
Green-Synthesized Silver Nanoparticles Using Filipendula ulmaria (L.) Maxim. and Salvia verticillata L. Extracts Inhibit Migration and Modulate Redox Homeostasis in Human Breast Cancer Cells via Nrf-2 Signaling Pathway

 , ,
, ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Culturing Cells and Treatment
2.2. Determination of Superoxide Anion Radical (NBT Test)
2.3. Determination of NO Concentration (Griess Method)
2.4. Determination of Reduced and Total Glutathione Concentration
2.5. Determination of Lipid Peroxidation (TBARS Test)
2.6. Determination of Migratory Potential (Transwell Method)
2.7. Measurement of MMP-9 Concentration
2.8. Measurement of Nrf-2 Concentration
2.9. Statistical Analyses
3. Results
3.1. Effects on Redox Status in Tumor Cells
3.1.1. Determination of Superoxide Anion Radical (NBT Assay)
3.1.2. Determination of Nitrites (Griess Assay)
3.1.3. Determination of Total and Reduced Glutathione
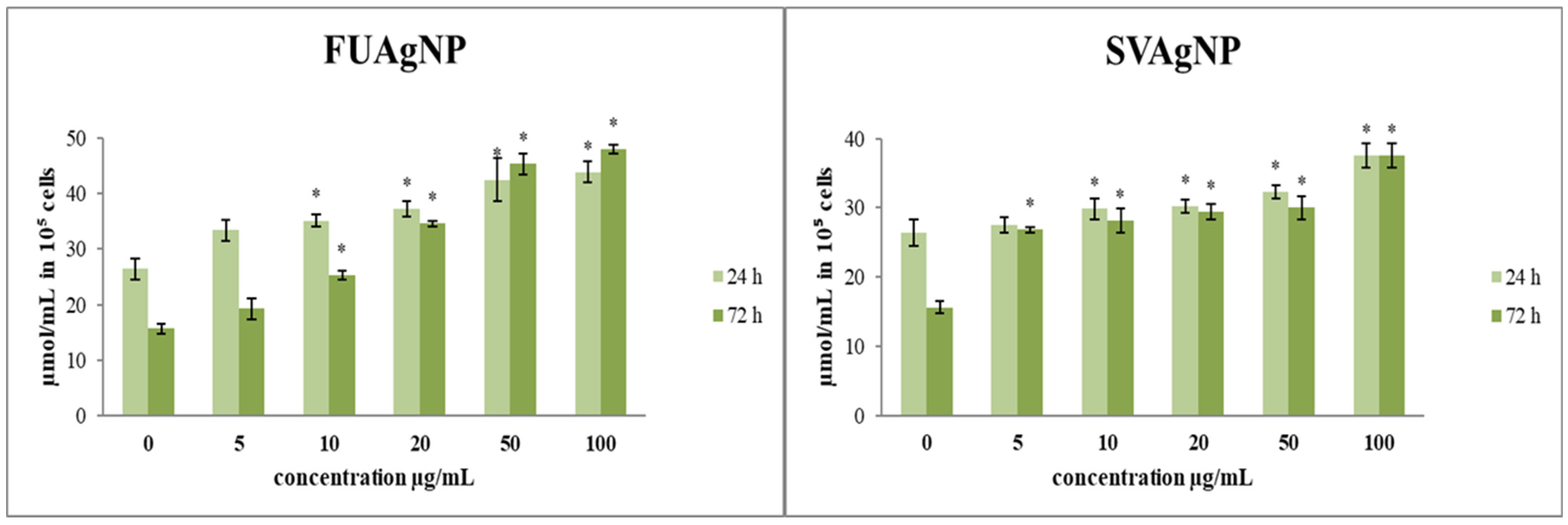
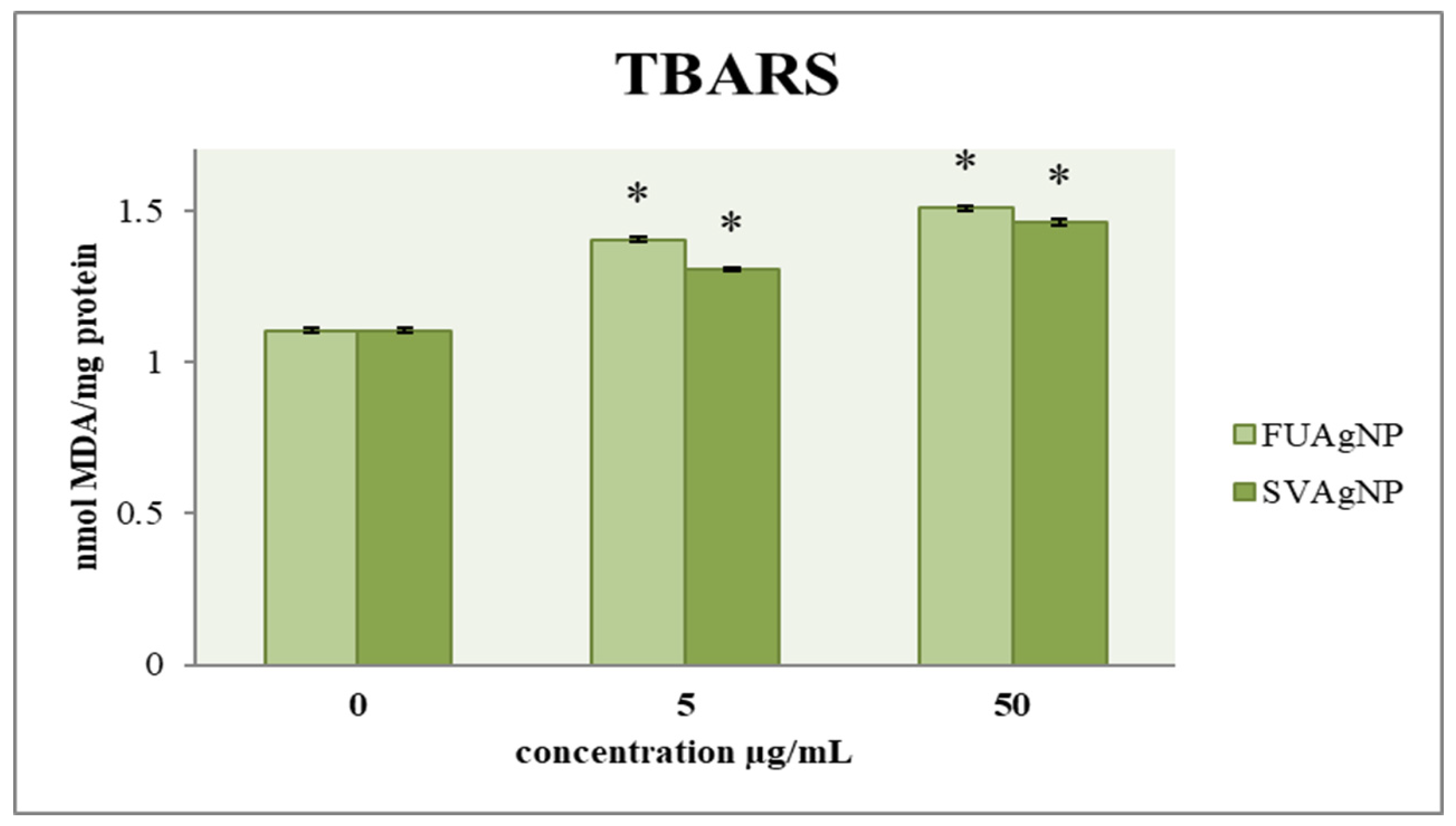
3.1.4. Determination of Lipid Peroxidation
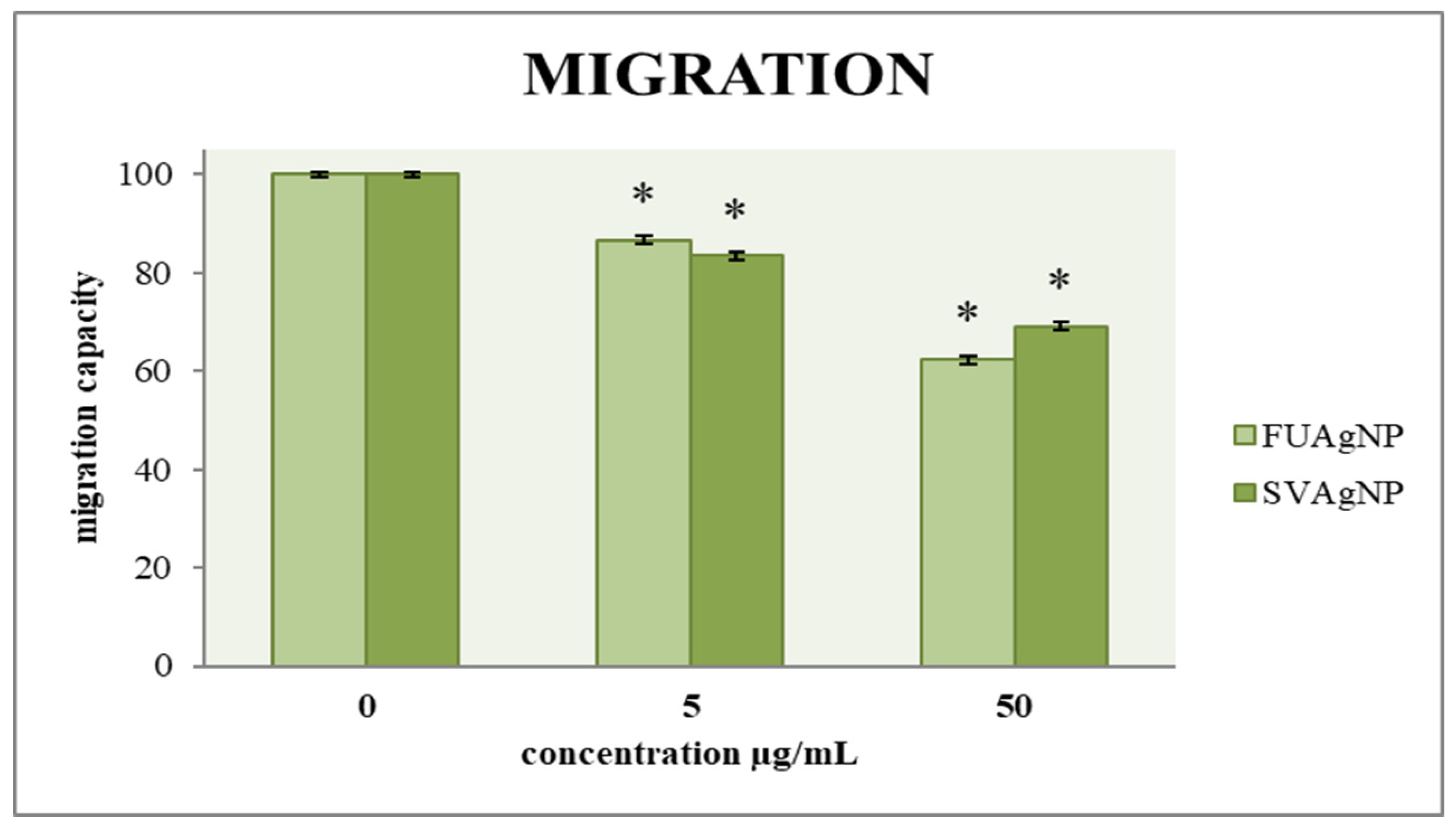
3.2. Transwell Assay for Cell Migration
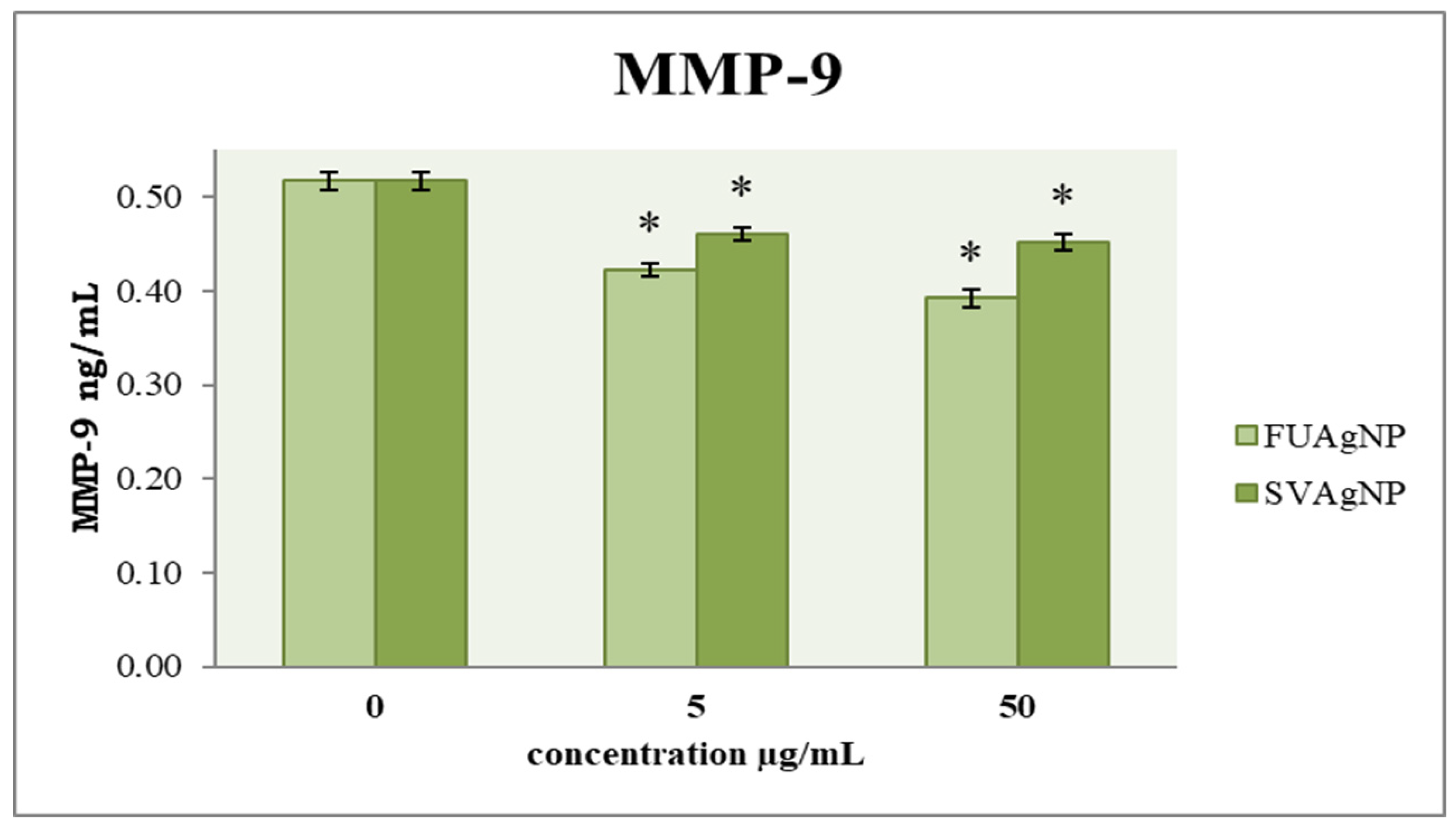
3.3. Concentration of MMP-9
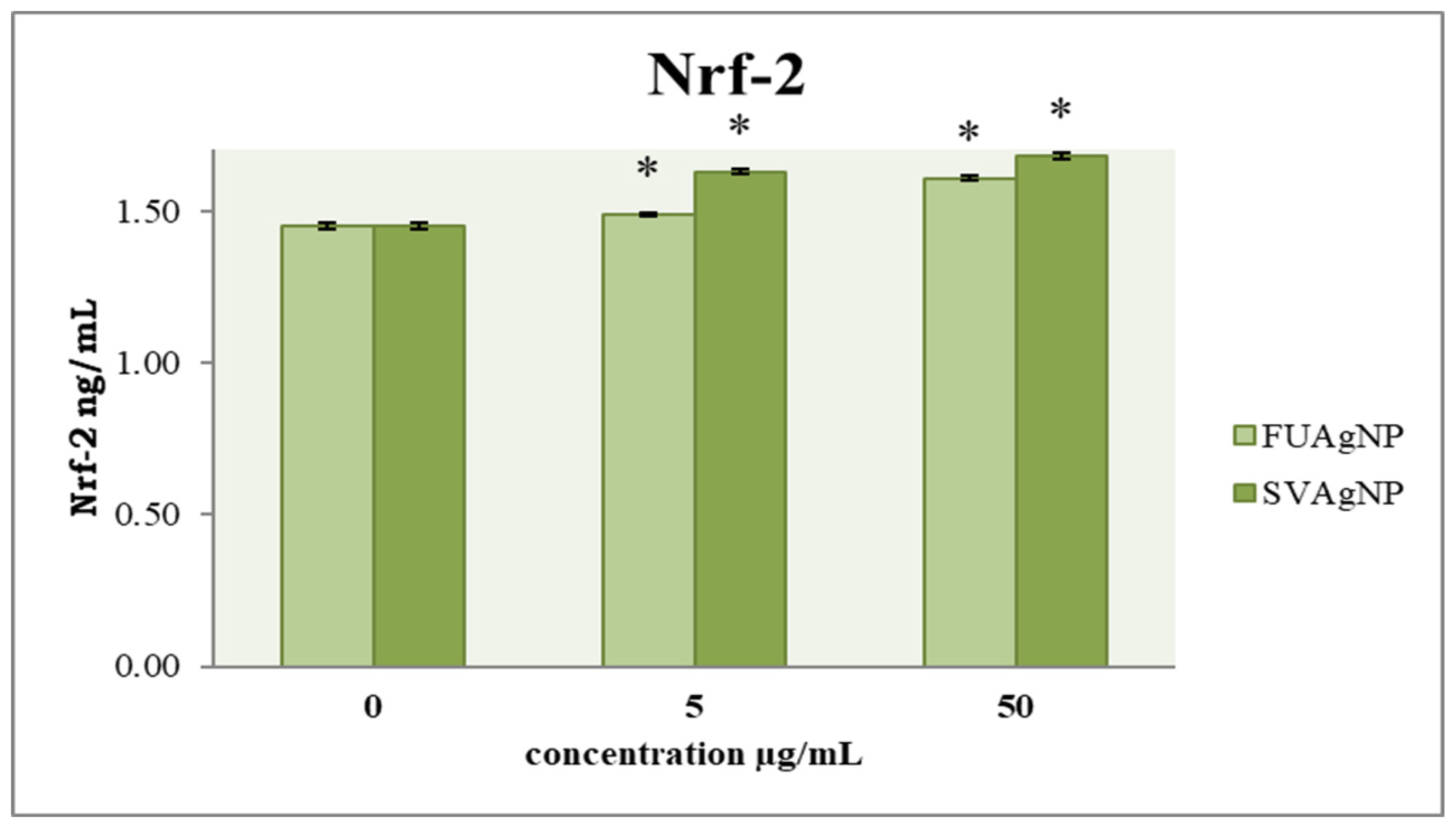
3.4. Concentration of Nrf-2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilkinson, L.; Gathani, T. Understanding breast cancer as a global health concern. Br. J. Radiol. 2022, 95, 20211033. [Google Scholar] [CrossRef]
- Lumachi, F.; Santeufemia, D.A.; Basso, S.M.M. Current medical treatment of estrogen receptorpositive breast cancer. World J. Biol. Chem. 2015, 6, 231–239. [Google Scholar]
- Wee, P.; Wang, Z. Epidermal Growth Factor Receptor Cell Proliferation Signaling Pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar]
- Di Carlo, E.; Sorrentino, C. Oxidative Stress and Age-Related Tumors. Antioxidants 2024, 13, 1109. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 4th ed.; Oxford Univerity Press Inc.: New York, NY, USA, 2007. [Google Scholar]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Arancibia-Hernández, Y.L.; Hernández-Cruz, E.Y.; Pedraza-Chaverri, J. RONS and Oxidative Stress: An Overview of Basic Concepts. Oxygen 2022, 2, 437–478. [Google Scholar] [CrossRef]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar]
- Kumar, H.; Kumar, R.M.; Bhattacharjee, D.; Somanna, P.; Jain, V. Role of Nrf2 Signaling Cascade in Breast Cancer: Strategies and Treatment. Front. Pharmacol. 2022, 13, 720076. [Google Scholar] [CrossRef]
- Mustafa, S.; Koran, S.; AlOmair, L. Insights into the Role of Matrix Metalloproteinases in Cancer and its Various Therapeutic Aspects: A Review. Front. Mol. Biosci. 2022, 9, 896099. [Google Scholar] [CrossRef]
- Mihailović, V.; Srećković, N.; Nedić, Z.P.; Dimitrijević, S.; Matić, M.; Obradović, A.; Selaković, D.; Rosić, G.; Katanić Stanković, J.S. Green Synthesis of Silver Nanoparticles Using Salvia verticillata and Filipendula ulmaria Extracts: Optimization of Synthesis, Biological Activities, and Catalytic Properties. Molecules 2023, 28, 808. [Google Scholar] [CrossRef]
- Auclair, C.; Voisin, E. Nitroblue Tetrazolium Reduction. In CRC Handbook of Methods for Oxygen Radical Research; Greenwald, R.E., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 123–132. [Google Scholar]
- Griess, P. Bemerkungen Zu Der Abhandlung Der HH. Weselsky Und Benedikt „UeberEinigeAzoverbindungen”. Berichte Der Dtsch. Ges. 1879, 12, 426–428. [Google Scholar] [CrossRef]
- Beutler, E. Reduced Glutathione (GSH). In Red Blood Cell Metabolism: A Manual of Biochemical Methods; Bergmeyen, H.V., Ed.; Grune and Stratton: New York, NY, USA, 1975; pp. 112–114. [Google Scholar]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Ohkawa, H.; Okishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Chen, H.C. Boyden chamber assay. Methods Mol. Biol. 2005, 294, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Obradović, A.; Paunović, M.; Ognjanović, B.; Vukić, M.; Vuković, N.; Kačániová, M.; Matić, M. Tanacetum balsamita essential oil from flower exerts antitumor effects by downregulating Nrf-2 and MMP-9 expression in human breast cancer cell. In Proceedings of the “Conference on Advances in Science And Technology” Coast, Herceg Novi, Montenegro, 29 May–1 June 2024; ISBN 978-9940-611-08-8. [Google Scholar]
- Malla, R.; Surepalli, N.; Farran, B.; Malhotra, S.V.; Nagaraju, G.P. Reactive oxygen species (ROS): Critical roles in breast tumor microenvironment. Crit. Rev. Oncol. Hematol. 2021, 160, 103285. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Sarkar, F. Signaling Mechanism(S) of Reactive Oxygen Species in Epithelial-Mesenchymal Transition Rem-iniscent of Cancer Stem Cells in Tumor Progression. Curr. Stem. Cell Res. Ther. 2010, 5, 74–80. [Google Scholar] [CrossRef]
- Çetin, R.; Devrim, E.; Kılıçoğlu, B.; Avcı, A.; Çandır, Ö.; Durak, İ. Cisplatin Impairs Antioxidant System and Causes Oxidation in Rat Kidney Tissues: Possible Protective Roles of Natural Antioxidant Foods. J. Appl. Toxicol. 2006, 26, 42–46. [Google Scholar] [CrossRef]
- Manda, G.; Isvoranu, G.; Comanescu, M.V.; Manea, A.; DebelecButuner, B.; Korkmaz, K.S. The redox biology network in cancer pathophysiology and therapeutics. Redox. Biol. 2015, 5, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.H.; Dervan, E.; Bhattacharyya, D.D.; McAuliffe, J.D.; Miranda, K.M.; Glynn, S.A. The Role of Nitric Oxide in Cancer: Master Regulator or NOt? Int. J. Mol. Sci. 2020, 21, 9393. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Kangari, P.; Zarnoosheh Farahany, T.; Golchin, A.; Ebadollahzadeh, S.; Salmaninejad, A.; Mahboob, S.A.; Nourazarian, A. Enzymatic Antioxidant and Lipid Peroxidation Evaluation in the Newly Diagnosed Breast Cancer Patients in Iran. Asian Pac. J. Cancer Prev. 2018, 19, 3511–3515. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Aykin-Burns, N.; Ahmad, I.M.; Zhu, Y.; Oberley, L.W.; Spitz, D.R. Increased levels of superoxide and H2O2 mediate the dif-ferential susceptibility of cancer cells versus normal cells to glucose deprivation. Biochem. J. 2009, 418, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Gautam, N.; Mantha, A.K.; Mittal, S. Essential Oils and Their Constituents as Anticancer Agents: A Mechanistic View. Bio. Med. Res. Int. 2014, 2014, 54106. [Google Scholar] [CrossRef]
- Katanić, J.; Boroja, T.; Stanković, N.; Mihailović, V.; Mladenović, M.; Kreft, S.; Vrvić, M.M. Bioactivity, stability and phenolic characterization of Filipendula ulmaria (L.) Maxim. Food Funct. 2015, 6, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Stanković, J.S.K.; Srećković, N.; Mišić, D.; Gašić, U.; Imbimbo, P.; Monti, D.M.; Mihailović, V. Bioactivity, biocompatibility and phytochemical assessment of lilac sage, Salvia verticillata L. (Lamiaceae)-A plant rich in rosmarinic acid. Ind. Crops Prod. 2020, 143, 111932. [Google Scholar] [CrossRef]
- Srećković, N.Z.; Nedić, Z.P.; Monti, D.M.; D’Elia, L.; Dimitrijević, S.B.; Mihailović, N.R.; Katanić Stanković, J.S.; Mihailović, V.B. Biosynthesis of Silver Nanoparticles Using Salvia pratensis L. Aerial Part and Root Extracts: Bioactivity, Biocompatibility, and Catalytic Potential. Molecules 2023, 28, 1387. [Google Scholar] [CrossRef]
- Takáč, P.; Michalková, R.; Čižmáriková, M.; Bedlovičová, Z.; Balážová, Ľ.; Takáčová, G. The Role of Silver Nanoparticles in the Diagnosis and Treatment of Cancer: Are There Any Perspectives for the Future? Life 2023, 13, 466. [Google Scholar] [CrossRef]
- Liu, H.; Liu, X.; Zhang, C.; Zhu, H.; Xu, Q.; Bu, Y.; Lei, Y. Redox Imbalance in the Development of Colorectal Cancer. J. Cancer 2017, 8, 1586–1597. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Nazıroglu, M. TRPV1 Channel: A Potential Drug Target for Treating Epilepsy. Curr. Neuropharmacol. 2015, 13, 239–247. [Google Scholar] [CrossRef]
- Nakamura, H.; Takada, K. Reactive oxygen species in cancer: Current findings and future directions. Cancer Sci. 2021, 112, 3945–3952. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Cai, S.; Wang, M.; Chen, Z.; Lu, C.; Yang, H. Photogenerated Holes Mediated Nitric Oxide Production for Hypoxic Tumor Treatment. Angew. Chem. 2021, 60, 7046–7050. [Google Scholar] [CrossRef]
- Rigas, B.; Williams, J.L. NO-donating NSAIDs and cancer: An overview with a note on whether NO is required for their action. Nitric Oxide 2008, 19, 199–204. [Google Scholar] [CrossRef]
- Xiao, L.; Xian, M.; Zhang, C.; Guo, Q.; Yi, Q. Lipid peroxidation of immune cells in cancer. Front. Immunol. 2024, 14, 1322746. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, L.; Sandhu, J.K.; Harper, M.E.; Cuperlovic-Culf, M. Role of Glutathione in Cancer: From Mechanisms to Therapies. Biomolecules. 2020, 10, 1429. [Google Scholar] [CrossRef]
- Vannini, F.; Kashfi, K.; Nath, N. The dual role of iNOS in cancer. Redox Biol. 2015, 6, 334–343. [Google Scholar] [CrossRef]
- Gupta, G.P.; Massagué, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Kang, Y. Epithelial-Mesenchymal Plasticity in Cancer Progression and Metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef]
- Huang, H. Matrix Metalloproteinase-9 (MMP-9) as a Cancer Biomarker and MMP-9 Biosensors: Recent Advances. Sensors 2018, 18, 3249. [Google Scholar] [CrossRef]
- Sathishkumar, P.; Vennila, K.; Jayakumar, R.; Yusoff, A.R.M.; Hadibarata, T.; Palvannan, T. Phyto-Synthesis of Silver Nanoparticles Using Alternanthera tenella Leaf Extract: An Effective Inhibitor for the Migration of Human Breast Adenocarcinoma (MCF-7) Cells. Bioprocess. Biosyst. Eng. 2016, 39, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Kavaz, D.; Umar, H.; Shehu, S. Synthesis, Characterization, Antimicrobial and Antimetastatic Activity of Silver Nanoparticles Synthesized from Ficus ingens Leaf. Artif. Cells Nanomed. Biotechnol. 2018, 46, S1193–S1203. [Google Scholar] [CrossRef] [PubMed]
- Ghareghomi, S.; Habibi-Rezaei, M.; Arese, M.; Saso, L.; Moosavi-Movahedi, A.A. Nrf2 Modulation in Breast Cancer. Biomedicines 2022, 10, 2668. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matić, M.; Obradović, A.; Paunović, M.; Ognjanović, B.; Mihailović, V.; Srećković, N.; Stanković, M. Green-Synthesized Silver Nanoparticles Using Filipendula ulmaria (L.) Maxim. and Salvia verticillata L. Extracts Inhibit Migration and Modulate Redox Homeostasis in Human Breast Cancer Cells via Nrf-2 Signaling Pathway. Antioxidants 2025, 14, 469. https://doi.org/10.3390/antiox14040469
Matić M, Obradović A, Paunović M, Ognjanović B, Mihailović V, Srećković N, Stanković M. Green-Synthesized Silver Nanoparticles Using Filipendula ulmaria (L.) Maxim. and Salvia verticillata L. Extracts Inhibit Migration and Modulate Redox Homeostasis in Human Breast Cancer Cells via Nrf-2 Signaling Pathway. Antioxidants. 2025; 14(4):469. https://doi.org/10.3390/antiox14040469
Chicago/Turabian StyleMatić, Miloš, Ana Obradović, Milica Paunović, Branka Ognjanović, Vladimir Mihailović, Nikola Srećković, and Milan Stanković. 2025. "Green-Synthesized Silver Nanoparticles Using Filipendula ulmaria (L.) Maxim. and Salvia verticillata L. Extracts Inhibit Migration and Modulate Redox Homeostasis in Human Breast Cancer Cells via Nrf-2 Signaling Pathway" Antioxidants 14, no. 4: 469. https://doi.org/10.3390/antiox14040469
APA StyleMatić, M., Obradović, A., Paunović, M., Ognjanović, B., Mihailović, V., Srećković, N., & Stanković, M. (2025). Green-Synthesized Silver Nanoparticles Using Filipendula ulmaria (L.) Maxim. and Salvia verticillata L. Extracts Inhibit Migration and Modulate Redox Homeostasis in Human Breast Cancer Cells via Nrf-2 Signaling Pathway. Antioxidants, 14(4), 469. https://doi.org/10.3390/antiox14040469