
Gross Antioxidant Capacity and Anti-Inflammatory Potential of Flavonol Oxidation Products: A Combined Experimental and Theoretical Study

 ,
,  ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemical Section
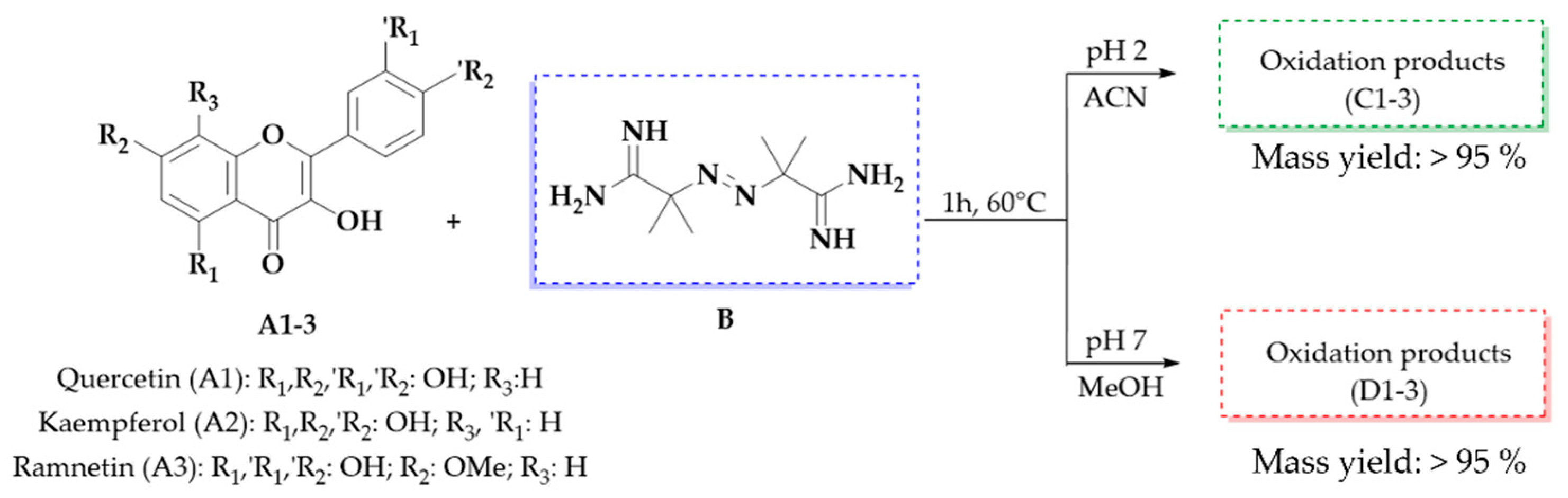
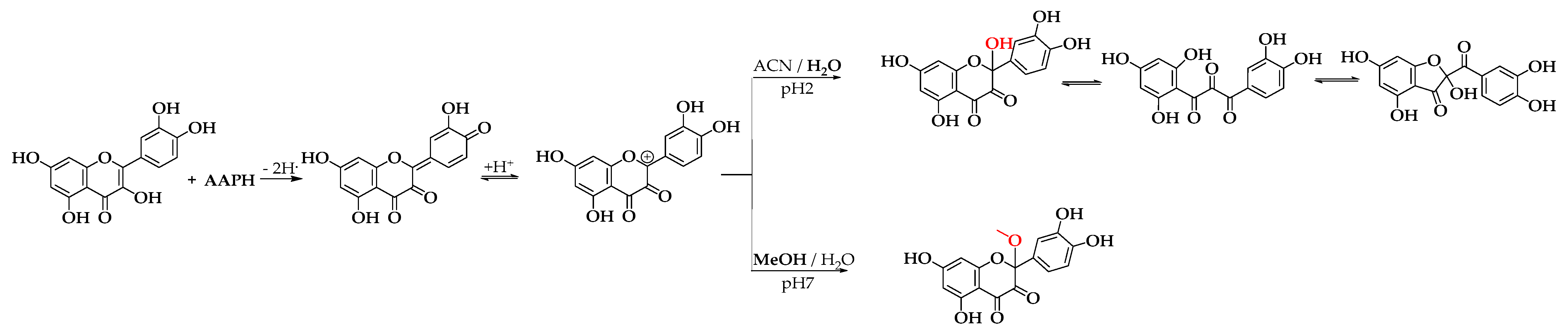
2.1.1. Obtaining Oxidation Products
2.1.2. HPLC-DAD Analysis
2.1.3. UPLC-ESI-MS/MS Analysis
2.2. Biological Section
2.2.1. ORAC-FL
2.2.2. ORAC-PGR
2.2.3. Viability Assay
2.2.4. Cellular Antioxidant Capacity (CAA) of Flavanols and Oxidation Products
2.2.5. Determination of COX-2 Expression by Western Blot
2.2.6. Statistical Analysis
2.3. Computational Studies
2.3.1. DFT Calculations
2.3.2. Molecular Docking
2.3.3. Molecular Dynamics Simulations and MM-GBSA Calculations
3. Results
3.1. Chemical Section
3.1.1. Obtaining Oxidation Products
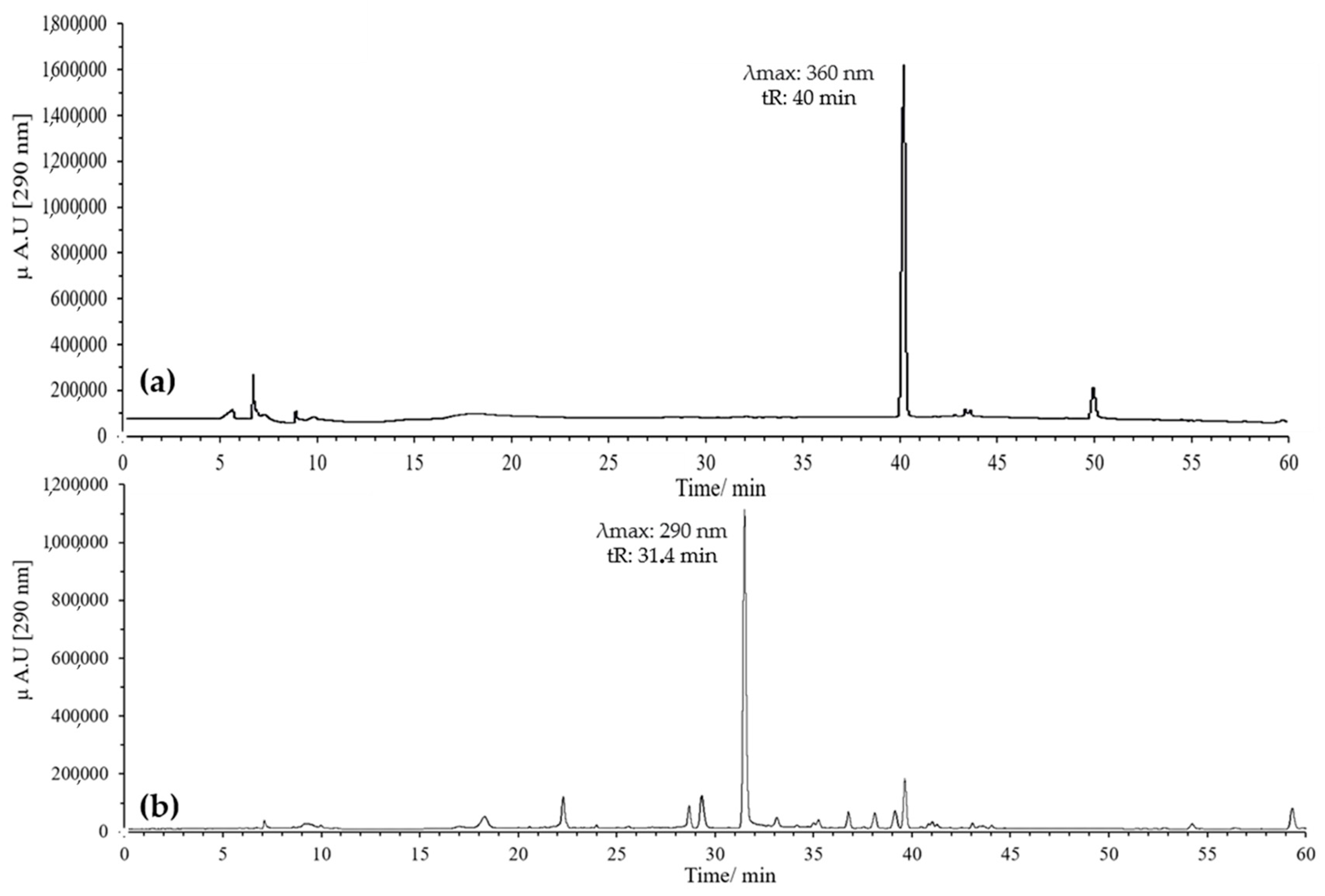
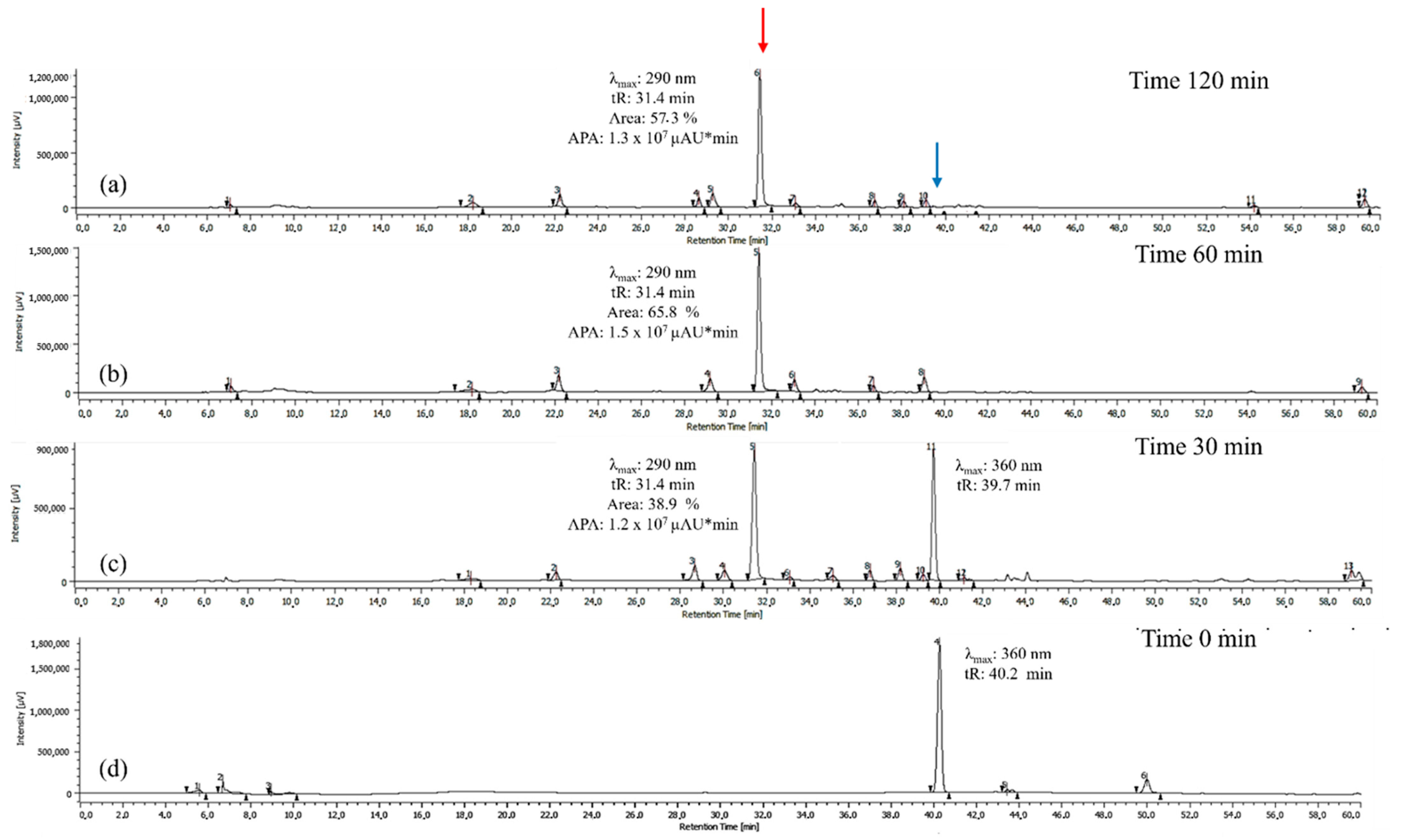
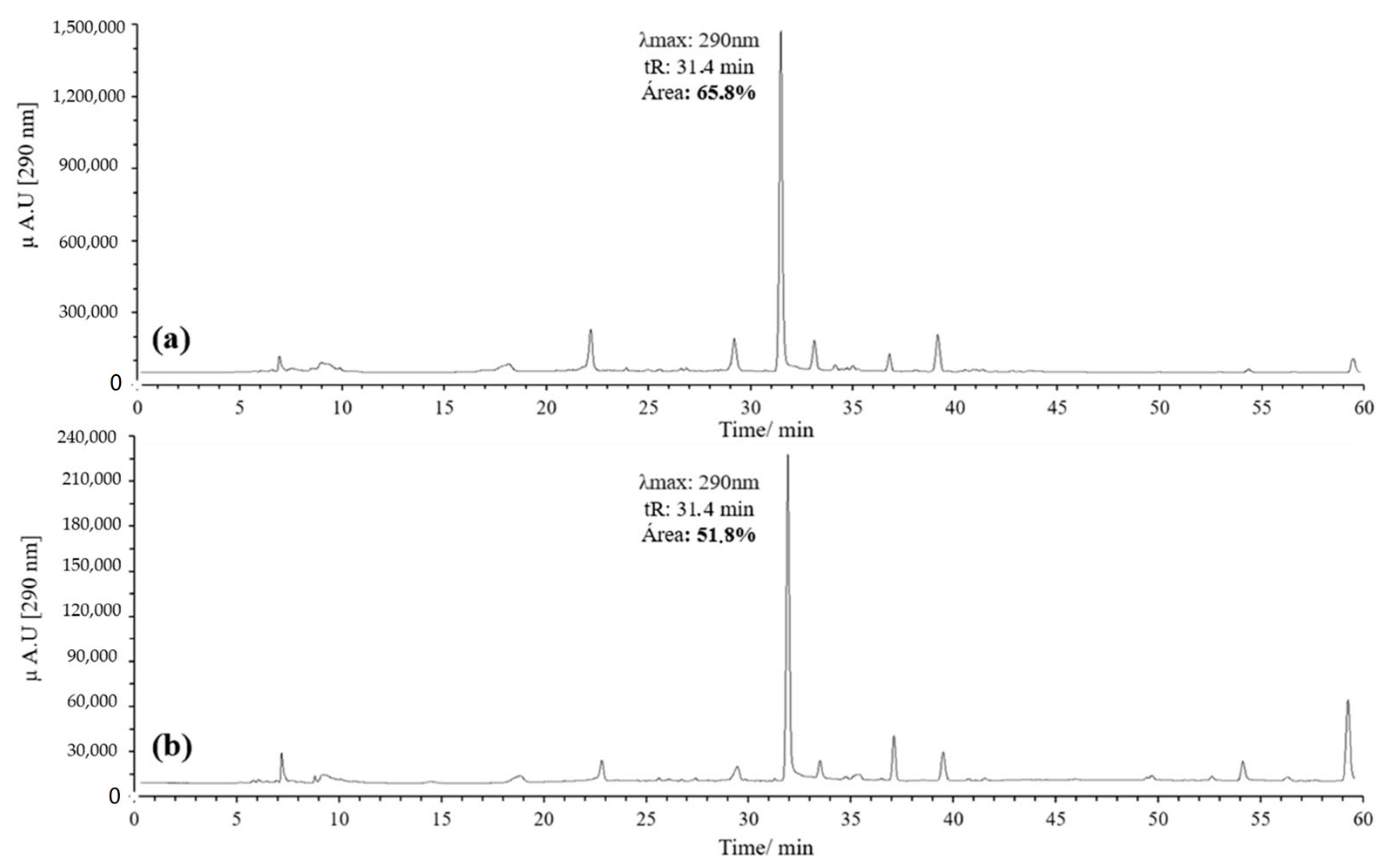
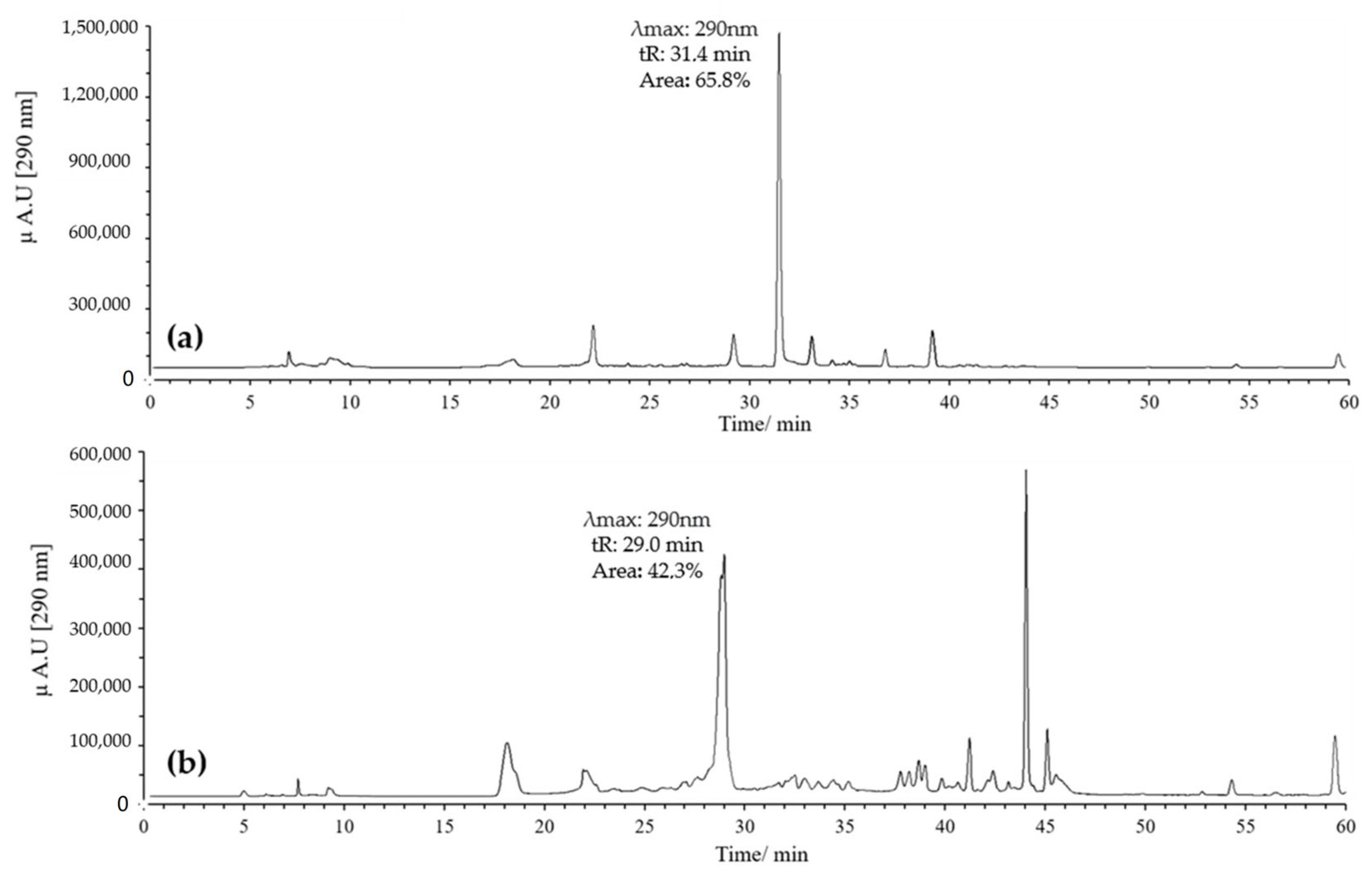
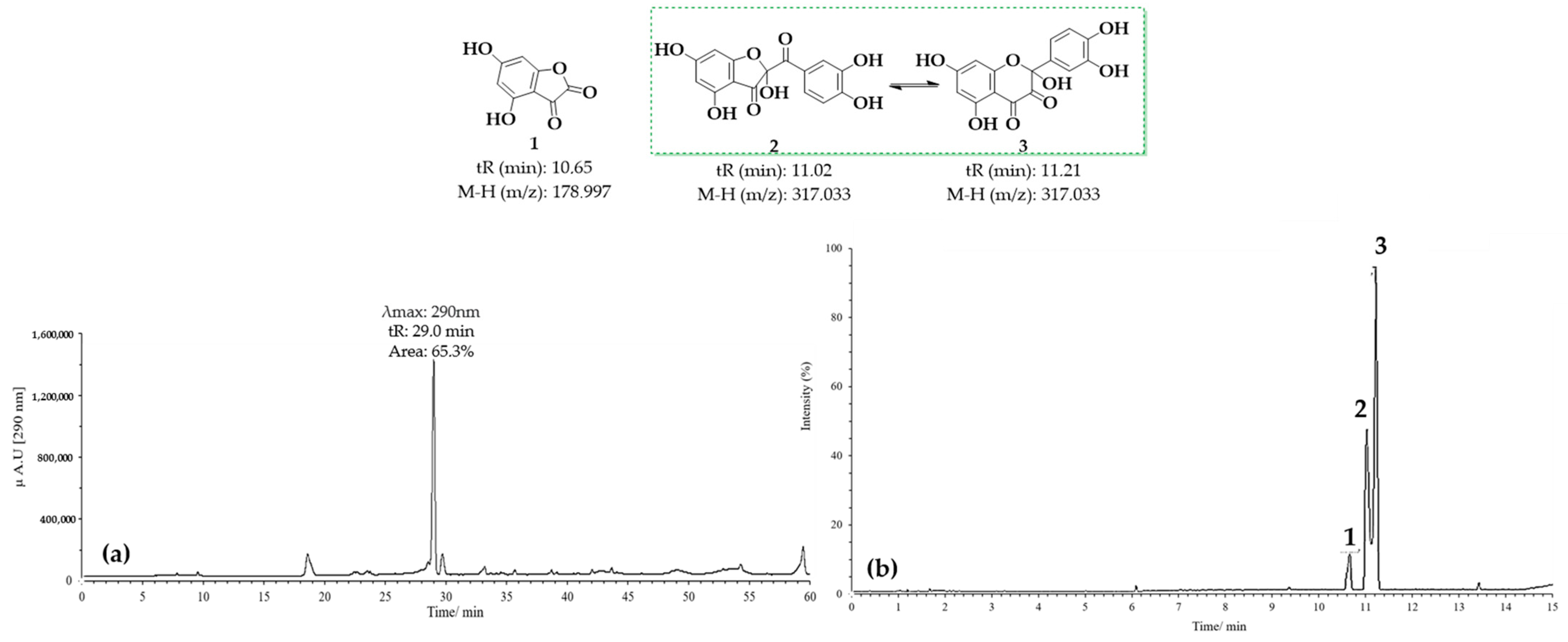
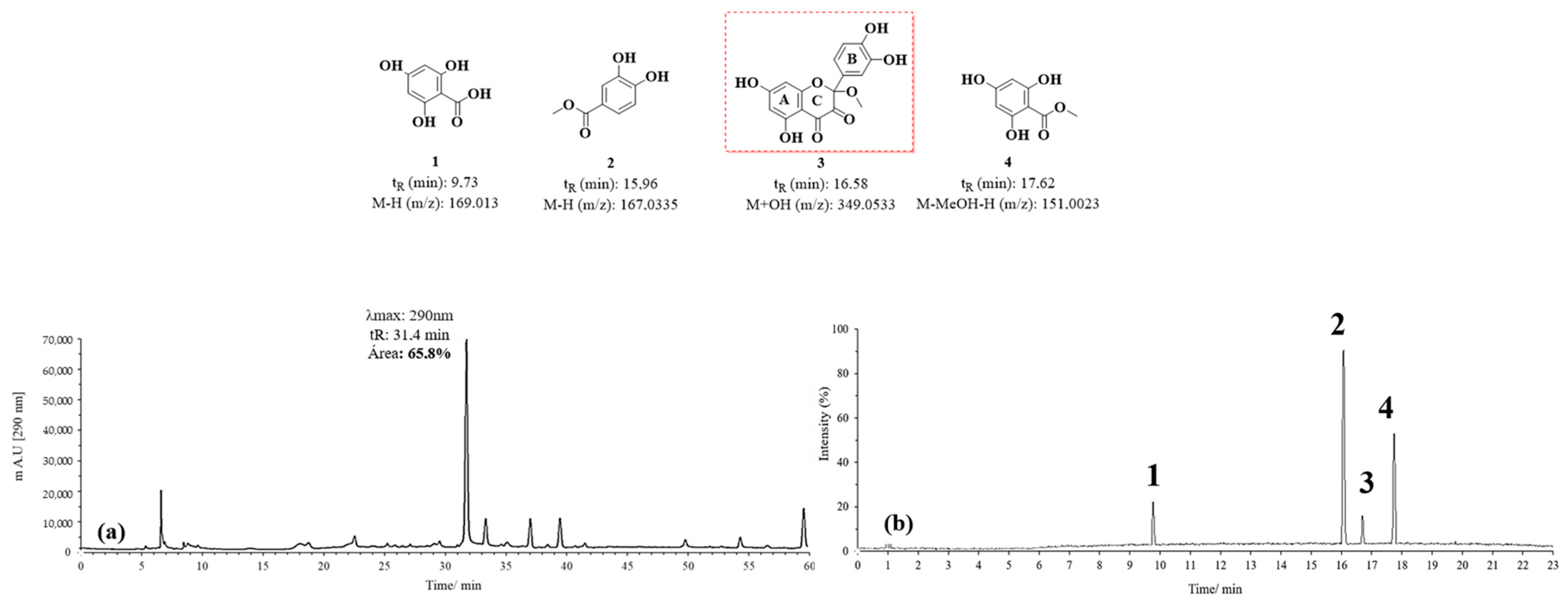
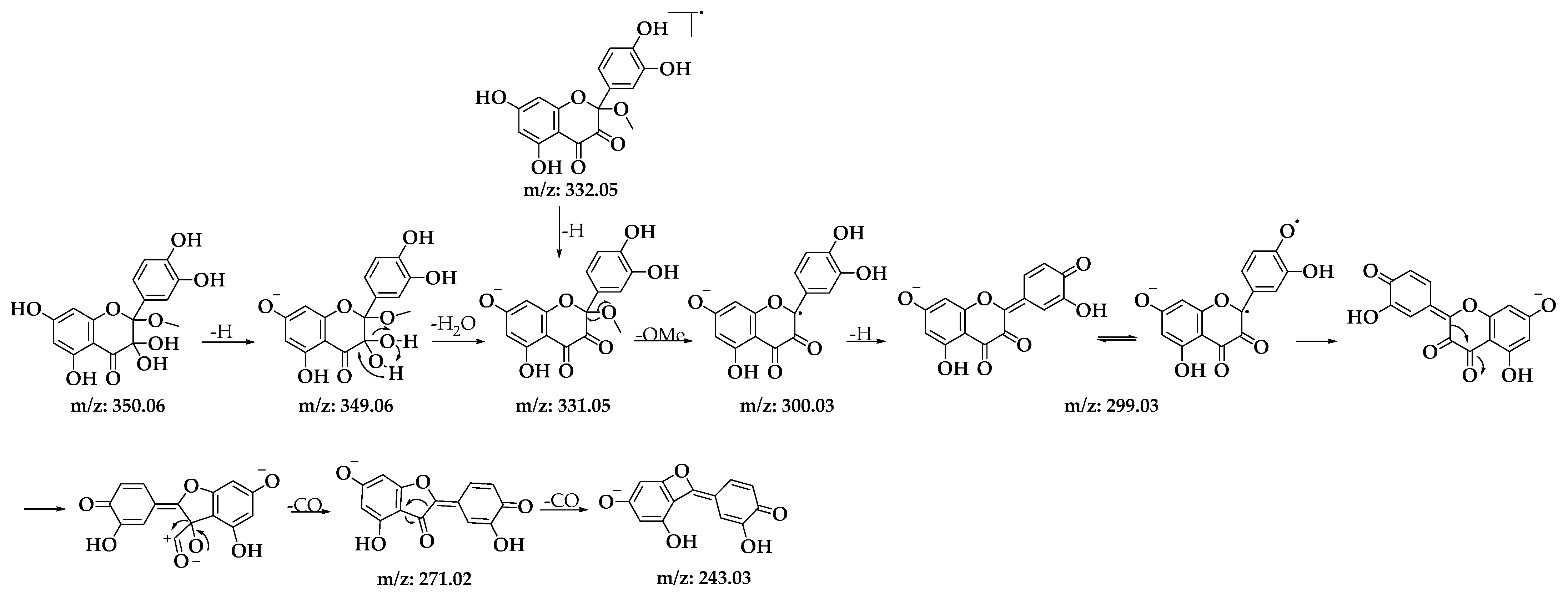
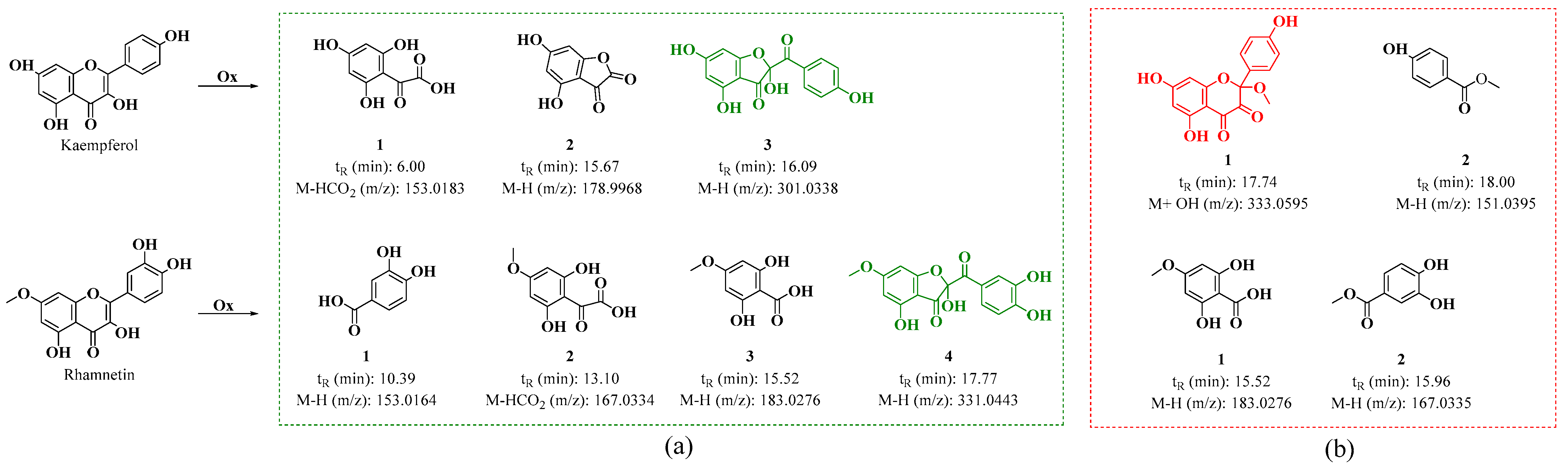
3.1.2. HPLC-DAD and UPLC-Ms/Ms Analysis of Oxidation Products
3.2. Biological Section
Analysis of the Antioxidant Capacity and Cell Viability of Oxidation Products
3.3. Computational Studies
3.3.1. DFT Analysis
3.3.2. Molecular Docking
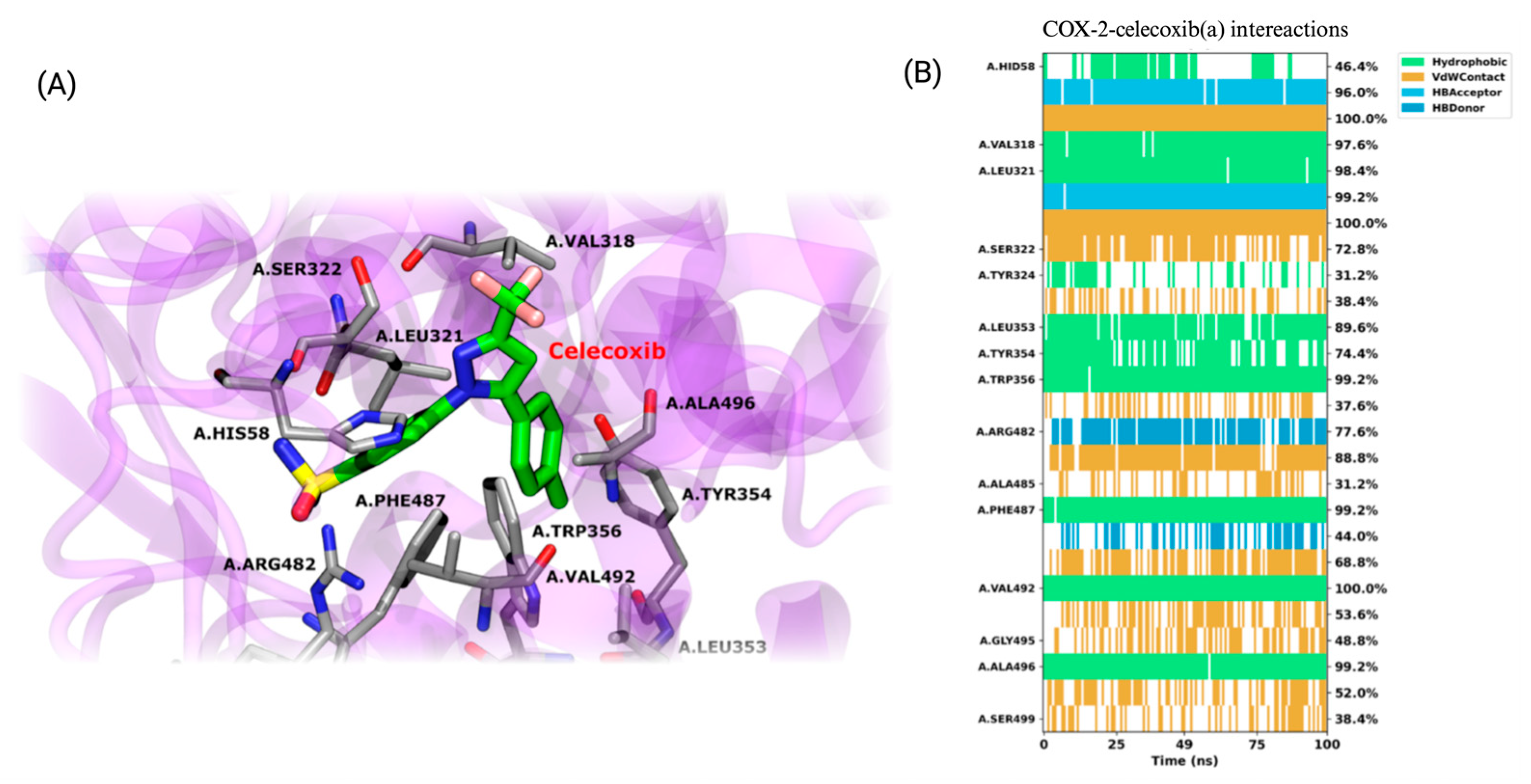
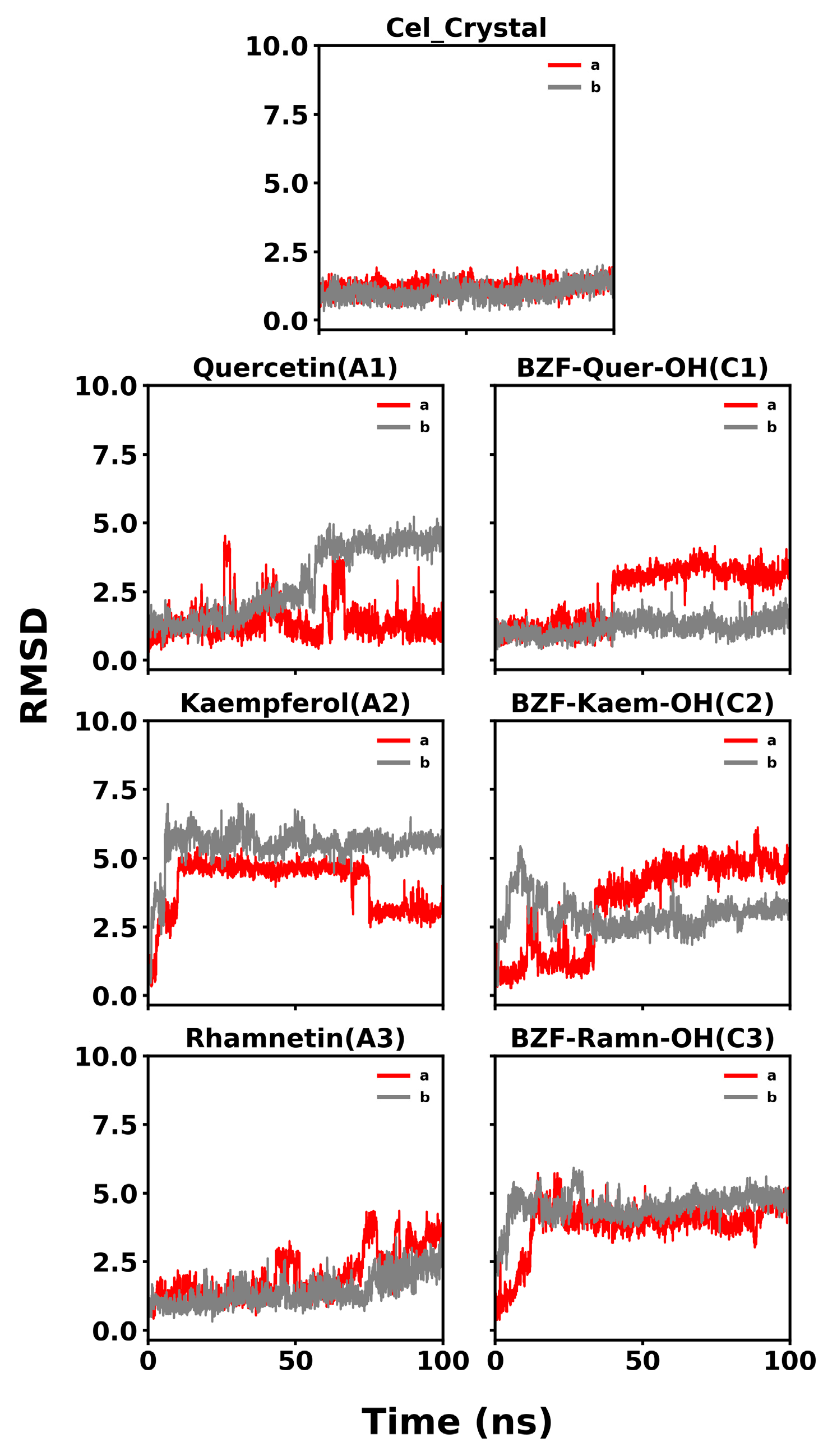
3.3.3. COX-2/Ligand Molecular Dynamics and MM-GBSA Calculations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Piacenza, L.; Trujillo, M.; Radi, R. Reactive Species and Pathogen Antioxidant Networks during Phagocytosis. J. Exp. Med. 2019, 216, 501–516. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Bulut, N.; Kocyigit, U.M.; Gecibesler, I.H.; Dastan, T.; Karci, H.; Taslimi, P.; Durna Dastan, S.; Gulcin, I.; Cetin, A. Synthesis of Some Novel Pyridine Compounds Containing Bis-1,2,4-Triazole/Thiosemicarbazide Moiety and Investigation of Their Antioxidant Properties, Carbonic Anhydrase, and Acetylcholinesterase Enzymes Inhibition Profiles. J. Biochem. Mol. Toxicol. 2018, 32, e22006. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Cetin Cakmak, K.; Gülçin, İ. Anticholinergic and Antioxidant Activities of Usnic Acid-an Activity-Structure Insight. Toxicol. Rep. 2019, 6, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Işık, M.; Beydemir, Ş.; Yılmaz, A.; Naldan, M.E.; Aslan, H.E.; Gülçin, İ. Oxidative Stress and MRNA Expression of Acetylcholinesterase in the Leukocytes of Ischemic Patients. Biomed. Pharmacother. 2017, 87, 561–567. [Google Scholar] [CrossRef]
- Krawczyk, H. The Stilbene Derivatives, Nucleosides, and Nucleosides Modified by Stilbene Derivatives. Bioorg. Chem. 2019, 90, 103073. [Google Scholar] [CrossRef]
- Stachowicz, K. Deciphering the Mechanisms of Regulation of an Excitatory Synapse via Cyclooxygenase-2. A Review. Biochem. Pharmacol. 2021, 192, 114729. [Google Scholar] [CrossRef]
- Polo-Cuadrado, E.; Acosta-Quiroga, K.; Rojas-Peña, C.; Rodriguez-Nuñez, Y.A.; Duarte, Y.; Brito, I.; Cisterna, J.; Gutiérrez, M. Molecular Modeling and Structural Analysis of Some Tetrahydroindazole and Cyclopentanepyrazole Derivatives as COX-2 Inhibitors. Arab. J. Chem. 2022, 15, 103540. [Google Scholar] [CrossRef]
- Muñoz, M.; Sánchez, A.; Pilar Martínez, M.; Benedito, S.; López-Oliva, M.E.; García-Sacristán, A.; Hernández, M.; Prieto, D. COX-2 Is Involved in Vascular Oxidative Stress and Endothelial Dysfunction of Renal Interlobar Arteries from Obese Zucker Rats. Free Radic. Biol. Med. 2015, 84, 77–90. [Google Scholar] [CrossRef]
- Onodera, Y.; Teramura, T.; Takehara, T.; Shigi, K.; Fukuda, K. Reactive Oxygen Species Induce Cox-2 Expression via TAK1 Activation in Synovial Fibroblast Cells. FEBS Open Bio 2015, 5, 492. [Google Scholar] [CrossRef] [PubMed]
- Virdis, A.; Bacca, A.; Colucci, R.; Duranti, E.; Fornai, M.; Materazzi, G.; Ippolito, C.; Bernardini, N.; Blandizzi, C.; Bernini, G.; et al. Endothelial Dysfunction in Small Arteries of Essential Hypertensive Patients: Role of Cyclooxygenase-2 in Oxidative Stress Generation. Hypertension 2013, 62, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Cordaro, M.; Siracusa, R.; D’amico, R.; Peritore, A.F.; Genovese, T.; Impellizzeri, D.; Di Paola, R.; Crupi, R.; Cuzzocrea, S.; et al. Novel Combination of COX-2 Inhibitor and Antioxidant Therapy for Modulating Oxidative Stress Associated with Intestinal Ischemic Reperfusion Injury and Endotoxemia. Antioxidants 2020, 9, 930. [Google Scholar] [CrossRef]
- Sies, H. Strategies of Antioxidant Defense. Eur. J. Biochem. 1993, 215, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Choi, S.W. Antioxidants in Food: Content, Measurement, Significance, Action, Cautions, Caveats, and Research Needs. Adv. Food Nutr. Res. 2014, 71, 1–53. [Google Scholar] [CrossRef]
- Hawrył, M.A.; Soczewiński, E. Separation of Some Flavonoids Using RP-HPLC-NP-TLC off-Line Coupled System. Chromatographia 2000, 52, 175–178. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Structure–Activity Relationships of Flavonoids in the Cellular Antioxidant Activity Assay. J. Agric. Food Chem. 2008, 56, 8404–8411. [Google Scholar] [CrossRef]
- Perez-Vizcaino, F.; Fraga, C.G. Research Trends in Flavonoids and Health. Arch. Biochem. Biophys. 2018, 646, 107–112. [Google Scholar] [CrossRef]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic Potential of Flavonoids in Cancer: ROS-Mediated Mechanisms. Biomed. Pharmacother. 2022, 146, 112442. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Miao, L.; Zhang, H.; Tan, L.; Zhao, Y.; Tu, Y.; Angel Prieto, M.; Simal-Gandara, J.; Chen, L.; He, C.; et al. Anti-Inflammatory Activity of Flavonols via Inhibiting MAPK and NF-ΚB Signaling Pathways in RAW264.7 Macrophages. Curr. Res. Food Sci. 2022, 5, 1176–1184. [Google Scholar] [CrossRef]
- López-Posadas, R.; Ballester, I.; Mascaraque, C.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; Sánchez De Medina, F. Flavonoids Exert Distinct Modulatory Actions on Cyclooxygenase 2 and NF-ΚB in an Intestinal Epithelial Cell Line (IEC18). Br. J. Pharmacol. 2010, 160, 1714. [Google Scholar] [CrossRef]
- Md Idris, M.H.; Mohd Amin, S.N.; Mohd Amin, S.N.; Nyokat, N.; Khong, H.Y.; Selvaraj, M.; Zakaria, Z.A.; Shaameri, Z.; Hamzah, A.S.; Teh, L.K.; et al. Flavonoids as Dual Inhibitors of Cyclooxygenase-2 (COX-2) and 5-Lipoxygenase (5-LOX): Molecular Docking and in Vitro Studies. Beni Suef Univ. J. Basic. Appl. Sci. 2022, 11, 117. [Google Scholar] [CrossRef]
- Banjarnahor, S.D.S.; Artanti, N. Antioxidant Properties of Flavonoids. Med. J. Indones. 2014, 23, 239–244. [Google Scholar] [CrossRef]
- Zeka, K.; Ruparelia, K.; Arroo, R.R.J.; Budriesi, R.; Micucci, M. Flavonoids and Their Metabolites: Prevention in Cardiovascular Diseases and Diabetes. Diseases 2017, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.; Arias-Santé, M.F.; Atala, E.; Pastene, E.; Kogan, M.J.; Speisky, H. Low Nanomolar Concentrations of a Quercetin Oxidation Product, Which Naturally Occurs in Onion Peel, Protect Cells against Oxidative Damage. Food Chem. 2020, 314, 126166. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.; Atala, E.; Pastene, E.; Carrasco-Pozo, C.; Speisky, H. Quercetin Oxidation Paradoxically Enhances Its Antioxidant and Cytoprotective Properties. J. Agric. Food Chem. 2017, 65, 11002–11010. [Google Scholar] [CrossRef]
- Speisky, H.; Shahidi, F.; Costa De Camargo, A.; Fuentes, J. Revisiting the Oxidation of Flavonoids: Loss, Conservation or Enhancement of Their Antioxidant Properties. Antioxidants 2022, 11, 133. [Google Scholar] [CrossRef]
- Krishnamachari, V.; Levine, L.H.; Paré, P.W. Flavonoid Oxidation by the Radical Generator AIBN: A Unified Mechanism for Quercetin Radical Scavenging. J. Agric. Food Chem. 2002, 50, 4357–4363. [Google Scholar] [CrossRef]
- Speisky, H.; Fuentes, J.; Atala, E.; Lopez, C. Antioxidant Activity of Quercetin-Oxidation Products. Free Radic. Biol. Med. 2015, 87, S96. [Google Scholar] [CrossRef]
- Speisky, H.; Fuentes, J.; Dorta, E.; Atala, E.; Lopez-Alarcon, C. Oxidation of Quercetin Can Markedly Enhance Its Antioxidant and Cytoprotective Properties. Free Radic. Biol. Med. 2016, 100, S110. [Google Scholar] [CrossRef]
- Atala, E.; Fuentes, J.; Wehrhahn, M.J.; Speisky, H. Quercetin and Related Flavonoids Conserve Their Antioxidant Properties despite Undergoing Chemical or Enzymatic Oxidation. Food Chem. 2017, 234, 479–485. [Google Scholar] [CrossRef]
- Fuentes, J.; Speisky, H.; De Camargo, A.C.; Atala, E.; Gotteland, M.; Olea-Azar, C. Quercetin Oxidation Metabolite Present in Onion Peel Protects Caco-2 Cells against the Oxidative Stress, Nf-Kb Activation, and Loss of Epithelial Barrier Function Induced by Nsaids. J. Agric. Food Chem. 2021, 69, 2157–2167. [Google Scholar] [CrossRef]
- Speisky, H.; Arias-Santé, M.F.; Fuentes, J. Oxidation of Quercetin and Kaempferol Markedly Amplifies Their Antioxidant, Cytoprotective, and Anti-Inflammatory Properties. Antioxidants 2023, 12, 155. [Google Scholar] [CrossRef] [PubMed]
- Pozo-Martínez, J.; Vázquez-Rodríguez, S.; Olea-Azar, C.; Moncada-Basualto, M. Evaluation of ORAC Methodologies in Determination of Antioxidant Capacity of Binary Combinations of Quercetin and 3-(3,4,5-Trihydroxybenzoyl) Coumarin Derivatives. Arab. J. Chem. 2022, 15, 104298. [Google Scholar] [CrossRef]
- López-Alarcón, C.; Lissi, E. A Novel and Simple ORAC Methodology Based on the Interaction of Pyrogallol Red with Peroxyl Radicals. Free Radic. Res. 2006, 40, 979–985. [Google Scholar] [CrossRef]
- Moncada-Basualto, M.; Saavedra-Olavarría, J.; Rivero-Jerez, P.S.; Rojas, C.; Maya, J.D.; Liempi, A.; Zúñiga-Bustos, M.; Olea-Azar, C.; Lapier, M.; Pérez, E.G.; et al. Assessment of the Activity of Nitroisoxazole Derivatives against Trypanosoma Cruzi. Molecules 2024, 29, 2762. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Rui, H.L. Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Lee, G.B.; Kim, Y.; Lee, K.E.; Vinayagam, R.; Singh, M.; Kang, S.G. Anti-Inflammatory Effects of Quercetin, Rutin, and Troxerutin Result From the Inhibition of NO Production and the Reduction of COX-2 Levels in RAW 264.7 Cells Treated with LPS. Appl. Biochem. Biotechnol. 2024, 196, 8431–8452. [Google Scholar] [CrossRef]
- Molina-Berríos, A.; Campos-Estrada, C.; Henriquez, N.; Faúndez, M.; Torres, G.; Castillo, C.; Escanilla, S.; Kemmerling, U.; Morello, A.; López-Muñoz, R.A.; et al. Protective Role of Acetylsalicylic Acid in Experimental Trypanosoma Cruzi Infection: Evidence of a 15-Epi-Lipoxin A4-Mediated Effect. PLoS Negl. Trop. Dis. 2013, 7, e2173. [Google Scholar] [CrossRef]
- Grimme, S.; Hansen, A.; Ehlert, S.; Mewes, J.M. R2SCAN-3c: A “Swiss Army Knife” Composite Electronic-Structure Method. J. Chem. Phys. 2021, 154, 064103. [Google Scholar] [CrossRef] [PubMed]
- Klamt, A. The COSMO and COSMO-RS Solvation Models. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2011, 1, 699–709. [Google Scholar] [CrossRef]
- Eichkorn, K.; Weigend, F.; Treutler, O.; Ahlrichs, R. Auxiliary Basis Sets for Main Row Atoms and Transition Metals and Their Use to Approximate Coulomb Potentials. Theor. Chem. Acc. 1997, 97, 119–124. [Google Scholar] [CrossRef]
- Furche, F.; Ahlrichs, R.; Hättig, C.; Klopper, W.; Sierka, M.; Weigend, F. Turbomole. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2014, 4, 91–100. [Google Scholar] [CrossRef]
- Neese, F. Software Update: The ORCA Program System—Version 5.0. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2022, 12, e1606. [Google Scholar] [CrossRef]
- Hernandez, D.A.; Rodriguez-Zavala, J.G.; Tenorio, F.J. DFT Study of Antioxidant Molecules from Traditional Japanese and Chinese Teas: Comparing Allylic and Phenolic Antiradical Activity. Struct. Chem. 2020, 31, 359–369. [Google Scholar] [CrossRef]
- Rohman, R.; Nath, R.; Kar, R. Revisiting the Hydrogen Atom Transfer Reactions through a Simple and Accurate Theoretical Model: Role of Hydrogen Bond Energy in Polyphenolic Antioxidants. Comput. Theor. Chem. 2023, 1223, 114097. [Google Scholar] [CrossRef]
- Parr, R.G.; Pearson, R.G. Absolute Hardness: Companion Parameter to Absolute Electronegativity. J. Am. Chem. Soc. 1983, 105, 7512–7516. [Google Scholar] [CrossRef]
- Pearson, R.G. Chemical Hardness and Density Functional Theory. J. Chem. Sci. 2005, 117, 369–377. [Google Scholar] [CrossRef]
- Osorio, E.; Pérez, E.G.; Areche, C.; Ruiz, L.M.; Cassels, B.K.; Flórez, E.; Tiznado, W. Why Is Quercetin a Better Antioxidant than Taxifolin? Theoretical Study of Mechanisms Involving Activated Forms. J. Mol. Model. 2013, 19, 2165–2172. [Google Scholar] [CrossRef] [PubMed]
- Ferraroni, M.; Matera, I.; Steimer, L.; Bürger, S.; Scozzafava, A.; Stolz, A.; Briganti, F. Crystal Structures of Salicylate 1,2-Dioxygenase-Substrates Adducts: A Step towards the Comprehension of the Structural Basis for Substrate Selection in Class III Ring Cleaving Dioxygenases. J. Struct. Biol. 2012, 177, 431–438. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.C.; Myers, J.B.; Folta, T.; Shoja, V.; Heath, L.S.; Onufriev, A. H++: A Server for Estimating p Ka s and Adding Missing Hydrogens to Macromolecules. Nucleic Acids Res. 2005, 33, W368–W371. [Google Scholar] [CrossRef]
- Feketeová, L.; Albright, A.L.; Sørensen, B.S.; Horsman, M.R.; White, J.; O’Hair, R.A.J.; Bassler, N. Formation of Radical Anions of Radiosensitizers and Related Model Compounds via Electrospray Ionization. Int. J. Mass Spectrom. 2014, 365–366, 56–63. [Google Scholar] [CrossRef]
- Morishetti, K.K.; Sripadi, P.; Mariappanadar, V.; Ren, J. Generation and Characterization of Distonic Dehydrophenoxide Radical Anions under Electrospray and Atmospheric Pressure Chemical Ionizations. Int. J. Mass Spectrom. 2011, 299, 169–177. [Google Scholar] [CrossRef]
- Holčapek, M.; Jirásko, R.; Lísa, M. Basic Rules for the Interpretation of Atmospheric Pressure Ionization Mass Spectra of Small Molecules. J. Chromatogr. A 2010, 1217, 3908–3921. [Google Scholar] [CrossRef]
- Tak, V.; Kanaujia, P.K.; Pardasani, D.; Kumar, R.; Srivastava, R.K.; Gupta, A.K.; Dubey, D.K. Application of Doehlert Design in Optimizing the Determination of Degraded Products of Nerve Agents by Ion-Pair Liquid Chromatography Electrospray Ionization Tandem Mass Spectrometry. J. Chromatogr. A 2007, 1161, 198–206. [Google Scholar] [CrossRef]
- Khairallah, G.; Peel, J.B. Identification of Dianions of C84 and C90 by Electrospray Mass Spectrometry. Chem. Phys. Lett. 1998, 296, 545–548. [Google Scholar] [CrossRef]
- Hiraoka, K.; Kudaka, I.; Fujimaki, S.; Shinohara, H. Observation of the Fullerene Anions C and C by Electrospray Ionization. Rapid Commun. Mass Spectrom. 1992, 6, 254–256. [Google Scholar] [CrossRef]
- López-Alarcón, C.; Aspée, A.; Lissi, E. Influence of the Target Molecule on the ORAC Index. ACS Symp. Ser. 2012, 1093, 417–429. [Google Scholar] [CrossRef]
- Dorta, E.; Fuentes-Lemus, E.; Aspée, A.; Atala, E.; Speisky, H.; Bridi, R.; Lissi, E.; López-Alarcón, C. The ORAC (Oxygen Radical Absorbance Capacity) Index Does Not Reflect the Capacity of Antioxidants to Trap Peroxyl Radicals. RSC Adv. 2015, 5, 39899–39902. [Google Scholar] [CrossRef]
- Dorta, E.; Atala, E.; Aspee, A.; Speisky, H.; Lissi, E.; Lopez-Alarcon, C. Role of Alkoxyl Radicals on the Fluorescein-Based ORAC (Oxygen Radical Absorbance Capacity) Assay. Free Radic. Biol. Med. 2014, 75, S38. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.; Brunser, O.; Atala, E.; Herranz, J.; de Camargo, A.C.; Zbinden-Foncea, H.; Speisky, H. Protection against Indomethacin-Induced Loss of Intestinal Epithelial Barrier Function by a Quercetin Oxidation Metabolite Present in Onion Peel: In Vitro and in Vivo Studies. J. Nutr. Biochem. 2022, 100, 108886. [Google Scholar] [CrossRef] [PubMed]
- Inhibition of COX-2 and PGE2 in LPS-Stimulated RAW264.7 Cells by Lonimacranthoide VI, a Chlorogenic Acid Ester Saponin. Available online: https://www.spandidos-publications.com/10.3892/br.2014.314 (accessed on 5 February 2025).
- Eliopoulos, A.G.; Dumitru, C.D.; Wang, C.C.; Cho, J.; Tsichlis, P.N. Induction of COX-2 by LPS in Macrophages Is Regulated by Tpl2-Dependent CREB Activation Signals. EMBO J. 2002, 21, 4831–4840. [Google Scholar] [CrossRef]
- Fujimoto, T.; Gotoh, H. Feature Selection for the Interpretation of Antioxidant Mechanisms in Plant Phenolics. Molecules 2023, 28, 1454. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, J.; Tan, J.; Liu, X.; Wang, B. Flavonoids from Agrimonia Pilosa Ledeb: Free Radical Scavenging and DNA Oxidative Damage Protection Activities and Analysis of Bioactivity-Structure Relationship Based on Molecular and Electronic Structures. Molecules 2017, 22, 195. [Google Scholar] [CrossRef]
- Amić, D.; Lučić, B. Reliability of Bond Dissociation Enthalpy Calculated by the PM6 Method and Experimental TEAC Values in Antiradical QSAR of Flavonoids. Bioorg. Med. Chem. 2010, 18, 28–35. [Google Scholar] [CrossRef]
- Atalay, V.E.; Atish, I.S.; Shahin, K.F.; Kashikchi, E.S.; Karahan, M.A. A DFT Study: Ranking of Antioxidant Activity of Various Candidate Molecules. UNEC J. Eng. Appl. Sci. 2022, 2, 33–40. [Google Scholar]
- Shaker, L.M.; Al-Amiery, A.A.; Abed, T.K.; Al-Azzawi, W.K.; Kadhum, A.A.H.; Sulaiman, G.M.; Mohammed, H.A.; Khan, M.; Khan, R.A. An Overview of the Density Functional Theory on Antioxidant Bioactivity Predictive Feasibilities: Insights from Natural Antioxidant Products. J. Mol. Struct. 2024, 1301, 137393. [Google Scholar] [CrossRef]
- Fernanda Arias-Santé, M.; Fuentes, J.; Ojeda, C.; Aranda, M.; Pastene, E.; Speisky, H. Amplification of the Antioxidant Properties of Myricetin, Fisetin, and Morin Following Their Oxidation. Food Chem. 2024, 435, 137487. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ORAC Index | CAA (%) | Viability (%) | |||
|---|---|---|---|---|---|---|
| PGR | FL | HFF (10 µg/mL) | RAW 264.7 | |||
| (10 µg/mL) | (100 µg/mL) | |||||
| Quercetin (A1) | 2.32 ± 0.20 **** | 12.43 ± 0.37 **** | 87.8 ± 0.4 **** | 100 ± 14 * | 98 ± 5 | 89 ± 4 |
| Mox-Quer pH2 (C1) | 0.88 ± 0.01 *** | 8.98 ± 1.14 **** | 91.2 ± 0.4 ** | 100 ± 7 * | 100 ± 3 | Nd 1 |
| Mox-Quer pH7 (D1) | Low reactive | 5.43 ± 0.07 **** | 55.3 ± 1.5 **** | 100 ± 12 | 100 ± 2 | Nd 1 |
| Kaempferol (A2) | 3.19 ± 0.18 **** | 11.05 ± 0.19 **** | 68.9 ± 1.1 **** | 100 ± 6 | 100 ± 5 | 98 ± 3 |
| Mox-Kam pH2 (C2) | 0.49 ± 0.08 ** | 6.80 ± 0.43 **** | 93.3 ± 2.3 * | 100 ± 3 | 95 ± 4 | Nd 1 |
| Mox-Kam pH7 (D2) | 0.46 ± 0.03 ** | 4.77 ± 0.13 **** | 76.1 ± 1.1 **** | 89 ± 6 | 100 ± 8 * | Nd 1 |
| Rhamnetin (A3) | 3.25 ± 0.16 **** | 5.02 ± 0.17 **** | 53.2 ± 2.9 **** | 70 ± 7 **** | 100 ± 3 | 93 ± 4 |
| Mox-Ram pH2 (C3) | 1.04 ± 0.09 **** | 6.63 ± 0.37 **** | 81.0 ± 2.5 **** | 88 ± 3 | 100 ±5 | Nd 1 |
| Mox-Ram pH7 (D3) | Low reactive | 5.23 ± 0.17 **** | 54.5 ± 3.9 **** | 85 ± 3 | 98 ± 2 | Nd 1 |
| Trolox | 1.00 ± 0.02 | 1.00 ± 0.05 | Nd 1 | Nd 1 | Nd 1 | Nd 1 |
| Luteolin | Nd 1 | 5.11 ± 0.45 **** | Nd 1 | Nd 1 | Nd 1 | Nd 1 |
| Rosmarinic acid | Nd 1 | 4.31 ± 0.28 **** | Nd 1 | Nd 1 | Nd 1 | Nd 1 |
| Ferulic acid | Nd 1 | 2.84 ± 0.36 *** | Nd 1 | Nd 1 | Nd 1 | Nd 1 |
| η (r2SCAN-D4/def2-TZVPP/M06-2X) | IP (r2SCAN-D4/def2-TZVPP/M06-2X) | Molecule |
|---|---|---|
| 89.8/122.1 | 123.2/91.0 | Quercetin |
| 90.2/124.9 | 124.0/99.5 | Kaempferol |
| 89.7/119.9 | 122.7/89.5 | Rhamnetin |
| 103.6/149.3 | 141.0/56.2 | Quer-BZF-OH |
| 106.1/179.3 | 146.4/117.5 | Kaem-BZF-OH |
| 102.5/151.6 | 140.8/60.8 | Ram-BZF-OH |
| 100.7/145.0 | 138.8/94.9 | Quer-BZF-OMe |
| 102.9/203.7 | 143.4/215.7 | Kam-BZF-OMe |
 | |||
|---|---|---|---|
| BDEηW (r2SCAN-D4/def2-TZVPP/M06-2X) | (r2SCAN-D4/def2-TZVPP/M06-2X) | ||
| 58.39/79.38 | 0.058/0.064 | 3-OH | Quercetin |
| 61.98/84.26 | 0.026/0.016 | 5-OH | |
| 61.98/84.26 | 0.026/0.018 | 7-OH | |
| 61.98/84.26 | 0.029/0.022 | 3’-OH | |
| 61.08/83.04 | 0.042/0.039 | 4’-OH | |
| 58.61/80.71 | 0.062/0.070 | 3-OH | Kaempferol |
| 62.22/85.68 | 0.029/0.017 | 5-OH | |
| 62.22/85.68 | 0.028/0.018 | 7-OH | |
| 62.22/85.68 | 0.042/0.036 | 4’-OH | |
| 58.28/77.90 | 0.057/0.065 | 3-OH | Rhamnetin |
| 61.87/82.69 | 0.028/0.017 | 5-OH | |
| 61.87/82.69 | 0.028/0.021 | 3’-OH | |
| 60.97/81.49 | 0.041/0.037 | 4’-OH | |
| 67.34/97.07 | 0.022/0.019 | 2-OH | BZF-OH-Quercetin |
| 70.44/101.55 | 0.028/0.004 | 4-OH | |
| 71.48/103.05 | 0.015/0.005 | 6-OH | |
| 70.44/101.55 | 0.059/0.86 | 3’-OH | |
| 69.41/100.06 | 0.055/0.069 | 4’-OH | |
| 68.97/115.89 | 0.025/0.023 | 2-OH | BZF-OH-Kaempferol |
| 72.16/121.25 | 0.042/0.056 | 4-OH | |
| 73.22/123.03 | 0.019/0.025 | 6-OH | |
| 73.22/123.03 | 0.054/0.021 | 4’-OH | |
| 66.64/98.52 | 0.023/0.019 | 2-OH | BZF-OH-Rhamnetin |
| 69.71/103.06 | 0.031/0.004 | 4-OH | |
| 69.71/103.06 | 0.055/0.086 | 3’-OH | |
| 68.69/101.55 | 0.052/0.069 | 4’-OH | |
| 68.50/98.62 | 0.026/0.004 | 4-OH | BZF-OMe-Quercetin |
| 69.51/100.07 | 0.019/0.005 | 6-OH | |
| 68.50/98.62 | 0.060/0.084 | 3’-OH | |
| 68.50/98.62 | 0.053/0.069 | 4’-OH | |
| 69.97/138.52 | 0.035/0.056 | 4-OH | BZF-OMe-Kaempferol |
| 71.00/140.56 | 0.023/0.026 | 6-OH | |
| 71.00/140.56 | 0.046/0.020 | 4’-OH | |
| Docking Score (kcal/mol) | Molecule |
|---|---|
| −10.5 | Celecoxib |
| −8.9 | Quercetin (A1) |
| −8.7 | Kaempferol (A2) |
| −8.4 | Rhamnetin (A3) |
| −9.1 | BZF-Quer-OH |
| −8.9 | BZF-Kaem-OH |
| −8.2 | BZF-Ram-OH |
| MM-GBSA(b) (kcal/mol) | MM-GBSA(a) (kcal/mol) | Molecule |
|---|---|---|
| −46.6 | −48.4 | Celecoxib |
| −27.7 | −28.2 | Quercetin (A1) |
| −29.7 | −29.9 | Kaempferol (A2) |
| −31.8 | −31.9 | Rhamnetin (A3) |
| −30.2 | −31.9 | BZF-Quer-OH |
| −28.3 | −32.2 | BZF-Kaem-OH |
| −30.9 | −44.7 | BZF-Ram-OH |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acosta-Quiroga, K.; Rocha-Valderrama, E.; Zúñiga-Bustos, M.; Mera-Adasme, R.; Cabrera-Barjas, G.; Olea-Azar, C.; Moncada-Basualto, M. Gross Antioxidant Capacity and Anti-Inflammatory Potential of Flavonol Oxidation Products: A Combined Experimental and Theoretical Study. Antioxidants 2025, 14, 479. https://doi.org/10.3390/antiox14040479
Acosta-Quiroga K, Rocha-Valderrama E, Zúñiga-Bustos M, Mera-Adasme R, Cabrera-Barjas G, Olea-Azar C, Moncada-Basualto M. Gross Antioxidant Capacity and Anti-Inflammatory Potential of Flavonol Oxidation Products: A Combined Experimental and Theoretical Study. Antioxidants. 2025; 14(4):479. https://doi.org/10.3390/antiox14040479
Chicago/Turabian StyleAcosta-Quiroga, Karen, Esteban Rocha-Valderrama, Matías Zúñiga-Bustos, Raúl Mera-Adasme, Gustavo Cabrera-Barjas, Claudio Olea-Azar, and Mauricio Moncada-Basualto. 2025. "Gross Antioxidant Capacity and Anti-Inflammatory Potential of Flavonol Oxidation Products: A Combined Experimental and Theoretical Study" Antioxidants 14, no. 4: 479. https://doi.org/10.3390/antiox14040479
APA StyleAcosta-Quiroga, K., Rocha-Valderrama, E., Zúñiga-Bustos, M., Mera-Adasme, R., Cabrera-Barjas, G., Olea-Azar, C., & Moncada-Basualto, M. (2025). Gross Antioxidant Capacity and Anti-Inflammatory Potential of Flavonol Oxidation Products: A Combined Experimental and Theoretical Study. Antioxidants, 14(4), 479. https://doi.org/10.3390/antiox14040479