidative effect. Previous research has demonstrated the beneficial effects of GAA on enhancing the antioxidant status in animals. For example, Li et al. [
12] observed that supplementing diets with GAA increased the antioxidant capacity in both the rumens and sera of lambs. Consistent with these findings, Yi et al. [
11] reported increased CAT concentrations and reduced MDA levels in the sera of Angus steers following GAA supplementation. These results align with the observations from the present research. T-AOC represents the overall levels of antioxidants, enzymes, and other components involved in oxidative defense within an animal’s body. It embodies the comprehensive antioxidant level derived from diverse antioxidant substances and antioxidant enzymes present in the animal system [
49]. Typically, T-AOC remains within a physiological range and can be influenced by diet, exercise, and metabolic conditions. In our study, despite the animals being healthy, the supplementation with GAA and Met significantly increased the T-AOC levels in both the serum and liver tissues. This suggests that GAA supplementation not only provides metabolic benefits but also enhances antioxidant defense mechanisms. One possible explanation for this significant increase in T-AOC is the upregulation of antioxidant enzyme activity, particularly SOD and CAT, which play distinct roles in mitigating oxidative stress. SOD converts superoxide anion radicals (O
2⁻) into hydrogen peroxide and oxygen, while CAT further decomposes hydrogen peroxide into water and oxygen [
50,
51]. The elevated activities of these enzymes in both serum and liver tissues indicate an enhanced endogenous antioxidant response. Notably, the increase in CAT activity suggests a potential metabolic adaptation to counteract oxidative stress associated with increased metabolic rates due to GAA supplementation. This supports the notion that while GAA promotes growth, it may also lead to increased metabolic activity, potentially inducing mild oxidative stress, which is then neutralized through an upregulated antioxidant defense system. Moreover, the reduction in MDA, a byproduct of lipid peroxidation [
52], further supports the hypothesis that GAA supplementation improves the body’s ability to manage oxidative stress. The observed increase in GSH levels in serum and liver tissues, without a corresponding significant increase in the GSH-Px activity, may be attributed to the increased consumption of GSH-Px in response to oxidative stress, resulting in a higher GSH turnover [
53]. This reflects a compensatory mechanism where the body maintains redox homeostasis through increased antioxidant production rather than solely relying on enzymatic activity. A substantial proportion of nutrients assimilated by the stomach and intestines are conveyed via the bloodstream to the liver, where they undergo metabolic transformations [
54] before being distributed to various tissues and organs throughout the body via the circulatory system. The consistency of the changes in the above antioxidant indicators in serum and liver tissues leads to the view that changes in liver metabolic function may affect the corresponding metabolic indicators in the blood [
55]. In addition, this study also observed significant changes in the levels of serum indicators related to protein metabolism and lipid metabolism to varying degrees. Hence, the metabolomic analysis of liver tissues in this study could facilitate the exploration of the effects of GAA supplementation on liver metabolic functions, while transcriptomic analysis could elucidate the precise mechanisms underlying metabolic function changes at the gene expression level.
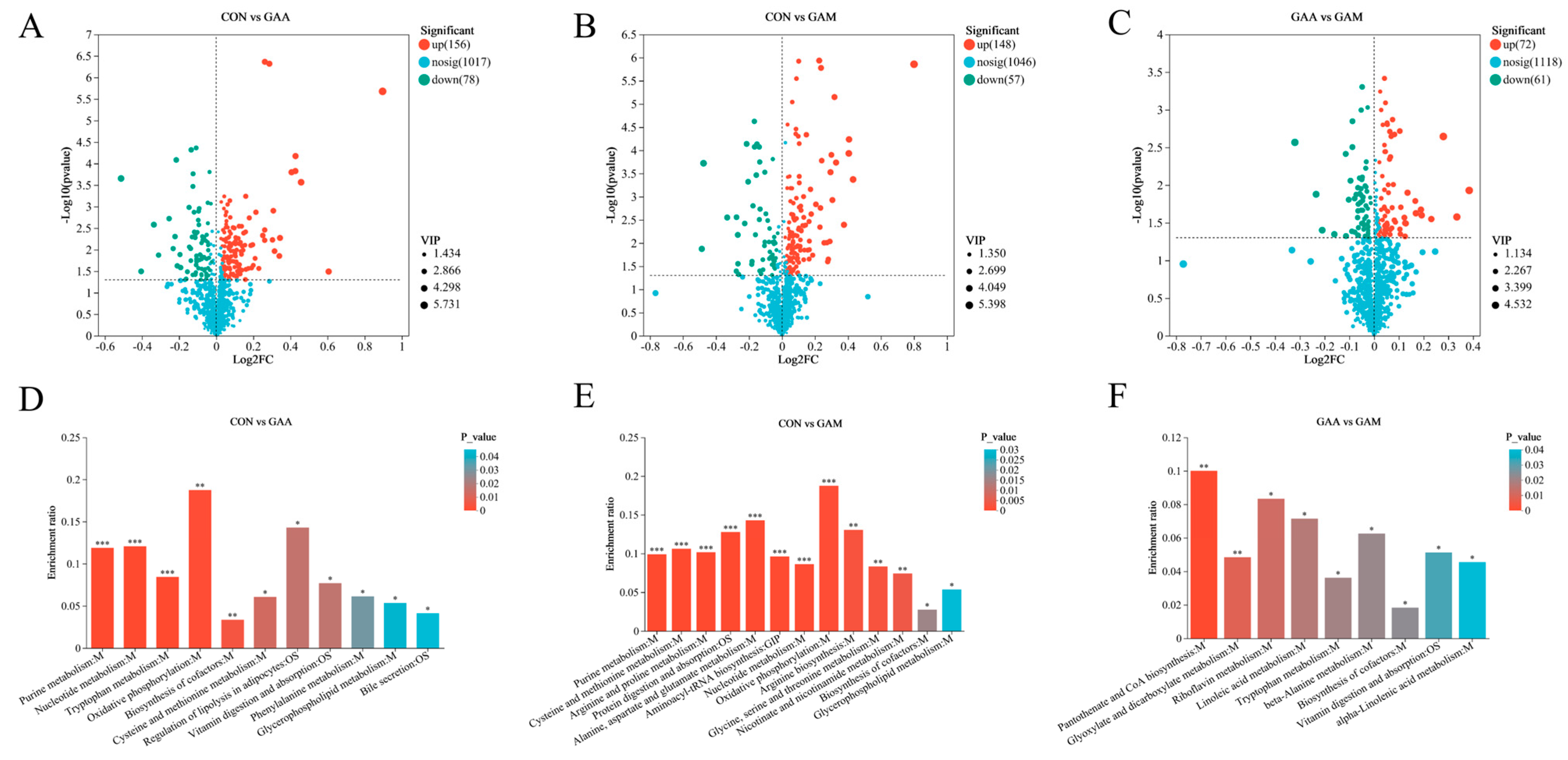
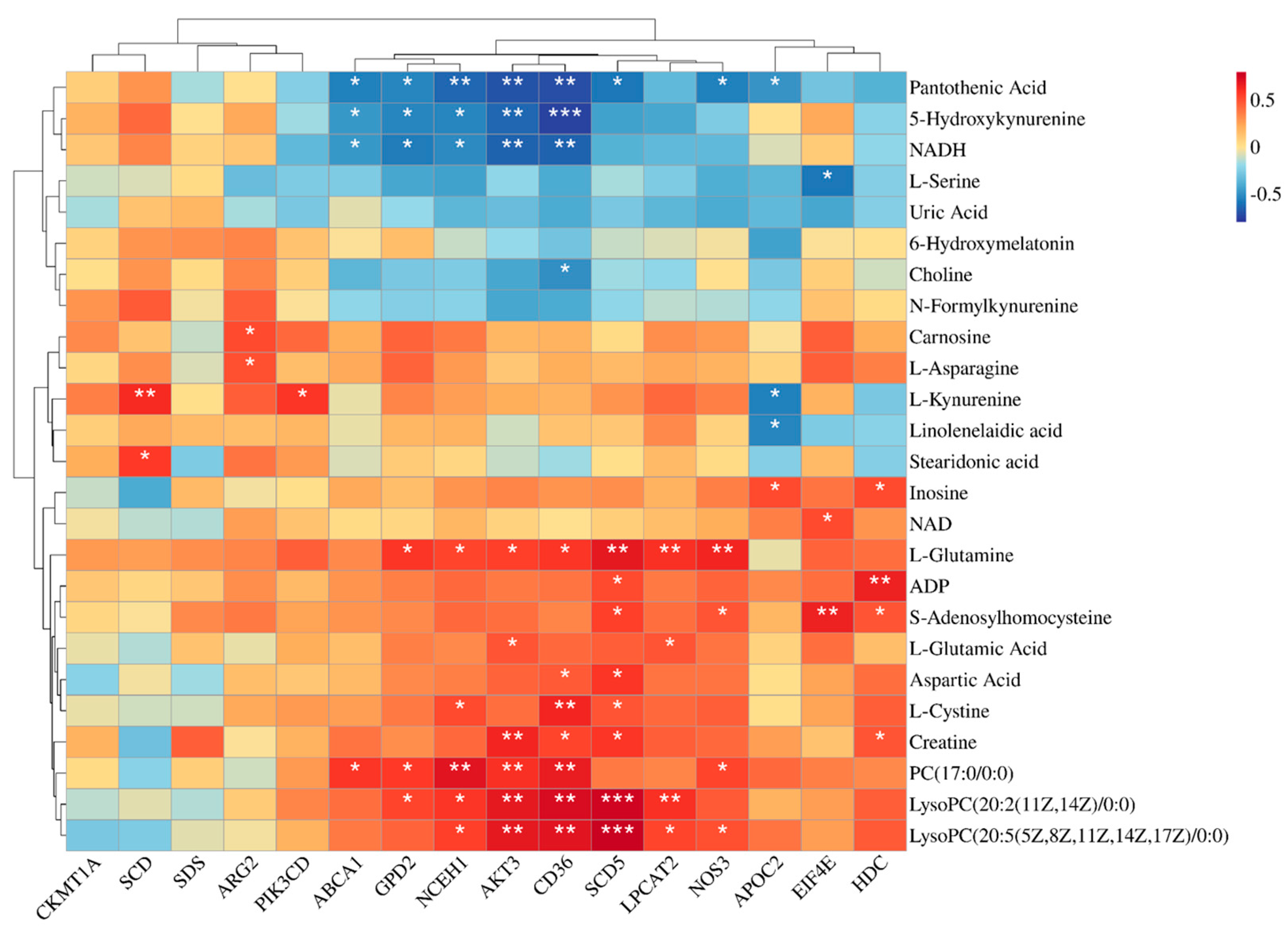
Relative to the CON group, the concentrations of Inosine, ADP, S-adenosylhomocysteine, L-cysteine, and PC (17:0/0:0) were elevated in both the GAA group and GAM group. Inosine is a key intermediate in purine metabolism and also participates in nucleotide metabolism and energy metabolism [
56]; ADP is also involved in the above metabolism and is also a marker of oxidative phosphorylation in the organism [
57]. The increased levels observed in both the GAA and GAM groups indicate that GAA supplementation enhances energy metabolism in beef cattle, providing abundant substrates for the synthesis of amino acids, sugars, nucleic acids, and fatty acids. S-adenosylhomocysteine (SAH), a crucial metabolite in creatine synthesis, is generated during the conversion of S-adenosylmethionine to creatine with GAA [
58]. Elevated SAH levels indicate increased creatine production in the hepatic tissues of both the GAA and GAM groups; in addition, SAH is metabolized to homocysteine, which reacts with betaine to be able to produce methionine, while choline is a precursor substance of betaine [
59]. The lower Choline levels observed in the GAA group, relative to the control, may indicate a higher requirement for betaine, indirectly suggesting that GAA supplementation enhanced the hepatic creatine metabolism. Furthermore, hepatic creatine levels in the GAM group were markedly elevated relative to the control, indicating that Met supplementation further enhanced the creatine synthesis in the liver. This suggests that the addition of Met further enhanced the creatine synthesis in the liver of the beef cattle. These changes corresponded to the changes in serum indicators of creatine metabolism. PC (17:0/0:0), LysoPC (20:2(11Z,14Z)/0:0), and LysoPC (20:5(5Z,8Z,11Z,14Z,17Z)/0:0) were all enriched in the glycerophospholipid metabolic pathway, which has the roles of constructing cell membranes, participating in lipid metabolism, and regulating the cholesterol metabolism in animals [
60]. The differing degrees of increase in the content of these three substances in the GAA group and GAM group indicate that the GAA supplementation stimulated the glycerophospholipid metabolic pathway, and the augmentation of this pathway suggests potential enhancement of lipid metabolism within the livers of the beef cattle. This finding also provides an explanation for the increased serum lipase content observed.
Previous research has demonstrated the impact of GAA supplementation on amino acid metabolism in animals [
10], which aligns with the results observed in the current investigation. In the hepatic tissues of the GAA group, the concentrations of 5-hydroxykynurenine and 6-hydroxymelatonin, which are both associated with the tryptophan metabolic pathway, were significantly reduced relative to the control. Tryptophan, an essential amino acid, is primarily used by the host for protein synthesis [
61]. The decreased levels of its related metabolites suggest that GAA supplementation suppresses tryptophan catabolism, thereby supporting improved protein synthesis. L-cystine, which is known for supporting hepatic detoxification [
62], significantly increased in both the GAA and GAM groups, suggesting that GAA supplementation boosts the liver detoxification capacity. Notably, Met supplementation significantly elevated the Aspartic acid, L-asparagine, L-glutamic acid, and L-glutamine levels while reducing L-serine in the liver of the GAM group cattle. These amino acids are integral to processes such as protein digestion and absorption. These alterations indicate increased activity in essential metabolic pathways related to protein synthesis, such as aminoacyl-tRNA biosynthesis; arginine and proline metabolisms; cysteine and methionine metabolisms; glycine, serine, and threonine metabolisms; and alanine, aspartate, and glutamate metabolisms. The rise in amino acid metabolites in the GAM group suggests that Met supplementation enhanced the hepatic amino acid metabolism. This reinforces the idea that beef cattle in the GAM group exhibited enhanced protein synthesis capacity, aligning with their greater daily weight gain.
NADH is known as the reduced coenzyme I and is involved in glycolysis and cellular respiration [
63], while NADPH is known as the reduced coenzyme II and plays energy supply, signal transduction, and antioxidant roles in cells [
64]. Although NADPH is widely considered to be the main reducing equivalent involved in the antioxidant defense system (especially through its role in the glutathione reductase and thioredoxin systems), recent studies have also emphasized that NADH has certain antioxidant properties [
65]. NADH can directly scavenge some reactive oxygen species (ROS) through its inherent reducing ability and can indirectly promote antioxidant regeneration, although to a lesser extent than NADPH; in addition, fluctuations in NADH/NAD⁺ levels can reflect the cellular redox status and metabolic activities, especially those within mitochondria [
63]. Therefore, the significant decrease in NADH levels observed in the GAA group and the significant increase in the level of its oxidized counterpart NAD+ in the GAM group in this study not only reflect changes in the energy metabolism but also partially contribute to redox regulation in liver tissue. Unlike NADH, NADPH is primarily produced through the pentose phosphate pathway (PPP), a key metabolic pathway that runs in parallel with glycolysis. The PPP plays a crucial role in cellular antioxidant defense by providing NADPH, which is essential for maintaining the reduced state of glutathione and supporting other antioxidant systems [
64]. Although NADPH was not specifically detected in our metabolomics analysis, its indirect role in redox homeostasis is well recognized. The enhanced antioxidant enzyme activities observed in the GAA and GAM groups may also reflect upstream activation of the metabolic pathways, including the PPP, which deserves further investigation in future studies. Additionally, hepatic concentrations of Pantothenic Acid and Carnosine, which are critical for Pantothenate and CoA biosynthesis and beta-Alanine metabolism, respectively, were markedly elevated in the GAM group relative to the GAA group. Pantothenic Acid is known to contribute to cellular energy metabolism, substance synthesis, and regulation [
66], whereas Carnosine is intimately tied to skeletal muscle energy metabolism and protein synthesis [
67]. Furthermore, L-Kynurenine and N′-Formylkynurenine, which are associated with tryptophan metabolism, as well as Stearidonic acid and Linolenelaidic acid, which are linked to α-linolenic acid metabolism, showed significant increases in the hepatic tissues of the GAM group. The changes in these metabolites suggest that protein and lipid metabolisms were more active in the livers of beef cattle in the GAM group than in the GAA group. This finding corresponds to the observed higher FBW and ADG in the GAM group.
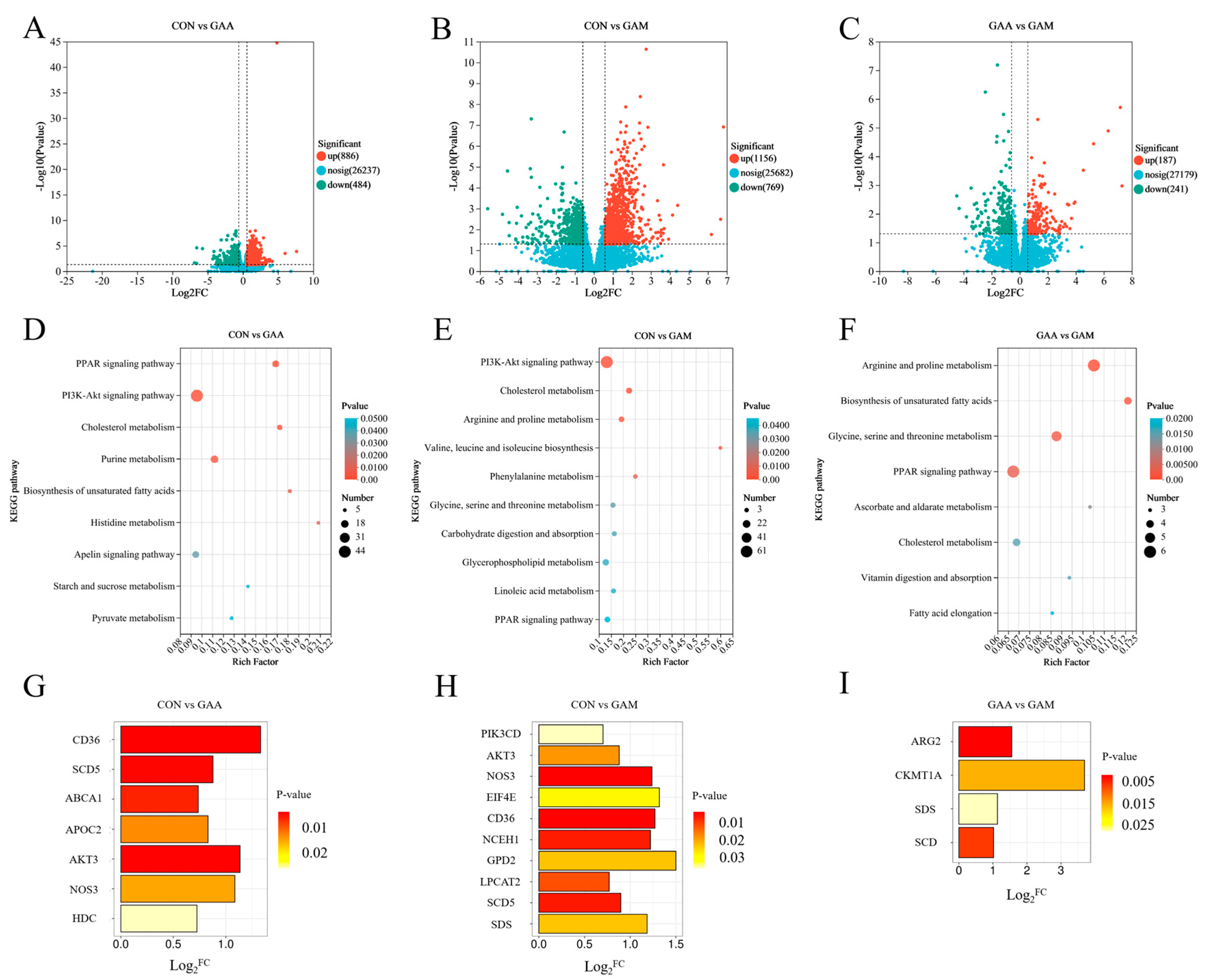
To explore the mechanisms underlying the observed metabolite changes, a transcriptomic analysis of liver tissues was conducted. The PI3K–AKT signaling pathway, which is known to regulate energy metabolism and protein synthesis in animals [
68,
69], was identified as a key contributor. In this pathway, the
AKT3 gene was significantly upregulated in the liver of beef cattle from both the GAA group and GAM group, while the
PIK3CD gene showed significant upregulation, specifically in the GAM group. This result indicates that the PI3K–AKT pathway was activated in both the GAA and GAM groups, suggesting that GAA supplementation promoted improved energy and protein metabolism in the hepatic tissues of beef cattle. This was flanked by the results showing that
AKT3 had significant positive correlations with Creatine, L-glutamic acid, and L-glutamine.
NOS3 is a downstream target gene of the PI3K–AKT pathway that is responsible for the transcriptional regulation of
eNOS production.
eNOS, in turn, catalyzes the conversion of L-arginine and reactive oxygen species (O
2) into L-citrulline and nitric oxide (NO), a process that aids in the elimination of reactive oxygen species from the body [
70]. At the same time, NO helps to dilate blood vessels so that more blood flows through the liver [
71], which can bring more nutrients to the liver for conversion and metabolism. The significant upregulation of
NOS3 in the liver tissues of beef cattle from both the GAA group and GAM group indicates that GAA supplementation can elevate
NOS3 expression by activating the PI3K–AKT pathway, thereby contributing to the improvement of the liver’s antioxidant capacity and the enhancement of the overall nutrient metabolism in beef cattle. The positive correlations between
NOS3 and phospholipids, creatine metabolism-related metabolites, and L-glutamine were also confirmed. The gene
EIF4E, which plays a key role downstream in the PI3K–AKT signaling pathway and is essential for regulating protein synthesis [
72], was significantly upregulated in the GAM group compared with the control group. These findings imply that the simultaneous administration of GAA and Met could improve hepatic protein synthesis in beef cattle. Moreover, this study observed notable changes in genes associated with amino acid metabolism. For example,
HDC was notably elevated in the GAA group, where it contributed to histidine metabolism by converting histidine into histamine [
73]. In contrast,
SDS, which is involved in serine metabolism by facilitating the conversion of serine to pyruvate [
74], exhibited significant upregulation, specifically in the GAM group. The gene expression patterns suggest that GAA supplementation enhances amino acid metabolism in the liver, and the addition of Met may further increase the protein synthesis efficiency.
Significant enrichment of both the PPAR signaling pathway and cholesterol metabolism was observed in the GAA and GAM groups. Central to the PPAR pathway are genes like
CD36 and
SCD5.
CD36 acts as a key regulator of fatty acid sensing and plays a pivotal role in modulating lipid metabolism [
75], while
SCD5 is essential as a rate-limiting enzyme for monounsaturated fatty acid production. Specifically,
SCD5 facilitates the conversion of stearoyl coenzyme A (C18:0) and palmitoyl coenzyme A (C16:0) into oleic acid (C18:1) and palmitoleic acid (C16:1), respectively [
76]. In the current research, both
CD36 and
SCD5 exhibited significant upregulation in the GAA and GAM groups, suggesting heightened lipid metabolic activity. These genes were also associated with specific lipid-related metabolites, including LysoPC (20:2(11Z,14Z)/0:0) and LysoPC (20:5(5Z,8Z,11Z,14Z,17Z)/0:0). These metabolites exhibited a significant positive correlation with one another. These findings collectively suggest that GAA supplementation effectively regulates hepatic lipid metabolism. In terms of cholesterol metabolism,
ABCA1 and
APOC2 were notably upregulated in the GAA group, whereas
NCEH1 showed significant upregulation in the GAM group. Additionally, genes involved in glycerophospholipid metabolism, such as
GPD2 and
LPCAT2, were significantly upregulated in the GAM group. These gene expression changes further support the regulatory role of GAA supplementation on lipid metabolism in beef cattle livers. Additionally, the expression of
SCD, a gene linked to the PPAR pathway, was notably higher in the GAM group compared with the GAA group, highlighting the added effect of Met supplementation. Analogously,
ARG2 and
CKMT1A, genes associated with arginine and proline metabolism, along with
SDS, a gene involved in serine metabolism, all displayed significant upregulation in the GAM group. These gene expression alterations collectively suggest that Met supplementation can further potentiate lipid metabolism and amino acid metabolism in the livers of beef cattle. Furthermore, additional transcriptomic and metabolomic analyses of the longissimus lumborum (LL) muscle revealed that dietary GAA supplementation improved the meat quality, antioxidant capacity, and modulated nutritional and fatty acid profiles. The addition of RPM further enhanced the antioxidant capacity and altered the fatty acid composition. These findings were associated with significant changes in genes and metabolites in amino acid and lipid metabolic pathways [
27]. Thus, GAA and RPM not only affect liver metabolism but also have meaningful effects on muscle metabolism, supporting improved growth performance and meat quality. Furthermore, given that beef is ultimately intended for human consumption, it is important to evaluate not only the effects of dietary interventions on animal performance and metabolism but also their potential impacts on the meat quality and food safety. Although the amount of methionine supplementation in this study was within the nutritional recommendations and physiological suitability for ruminants, the long-term consequences of such interventions—particularly the accumulation of amino acids or related metabolites in edible tissues—need careful consideration. Although no adverse effects on the liver function were observed in the current trial and improvements in the antioxidant capacity and growth performance were evident, further studies are needed to assess the nutritional safety of methionine-enriched meat products for human consumption. Future research should combine comprehensive food safety assessments with nutritional assessments relevant to humans to ensure that dietary supplementation strategies for livestock not only benefit production efficiency but also comply with public health considerations.


 ,
, 

{kind=link}
{kind=link}
{kind=link}