
Natural Compounds with Beneficial Effects on Skin Collagen Type I and Mechanisms of Their Action


Abstract
1. Introduction
2. Collagen Biosynthesis and Regulation
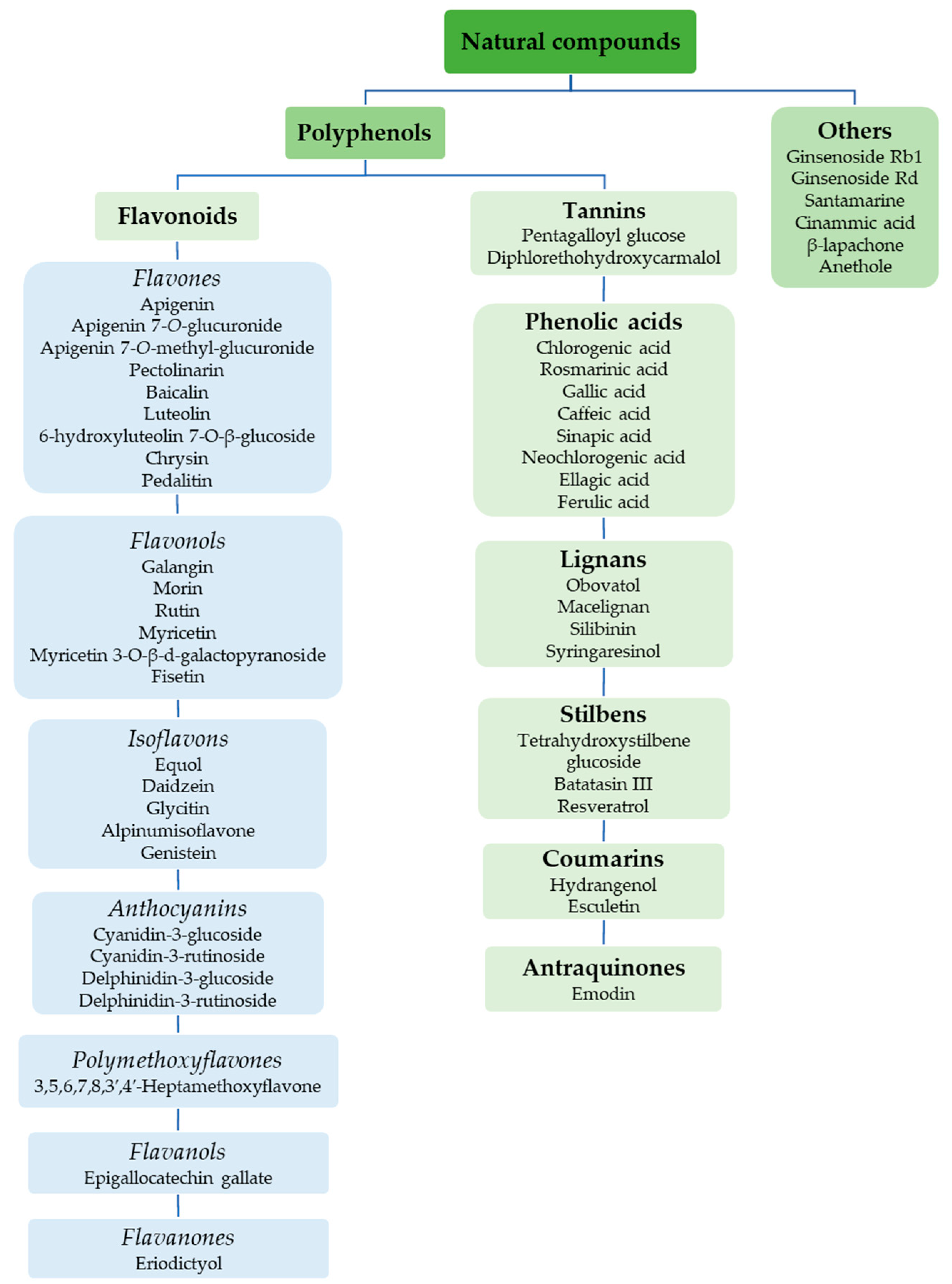
3. Natural Compounds with Stimulating and Protective Effects on Skin Collagen Type I
3.1. Mechanisms of Action of Natural Compounds on Skin Collagen Type I
3.1.1. TGF-β/Smad Pathway
3.1.2. MAPK Signaling Pathways
The MAPK/ERK1/2 Pathway
The MAPK/p38/JNK/AP-1 Pathway
The MAPK/NF-κB Pathway
3.1.3. The PI3K/Akt/mTOR Pathway
3.1.4. IGF-1/IGF-1R
3.1.5. The Nrf2/ARE Pathway
3.1.6. Other Mechanisms
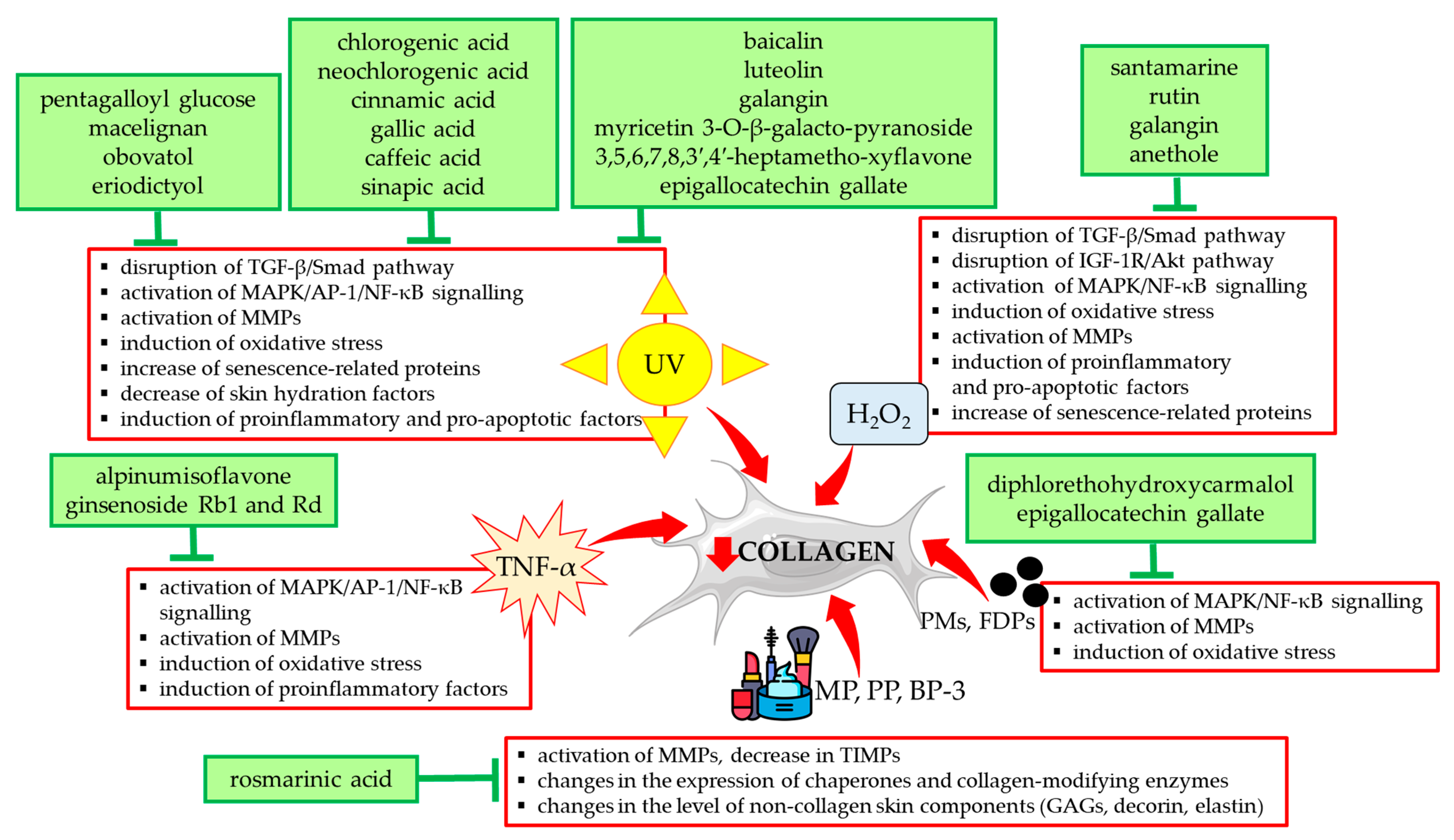
3.2. The Protective Effect of Natural Compounds Against the Unfavorable Influence of External Factors on Collagen Type I
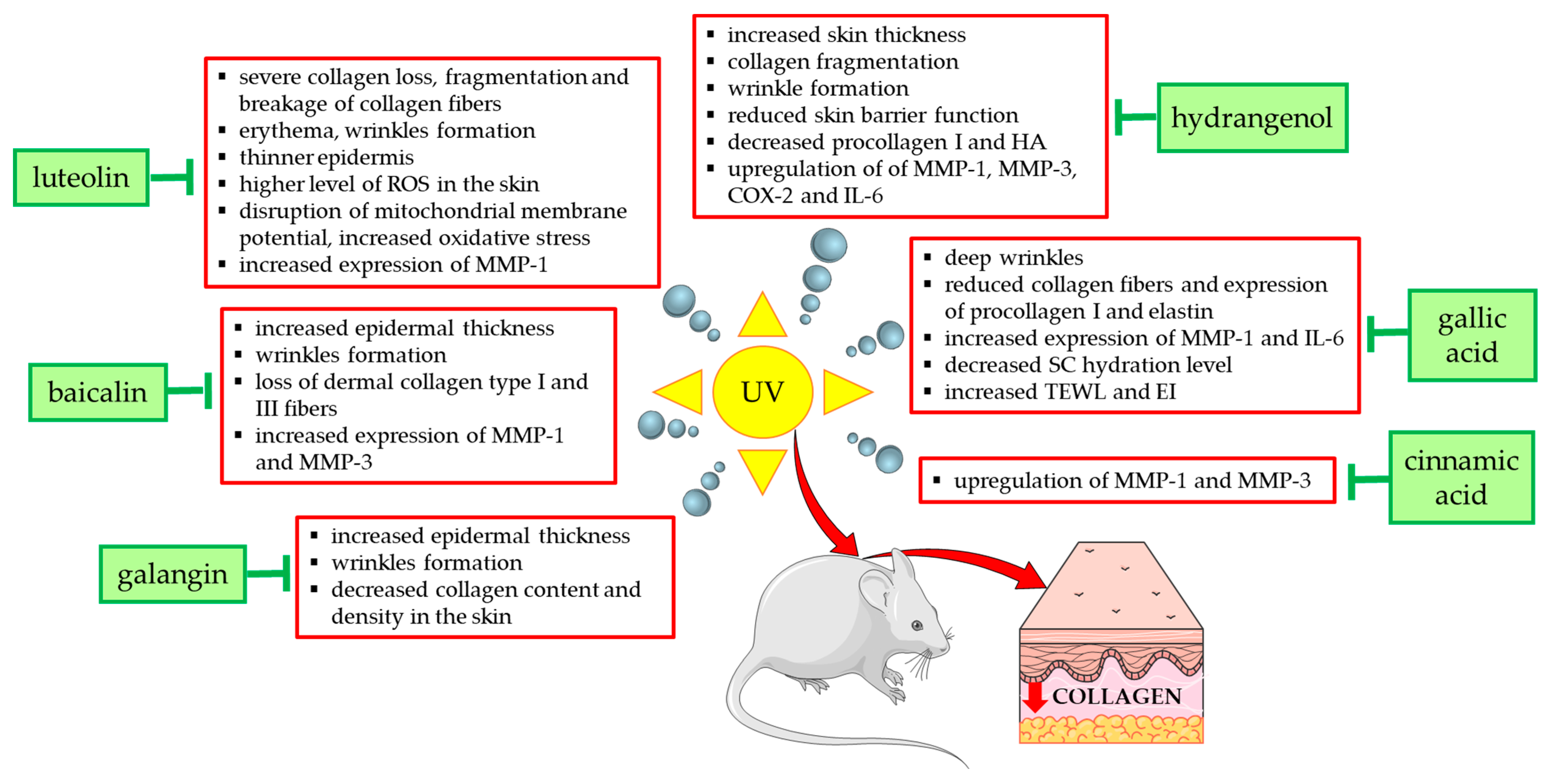
3.2.1. UV
3.2.2. Air Pollution
3.2.3. Chemicals from Cosmetics Products
3.3. Examples of Clinical Studies on the Use of Collagen Type I Stimulating Natural Compounds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Kadler, K.; Baldock, C.; Bella, J.; Boot-Handford, R. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar]
- Naomi, R.; Ridzuan, P.M.; Bahari, H. Current Insights into Collagen Type I. Polymers 2021, 13, 2642. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Baffet, G.; Théret, N. Molecular and tissue alterations of collagens in fibrosis. Matrix Biol. 2018, 68–69, 122–149. [Google Scholar] [CrossRef]
- Davison-Kotler, E.; Marshall, W.S.; García-Gareta, E. Sources of collagen for biomaterials in skin wound healing. Bioengineering 2019, 6, 56. [Google Scholar] [CrossRef]
- Amirrah, I.N.; Lokanathan, Y.; Zulkiflee, I.; Wee, M.F.M.R.; Motta, A.; Fauzi, M.B. A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold. Biomedicines 2022, 10, 2307. [Google Scholar] [CrossRef]
- Sklenářová, R.; Akla, N.; Latorre, M.J.; Ulrichová, J.; Franková, J. Collagen as a Biomaterial for Skin and Corneal Wound Healing. J. Funct. Biomater. 2022, 13, 249. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar]
- Holmes, D.F.; Lu, Y.; Starborg, T.; Kadler, K.E. Collagen Fibril Assembly and Function. Curr. Top. Dev. Biol. 2018, 130, 107–142. [Google Scholar]
- Reilly, D.M.; Lozano, J. Skin collagen through the lifestages: Importance for skin health and beauty. Plast. Aesthet. Res. 2021, 8, 2. [Google Scholar]
- Chowdhury, S.R.; Mh Busra, M.F.; Lokanathan, Y.; Ng, M.H.; Law, J.X.; Cletus, U.C.; Haji Idrus, R.B. Collagen Type I: A Versatile Biomaterial. Adv. Exp. Med. Biol. 2018, 1077, 389–414. [Google Scholar]
- Wolf, K.; Alexander, S.; Schacht, V.; Coussens, L.M.; von Andrian, U.H.; van Rheenen, J.; Deryugina, E.; Friedl, P. Collagen-based cell migration models In Vitro and In Vivo. Semin. Cell Dev. Biol. 2009, 20, 931–941. [Google Scholar]
- Elango, J.; Hou, C.; Bao, B.; Wang, S.; Maté Sánchez de Val, J.E.; Wenhui, W. The Molecular Interaction of Collagen with Cell Receptors for Biological Function. Polymers 2022, 14, 876. [Google Scholar] [CrossRef]
- Wiegand, C.; Schönfelder, U.; Abel, M.; Ruth, P.; Kaatz, M.; Hipler, U.C. Protease and pro-inflammatory cytokine concentrations are elevated in chronic compared to acute wounds and can be modulated by collagen type I In Vitro. Arch. Dermatol. Res. 2009, 302, 419–428. [Google Scholar] [PubMed]
- Twardowski, T.; Fertala, A.; Orgel, J.; Antonio, J.S. Type I Collagen and Collagen Mimetics as Angiogenesis Promoting Superpolymers. Curr. Pharm. Des. 2007, 13, 3608–3621. [Google Scholar]
- Fisher, G.J.; Wang, Z.Q.; Datta, S.C.; Varani, J.; Kang, S.; Voorhees, J.J. Pathophysiology of premature skin aging induced by ultraviolet light. N. Engl. J. Med. 1997, 337, 1419–1428. [Google Scholar]
- Lan, C.E.; Hung, Y.T.; Fang, A.H.; Ching-Shuang, W. Effects of irradiance on UVA-induced skin aging. J. Dermatol. Sci. 2019, 94, 220–228. [Google Scholar] [PubMed]
- Laronha, H.; Caldeira, J. Structure and function of human matrix metalloproteinases. Cells 2020, 9, 1076. [Google Scholar] [CrossRef]
- Potekaev, N.N.; Borzykh, O.B.; Medvedev, G.V.; Petrova, M.M.; Gavrilyuk, O.A.; Karpova, E.I.; Trefilova, V.V.; Demina, O.M.; Popova, T.E.; Shnayder, N.A. Genetic and Epigenetic Aspects of Skin Collagen Fiber Turnover and Functioning. Cosmetics 2021, 8, 92. [Google Scholar] [CrossRef]
- Vierkötter, A.; Krutmann, J. Environmental influences on skin aging and ethnic-specific manifestations. Dermatoendocrinology 2012, 4, 227–231. [Google Scholar]
- Parrado, C.; Mercado-Saenz, S.; Perez-Davo, A.; Gilaberte, Y.; Gonzalez, S.; Juarranz, A. Environmental Stressors on Skin Aging. Mechanistic Insights. Front. Pharmacol. 2019, 10, 759. [Google Scholar]
- Majewska, N.; Zaręba, I.; Surażyński, A.; Galicka, A. Methylparaben-induced decrease in collagen production and viability of cultured human dermal fibroblasts. J. Appl. Toxicol. 2017, 37, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Matwiejczuk, N.; Galicka, A.; Brzóska, M.M. Review of the safety of application of cosmetic products containing parabens. J. Appl. Toxicol. 2020, 40, 176–210. [Google Scholar] [CrossRef] [PubMed]
- Santander Ballestín, S.; Luesma Bartolomé, M.J. Toxicity of Different Chemical Components in Sun Cream Filters and Their Impact on Human Health: A Review. Appl. Sci. 2023, 13, 712. [Google Scholar] [CrossRef]
- Shin, J.W.; Kwon, S.H.; Choi, J.Y.; Na, J.I.; Huh, C.H.; Choi, H.R.; Park, K.C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef]
- Al-Atif, H. Collagen Supplements for Aging and Wrinkles: A Paradigm Shift in the Fields of Dermatology and Cosmetics. Dermatol. Pract. Concept. 2022, 12, e2022018. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, J.; Shin, D.W. The Molecular Mechanism of Polyphenols with Anti-Aging Activity in Aged Human Dermal Fibroblasts. Molecules 2022, 27, 4351. [Google Scholar] [CrossRef]
- Arct, J.; Pytkowska, K. Flavonoids as components of biologically active cosmeceuticals. Clin. Dermatol. 2008, 26, 347–357. [Google Scholar] [CrossRef]
- Dunaway, S.; Odin, R.; Zhou, L.; Ji, L.; Zhang, Y.; Kadekaro, A.L. Natural Antioxidants: Multiple Mechanisms to Protect Skin From Solar Radiation. Front. Pharmacol. 2018, 9, 392. [Google Scholar] [CrossRef]
- Liu, J.K. Natural products in cosmetics. Nat. Prod. Bioprospect. 2022, 12, 40. [Google Scholar] [CrossRef]
- Kun, N.; Tsuchiya, K.; Nakajima, S. New anti-aging cosmetic ingredients. J. Soc. Cosmet. Chem. Jpn. 1999, 33, 245–253. [Google Scholar] [CrossRef]
- Bou-Gharios, G.; Abraham, D.; de Crombrugghe, B. Type I collagen structure, synthesis, and regulation. In Principles of Bone Biology; Elsevier: Alpharetta, GA, USA, 2020; pp. 295–337. [Google Scholar]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens-Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef]
- Gjaltema, R.A.; Bank, R.A. Molecular insights into prolyl and lysyl hydroxylation of fibrillar collagens in health and disease. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 74–95. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.L.; Kehlet, S.N.; Siebuhr, A.S.; Gudmann, N.S.; Karsdal, M.A. Chapter 35—Collagen chaperones. In Biochemistry of Collagens, Laminins and Elastin, 3rd ed.; Karsdal, M.A., Ed.; Academic Press: Cambridge, MA, USA, 2024; pp. 351–369. [Google Scholar]
- Smith, T.; Ferreira, L.R.; Hebert, C.; Norris, K.; Sauk, J.J. Hsp47 and cyclophilin B traverse the endoplasmic reticulum with procollagen into pre-Golgi intermediate vesicles. A role for Hsp47 and cyclophilin B in the export of procollagen from the endoplasmic reticulum. J. Biol. Chem. 1995, 270, 18323–18328. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Rubin, K.; Bächinger, H.P.; Kalamajski, S. The endoplasmic reticulum-resident collagen chaperone Hsp47 interacts with and promotes the secretion of decorin, fibromodulin, and lumican. J. Biol. Chem. 2018, 293, 13707–13716. [Google Scholar] [CrossRef]
- Kirkness, M.W.; Lehmann, K.; Forde, N.R. Mechanics and structural stability of the collagen triple helix. Curr. Opin. Chem. Biol. 2019, 53, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Devos, H.; Zoidakis, J.; Roubelakis, M.G.; Latosinska, A.; Vlahou, A. Reviewing the regulators of COL1A1. Int. J. Mol. Sci. 2023, 24, 10004. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, X.F.; Wang, Z.C.; Lou, D.; Fang, Q.Q.; Hu, Y.Y.; Zhao, W.Y.; Zhang, L.Y.; Wu, L.H.; Tan, W.Q. Current potential therapeutic strategies targeting the TGF-β/Smad signaling pathway to attenuate keloid and hypertrophic scar formation. Biomed. Pharmacother. 2020, 129, 110287. [Google Scholar] [CrossRef]
- Frangogiannis, N. Transforming growth factor-β in tissue fibrosis. J. Exp. Med. 2020, 217, e20190103. [Google Scholar] [CrossRef]
- Choi, M.E. Mechanism of transforming growth factor-beta1 signaling. Kidney Int. Suppl. 2000, 77, S53–S58. [Google Scholar] [CrossRef]
- Hayashida, T.; Decaestecker, M.; Schnaper, H.W. Cross-talk between ERK MAP kinase and Smad signaling pathways enhances TGF-beta-dependent responses in human mesangial cells. FASEB J. 2003, 17, 1576–1578. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Zhang, Y.; Liu, J.; Zhang, W.; Sha, Y.; Zhan, Y.; Xiang, M. Mechanism of Akt regulation of the expression of collagens and MMPs in conjunctivochalasis. Exp. Eye Res. 2023, 226, 109313. [Google Scholar] [PubMed]
- Gillery, P.; Leperre, A.; Maquart, F.X.; Borel, J.P. Insulin-like growth factor-I (IGF-I) stimulates protein synthesis and collagen gene expression in monolayer and lattice cultures of fibroblasts. J. Cell Physiol. 1992, 152, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Hakuno, F.; Takahashi, S.I. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2016, 20, 1689–1699. [Google Scholar]
- El Oirdi, M. Harnessing the Power of Polyphenols: A New Frontier in Disease Prevention and Therapy. Pharmaceuticals 2024, 17, 692. [Google Scholar] [CrossRef]
- Archivio, D.; Filesi, M.; Di Benedetto, C.; Gargiulo, R.; Giovannini, R.; Masella, C.R. Polyphenols, Dietary Sources and Bioavailability. Ann. Ist. Super. Sanità 2007, 43, 348. [Google Scholar] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar]
- Zhang, Y.; Wang, J.; Cheng, X.; Yi, B.; Zhang, X.; Li, Q. Apigenin induces dermal collagen synthesis via smad2/3 signaling pathway. Eur. J. Histochem. 2015, 59, 98–106. [Google Scholar]
- Galicka, A.; Nazaruk, J. Stimulation of collagen biosynthesis by flavonoid glycosides in skin fibroblasts of osteogenesis imperfecta type I and the potential mechanism of their action. Int. J. Mol. Med. 2007, 20, 889–895. [Google Scholar]
- Zhang, J.A.; Yin, Z.; Ma, L.W.; Yin, Z.Q.; Hu, Y.Y.; Xu, Y.; Wu, D.; Permatasari, F.; Luo, D.; Zhou, B.R. The protective effect of baicalin against UVB irradiation induced photoaging: An In Vitro and In Vivo study. PLoS ONE 2014, 9, e99703. [Google Scholar]
- Mu, J.; Ma, H.; Chen, H.; Zhang, X.; Ye, M. Luteolin Prevents UVB-Induced Skin Photoaging Damage by Modulating SIRT3/ROS/MAPK Signaling: An In Vitro and In Vivo Studies. Front. Pharmacol. 2021, 12, 728261. [Google Scholar]
- Nazaruk, J.; Galicka, A. The influence of selected flavonoids from the leaves of Cirsium palustre (L.) Scop. on collagen expression in human skin fibroblasts. Phytother. Res. 2014, 28, 1399–1405. [Google Scholar]
- Stipcevic, T.; Piljac, J.; Vanden Berghe, D. Effect of different flavonoids on collagen synthesis in human fibroblasts. Plant Foods Hum. Nutr. 2006, 61, 29–34. [Google Scholar]
- Choi, S.J.; Lee, S.N.; Kim, K.; Joo, D.H.; Shin, S.; Lee, J.; Lee, H.K.; Kim, J.; Kwon, S.B.; Kim, M.J.; et al. Biological effects of rutin on skin aging. Int. J. Mol. Med. 2016, 38, 357–363. [Google Scholar] [PubMed]
- Lee, J.J.; Ng, S.C.; Ni, Y.T.; Liu, J.S.; Chen, C.J.; Padma, V.V.; Huang, C.Y.; Kuo, W.W. Protective effects of galangin against H2O2/UVB-induced dermal fibroblast collagen degradation via hsa-microRNA-4535-mediated TGFβ/Smad signaling. Aging 2021, 13, 25342–25364. [Google Scholar] [PubMed]
- Wen, S.Y.; Chen, J.Y.; Weng, Y.; Aneja, R.; Chen, C.J.; Huang, C.Y.; Kuo, W.W. Galangin suppresses H2O2—induced aging in human dermal fibroblasts. Environ. Toxicol. 2017, 32, 2419–2427. [Google Scholar] [CrossRef]
- Wen, S.Y.; Chen, J.Y.; Chen, C.J.; Huang, C.Y.; Kuo, W.W. Protective effects of galangin against H2O2-induced aging via the IGF-1 signaling pathway in human dermal fibroblasts. Environ. Toxicol. 2020, 35, 115–123. [Google Scholar] [PubMed]
- Wu, Z.; Zheng, X.; Gong, M.; Li, Y. Myricetin, a potent natural agent for treatment of diabetic skin damage by modulating TIMP/MMPs balance and oxidative stress. Oncotarget 2016, 7, 71754–71760. [Google Scholar]
- Oh, J.H.; Karadeniz, F.; Lee, J.I.; Park, S.Y.; Seo, Y.; Kong, C.S. Anticatabolic and Anti-Inflammatory Effects of Myricetin 3-O-β-d-Galactopyranoside in UVA-Irradiated Dermal Cells via Repression of MAPK/AP-1 and Activation of TGFβ/Smad. Molecules 2020, 25, 1331. [Google Scholar] [CrossRef]
- Shon, M.S.; Kim, R.H.; Kwon, O.J.; Roh, S.S.; Kim, G.N. Beneficial role and function of fisetin in skin health via regulation of the CCN2/TGF-β signaling pathway. Food Sci. Biotechnol. 2016, 25, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Gopaul, R.; Knaggs, H.E.; Lephart, E.D. Biochemical investigation and gene analysis of equol: A plant and soy-derived isoflavonoid with antiaging and antioxidant properties with potential human skin applications. BioFactors 2012, 38, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Shi, Y.; Dang, Y.; Zhai, Y.; Ye, X. Daidzein stimulates collagen synthesis by activating the TGF-β/smad signal pathway. Australas. J. Dermatol. 2015, 56, 7–14. [Google Scholar] [CrossRef]
- Kim, Y.M.; Huh, J.S.; Lim, Y.; Cho, M. Soy Isoflavone Glycitin (4′-Hydroxy-6-Methoxyisoflavone-7-D-Glucoside) Promotes Human Dermal Fibroblast Cell Proliferation and Migration via TGF-β Signaling. Phytother. Res. 2015, 29, 757–769. [Google Scholar] [CrossRef]
- Lee, S.; Hoang, G.D.; Kim, D.; Song, H.S.; Choi, S.; Lee, D.; Kang, K.S. Efficacy of Alpinumisoflavone Isolated from Maclura tricuspidata Fruit in Tumor Necrosis Factor-α-Induced Damage of Human Dermal Fibroblasts. Antioxidants 2021, 10, 514. [Google Scholar] [CrossRef] [PubMed]
- Polito, F.; Marini, H.; Bitto, A.; Irrera, N.; Vaccaro, M.; Adamo, E.B.; Micali, A.; Squadrito, F.; Minutoli, L.; Altavilla, D. Genistein aglycone, a soy-derived isoflavone, improves skin changes induced by ovariectomy in rats. Br. J. Pharmacol. 2012, 165, 994–1005. [Google Scholar] [CrossRef]
- Nanashima, N.; Horie, K.; Maeda, H.; Tomisawa, T.; Kitajima, M.; Nakamura, T. Blackcurrant Anthocyanins Increase the Levels of Collagen, Elastin, and Hyaluronic Acid in Human Skin Fibroblasts and Ovariectomized Rats. Nutrients 2018, 10, 495. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.I.; Jeong, Y.U.; Kim, J.H.; Park, Y.J. 3,5,6,7,8,3′,4′-Heptamethoxyflavone, a Citrus Flavonoid, Inhibits Collagenase Activity and Induces Type I Procollagen Synthesis in HDFn Cells. Int. J. Mol. Sci. 2018, 19, 620. [Google Scholar] [CrossRef]
- Nisar, M.F.; Liu, T.; Wang, M.; Chen, S.; Chang, L.; Karisma, V.W.; Weixu; Diao, Q.; Xue, M.; Tang, X.; et al. Eriodictyol protects skin cells from UVA irradiation-induced photodamage by inhibition of the MAPK signaling pathway. J. Photochem. Photobiol. B 2022, 226, 112350. [Google Scholar]
- Chowdhury, A.; Nosoudi, N.; Karamched, S.; Parasaram, V.; Vyavahare, N. Polyphenol treatments increase elastin and collagen deposition by human dermal fibroblasts; Implications to improve skin health. J. Dermatol. Sci. 2021, 102, 94–100. [Google Scholar]
- Wang, L.; Lee, W.; Cui, Y.R.; Ahn, G.; Jeon, Y.J. Protective effect of green tea catechin against urban fine dust particle-induced skin aging by regulation of NF-κB, AP-1, and MAPKs signaling pathways. Environ. Pollut. 2019, 252, 1318–1324. [Google Scholar] [PubMed]
- Wang, L.; Kim, H.S.; Je, J.G.; Oh, J.Y.; Kim, Y.S.; Cha, S.H.; Jeon, Y.J. Protective Effect of Diphlorethohydroxycarmalol Isolated from Ishige okamurae Against Particulate Matter-Induced Skin Damage by Regulation of NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts. Molecules 2020, 25, 1055. [Google Scholar] [CrossRef] [PubMed]
- Xue, N.; Liu, Y.; Jin, J.; Ji, M.; Chen, X. Chlorogenic Acid Prevents UVA-Induced Skin Photoaging Through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts. Int. J. Mol. Sci. 2022, 23, 6941. [Google Scholar] [CrossRef] [PubMed]
- Matwiejczuk, N.; Galicka, A.; Zaręba, I.; Brzóska, M.M. The Protective Effect of Rosmarinic Acid Against Unfavorable Influence of Methylparaben and Propylparaben on Collagen in Human Skin Fibroblasts. Nutrients 2020, 12, 1282. [Google Scholar] [CrossRef] [PubMed]
- Galicka, A.; Sutkowska-Skolimowska, J. The Beneficial Effect of Rosmarinic Acid on Benzophenone-3-Induced Alterations in Human Skin Fibroblasts. Int. J. Mol. Sci. 2021, 22, 11451. [Google Scholar] [CrossRef]
- Sutkowska, J.; Hupert, N.; Gawron, K.; Strawa, J.W.; Tomczyk, M.; Forlino, A.; Galicka, A. The Stimulating Effect of Rosmarinic Acid and Extracts from Rosemary and Lemon Balm on Collagen Type I Biosynthesis in Osteogenesis Imperfecta Type I Skin Fibroblasts. Pharmaceutics 2021, 13, 938. [Google Scholar] [CrossRef]
- Hwang, E.; Park, S.Y.; Lee, H.J.; Lee, T.Y.; Sun, Z.W.; Yi, T.H. Gallic acid regulates skin photoaging in UVB-exposed fibroblast and hairless mice. Phytother. Res. 2014, 28, 1778–1788. [Google Scholar] [PubMed]
- Jeon, J.; Sung, J.; Lee, H.; Kim, Y.; Jeong, H.S.; Lee, J. Protective activity of caffeic acid and sinapic acid against UVB-induced photoaging in human fibroblasts. J. Food Biochem. 2019, 43, e12701. [Google Scholar]
- Ahn, H.S.; Kim, H.J.; Na, C.; Jang, D.S.; Shin, Y.K.; Lee, S.H. The Protective Effect of Adenocaulon himalaicum Edgew. and Its Bioactive Compound Neochlorogenic Acid against UVB-Induced Skin Damage in Human Dermal Fibroblasts and Epidermal Keratinocytes. Plants 2021, 10, 1669. [Google Scholar] [CrossRef]
- Duckworth, C.; Stutts, J.; Clatterbuck, K.; Nosoudi, N. Effect of ellagic acid and retinoic acid on collagen and elastin production by human dermal fibroblasts. Biomed. Mater. Eng. 2023, 34, 473–480. [Google Scholar]
- Park, H.J.; Cho, J.H.; Hong, S.H.; Kim, D.H.; Jung, H.Y.; Kang, I.K.; Cho, Y.J. Whitening and anti-wrinkle activities of ferulic acid isolated from Tetragonia tetragonioides in B16F10 melanoma and CCD-986sk fibroblast cells. J. Nat. Med. 2018, 72, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Hahn, H.J.; Kim, K.B.; Bae, S.; Choi, B.G.; An, S.; Ahn, K.J.; Kim, S.Y. Pretreatment of Ferulic Acid Protects Human Dermal Fibroblasts against Ultraviolet A Irradiation. Ann. Dermatol. 2016, 28, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Yoo, M.S.; Son, D.J.; Jung, H.Y.; Lee, S.H.; Jung, J.K.; Lee, B.C.; Yun, Y.P.; Pyo, H.B.; Hong, J.T. Increase of collagen synthesis by obovatol through stimulation of the TGF-beta signaling and inhibition of matrix metalloproteinase in UVB-irradiated human fibroblast. J. Dermatol. Sci. 2007, 46, 127–137. [Google Scholar] [PubMed]
- Lee, K.E.; Mun, S.; Pyun, H.B.; Kim, M.S.; Hwang, J.K. Effects of macelignan isolated from Myristica fragrans (Nutmeg) on expression of matrix metalloproteinase-1 and type I procollagen in UVB-irradiated human skin fibroblasts. Biol. Pharm. Bull. 2012, 35, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Tabandeh, M.R.; Oryan, A.; Mohhammad-Alipour, A.; Tabatabaei-Naieni, A. Silibinin regulates matrix metalloproteinase 3 (stromelysine1) gene expression, hexoseamines and collagen production during rat skin wound healing. Phytother. Res. 2013, 27, 1149–1153. [Google Scholar] [PubMed]
- Oh, J.H.; Joo, Y.H.; Karadeniz, F.; Ko, J.; Kong, C.S. Syringaresinol Inhibits UVA-Induced MMP-1 Expression by Suppression of MAPK/AP-1 Signaling in HaCaT Keratinocytes and Human Dermal Fibroblasts. Int. J. Mol. Sci. 2020, 21, 3981. [Google Scholar] [CrossRef]
- Zhou, X.; Ge, L.; Yang, Q.; Xie, Y.; Sun, J.; Cao, W.; Wang, S. Thinning of dermas with the increasing age may be against by tetrahydroxystilbene glucoside in mice. Int. J. Clin. Exp. Med. 2014, 7, 2017–2024. [Google Scholar]
- Ma, R.J.; Yang, L.; Bai, X.; Li, J.Y.; Yuan, M.Y.; Wang, Y.Q.; Xie, Y.; Hu, J.M.; Zhou, J. Phenolic Constituents with Antioxidative, Tyrosinase Inhibitory and Anti-aging Activities from Dendrobium loddigesii Rolfe. Nat. Prod. Bioprospect. 2019, 9, 329–336. [Google Scholar]
- Xia, Y.; Zhang, H.; Wu, X.; Xu, Y.; Tan, Q. Resveratrol activates autophagy and protects from UVA-induced photoaging in human skin fibroblasts and the skin of male mice by regulating the AMPK pathway. Biogerontology 2024, 25, 649–664. [Google Scholar] [CrossRef]
- Myung, D.B.; Han, H.S.; Shin, J.S.; Park, J.Y.; Hwang, H.J.; Kim, H.J.; Ahn, H.S.; Lee, S.H.; Lee, K.T. Hydrangenol Isolated from the Leaves of Hydrangea serrata Attenuates Wrinkle Formation and Repairs Skin Moisture in UVB-Irradiated Hairless Mice. Nutrients 2019, 11, 2354. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, S.R.; An, H.J.; Kim, W.J.; Choe, M.; Han, J.A. Esculetin promotes type I procollagen expression in human dermal fibroblasts through MAPK and PI3K/Akt pathways. Mol. Cell Biochem. 2012, 368, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Jo, H.S.; Shim, W.; Kwon, Y.W.; Bae, S.; Kwon, Y.; Azamov, B.; Hur, J.; Lee, D.; Ryu, S.H.; et al. Emodin induces collagen type I synthesis in Hs27 human dermal fibroblasts. Exp. Ther. Med. 2021, 21, 420. [Google Scholar] [PubMed]
- Kim, J.; Phung, H.M.; Lee, S.; Kim, K.T.; Son, T.K.; Kang, K.S.; Lee, S. Anti-skin-aging effects of tissue-cultured mountain-grown ginseng and quantitative HPLC/ELSD analysis of major ginsenosides. J. Nat. Med. 2022, 76, 811–820. [Google Scholar]
- Oh, J.H.; Kim, J.; Karadeniz, F.; Kim, H.R.; Park, S.Y.; Seo, Y.; Kong, C.S. Santamarine Shows Anti-Photoaging Properties via Inhibition of MAPK/AP-1 and Stimulation of TGF-β/Smad Signaling in UVA-Irradiated HDFs. Molecules 2021, 26, 3585. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.C.; Korivi, M.; Lin, F.Y.; Li, M.L.; Lin, R.W.; Wu, J.J.; Yang, H.L. Trans-cinnamic acid attenuates UVA-induced photoaging through inhibition of AP-1 activation and induction of Nrf2-mediated antioxidant genes in human skin fibroblasts. J. Dermatol. Sci. 2018, 90, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Jeong, S.H.; Kim, S.W. β-Lapachone Regulates the Transforming Growth Factor-β-Smad Signaling Pathway Associated with Collagen Biosynthesis in Human Dermal Fibroblasts. Biol. Pharm. Bull. 2016, 39, 524–531. [Google Scholar]
- Andrulewicz-Botulińska, E.; Kuźmicz, I.; Nazaruk, J.; Wosek, J.; Galicka, A. The concentration-dependent effect of anethole on collagen, MMP-2 and GAG in human skin fibroblast cultures. Adv. Med. Sci. 2019, 64, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.K. MAP kinase pathways. Cold Spring Harb. Perspect. Biol. 2012, 4, a011254. [Google Scholar] [CrossRef]
- Papakrivopoulou, J.; Lindahl, G.E.; Bishop, J.E.; Laurent, G.J. Differential roles of extracellular signal-regulated kinase 1/2 and p38MAPK in mechanical load-induced procollagen α1(I) gene expression in cardiac fibroblasts. Cardiovasc. Res. 2004, 61, 736–744. [Google Scholar]
- Yang, Y.; Kim, H.J.; Woo, K.J.; Cho, D.; Bang, S.I. Lipo-PGE1 suppresses collagen production in human dermal fibroblasts via the ERK/Ets-1 signaling pathway. PLoS ONE 2017, 12, e0179614. [Google Scholar] [CrossRef]
- Bhogal, R.K.; Bona, C.A. Regulatory effect of extracellular signal-regulated kinases (ERK) on type I collagen synthesis in human dermal fibroblasts stimulated by IL-4 and IL-13. Int. Rev. Immunol. 2008, 27, 472–496. [Google Scholar] [CrossRef] [PubMed]
- Reunanen, N.; Li, S.P.; Ahonen, M.; Foschi, M.; Han, J.; Kähäri, V.M. Activation of p38 alpha MAPK enhances collagenase-1 (matrix metalloproteinase (MMP)-1) and stromelysin-1 (MMP-3) expression by mRNA stabilization. J. Biol. Chem. 2002, 277, 32360–32368. [Google Scholar] [CrossRef]
- Cortez, D.M.; Feldman, M.D.; Mummidi, S.; Valente, A.J.; Steffensen, B.; Vincenti, M.; Barnes, J.L.; Chandrasekar, B. IL-17 stimulates MMP-1 expression in primary human cardiac fibroblasts via p38 MAPK- and ERK1/2-dependent C/EBP-beta, NF-kappaB, and AP-1 activation. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3356–H3365. [Google Scholar] [CrossRef]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed]
- Kook, S.H.; Jang, Y.S.; Lee, J.C. Involvement of JNK-AP-1 and ERK-NF-κB signaling in tension-stimulated expression of type I collagen and MMP-1 in human periodontal ligament fibroblasts. J. Appl. Physiol. 2011, 111, 1575–1583. [Google Scholar] [PubMed]
- Han, X.; Lei, Q.; Xie, J.; Liu, H.; Li, J.; Zhang, X.; Zhang, T.; Gou, X. Potential Regulators of the Senescence-Associated Secretory Phenotype During Senescence and Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 77, 2207–2218. [Google Scholar] [CrossRef]
- Salminen, A.; Kauppinen, A.; Kaarniranta, K. Emerging role of NF-κB signaling in the induction of senescence-associated secretory phenotype (SASP). Cell Signal. 2012, 24, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Park, H.; Kim, H.P. Effects of flavonoids on senescence-associated secretory phenotype formation from bleomycinin duced senescence in BJ fibroblasts. Biochem. Pharmacol. 2015, 96, 337–348. [Google Scholar] [CrossRef]
- Asano, Y.; Ihn, H.; Yamane, K.; Jinnin, M.; Mimura, Y.; Tamaki, K. Phosphatidylinositol 3-kinase is involved in alpha2(I) collagen gene expression in normal and scleroderma fibroblasts. J. Immunol. 2004, 172, 7123–7135. [Google Scholar] [CrossRef]
- Chetty, A.; Cao, G.J.; Nielsen, H.C. Insulin-like growth factor-I signaling mechanisms, type I collagen and alpha smooth muscle actin in human fetal lung fibroblasts. Pediatr. Res. 2006, 60, 389–394. [Google Scholar] [CrossRef]
- Kim, J.; Cha, Y.N.; Surh, Y.J. A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in inflammatory disorders. Mutat. Res. 2010, 690, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Panieri, E.; Telkoparan-Akillilar, P.; Saso, L. NRF2, a crucial modulator of skin cells protection against vitiligo, psoriasis, and cancer. Biofactors 2023, 49, 228–250. [Google Scholar] [CrossRef]
- Gruber, F.; Ornelas, C.M.; Karner, S.; Narzt, M.S.; Nagelreiter, M.I.; Gschwandtner, M.; Bochkov, V.; Tschachler, E. Nrf2 deficiency causes lipid oxidation, inflammation, and matrix-protease expression in DHA-supplemented and UVA-irradiated skin fibroblasts. Free Radic. Biol. Med. 2015, 88, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.S.; Crocker, S.J. An alternate perspective on the roles of TIMPs and MMPs in pathology. Am. J. Pathol. 2012, 180, 12–16. [Google Scholar] [CrossRef]
- Li, F.; Zhi, J.; Zhao, R.; Sun, Y.; Wen, H.; Cai, H.; Chen, W.; Jiang, X.; Bai, R. Discovery of matrix metalloproteinase inhibitors as anti-skin photoaging agents. Eur. J. Med. Chem. 2024, 267, 116152. [Google Scholar] [CrossRef] [PubMed]
- Surazynski, A.; Miltyk, W.; Palka, J.; Phang, J.M. Prolidase-dependent regulation of collagen biosynthesis. Amino Acids 2008, 35, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Gajko-Galicka, A. Mutations in type I collagen genes resulting in osteogenesis imperfecta in humans. Acta Biochim. Pol. 2002, 49, 433–441. [Google Scholar] [CrossRef]
- Tang, X.; Yang, T.; Yu, D.; Xiong, H.; Zhang, S. Current insights and future perspectives of ultraviolet radiation (UV) exposure: Friends and foes to the skin and beyond the skin. Environ. Int. 2024, 185, 108535. [Google Scholar] [CrossRef] [PubMed]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Photoaging: UV radiation-induced inflammation and immunosuppression accelerate the aging process in the skin. Inflamm. Res. 2022, 71, 817–831. [Google Scholar] [CrossRef]
- Papaccio, F.; Arino, A.D.; Caputo, S.; Bellei, B. Focus on the Contribution of Oxidative Stress in Skin Aging. Antioxidants 2022, 11, 1121. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef]
- Biskanaki, F.; Rallis, E.; Skouras, G.; Stofas, A.; Thymara, E.; Kavantzas, N.; Lazaris, A.C.; Kefala, V. Impact of Solar Ultraviolet Radiation in the Expression of Type I Collagen in the Dermis. Cosmetics 2021, 8, 46. [Google Scholar] [CrossRef]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.H.; Kim, S.Y. Resveratrol-enriched rice attenuates UVB-ROS-induced skin aging via downregulation of inflammatory cascades. Oxidat. Med. Cell Longevit. 2017, 2017, 8379539. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Lim, D.; Han, H.; Lee, S.; Choi, K.H.; Park, B.J. Emulsions stabilized by fine dust particles. J. Ind. Eng. Chem. 2020, 82, 190–196. [Google Scholar] [CrossRef]
- Anderson, J.O.; Thundiyil, J.G.; Stolbach, A. Clearing the air: A review of the effects of particulate matter air pollution on human health. J. Med. Toxicol. 2012, 8, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.; Feng, B.; Liang, H.; Zhao, X.; Song, J. Particulate matter and inflammatory skin diseases: From epidemiological and mechanistic studies. Sci. Total Environ. 2023, 905, 167111. [Google Scholar] [CrossRef]
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life Sci. 2016, 152, 126–134. [Google Scholar] [CrossRef]
- Reynolds, W.J.; Bowman, A.; Hanson, P.S.; Critchley, A.; Griffiths, B.; Chavan, B.; Birch-Machin, M.A. Adaptive responses to air pollution in human dermal fibroblasts and their potential roles in aging. FASEB Bioadv. 2021, 3, 855–865. [Google Scholar] [CrossRef]
- Vierkötter, A.; Schikowski, T.; Ranft, U.; Sugiri, D.; Matsui, M.; Krämer, U.; Krutmann, J. Airborne particle exposure and extrinsic skin aging. J. Investig. Dermatol. 2010, 130, 2719–2726. [Google Scholar] [CrossRef]
- Reynolds, W.J.; Hanson, P.S.; Critchley, A.; Griffiths, B.; Chavan, B.; Birch-Machin, M.A. Exposing human primary dermal fibroblasts to particulate matter induces changes associated with skin aging. FASEB J. 2020, 34, 14725–14735. [Google Scholar] [CrossRef]
- Darbre, P.D.; Harvey, P.W. Paraben esters: Review of recent studies of endocrine toxicity, absorption, esterase and human exposure, and discussion of potential human health risks. J. Appl. Toxicol. 2008, 28, 561–578. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.J.; Bae, S.; Kim, K.; Kwon, S.B.; An, I.S.; Ahn, K.J.; Ryu, J.; Kim, H.S.; Ye, S.K.; Kim, B.H.; et al. Overdosage of methylparaben induces cellular senescence In Vitro and In Vivo. J. Investig. Dermatol. 2015, 135, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Adhikary, S.; Bhattacharya, S.; Chakraborty, A.; Dutta, S.; Roy, D.; Ganguly, A.; Nanda, S.; Rajak, P. Parabens as the double-edged sword: Understanding the benefits and potential health risks. Sci. Total Environ. 2024, 954, 176547. [Google Scholar]
- Dubey, D.; Chopra, D.; Singh, J.; Srivastav, A.K.; Kumari, S.; Verma, A.; Ray, R.S. Photosensitized methyl paraben induces apoptosis via caspase dependent pathway under ambient UVB exposure in human skin cells. Food Chem. Toxicol. 2017, 108, 171–185. [Google Scholar] [CrossRef]
- Ngoc, L.T.N.; Tran, V.V.; Moon, J.Y.; Chae, M.; Park, D.; Lee, Y.C. Recent trends of sunscreen cosmetic: An update review. Cosmetics 2019, 6, 64. [Google Scholar] [CrossRef]
- Janjua, N.R.; Mogensen, B.; Andersson, A.M.; Petersen, J.H.; Henriksen, M.; Skakkebaek, N.E.; Wulf, H.C. Systemic absorption of the sunscreens benzophenone-3, octyl-methoxycinnamate, and 3-(4-methylbenzylidene) camphor after whole-body topical application and reproductive hormone levels in humans. J. Investig. Dermatol. 2004, 123, 57–61. [Google Scholar]
- Barr, L.; Alamer, M.; Darbre, P.D. Measurement of concentrations of four chemical ultraviolet filters in human breast tissue at serial locations across the breast. J. Appl. Toxicol. 2018, 38, 1112–1120. [Google Scholar] [CrossRef]
- Hayden, C.G.; Roberts, M.S.; Benson, H.A. Systemic absorption of sunscreen after topical application. Lancet 1997, 350, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Zduńska-Pęciak, K.; Dębowska, R.; Kołodziejczak, A.; Rotsztejn, H. Ferulic acid—A novel topical agent in reducing signs of photoaging. Dermatol. Ther. 2022, 35, e15543. [Google Scholar]
- Cavalcanti, G.R.; Duarte, F.I.C.; Converti, A.; de Lima, Á.A.N. Ferulic Acid Activity in Topical Formulations: Technological and Scientific Prospecting. Curr. Pharm. Des. 2021, 27, 2289–2298. [Google Scholar] [CrossRef] [PubMed]
- Igielska-Kalwat, J.; Firlej, M.; Lewandowska, A.; Biedziak, B. In vivo studies of resveratrol contained in cosmetic emulsions. Acta Biochim. Pol. 2019, 66, 371–374. [Google Scholar] [CrossRef]
- Bonato Alves Oliveira, L.; Oliveira, R.P.d.; Oliveira, C.; Raposo, N.R.B.; Brandão, M.A.F.; Ferreira, A.D.O.; Polonini, H. Cosmetic Potential of a Liotropic Liquid Crystal Emulsion Containing Resveratrol. Cosmetics 2017, 4, 54. [Google Scholar] [CrossRef]
- Moyano-Mendez, J.R.; Fabbrocini, G.; De Stefano, D.; Mazzella, C.; Mayol, L.; Scognamiglio, I.; Carnuccio, R.; Ayala, F.; La Rotonda, M.I.; De Rosa, G. Enhanced antioxidant effect of trans-resveratrol: Potential of binary systems with polyethylene glycol and cyclodextrin. Drug Dev. Ind. Pharm. 2014, 40, 1300–1307. [Google Scholar] [CrossRef]
- Farris, P.; Yatskayer, M.; Chen, N.; Krol, Y.; Oresajo, C. Evaluation of efficacy and tolerance of a nighttime topical antioxidant containing resveratrol, baicalin, and vitamin e for treatment of mild to moderately photodamaged skin. J. Drugs Dermatol. 2014, 13, 1467–1472. [Google Scholar] [PubMed]
- Wu, Y.; Jia, L.L.; Zheng, Y.N.; Xu, X.G.; Luo, Y.J.; Wang, B.; Chen, J.Z.S.; Gao, X.H.; Chen, H.D.; Matsui, M.; et al. Resveratrate protects human skin from damage due to repetitive ultraviolet irradiation. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 345–350. [Google Scholar] [CrossRef]
- Woon, C.G.; Jin, J.H.; Kyung, S.J.; Hwoon, B.J.; Mi, K.Y.; Chool, B.Y. Skin Anti-aging Effects of a Cream Containing Resveratryl Triacetate (RTA). J. Soc. Cosmet. Sci. Korea 2018, 44, 161–170. [Google Scholar]
- Brinke, A.S.; Janssens-Böcker, C.; Kerscher, M. Skin Anti-Aging Benefits of a 2% Resveratrol Emulsion. J. Cosmet. Dermatol. Sci. Appl. 2021, 11, 155–168. [Google Scholar] [CrossRef]
- Silva, L.A.; Ferraz Carbonel, A.A.; de Moraes, A.R.B.; Simões, R.S.; Sasso, G.R.D.S.; Goes, L.; Nunes, W.; Simões, M.J.; Patriarca, M.T. Collagen concentration on the facial skin of postmenopausal women after topical treatment with estradiol and genistein: A randomized double-blind controlled trial. Gynecol. Endocrinol. 2017, 33, 845–848. [Google Scholar] [CrossRef]
- Ud-Din, S.; Foden, P.; Mazhari, M.; Al-Habba, S.; Baguneid, M.; Bulfone-Paus, S.; McGeorge, D.; Bayat, A. A Double-Blind, Randomized Trial Shows the Role of Zonal Priming and Direct Topical Application of Epigallocatechin-3-Gallate in the Modulation of Cutaneous Scarring in Human Skin. J. Investig. Dermatol. 2019, 139, 1680–1690.e16. [Google Scholar] [CrossRef]
- Magnet, U.; Urbanek, C.; Gaisberger, D.; Tomeva, E.; Dum, E.; Pointner, A.; Haslberger, A.G. Topical equol preparation improves structural and molecular skin parameters. Int. J. Cosmet. Sci. 2017, 39, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Skibola, C.F.; Smith, M.T. Potential health impacts of excessive flavonoid intake. Free Radic. Biol. Med. 2000, 29, 375–383. [Google Scholar] [PubMed]
- Tang, Z.; Zhang, Q. The potential toxic side effects of flavonoids. Biocell 2021, 46, 357–366. [Google Scholar]
 inhibition (in green indicates upregulation and in red, downregulation of collagen biosynthesis). Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COL1A1—gene encoding the pro-alpha1 chains of type I collagen; COL1A2—gene encoding the pro-alpha2 chains of type I collagen; ERK—extracellular signal-regulated kinase; Gly—glycine; GSH—gene encoding glutathione; HO-1—gene encoding heme oxygenase 1; IGF-1—insulin-like growth factor 1; IL—gene encoding interleukin; IκBα—inhibitor of NF-κB; Keap-1—Kelch-like ECH-associated protein 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; MMP—gene encoding matrix metalloproteinase; mTOR—mammalian target of rapamycin; NF-κB—nuclear factor-kappa B; Nrf-2—nuclear factor erythroid 2-related factor 2; PI3K—phosphoinositide 3-kinase; IRS-1/2—insulin receptor substrate 1/2; JNK—Jun N-terminal kinase; P—phosphorylated; Pro—proline; Smad—suppressor of mothers against decapentaplegic; Raf—rapidly accelerated fibrosarcoma; Ras—retrovirus-associated sequence; ROS—reactive oxygen species; SOD—gene encoding superoxide dismutase; TGF-β1—transforming growth factor beta 1; TNF-α—gene encoding tumor necrosis factor alpha; Ub—ubiquitin; α, β—subunits of integrin receptor. The figure was made using the templates available on the Servier Medical Art website (Creative Commons Attribution 4.0 unported license). (b) Mechanisms of action of the natural compounds on collagen type I. ↑—stimulation; ↓—inhibition. Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COX-2—cyclooxygenase-2; ERK—extracellular signal-regulated kinase; GCLC—glutamate cysteine ligase catalysis subunit; GCLM—glutamate cysteine ligase modifier subunit; GLT25D1—gene encoding collagen galactosyltransferase 1; hsa-miR—homo sapiens microRNA; HO-1—heme oxygenase 1; HSP47—heat shock protein 47; IGF-1—insulin-like growth factor 1; IGF-1R—insulin-like growth factor 1 receptor; IL—interleukin 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; NF-κB—nuclear factor-kappa B; NQO-1—NAD(P)H quinone oxidoreductase 1; Nrf-2—nuclear factor erythroid 2-related factor 2; pAkt—phosphorylated protein kinase B; pc-Fos—phosphorylated c-Fos protein; pc-Jun—phosphorylated c-Jun protein; PDI—gene encoding protein disulfide isomerase; pERK1/2—phosphorylated extracellular signal-regulated kinase 1/2; PI3K—phosphoinositide 3-kinase; pJNK—phosphorylated Jun N-terminal kinase; pmTOR—phosphorylated mammalian target of rapamycin; pNF-κB—phosphorylated nuclear factor-kappa B; pNrf-2—phosphorylated nuclear factor erythroid 2-related factor 2; pp38—phosphorylated p38 protein; pSmad—phosphorylated suppressor of mothers against decapentaplegic; ROS—reactive oxygen species; Smad—suppressor of mothers against decapentaplegic; SOD-1—superoxide dismutase 1; TGF-β1—transforming growth factor beta 1; TIMP—tissue inhibitor of metalloproteinase; TNF-α—tumor necrosis factor alpha.
inhibition (in green indicates upregulation and in red, downregulation of collagen biosynthesis). Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COL1A1—gene encoding the pro-alpha1 chains of type I collagen; COL1A2—gene encoding the pro-alpha2 chains of type I collagen; ERK—extracellular signal-regulated kinase; Gly—glycine; GSH—gene encoding glutathione; HO-1—gene encoding heme oxygenase 1; IGF-1—insulin-like growth factor 1; IL—gene encoding interleukin; IκBα—inhibitor of NF-κB; Keap-1—Kelch-like ECH-associated protein 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; MMP—gene encoding matrix metalloproteinase; mTOR—mammalian target of rapamycin; NF-κB—nuclear factor-kappa B; Nrf-2—nuclear factor erythroid 2-related factor 2; PI3K—phosphoinositide 3-kinase; IRS-1/2—insulin receptor substrate 1/2; JNK—Jun N-terminal kinase; P—phosphorylated; Pro—proline; Smad—suppressor of mothers against decapentaplegic; Raf—rapidly accelerated fibrosarcoma; Ras—retrovirus-associated sequence; ROS—reactive oxygen species; SOD—gene encoding superoxide dismutase; TGF-β1—transforming growth factor beta 1; TNF-α—gene encoding tumor necrosis factor alpha; Ub—ubiquitin; α, β—subunits of integrin receptor. The figure was made using the templates available on the Servier Medical Art website (Creative Commons Attribution 4.0 unported license). (b) Mechanisms of action of the natural compounds on collagen type I. ↑—stimulation; ↓—inhibition. Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COX-2—cyclooxygenase-2; ERK—extracellular signal-regulated kinase; GCLC—glutamate cysteine ligase catalysis subunit; GCLM—glutamate cysteine ligase modifier subunit; GLT25D1—gene encoding collagen galactosyltransferase 1; hsa-miR—homo sapiens microRNA; HO-1—heme oxygenase 1; HSP47—heat shock protein 47; IGF-1—insulin-like growth factor 1; IGF-1R—insulin-like growth factor 1 receptor; IL—interleukin 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; NF-κB—nuclear factor-kappa B; NQO-1—NAD(P)H quinone oxidoreductase 1; Nrf-2—nuclear factor erythroid 2-related factor 2; pAkt—phosphorylated protein kinase B; pc-Fos—phosphorylated c-Fos protein; pc-Jun—phosphorylated c-Jun protein; PDI—gene encoding protein disulfide isomerase; pERK1/2—phosphorylated extracellular signal-regulated kinase 1/2; PI3K—phosphoinositide 3-kinase; pJNK—phosphorylated Jun N-terminal kinase; pmTOR—phosphorylated mammalian target of rapamycin; pNF-κB—phosphorylated nuclear factor-kappa B; pNrf-2—phosphorylated nuclear factor erythroid 2-related factor 2; pp38—phosphorylated p38 protein; pSmad—phosphorylated suppressor of mothers against decapentaplegic; ROS—reactive oxygen species; Smad—suppressor of mothers against decapentaplegic; SOD-1—superoxide dismutase 1; TGF-β1—transforming growth factor beta 1; TIMP—tissue inhibitor of metalloproteinase; TNF-α—tumor necrosis factor alpha.
inhibition (in green indicates upregulation and in red, downregulation of collagen biosynthesis). Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COL1A1—gene encoding the pro-alpha1 chains of type I collagen; COL1A2—gene encoding the pro-alpha2 chains of type I collagen; ERK—extracellular signal-regulated kinase; Gly—glycine; GSH—gene encoding glutathione; HO-1—gene encoding heme oxygenase 1; IGF-1—insulin-like growth factor 1; IL—gene encoding interleukin; IκBα—inhibitor of NF-κB; Keap-1—Kelch-like ECH-associated protein 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; MMP—gene encoding matrix metalloproteinase; mTOR—mammalian target of rapamycin; NF-κB—nuclear factor-kappa B; Nrf-2—nuclear factor erythroid 2-related factor 2; PI3K—phosphoinositide 3-kinase; IRS-1/2—insulin receptor substrate 1/2; JNK—Jun N-terminal kinase; P—phosphorylated; Pro—proline; Smad—suppressor of mothers against decapentaplegic; Raf—rapidly accelerated fibrosarcoma; Ras—retrovirus-associated sequence; ROS—reactive oxygen species; SOD—gene encoding superoxide dismutase; TGF-β1—transforming growth factor beta 1; TNF-α—gene encoding tumor necrosis factor alpha; Ub—ubiquitin; α, β—subunits of integrin receptor. The figure was made using the templates available on the Servier Medical Art website (Creative Commons Attribution 4.0 unported license). (b) Mechanisms of action of the natural compounds on collagen type I. ↑—stimulation; ↓—inhibition. Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COX-2—cyclooxygenase-2; ERK—extracellular signal-regulated kinase; GCLC—glutamate cysteine ligase catalysis subunit; GCLM—glutamate cysteine ligase modifier subunit; GLT25D1—gene encoding collagen galactosyltransferase 1; hsa-miR—homo sapiens microRNA; HO-1—heme oxygenase 1; HSP47—heat shock protein 47; IGF-1—insulin-like growth factor 1; IGF-1R—insulin-like growth factor 1 receptor; IL—interleukin 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; NF-κB—nuclear factor-kappa B; NQO-1—NAD(P)H quinone oxidoreductase 1; Nrf-2—nuclear factor erythroid 2-related factor 2; pAkt—phosphorylated protein kinase B; pc-Fos—phosphorylated c-Fos protein; pc-Jun—phosphorylated c-Jun protein; PDI—gene encoding protein disulfide isomerase; pERK1/2—phosphorylated extracellular signal-regulated kinase 1/2; PI3K—phosphoinositide 3-kinase; pJNK—phosphorylated Jun N-terminal kinase; pmTOR—phosphorylated mammalian target of rapamycin; pNF-κB—phosphorylated nuclear factor-kappa B; pNrf-2—phosphorylated nuclear factor erythroid 2-related factor 2; pp38—phosphorylated p38 protein; pSmad—phosphorylated suppressor of mothers against decapentaplegic; ROS—reactive oxygen species; Smad—suppressor of mothers against decapentaplegic; SOD-1—superoxide dismutase 1; TGF-β1—transforming growth factor beta 1; TIMP—tissue inhibitor of metalloproteinase; TNF-α—tumor necrosis factor alpha.
inhibition (in green indicates upregulation and in red, downregulation of collagen biosynthesis). Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COL1A1—gene encoding the pro-alpha1 chains of type I collagen; COL1A2—gene encoding the pro-alpha2 chains of type I collagen; ERK—extracellular signal-regulated kinase; Gly—glycine; GSH—gene encoding glutathione; HO-1—gene encoding heme oxygenase 1; IGF-1—insulin-like growth factor 1; IL—gene encoding interleukin; IκBα—inhibitor of NF-κB; Keap-1—Kelch-like ECH-associated protein 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; MMP—gene encoding matrix metalloproteinase; mTOR—mammalian target of rapamycin; NF-κB—nuclear factor-kappa B; Nrf-2—nuclear factor erythroid 2-related factor 2; PI3K—phosphoinositide 3-kinase; IRS-1/2—insulin receptor substrate 1/2; JNK—Jun N-terminal kinase; P—phosphorylated; Pro—proline; Smad—suppressor of mothers against decapentaplegic; Raf—rapidly accelerated fibrosarcoma; Ras—retrovirus-associated sequence; ROS—reactive oxygen species; SOD—gene encoding superoxide dismutase; TGF-β1—transforming growth factor beta 1; TNF-α—gene encoding tumor necrosis factor alpha; Ub—ubiquitin; α, β—subunits of integrin receptor. The figure was made using the templates available on the Servier Medical Art website (Creative Commons Attribution 4.0 unported license). (b) Mechanisms of action of the natural compounds on collagen type I. ↑—stimulation; ↓—inhibition. Akt—protein kinase B; AP-1—activator protein 1; ARE—antioxidant response element; COX-2—cyclooxygenase-2; ERK—extracellular signal-regulated kinase; GCLC—glutamate cysteine ligase catalysis subunit; GCLM—glutamate cysteine ligase modifier subunit; GLT25D1—gene encoding collagen galactosyltransferase 1; hsa-miR—homo sapiens microRNA; HO-1—heme oxygenase 1; HSP47—heat shock protein 47; IGF-1—insulin-like growth factor 1; IGF-1R—insulin-like growth factor 1 receptor; IL—interleukin 1; MAPK—mitogen-activated protein kinase; MMP—matrix metalloproteinase; NF-κB—nuclear factor-kappa B; NQO-1—NAD(P)H quinone oxidoreductase 1; Nrf-2—nuclear factor erythroid 2-related factor 2; pAkt—phosphorylated protein kinase B; pc-Fos—phosphorylated c-Fos protein; pc-Jun—phosphorylated c-Jun protein; PDI—gene encoding protein disulfide isomerase; pERK1/2—phosphorylated extracellular signal-regulated kinase 1/2; PI3K—phosphoinositide 3-kinase; pJNK—phosphorylated Jun N-terminal kinase; pmTOR—phosphorylated mammalian target of rapamycin; pNF-κB—phosphorylated nuclear factor-kappa B; pNrf-2—phosphorylated nuclear factor erythroid 2-related factor 2; pp38—phosphorylated p38 protein; pSmad—phosphorylated suppressor of mothers against decapentaplegic; ROS—reactive oxygen species; Smad—suppressor of mothers against decapentaplegic; SOD-1—superoxide dismutase 1; TGF-β1—transforming growth factor beta 1; TIMP—tissue inhibitor of metalloproteinase; TNF-α—tumor necrosis factor alpha.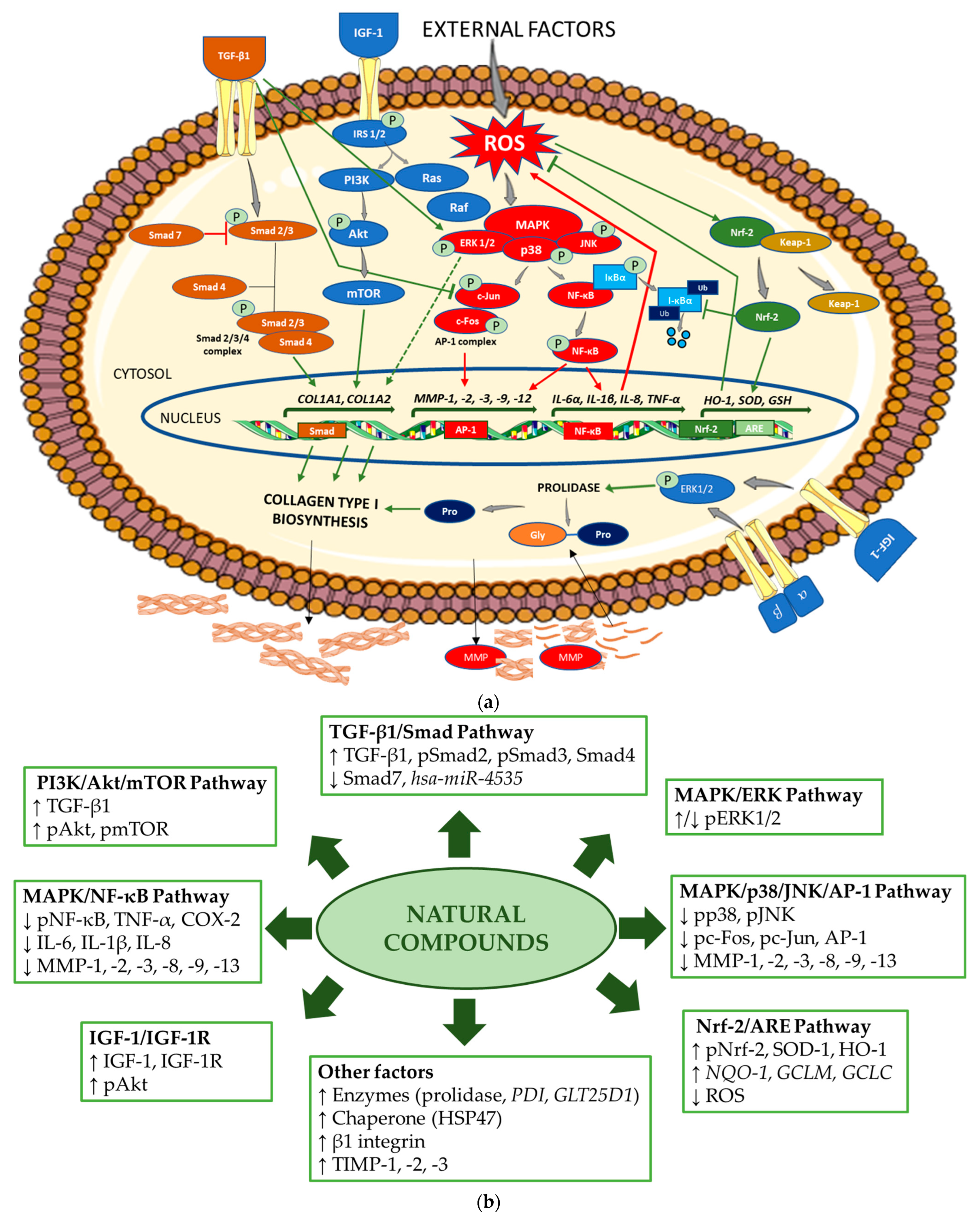

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Compound/ Structural Formula | Research Model (Concentration) | Stimuli (Intensity) | Effects/Mechanisms of Action | Ref. |
|---|---|---|---|---|
| FLAVONOIDS | ||||
| Flavones | ||||
Apigenin | HDFs (0.1–10 µM) C57BL/6 mice (5 µM) | None | ↑ COL1A2, COL3A1 ↑ pSmad2, pSmad3 (-) MMP-1, MMP-2, MMP-9, TIMP-1 (-) pJNK, pERK, p38 ↑ collagen density, dermal thickness | [51] |
Apigenin 7-O-glucuronide Apigenin 7-O-methyl- glucuronide  Pectolinarin  | HDFs OI (30 µM) | None | ↑ total collagen synthesis (assay with 5-[3H]proline) ↑ secretion of collagen ↑ prolidase activity, IGF-1R, β1 integrin (-) MMP-2, MMP-9, MMP-3 | [52] |
Baicalin | C57BL/6 mice (0.5 and 1 mg/cm2 skin area) HDFs (6.25–25 µg/mL) | UVB (6.9 J/cm2) UVB (10 mJ/cm2) | ↑ COL1A1, COL3A1 ↑ procollagen type I and III in the skin ↓ MMP-1, MMP-3, MMP-1, MMP-3 ↓ epidermal thickness ↓ premature senescence ↓ p53, p21, p16, γ-H2AX | [53] |
Luteolin | HDFs (10, 20, 40 µM) SD rats (60 and 120 mg/kg) | UVB (20 mJ/cm2) UVB (300 mJ/cm2) | ↑ collagen type I ↑ TGF-β, Smad3, SIRT3, SOD, ROS ↓ p38, pJNK, c-Jun, MMP-1, MMP-3 ↑ collagen type I ↑ TGF-β, Smad3, SIRT3, SOD, ROS ↓ p38, pJNK, c-Jun, MMP-1, MMP-3 ↓ skin damage, erythema, wrinkles | [54] |
| 6-hydroxyluteolin 7-O-β- glucoside  Pedalitin 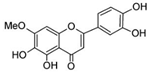 | HDFs (1, 2, 40 μM) | None | ↑ COL1A1, COL1A2, COL3A1 ↑ total soluble collagen ↑ HSP47 ↓ MMP-2 | [55] |
Chrysin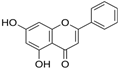 | HDFs (20, 40, 80 µM) | None | ↑ total collagen (hydroxyproline content) | [56] |
| Flavonols | ||||
Morin 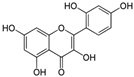 | HDFs (80 µM) | None | ↑ total collagen (hydroxyproline content) | [56] |
Rutin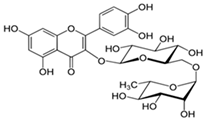 | HDFs (80 µM) | None | ↑ total collagen (hydroxyproline content) | [56] |
| HDFs (1, 10, 50 µM) | H2O2 (0.2 mM) | ↑ COL1A1 ↓ MMP-1 ↓ ROS | [57] | |
Galangin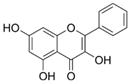 | HDF HS68 (30 μM) C57BL/6J mice (12 and 24 mg/kg) | H2O2 (0.2 mM) UVB (40 mJ/cm2) UVB (150 mJ/cm2) | ↑ collagen type I and III ↑ TGF-β, Smad4, p-Smad2/3 ↓ hsa-miR-4535, MMP-1 ↑ collagen content and density ↓ wrinkle formation ↑ TGF-β, Smad4, pSmad2/3 ↓ hsa-miR-4535, MMP1 | [58] |
| HDF Hs68 (10 and 30 µM) | H2O2 (0.2 mM) | ↑ collagen type I and III ↑ IGF1-R, pAkt, pERK, pNrf2, HO-1, Bcl-xL ↓ pNF-κB, IL-6, IL-1β, TNF-α ↓ p65, p53, p16, p21 | [59] | |
| HDF Hs68 (30 μM) | None H2O2 (0.2 mM) | ↑ collagen type I and III ↑ IGF-1R, p-ERK ↑ collagen type I and III ↑ IGF-1R, p-ERK ↓ pNF-κB, IL-1β, IL-6, TNF-α, MMP-1 ↓ SA-β-gal, p53, p21, p16 | [60] | |
Myricetin  | HDFs (3 μM) | None | ↑ procollagen type I and III ↓ MMP-1, MMP-2, MMP-9 ↑ TIMP-1, TIMP1/MMP ratio | [61] |
| Myricetin 3-O-β-galacto- pyranoside 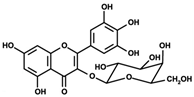 | HDFs (1, 5, 25 μM) | UVA (19 J/cm2) | ↑ collagen type I ↑ TGF-β, pSmad2/3, Smad4, pERK ↓ MMP-1, MMP-3, MMP-9 ↓ COX-2, iNOS, TNF-α, IL-1β ↓ pp38, pJNK, pc-Fos, pc-Jun | [62] |
Fisetin | HDFs (10, 25, 50 µM) | None | ↑ COL1A2 ↑ CCN2, Smad 2 ↑ CCN2, TGF-β1, TGF-β2, TGF-β3 | [63] |
| Isoflavons | ||||
Equol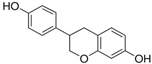 | HDFs and 3D HDFs (10 nM) Human skin barrier equivalents (EFT cultures) (1.2%) | None | ↑ collagen type I C-terminal propeptide ↑ collagen type I and III ↓ MMP-1, MMP-3 ↑ COL1A1, TIMP-1 ↓ MMP-1, MMP-3, MMP-9 | [64] |
Daidzein | nHDFs (0.5, 5, 50 μg/mL) BALB/C mice (200 μg/mL) | None | ↑ COL1A1 ↑ TGF-β, p-Smad2/3, TIMP-1 ↓ MMP-1, MMP-2 ↑ COL1A1, collagen type I ↑ TGF-β ↓ MMP-1, MMP-2 | [65] |
Glycitin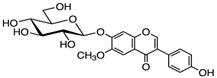 | HDFs (20 µM) | None | ↑ COL1A1, COL3A1, collagen type I and III ↑ TGFβ-1, TGFβ-1, pAKT, pmTOR, MMP-2 ↓ MMP-1 | [66] |
Alpinumisoflavone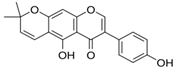 | HDFs (25 and 50 µM) | TNF-α (20 ng/mL) | ↑ COL1A1 ↓ MMP-1, MMP-1, ROS, iNOS, COX-2 ↓ IL-1β, IL-6, IL-8, IL-1β, IL-6, IL-8, ↓ NF-κB (p65), AP-1 ↓ pERK, pJNK, pp38 | [67] |
Genistein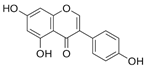 | SD rats (1 and 10 mg/kg) daily, for 12 weeks, subcutaneous administration | None | ↑ collagen thickness ↑ TGF-β1, MMP-2, MMP-9 ↑ TIMP-1, TIMP-2 ↑ skin breaking strength | [68] |
| Anthocyanins | ||||
Cyanidin-3-glucoside (C3G)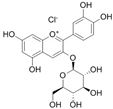 Cyanidin-3-rutinoside (C3R)  Delphinidin-3-glucoside (D3G)  Delphinidin-3-rutinoside (D3R)  | HDFs TIG113 (C3G, C3R, D3G and D3R—10 µM) BCE (1–10 µg/mL) OVX SD rats (BCE 3%—dietary administration) | None | ↑ COL1A1, COL3A1, TIMP-3 ↑ collagen type I and III ↑ TGF-β, IGF-2, IGFBP2, IGFBP5 ↑ collagen thickness | [69] |
| Polymethoxyflavones | ||||
| 3,5,6,7,8,3′,4′-Heptametho- xyflavone 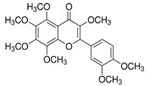 | nHDFs (50, 100, 200 μg/mL) | UVB (20 mJ/cm2) | ↑ procollagen type I C-peptide ↑ Smad3 ↓ Smad7, MMP1, pERK, pJNK, pc-Jun, c-Fos (-) pp38 | [70] |
| Flavanones | ||||
Eriodictyol | HDFs FEK-4 (2.5, 5, 10, 20, 40 µM) | UVA (150 KJ/m2) | ↑ COL1A1 ↓ MMP-1, ROS, pp38, pERK, pJNK ↓ IL-1β, IL-6, TNFα, TGFβ, COX-2, NF-κB ↑ TIMP-1, SOD | [71] |
| Flavanols | ||||
Epigallocatechin gallate  | HDFs (10 µg/mL) | UVA (10 mW/cm2) | ↑ total collagen (soluble and matrix) | [72] |
| HDFs (12.5, 25, 50 µM) | FDPs (ERM-CZ100) (200 µg/mL) | ↑ procollagen content ↓ MMP-1, MMP-2, MMP-8, MMP-9, MMP-13 ↓ collagenase activity ↓ ROS, p50, p65, pc-Jun, pERK, pJNK, pp38 | [73] | |
| TANNINS | ||||
Pentagalloyl glucose  | HDFs (10 µg/mL) | UVA (10 mW/cm2) | ↑ total collagen (soluble and matrix) | [72] |
Diphlorethohydroxycarmalol | HDFs (25, 50, 100 µM) | PMs (ERM-CZ100) (200 µg/mL) | ↑ collagen content ↓ MMP-1, MMP-2, MMP-8, MMP-9, MMP-13 ↓ collagenase activity ↓ ROS, p50, p65, pc-Jun, pERK, pJNK, pp38 | [74] |
| PHENOLIC ACIDS | ||||
Chlorogenic acid  | eHDFs CCC-ESF-1 (0.1–10 µM) HDFα (3 and 30 µM) | None UVA (12 J/cm2) | ↑ COL1A2, collagen type I ↑ collagen type I secretion ↑ COL1A1, collagen type I ↑ p-Smad2/3, Rad51 ↓ MMP-3, ROS, C-PARP, γ-H2AX (-) COL1A2, COL3A1, COL5A1, MMP-1, MMP-3 (-) pERK | [75] |
Rosmarinic acid | HDFs CRL-1474 (50–150 μM) | MP and PP (0.001% and 0.0003%; 0.003% and 0.001%; 0.01% and 0.003%) | ↑ COL1A1, COL1A2, COL3A1, HSP47 ↑ collagen type I, HSP47 ↓ MMP-1, MMP-2, MT1-MMP ↓ MMP-1, MMP-2, MT1-MMP ↑ TIMP-1, TIMP-2 ↓ pERK1/2 | [76] |
| HDFs CRL-1474 (100 μM) | BP-3 (0.1–100 µM) | ↑ COL1A1, PDI, GLT25D1 ↑ collagen type I ↓ MMP-1, MMP-2, MMP-1, MMP-2 | [77] | |
| HDFs OI (0.1, 1, 10 µM) | None | ↑ COL1A1 ↑ procollagen type I, collagen type I ↓ MMP-2, MMP-9 | [78] | |
Gallic acid | HDFs (1 and 10 µM) HDFs (1 and 10 µM) SKH-1 mice (1 and 5%) | None UVB (144 mJ/cm2) UVB (100–200 mJ/cm2) | ↑ procollagen type I ↓ pc-Fos, pc-Jun, MMP-1, IL-6 ↑ procollagen type I ↓ pc-Fos, pc-Jun, MMP-1, IL-6 ↑ procollagen type I, TGF-β1 ↓ ROS, MMP-1, IL-6 ↓ wrinkle formation | [79] |
Caffeic acid  Sinapic acid  | HDF Hs68 (100 µM) | UVB (30 mJ/cm2) | ↑ total soluble collagen ↓ MMP-1, pERK, pJNK, pp38, NF-KB p50, ROS | [80] |
Neochlorogenic acid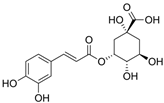 | HDF Hs68 (50, 100, 200 μM) | UVB (15 mJ/cm2) | ↑ procollagen type I ↓ MMP-1, pp38, pERK, pc-Fos, pc-Jun (-) pJNK | [81] |
Ellagic acid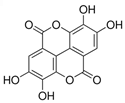 | HDFs (2 µg/mL) | None | ↑ COL1A1 ↑ collagen deposition in the ECM | [82] |
Ferulic acid | HDF CCD-986sk (5, 10, 20 µg/mL) | UVB (20 mJ/cm2) | ↑ procollagen type I ↑ TIMP-1 ↓ MMP-1 | [83] |
| HDFs (10 and 20 µM) | UVA (10 J/cm2) | ↑ SOD, CAT ↓ MMP-1, MMP-3, p16 ↓ ROS ↓ % of senescent cells | [84] | |
| LIGNANS | ||||
Obovatol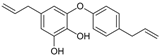 | HDFs (1, 2.5, 5 μM) | UVB (40 mJ/cm2) | ↑ procollagen type I ↑ TGF-β, Smad3 ↓ MMP-3, Smad7, AP-1, c-Fos, c-Jun ↓ pERK, pJNK, pp38 | [85] |
Macelignan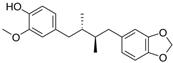 | HDF Hs68 (1, 5, 10 μM) | UVB (20 mJ/cm2) | ↑ COL1A1, procollagen type I C-peptide ↑ Smad3 ↓ Smad7, MMP1, MMP-1, ROS ↓ pERK, pJNK, pp38, pc-Jun (-) c-Fos | [86] |
Silibinin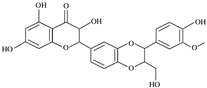 | Wistar rats with skin wound (10% and 20% (w/v) powder) | None | ↑ collagen (hydroxyproline content) ↑ MMP-3 ↑ closure of wounded skin | [87] |
Syringaresinol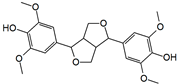 | HDFs (1, 5, 20 µM) | UVA (10 J/cm2) | ↑ procollagen type I, pERK ↓ MMP-1, MMP-9, TNF-α, COX-2, IL-1β ↓ JNK, pc-Fos, pc-Jun | [88] |
| STILBENS | ||||
Tetrahydroxystilbene glucoside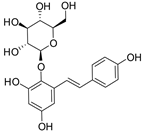 | Kunming mice (180 mg/kg, gastric lavage) | None | ↑ collagen content ↑ thickness of dermal layer ↓ IGF-1, IGF-1R | [89] |
Batatasin III | HDFs (3.182 μg/mL) | ↑ procollagen | [90] | |
Resveratrol | HDFs (100 μM) BALB/C mice (100 μM) | UVA (16 J/cm2) UVA (0.35 J/cm2, with increase of 5% per day) | ↑ collagen type I ↓ MMP-1, SA-β-gal, p21, ROS ↑ collagen fiber content ↑ pAMPK ↓ MMP-1, p21 ↓ epidermal layer thickness | [91] |
| COUMARINS | ||||
Hydrangenol | HR-1 hairless mice (5, 10, 20, 40 mg/kg), oral administration | UVB (60–240 mJ/cm2, stepwise increased irradiation) | ↑ COL1A1, procollagen type I ↑ Nrf2, HO-1, NQO-1, GCLM, GCLC ↓ MMP-1, MMP-3, COX-2, IL-6, pSTAT1 ↓ pp38, pERK, pc-Fos, pc-Jun ↓ wrinkle formation, dermis thickness | [92] |
Esculetin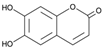 | HDFs (10 and 100 µg/mL) | ↑ COL1A1, procollagen type I ↑ pERK, pp38, pJNK ↑ pAkt ↑ Sp1 | [93] | |
| ANTHRAQUINONES | ||||
Emodin | HDFs Hs27 (0.01–1 μM) | None | ↑ COL1A1, COL1A2, collagen type I ↑ pAMPK, pERK1/2 (-) COL3A1, COL5A1 (-) pFAK, pp38, pSMAD2 | [94] |
| OTHERS (NON-POLYPHENOLIC COMPOUNDS) | ||||
Ginsenoside Rb1 Ginsenoside Rd  | HDFs (50 and 100 μM) | TNF-α (20 μg/mL) | ↑ collagen type I in medium (-) MMP-1 | [95] |
Santamarine | HDFs (1, 5, 10 μM) | UVA (8 J/cm2) H2O2 (500 µM) | ↑ COL1A1, procollagen type I ↑ TGF-β, pSmad2/3, Smad4, pERK, ↑ Nrf2, SOD-1, HO-1, Nrf2, SOD-1, HO-1 ↓ MMP-1, MMP-1, MMP-3, MMP-9 ↓ pp38, pJNK, pc-Fos, pc-Jun, Smad7 ↓ ROS | [96] |
Trans-Cinnamic acid  | HDF Hs68 (20, 60, 100 μM) HDF Hs68 (20, 60, 100 μM) BALB/c-nu mice (20 and 100 mM) | UVA (3 J/cm2) None UVA (3 J/cm2) | ↑ procollagen type I ↓ MMP-1, MMP-3, ROS, pc-Fos ↑ Nrf2, HO-1 and γ-GCLC ↑ procollagen type I ↓ MMP-1, MMP-3 | [97] |
Β-lapachone | HDFs (0.01–0.1 μg/mL) | None | ↑ COL1A1, procollagen type I C-peptide ↑ collagen type I ↑ p-Smad2/3 | [98] |
Anethole | HDFs (1 μM) | None | ↑ COL1A1, total collagen ↓ MMP-2 | [99] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żynda, W.; Ruczaj, A.; Galicka, A. Natural Compounds with Beneficial Effects on Skin Collagen Type I and Mechanisms of Their Action. Antioxidants 2025, 14, 389. https://doi.org/10.3390/antiox14040389
Żynda W, Ruczaj A, Galicka A. Natural Compounds with Beneficial Effects on Skin Collagen Type I and Mechanisms of Their Action. Antioxidants. 2025; 14(4):389. https://doi.org/10.3390/antiox14040389
Chicago/Turabian StyleŻynda, Wioleta, Agnieszka Ruczaj, and Anna Galicka. 2025. "Natural Compounds with Beneficial Effects on Skin Collagen Type I and Mechanisms of Their Action" Antioxidants 14, no. 4: 389. https://doi.org/10.3390/antiox14040389
APA StyleŻynda, W., Ruczaj, A., & Galicka, A. (2025). Natural Compounds with Beneficial Effects on Skin Collagen Type I and Mechanisms of Their Action. Antioxidants, 14(4), 389. https://doi.org/10.3390/antiox14040389