

A Highly Potent Apomorphine Derivative Enhancing Neurite Outgrowth via Nrf2 Activation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
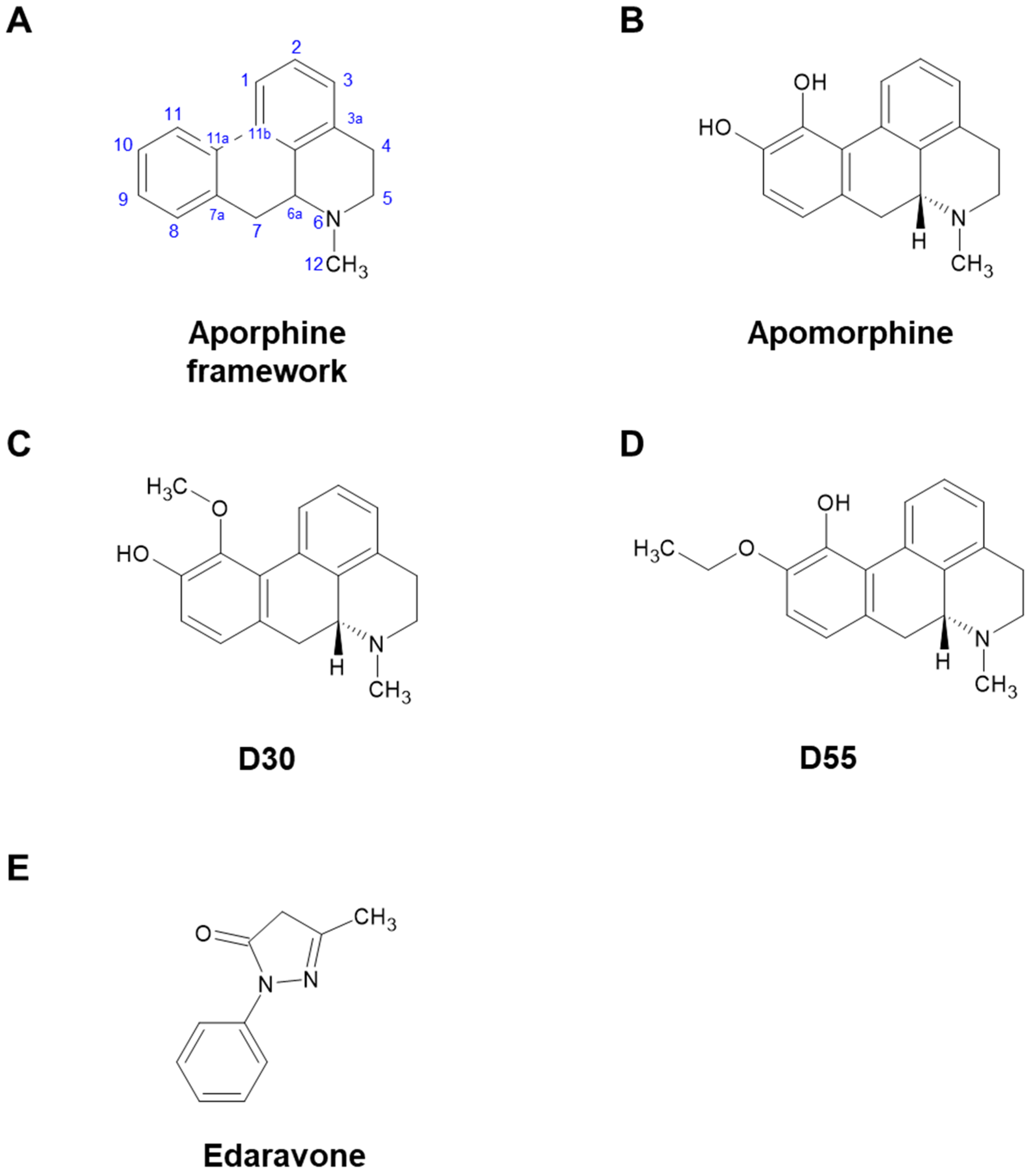
2.1. Compounds
2.2. Quantification of Neurite Outgrowth in PC12 Cells
2.3. Statistical Analysis
3. Results
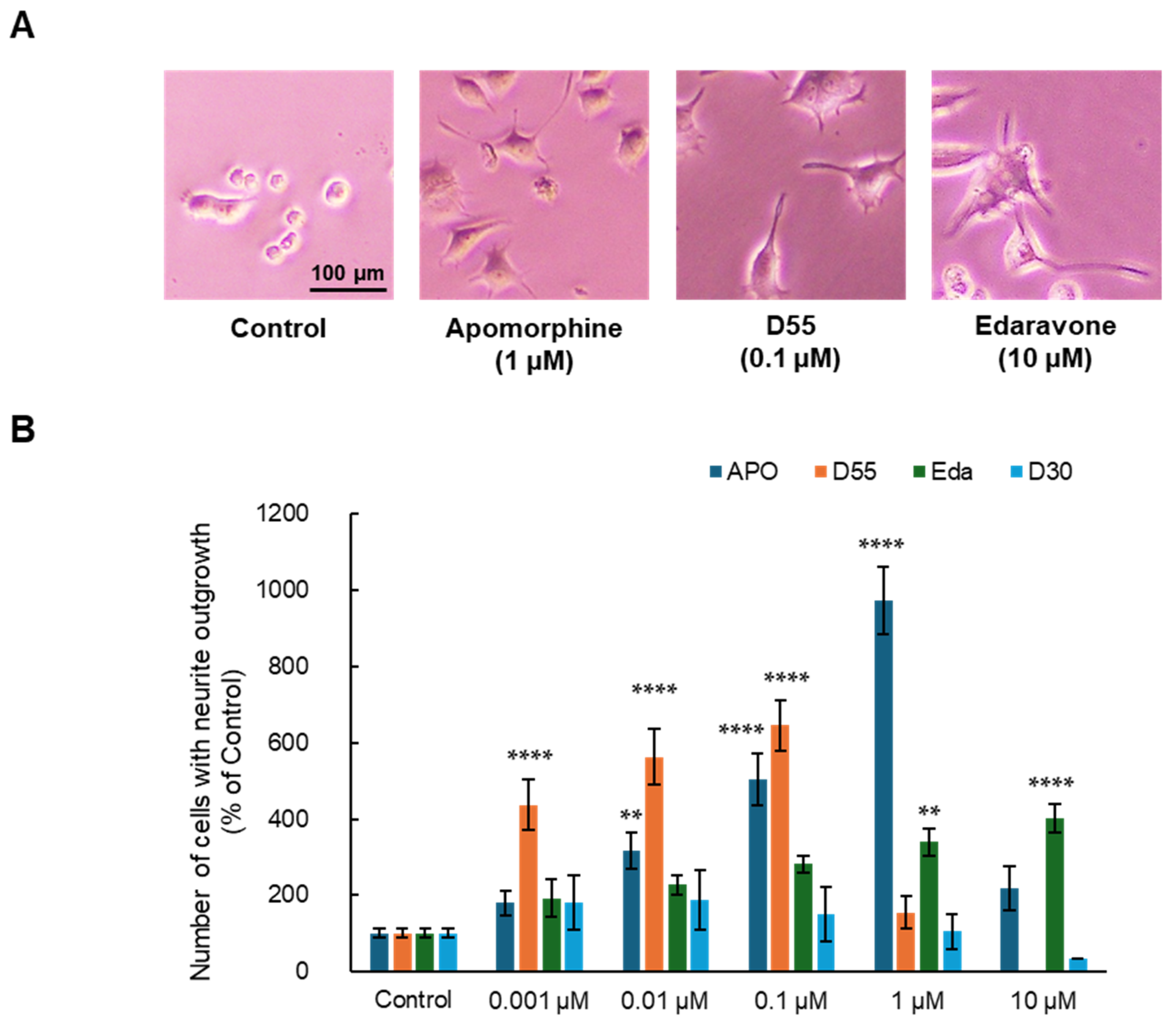
3.1. Effects of APO, Its Derivatives, and Eda on NGF-Induced Neurite Outgrowth in PC12 Cells
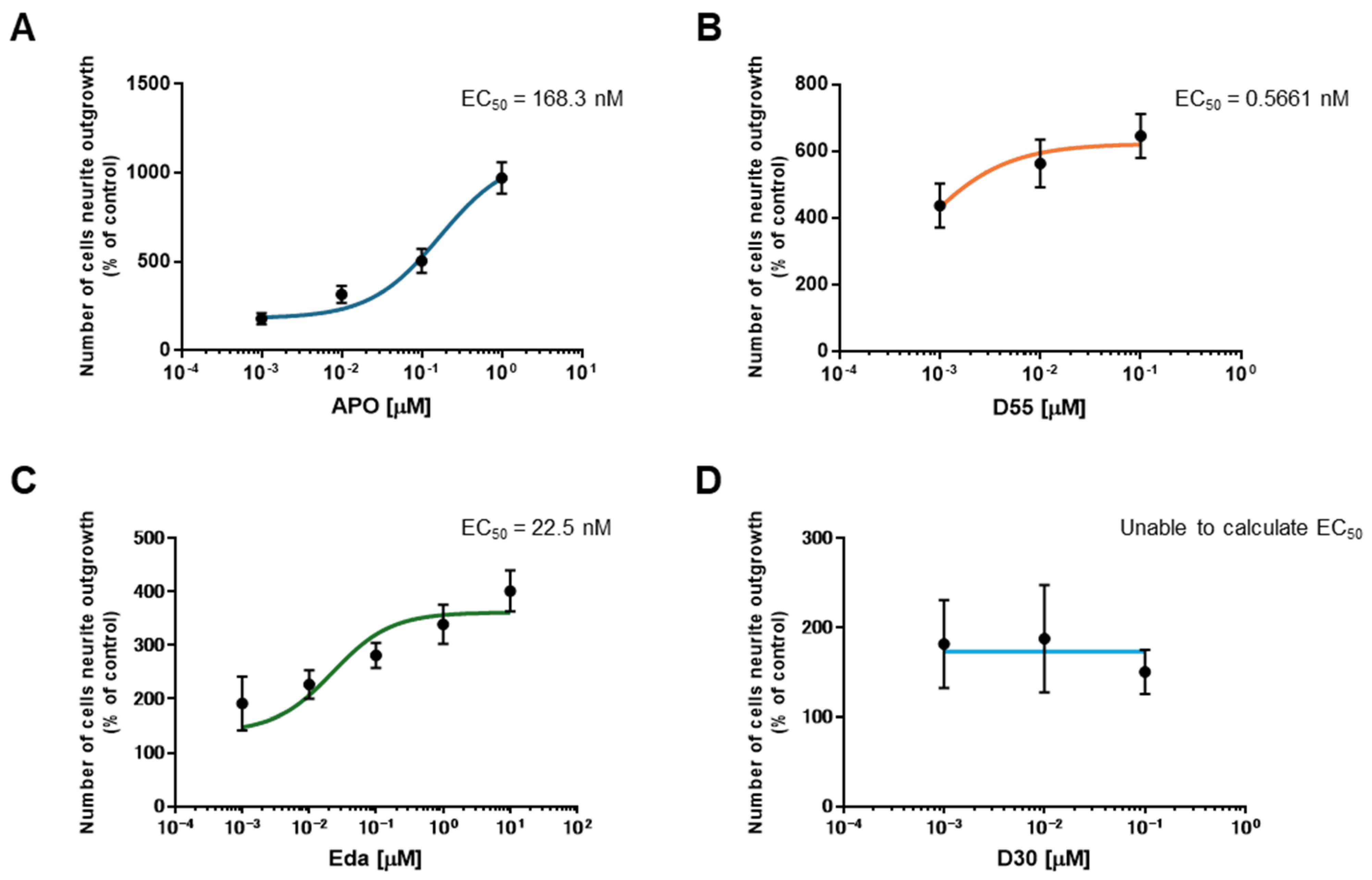
3.2. EC50 of APO, Its Derivatives, and Eda
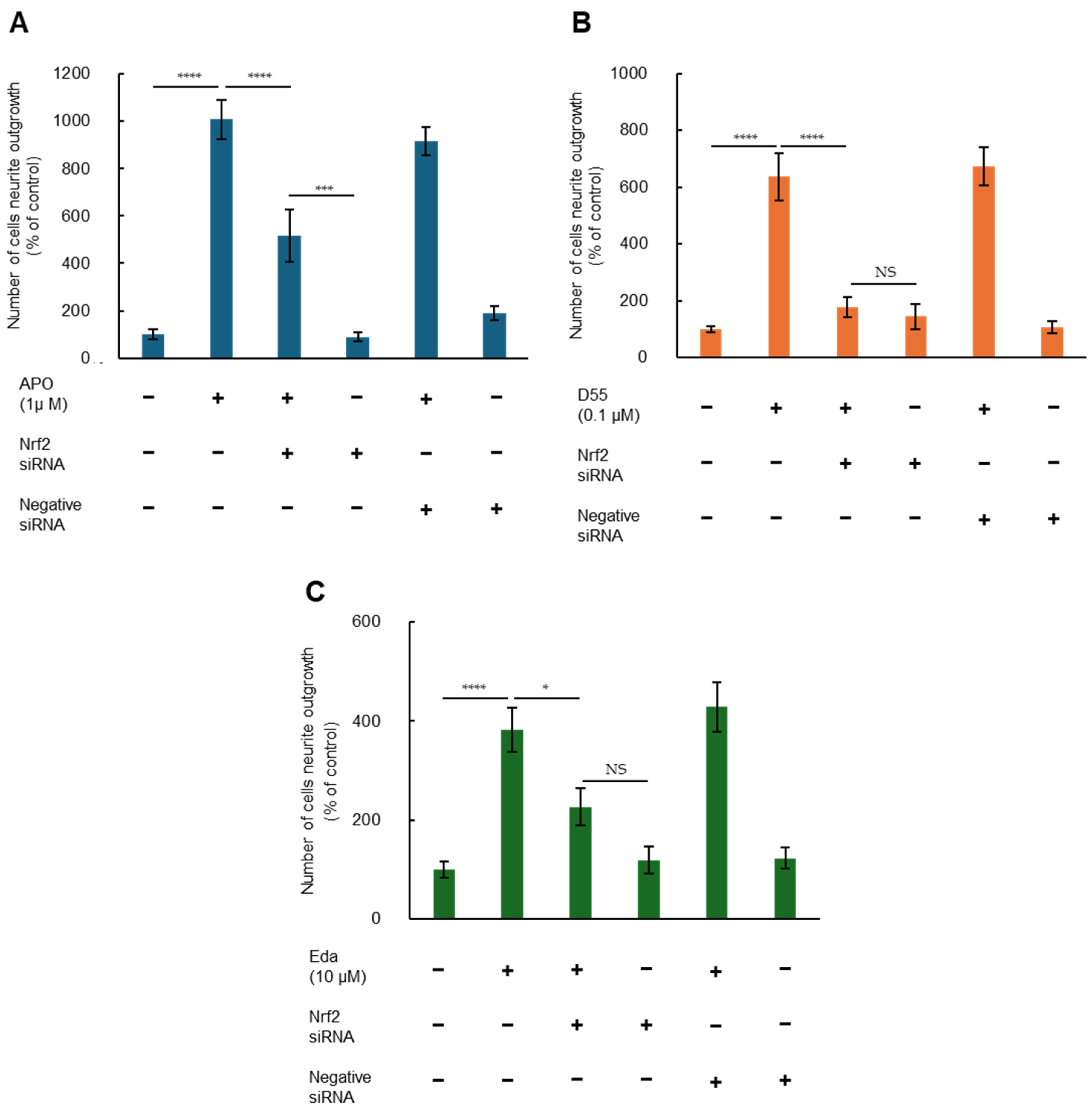
3.3. Role of Nrf2 in Potentiating NGF-Induced Neurite Outgrowth by APO, D55, and Eda
4. Discussion
4.1. Neuroprotective Effect of APO
4.2. Cell-Based In Vitro Model Using PC12 Cells
4.3. Neurite Outgrowth and Nrf2
4.4. Neuroplasticity
4.5. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APO | Apomorphine |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| Eda | Edaravone |
| DA | dopaminergic |
| ARE | Antioxidant Response Element |
| ALS | Amyotrophic lateral sclerosis |
| NGF | Nerve Growth Factor |
| MASH | Metabolic dysfunction steatohepatitis |
| 5-HT | 5-hydroxytryptamine |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
References
- Auffret, M.; Drapier, S.; Vérin, M. Pharmacological Insights into the Use of Apomorphine in Parkinson’s Disease: Clinical Relevance. Clin. Drug Investig. 2018, 38, 287–312. [Google Scholar] [CrossRef]
- Ribarič, S. The pharmacological properties and therapeutic use of apomorphine. Molecules 2012, 17, 5289–5309. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, A.; Kouga, T.; Jimbo, E.F.; Matsuhashi, T.; Abe, T.; Yamagata, T.; Osaka, H. Apomorphine rescues reactive oxygen species-induced apoptosis of fibroblasts with mitochondrial disease. Mitochondrion 2019, 49, 111–120. [Google Scholar] [CrossRef]
- Kobayashi, M.; Miyauchi, A.; Jimbo, E.F.; Oishi, N.; Aoki, S.; Watanabe, M.; Yoshikawa, Y.; Akiyama, Y.; Yamagata, T.; Osaka, H. Synthetic aporphine alkaloids are potential therapeutics for Leigh syndrome. Sci. Rep. 2024, 14, 11561. [Google Scholar] [CrossRef]
- Hara, H.; Ohta, M.; Adachi, T. Apomorphine protects against 6-hydroxydopamine-induced neuronal cell death through activation of the Nrf2-ARE pathway. J. Neurosci. Res. 2006, 84, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Miura, K.; Aizawa, K.; Bat-Erdene, O.; Sashikawa-Kimura, M.; Noguchi, E.; Watanabe, M.; Yamada, N.; Osaka, H.; Morimoto, N.; et al. Apomorphine Suppresses the Progression of Steatohepatitis by Inhibiting Ferroptosis. Antioxidants 2024, 13, 805. [Google Scholar] [CrossRef] [PubMed]
- Barati, A.; Moghimi, S.; Taghavi Zanjani, K.; Rohani, M.; Sohrabi Hesar, M.; Arfaie, A.; Ghezelche Khamsiyan, M.; Mahmoudi, J.; Sadigh-Eteghad, S. Acute Administration of Edaravone Improves Cognitive Impairment in a Mouse Model of mPFC Ischemia: Crosstalk Between Necroptosis, Neuroinflammation, and Antioxidant Defense. Mol. Neurobiol. 2025, 62, 4420–4434. [Google Scholar] [CrossRef]
- Bajpai, A.; Bharathi, V.; Patel, B.K. Therapeutic targeting of the oxidative stress generated by pathological molecular pathways in the neurodegenerative diseases, ALS and Huntington’s. Eur. J. Pharmacol. 2025, 987, 177187. [Google Scholar] [CrossRef]
- Soejima-Kusunoki, A.; Okada, K.; Saito, R.; Watabe, K. The Protective Effect of Edaravone on TDP-43 Plus Oxidative Stress-Induced Neurotoxicity in Neuronal Cells: Analysis of Its Neuroprotective Mechanisms Using RNA Sequencing. Pharmaceuticals 2022, 15, 842. [Google Scholar] [CrossRef]
- Hashimoto, K. Essential Role of Keap1-Nrf2 Signaling in Mood Disorders: Overview and Future Perspective. Front. Pharmacol. 2018, 9, 1182. [Google Scholar] [CrossRef]
- Meng, P.; Zhu, L.; Guo, J.; Li, Y.; Wei, Y.; Sun, J.; Zhu, J. Preparation of recombinant neuritin protein. Protein Expr. Purif. 2024, 223, 106554. [Google Scholar] [PubMed]
- Wang, F.; Wang, H.; Shan, X.; Mei, J.; Wei, P.; Song, Q.; Chen, W. High-strength and high-toughness ECM films with the potential for peripheral nerve repair. Biomed. Mater. 2023, 19, 015010. [Google Scholar] [CrossRef]
- Johansen, V.B.I.; Hampson, E.; Tsonou, E.; Pantarelli, C.; Chu, J.Y.; Crossland, L.; Okkenhaug, H.; Massey, A.J.; Hornigold, D.C.; Welch, H.C.E.; et al. The GPCR adaptor protein Norbin regulates S1PR1 trafficking and the morphology, cell cycle and survival of PC12 cells. Sci. Rep. 2023, 13, 18237. [Google Scholar] [CrossRef] [PubMed]
- Gassen, M.; Gross, A.; Youdim, M.B. Apomorphine enantiomers protect cultured pheochromocytoma (PC12) cells from oxidative stress induced by H2O2 and 6-hydroxydopamine. Mov. Disord. Off. J. Mov. Disord. Soc. 1998, 13, 661–667. [Google Scholar]
- Hashimoto, K.; Ishima, T. A novel target of action of minocycline in NGF-induced neurite outgrowth in PC12 cells: Translation initiation [corrected] factor eIF4AI. PLoS ONE 2010, 5, e15430. [Google Scholar]
- Ishima, T.; Iyo, M.; Hashimoto, K. Neurite outgrowth mediated by the heat shock protein Hsp90α: A novel target for the antipsychotic drug aripiprazole. Transl. Psychiatry 2012, 2, e170. [Google Scholar] [CrossRef]
- Ishima, T.; Futamura, T.; Ohgi, Y.; Yoshimi, N.; Kikuchi, T.; Hashimoto, K. Potentiation of neurite outgrowth by brexpiprazole, a novel serotonin-dopamine activity modulator: A role for serotonin 5-HT1A and 5-HT2A receptors. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2015, 25, 505–511. [Google Scholar]
- Yao, W.; Zhang, J.C.; Ishima, T.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Wu, J.; Suganuma, H.; et al. Role of Keap1-Nrf2 signaling in depression and dietary intake of glucoraphanin confers stress resilience in mice. Sci. Rep. 2016, 6, 30659. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, J.C.; Ishima, T.; Ren, Q.; Yang, C.; Dong, C.; Ma, M.; Saito, A.; Honda, T.; Hashimoto, K. Antidepressant effects of TBE-31 and MCE-1, the novel Nrf2 activators, in an inflammation model of depression. Eur. J. Pharmacol. 2016, 793, 21–27. [Google Scholar] [CrossRef]
- Yuan, H.; Sarre, S.; Ebinger, G.; Michotte, Y. Neuroprotective and neurotrophic effect of apomorphine in the striatal 6-OHDA-lesion rat model of Parkinson’s disease. Brain Res. 2004, 1026, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Fornai, F.; Battaglia, G.; Gesi, M.; Orzi, F.; Nicoletti, F.; Ruggieri, S. Dose-dependent protective effects of apomorphine against methamphetamine-induced nigrostriatal damage. Brain Res. 2001, 898, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Castri, P.; Busceti, C.; Battaglia, G.; Girardi, F.; Cavallari, M.; Orzi, F.; Fornai, F. Protection by apomorphine in two independent models of acute inhibition of oxidative metabolism in rodents. Clin. Exp. Hypertens. 2006, 28, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Maeda, A.; Kamiya, T.; Adachi, T. Protective effects of apomorphine against zinc-induced neurotoxicity in cultured cortical neurons. Biol. Pharm. Bull. 2013, 36, 585–591. [Google Scholar] [CrossRef]
- Ikram, H.; Zakir, R.; Haleem, D.J. Memory enhancing and neuroprotective effects of apomorphine in a rat model of dementia. Metab. Brain Dis. 2024, 39, 1051–1063. [Google Scholar] [CrossRef]
- Miyauchi, A.; Watanabe, C.; Yamada, N.; Jimbo, E.F.; Kobayashi, M.; Ohishi, N.; Nagayoshi, A.; Aoki, S.; Kishita, Y.; Ohtake, A.; et al. Apomorphine is a potent inhibitor of ferroptosis independent of dopaminergic receptors. Sci. Rep. 2024, 14, 4820. [Google Scholar] [CrossRef] [PubMed]
- Ogiso, H.; Miura, K.; Nagai, R.; Osaka, H.; Aizawa, K. Non-Targeted Metabolomics Reveal Apomorphine’s Therapeutic Effects and Lysophospholipid Alterations in Steatohepatitis. Antioxidants 2024, 13, 1293. [Google Scholar] [CrossRef]
- Gassen, M.; Glinka, Y.; Pinchasi, B.; Youdim, M.B. Apomorphine is a highly potent free radical scavenger in rat brain mitochondrial fraction. Eur. J. Pharmacol. 1996, 308, 219–225. [Google Scholar] [CrossRef]
- Greene, L.A.; Tischler, A.S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA. 1976, 73, 2424–2428. [Google Scholar] [CrossRef]
- Zachor, D.A.; Moore, J.F.; Brezausek, C.M.; Theibert, A.B.; Percy, A.K. Cocaine inhibition of neuronal differentiation in NGF-induced PC12 cells is independent of ras signaling. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2000, 18, 765–772. [Google Scholar] [CrossRef]
- Xiong, A.; Yan, A.L.; Bi, C.W.; Lam, K.Y.; Chan, G.K.; Lau, K.K.; Dong, T.T.; Lin, H.; Yang, L.; Wang, Z.; et al. Clivorine, an otonecine pyrrolizidine alkaloid from Ligularia species, impairs neuronal differentiation via NGF-induced signaling pathway in cultured PC12 cells. Phytomed. Int. J. Phytother. Phytopharm. 2016, 23, 931–938. [Google Scholar] [CrossRef]
- Yu, C.L.; Zhao, X.M.; Niu, Y.C. Ferulic Acid Protects Against Lead Acetate-Induced Inhibition of Neurite Outgrowth by Upregulating HO-1 in PC12 Cells: Involvement of ERK1/2-Nrf2 Pathway. Mol. Neurobiol. 2016, 53, 6489–6500. [Google Scholar] [CrossRef]
- Greene, L.A. Nerve growth factor prevents the death and stimulates the neuronal differentiation of clonal PC12 pheochromocytoma cells in serum-free medium. J. Cell Biol. 1978, 78, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Gassen, M.; Pergande, G.; Youdim, M.B. Antioxidant properties of the triaminopyridine, flupirtine. Biochem. Pharmacol. 1998, 56, 1323–1329. [Google Scholar] [CrossRef]
- Wei, H.; Leeds, P.R.; Qian, Y.; Wei, W.; Chen, R.; Chuang, D. beta-amyloid peptide-induced death of PC 12 cells and cerebellar granule cell neurons is inhibited by long-term lithium treatment. Eur. J. Pharmacol. 2000, 392, 117–123. [Google Scholar] [CrossRef]
- Matsumoto, G.; Stojanovic, A.; Holmberg, C.I.; Kim, S.; Morimoto, R.I. Structural properties and neuronal toxicity of am-yotrophic lateral sclerosis-associated Cu/Zn superoxide dismutase 1 aggregates. J. Cell Biol. 2005, 171, 75–85. [Google Scholar] [CrossRef]
- Homma, T.; Kobayashi, S.; Sato, H.; Fujii, J. Edaravone, a free radical scavenger, protects against ferroptotic cell death in vitro. Exp. Cell Res. 2019, 384, 111592. [Google Scholar] [CrossRef] [PubMed]
- Picada, J.N.; Roesler, R.; Henriques, J.A. Genotoxic, neurotoxic and neuroprotective activities of apomorphine and its oxidized derivative 8-oxo-apomorphine. Braz. J. Med. Biol. Res. 2005, 38, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; De Fazio, S.; De Fazio, P.; Amorosi, A.; Perrotta, I.; De Sarro, G.; Donato, G. Apomorphine-induced neurodegen-eration in Mongolian gerbil hippocampus. Schizophr. Res. 2007, 95, 223–227. [Google Scholar] [CrossRef]
- Lee, J.Y.; Ham, J.; Lim, W.; Song, G. Apomorphine induces mitochondrial-dysfunction-dependent apoptosis in choriocarcinoma. Reproduction 2020, 160, 367–377. [Google Scholar] [CrossRef]
- Yu, C.; Pan, S.; Dong, M.; Niu, Y. Astragaloside IV attenuates lead acetate-induced inhibition of neurite outgrowth through activation of Akt-dependent Nrf2 pathway in vitro. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1195–1203. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Hirata, N.; Kurone, Y.; Shimoeda, S. Edaravone is a Therapeutic Candidate for Doxorubicin-Induced Cardiomyopathy by Activating the Nrf2 Pathway. Pharmacol. Res. Perspect. 2025, 13, e70066. [Google Scholar] [CrossRef] [PubMed]
- Dang, R.; Wang, M.; Li, X.; Wang, H.; Liu, L.; Wu, Q.; Zhao, J.; Ji, P.; Zhong, L.; Licinio, J.; et al. Edaravone ameliorates de-pressive and anxiety-like behaviors via Sirt1/Nrf2/HO-1/Gpx4 pathway. J. Neuroinflamm. 2022, 19, 41. [Google Scholar] [CrossRef]
- Newman-Tancredi, A.; Cussac, D.; Quentric, Y.; Touzard, M.; Verrièle, L.; Carpentier, N.; Millan, M.J. Differential actions of antiparkinson agents at multiple classes of monoaminergic receptor. III. Agonist and antagonist properties at serotonin, 5-HT(1) and 5-HT(2), receptor subtypes. J. Pharmacol. Exp. Ther. 2002, 303, 815–822. [Google Scholar] [CrossRef]
- Sethi, P.; Mehan, S.; Khan, Z.; Maurya, P.K.; Kumar, N.; Kumar, A.; Tiwari, A.; Sharma, T.; Das Gupta, G.; Narula, A.S.; et al. The SIRT-1/Nrf2/HO-1 axis: Guardians of neuronal health in neurological disorders. Behav. Brain Res. 2025, 476, 115280. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.R.; Jee, H.J.; Jung, Y.S. Neuroprotective Effect of β-Lapachone against Glutamate-Induced Injury in HT22 Cells. Biomol. Ther. 2025, 33, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Grünblatt, E.; Mandel, S.; Gassen, M.; Youdim, M.B. Potent neuroprotective and antioxidant activity of apomorphine in MPTP and 6-hydroxydopamine induced neurotoxicity. J. Neural Transm. Suppl. 1999, 55, 57–70. [Google Scholar]
- Kim, S.; Indu Viswanath, A.N.; Park, J.H.; Lee, H.E.; Park, A.Y.; Choi, J.W.; Kim, H.J.; Londhe, A.M.; Jang, B.K.; Lee, J.; et al. Nrf2 activator via interference of Nrf2-Keap1 interaction has antioxidant and anti-inflammatory properties in Parkinson’s disease animal model. Neuropharmacology 2020, 167, 107989. [Google Scholar] [CrossRef]
- Kaur, M.; Aran, K.R. Unraveling the role of Nrf2 in dopaminergic neurons: A review of oxidative stress and mitochondrial dysfunction in Parkinson’s disease. Metab. Brain Dis. 2025, 40, 123. [Google Scholar] [CrossRef]
- Pajares, M.; Cuadrado, A.; Rojo, A.I. Modulation of proteostasis by transcription factor NRF2 and impact in neurodegenerative diseases. Redox Biol. 2017, 11, 543–553. [Google Scholar] [CrossRef]
- Bahn, G.; Park, J.S.; Yun, U.J.; Lee, Y.J.; Choi, Y.; Park, J.S.; Baek, S.H.; Choi, B.Y.; Cho, Y.S.; Kim, H.K.; et al. NRF2/ARE pathway negatively regulates BACE1 expression and ameliorates cognitive deficits in mouse Alzheimer’s models. Proc. Natl. Acad. Sci. USA 2019, 116, 12516–12523. [Google Scholar] [CrossRef]
- Hiramatsu, G.; Mizutani, R.; Toume, K.; Inada, Y.; Sawahata, M.; Uta, D.; Komatsu, K.; Kume, T. Preventive Effects of Psora-leae Semen Extracts on Cognitive Dysfunction in Alzheimer’s Disease Model App(NL-P-F) Mice. Biol. Pharm. Bull. 2025, 48, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Arthur, R.; Navik, U.; Kumar, P. Artemisinin Ameliorates the Neurotoxic Effect of 3-Nitropropionic Acid: A Possible Involvement of the ERK/BDNF/Nrf2/HO-1 Signaling Pathway. Mol. Neurobiol. 2025, 62, 3583–3600. [Google Scholar] [CrossRef]
- Tucci, P.; Lattanzi, R.; Severini, C.; Saso, L. Nrf2 Pathway in Huntington’s Disease (HD): What Is Its Role? Int. J. Mol. Sci. 2022, 23, 15272. [Google Scholar] [CrossRef]
- Yang, B.; Pan, J.; Zhang, X.N.; Wang, H.; He, L.; Rong, X.; Li, X.; Peng, Y. NRF2 activation suppresses motor neuron ferroptosis induced by the SOD1(G93A) mutation and exerts neuroprotection in amyotrophic lateral sclerosis. Neurobiol. Dis. 2023, 184, 106210. [Google Scholar] [CrossRef]
- Ito, T.; Ohuchi, K.; Kurita, H.; Murakami, T.; Takizawa, S.; Fujimaki, A.; Murata, J.; Oida, Y.; Hozumi, I.; Kitaichi, K.; et al. Activated Fibroblast Growth Factor Receptor 1 Mitigated Poly-PR-Induced Oxidative Stress and Protein Translational Impairment. Biol. Pharm. Bull. 2025, 48, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Vo, A.T.T.; Khan, U.; Liopo, A.V.; Mouli, K.; Olson, K.R.; McHugh, E.A.; Tour, J.M.; Pooparayil Manoj, M.; Derry, P.J.; Kent, T.A. Harshly Oxidized Activated Charcoal Enhances Protein Persulfidation with Implications for Neurodegeneration as Exemplified by Friedreich’s Ataxia. Nanomaterials 2024, 14, 2007. [Google Scholar] [CrossRef]
- Wu, Y.L.; Sun, H.L.; Chang, J.C.; Lin, W.Y.; Chen, P.Y.; Chen, C.C.; Lee, L.Y.; Li, C.C.; Hsieh, M.; Chen, H.W.; et al. Erinacine A-Enriched Hericium erinaceus Mycelium Ethanol Extract Lessens Cellular Damage in Cell and Drosophila Models of Spinocerebellar Ataxia Type 3 by Improvement of Nrf2 Activation. Antioxidants 2024, 13, 1495. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Feng, H.Y.; Fan, C.N.; Wang, J.; Yuan, Z.Y.; Xu, G.H.; Li, C.F.; Huang, W.F.; Yi, L.T. Asiaticoside Attenuates Chronic Restraint Stress-Induced Hippocampal CA1 Neuronal Ferroptosis via Activating BDNF/Nrf2/GPX4 Signaling Pathway. Drug Des. Dev. Ther. 2025, 19, 793–810. [Google Scholar] [CrossRef]
- Herbet, M.; Szumełda, I.; Piątkowska-Chmiel, I.; Gawrońska-Grzywacz, M.; Dudka, J. Beneficial effects of combined admin-istration of fluoxetine and mitochondria-targeted antioxidant at in behavioural and molecular studies in mice model of de-pression. Behav. Brain Res. 2021, 405, 113185. [Google Scholar] [CrossRef]
- Shirai, Y.; Fujita, Y.; Hashimoto, R.; Ohi, K.; Yamamori, H.; Yasuda, Y.; Ishima, T.; Suganuma, H.; Ushida, Y.; Takeda, M.; et al. Dietary Intake of Sulforaphane-Rich Broccoli Sprout Extracts during Juvenile and Adolescence Can Prevent Phencyclidine-Induced Cognitive Deficits at Adulthood. PLoS ONE 2015, 10, e0127244. [Google Scholar] [CrossRef]
- Peng, Z.; Jia, Q.; Mao, J.; Jiang, S.; Ma, Q.; Luo, X.; An, Z.; Huang, A.; Ma, C.; Yi, Q. The role of ferroptosis and oxidative stress in cognitive deficits among chronic schizophrenia patients: A multicenter investigation. Schizophrenia 2025, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, R.; Minato, Y.; Maeda, S.; Yagi, H. Nrf2 phosphorylation contributes to acquisition of pericyte reprogramming via the PKCδ pathway. Neurobiol. Dis. 2025, 206, 106824. [Google Scholar] [CrossRef] [PubMed]
- Bink, D.I.; Ritz, K.; Mackaaij, C.; Stam, O.; Scheffer, S.; Mizee, M.R.; Ploegmakers, H.J.; van Het Hof, B.J.; de Boer, O.J.; Sluimer, J.C.; et al. Lack of intracranial atherosclerosis in various atherosclerotic mouse models. Vasc. Biol. 2025, 7, e230013. [Google Scholar] [CrossRef]
- Fang, J.; Wang, H.; Zhou, J.; Dai, W.; Zhu, Y.; Zhou, Y.; Wang, X.; Zhou, M. Baicalin provides neuroprotection in traumatic brain injury mice model through Akt/Nrf2 pathway. Drug Des. Dev. Ther. 2018, 12, 2497–2508. [Google Scholar] [CrossRef]
- Shi, G.; Cao, Y.; Xu, J.; Chen, B.; Zhang, X.; Zhu, Y.; Liu, L.; Liu, X.; Zhang, L.; Zhou, Y.; et al. Inhibition of S100A8/A9 ameliorates neuroinflammation by blocking NET formation following traumatic brain injury. Redox Biol. 2025, 81, 103532. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sanz, A.; Coronado-Albi, M.J.; Muñoz-Viana, R.; García-Merino, A.; Sánchez-López, A.J. Neuroprotective and Anti-Inflammatory Effects of Dimethyl Fumarate, Monomethyl Fumarate, and Cannabidiol in Neurons and Microglia. Int. J. Mol. Sci. 2024, 25, 13082. [Google Scholar] [CrossRef]
- El Ali, Z.; Ollivier, A.; Manin, S.; Rivard, M.; Motterlini, R.; Foresti, R. Therapeutic effects of CO-releaser/Nrf2 activator hybrids (HYCOs) in the treatment of skin wound, psoriasis and multiple sclerosis. Redox Biol. 2020, 34, 101521. [Google Scholar] [CrossRef]
- Tebay, L.E.; Robertson, H.; Durant, S.T.; Vitale, S.R.; Penning, T.M.; Dinkova-Kostova, A.T.; Hayes, J.D. Mechanisms of activation of the transcription factor Nrf2 by redox stressors, nutrient cues, and energy status and the pathways through which it attenuates degenerative disease. Free Radic. Biol. Med. 2015, 88 Pt B, 108–146. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishima, T.; Osaka, H.; Nagai, R.; Aizawa, K. A Highly Potent Apomorphine Derivative Enhancing Neurite Outgrowth via Nrf2 Activation. Antioxidants 2025, 14, 537. https://doi.org/10.3390/antiox14050537
Ishima T, Osaka H, Nagai R, Aizawa K. A Highly Potent Apomorphine Derivative Enhancing Neurite Outgrowth via Nrf2 Activation. Antioxidants. 2025; 14(5):537. https://doi.org/10.3390/antiox14050537
Chicago/Turabian StyleIshima, Tamaki, Hitoshi Osaka, Ryozo Nagai, and Kenichi Aizawa. 2025. "A Highly Potent Apomorphine Derivative Enhancing Neurite Outgrowth via Nrf2 Activation" Antioxidants 14, no. 5: 537. https://doi.org/10.3390/antiox14050537
APA StyleIshima, T., Osaka, H., Nagai, R., & Aizawa, K. (2025). A Highly Potent Apomorphine Derivative Enhancing Neurite Outgrowth via Nrf2 Activation. Antioxidants, 14(5), 537. https://doi.org/10.3390/antiox14050537