
Myrobalan Fruit Extracts Modulate Immunobiochemical Pathways In Vitro


 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Ethanol Extracts
2.2. Cell Culture
2.3. Oxygen Radical Antioxidant Capacity (ORAC) Assay
2.4. Measurement of NF-κB Activity in THP-1-BlueTM Cells
2.5. Measurements of Tryptophan (Trp), Kynurenine (Kyn), and Neopterin
2.6. Nrf-2/ARE Reporter Gene Assay
2.7. Measurement of Intracellular ROS
2.8. Cell Viability Assay
2.9. Statistical Analysis
2.10. RNA Extraction
2.11. Reverse Transcription Quantitative PCR (RT-qPCR)
2.12. Transcriptomics
2.13. Functional Analysis
3. Results
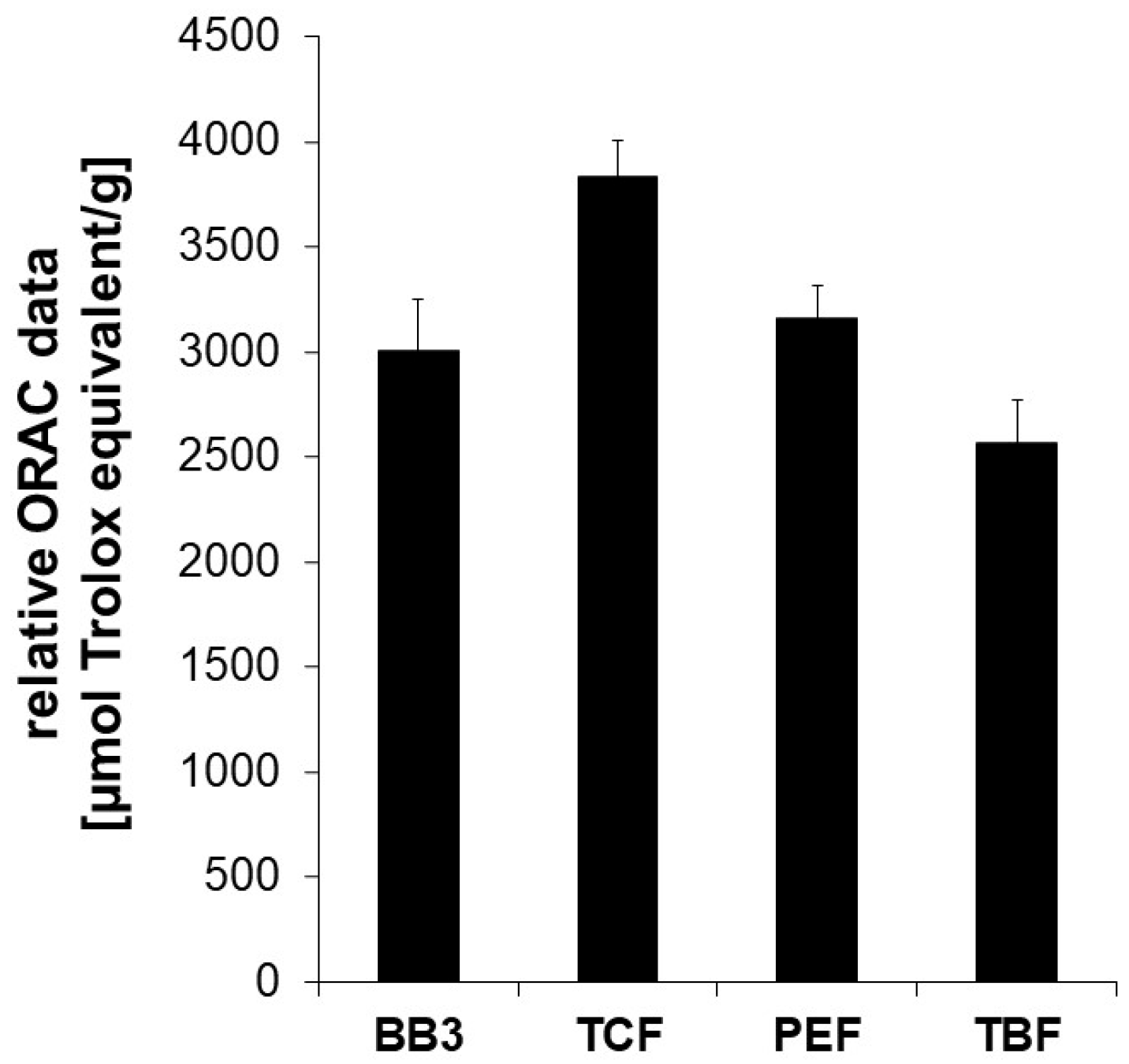
3.1. Antioxidant Capacity
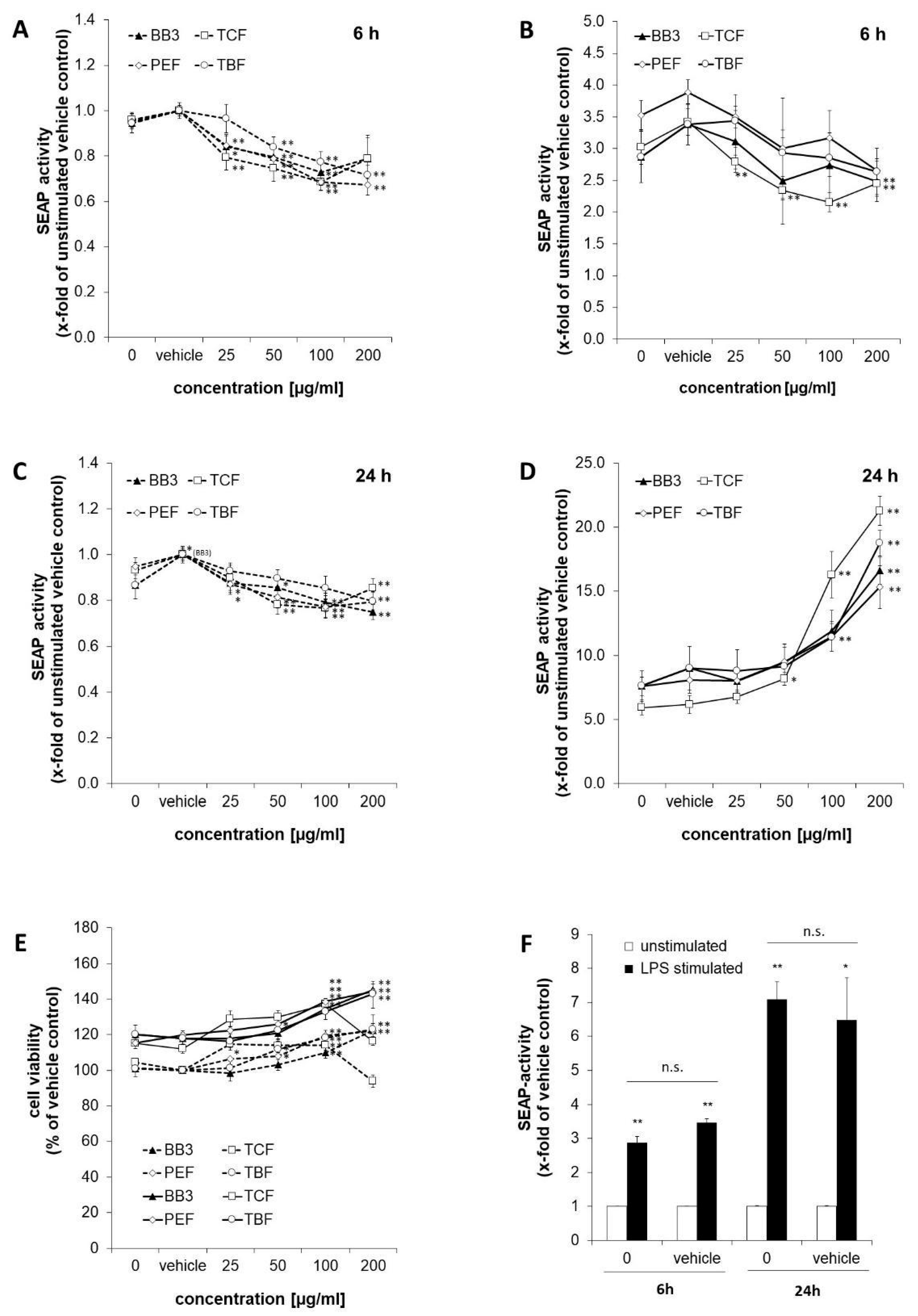
3.2. Effects of BB3 and Constituent Extracts on NF-κB Activity
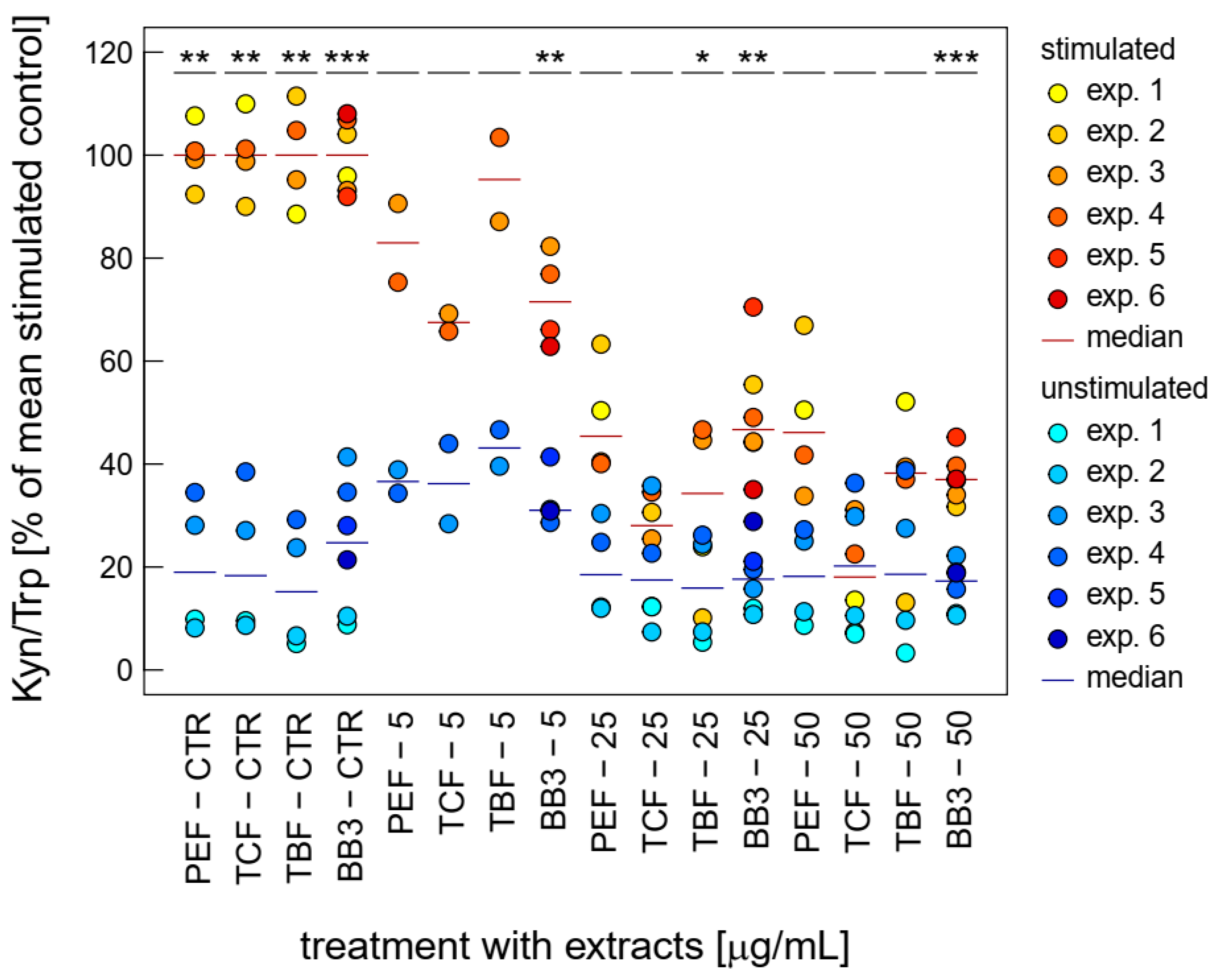
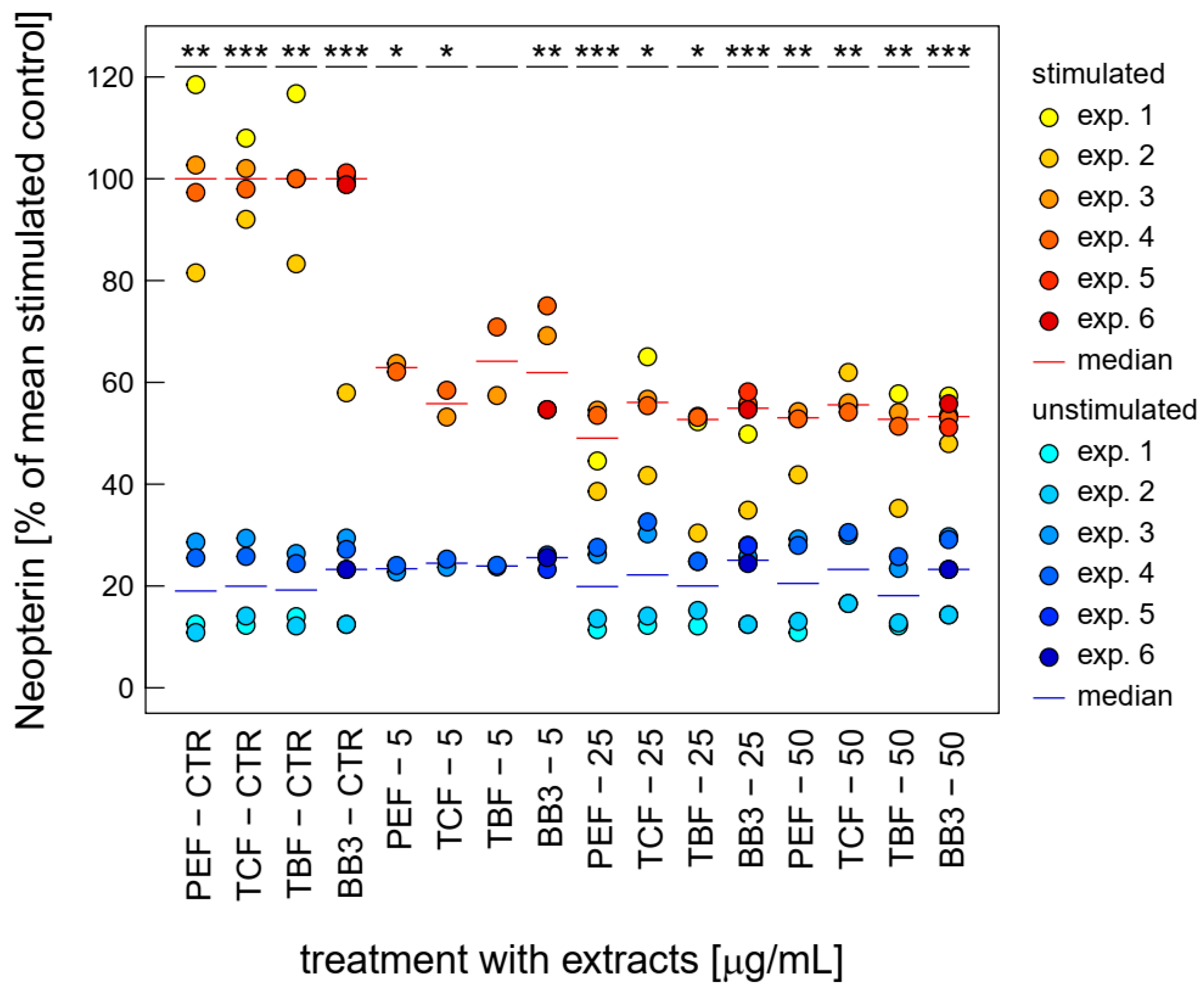
3.3. Effects of BB3 and Constituent Extracts on Tryptophan and Neopterin Metabolism
3.4. Viability of HepG2 Cells
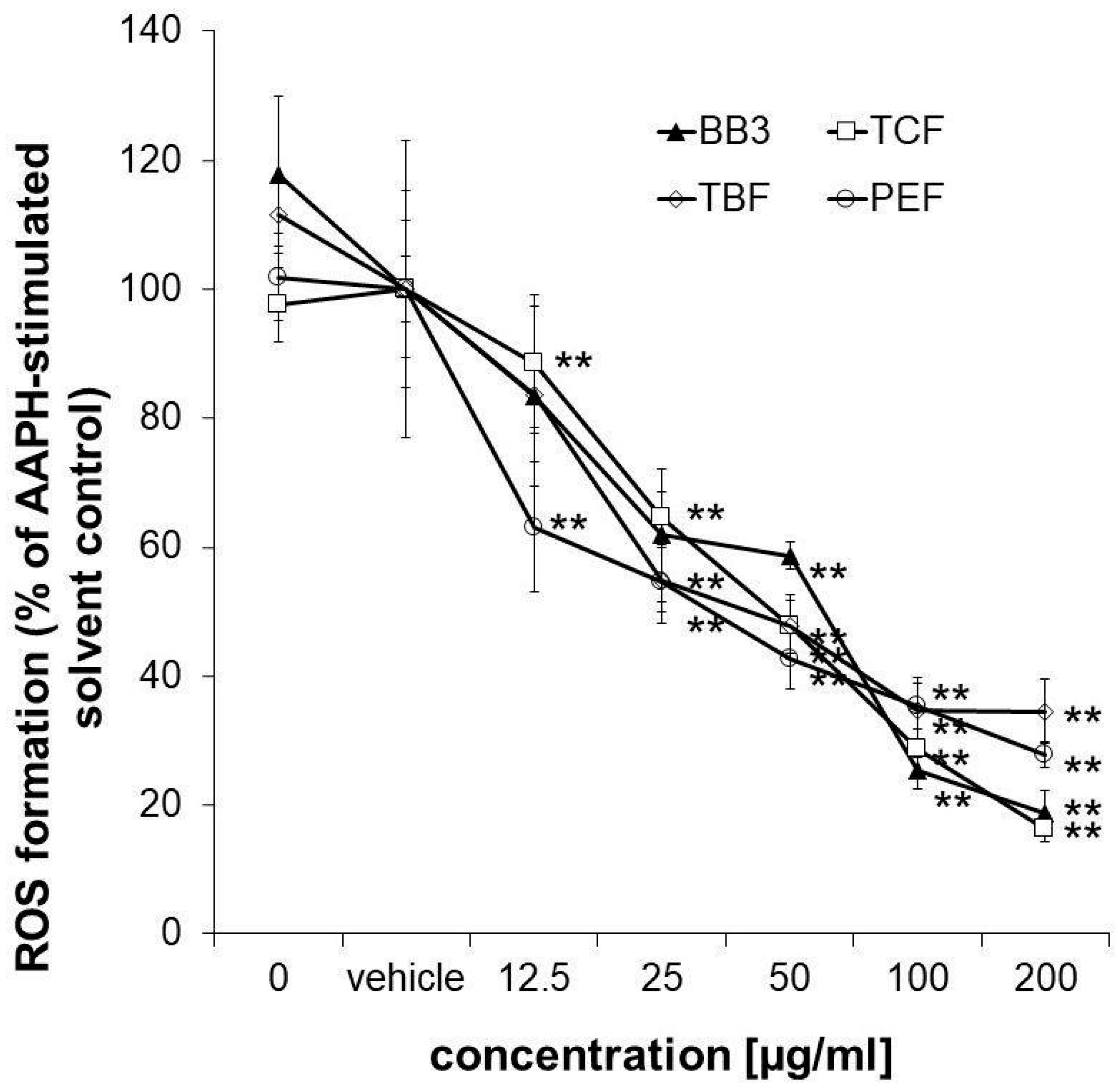
3.5. Cellular Antioxidant Activity in HepG2 Cells
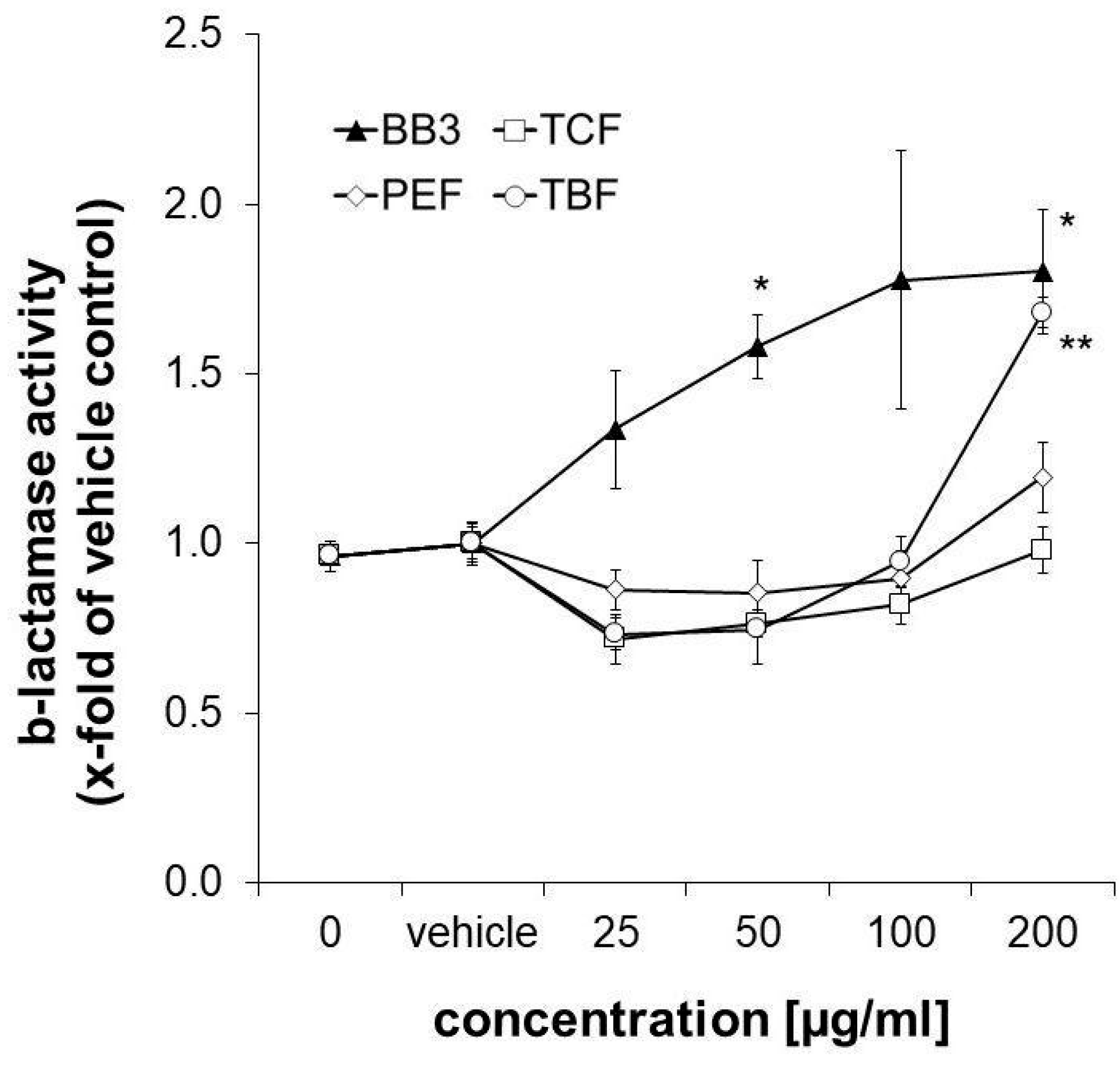
3.6. Synergistic Induction of ARE-Dependent Gene Expression
3.7. Gene Expression Changes
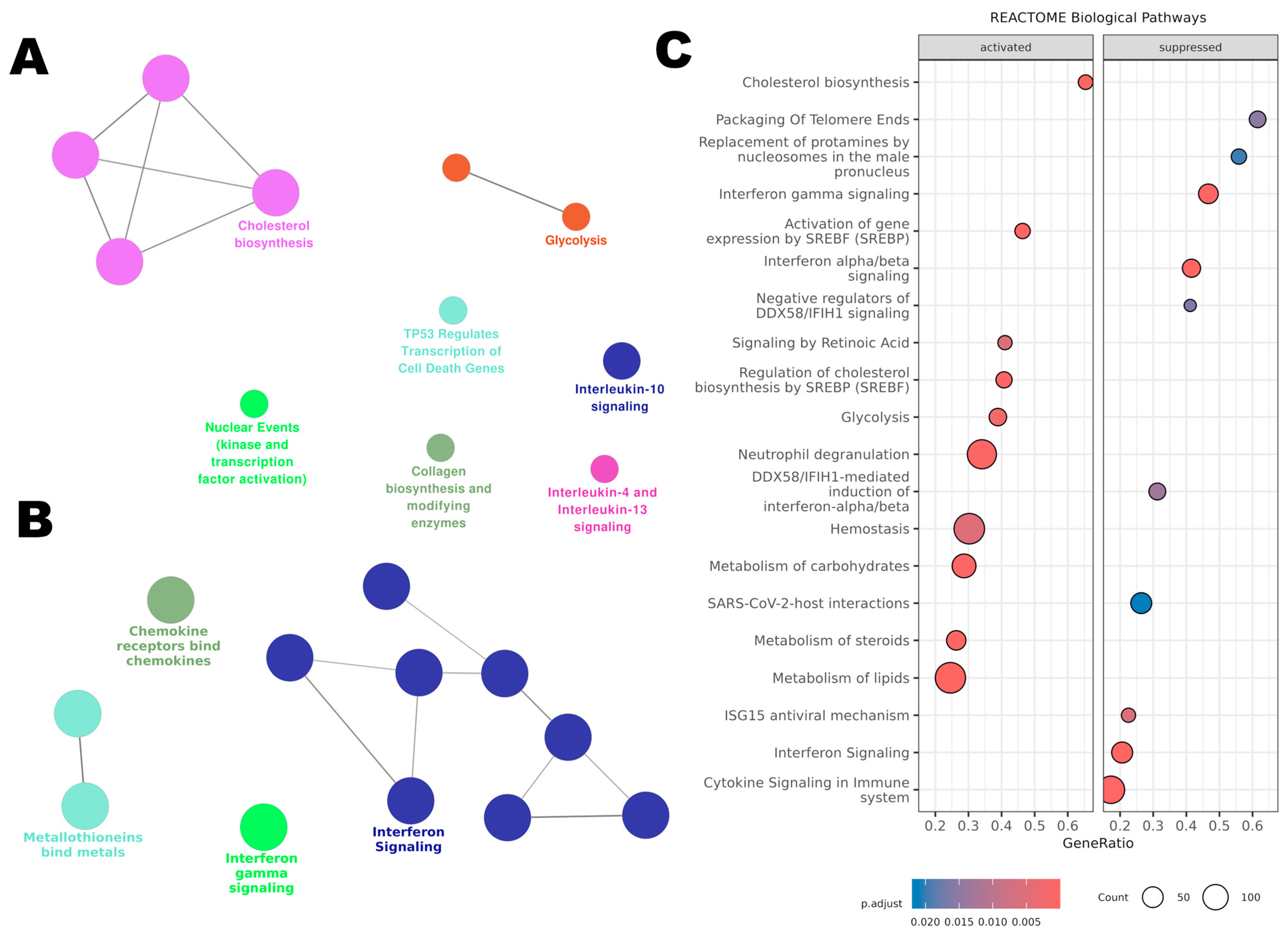
3.7.1. Pathway Enrichment Analysis with ClueGO
3.7.2. Validation of Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gostner, J.M.; Wrulich, O.A.; Jenny, M.; Fuchs, D.; Ueberall, F. An Update on the Strategies in Multicomponent Activity Monitoring within the Phytopharmaceutical Field. BMC Complement. Altern. Med. 2012, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolutionary Advantage and Molecular Modes of Action of Multi-Component Mixtures Used in Phytomedicine. Curr. Drug Metab. 2008, 9, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Talalay, P. Direct and Indirect Antioxidant Properties of Inducers of Cytoprotective Proteins. Mol. Nutr. Food Res. 2008, 52 (Suppl. S1), S128–S138. [Google Scholar] [CrossRef]
- Klein, A.; Wrulich, O.A.; Jenny, M.; Gruber, P.; Becker, K.; Fuchs, D.; Gostner, J.M.; Uberall, F. Pathway-Focused Bioassays and Transcriptome Analysis Contribute to a Better Activity Monitoring of Complex Herbal Remedies. BMC Genom. 2013, 14, 133. [Google Scholar] [CrossRef]
- Ulrich-Merzenich, G.; Panek, D.; Zeitler, H.; Wagner, H.; Vetter, H. New Perspectives for Synergy Research with the “Omic”-Technologies. Phytomed. Int. J. Phytother. Phytopharm. 2009, 16, 495–508. [Google Scholar] [CrossRef]
- Singh, R.; Singh, B.; Kumarb, N.; Arora, S. Antioxidant Activity of Triphala a Combination of Terminalia chebula, Terminalia bellerica and Emblica officinalis. J. Food Biochem. 2010, 34, 222–232. [Google Scholar] [CrossRef]
- Peterson, C.T.; Denniston, K.; Chopra, D. Therapeutic Uses of Triphala in Ayurvedic Medicine. J. Altern. Complement. Med. 2017, 23, 607–614. [Google Scholar] [CrossRef]
- Long, X.-M.; Li, R.; Liu, H.-P.; Xia, Z.-X.; Guo, S.; Gu, J.-X.; Zhang, L.-J.; Fan, Y.; Chen, Z.-K. Chemical Fingerprint Analysis and Quality Assessment of Tibetan Medicine Triphala from Different Origins by High-Performance Liquid Chromatography. Phytochem. Anal. 2023, 34, 476–486. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. In Vitro Bioaccessibility, Human Gut Microbiota Metabolites and Hepatoprotective Potential of Chebulic Ellagitannins: A Case of Padma Hepaten® Formulation. Nutrients 2015, 7, 8456–8477. [Google Scholar] [CrossRef]
- Tarasiuk, A.; Mosińska, P.; Fichna, J. Triphala: Current Applications and New Perspectives on the Treatment of Functional Gastrointestinal Disorders. Chin. Med. 2018, 13, 39. [Google Scholar] [CrossRef]
- Naik, G.H.; Priyadarsini, K.I.; Bhagirathi, R.G.; Mishra, B.; Mishra, K.P.; Banavalikar, M.M.; Mohan, H. In Vitro Antioxidant Studies and Free Radical Reactions of Triphala, an Ayurvedic Formulation and Its Constituents. Phytother. Res. 2005, 19, 582–586. [Google Scholar] [CrossRef]
- Kaur, S.; Michael, H.; Arora, S.; Härkönen, P.L.; Kumar, S. The in Vitro Cytotoxic and Apoptotic Activity of Triphala—An Indian Herbal Drug. J. Ethnopharmacol. 2005, 97, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Gostner, J.M.; Becker, K.; Fuchs, D.; Sucher, R. Redox Regulation of the Immune Response. Redox Rep. Commun. Free Radic. Res. 2013, 18, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.R.; Werner-Felmayer, G.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Wachter, H. Parallel Induction of Tetrahydrobiopterin Biosynthesis and Indoleamine 2,3-Dioxygenase Activity in Human Cells and Cell Lines by Interferon-Gamma. Biochem. J. 1989, 262, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Jyothi Prasanna, S.; Chandrasekar, B.; Nandi, D. Gene Modulation and Immunoregulatory Roles of Interferon Gamma. Cytokine 2010, 50, 1–14. [Google Scholar] [CrossRef]
- Hatano, E.; Bennett, B.L.; Manning, A.M.; Qian, T.; Lemasters, J.J.; Brenner, D.A. NF-kappaB Stimulates Inducible Nitric Oxide Synthase to Protect Mouse Hepatocytes from TNF-Alpha- and Fas-Mediated Apoptosis. Gastroenterology 2001, 120, 1251–1262. [Google Scholar] [CrossRef]
- Jaiswal, A.K. Nrf2 Signaling in Coordinated Activation of Antioxidant Gene Expression. Free Radic. Biol. Med. 2004, 36, 1199–1207. [Google Scholar] [CrossRef]
- Funes, S.C.; Rios, M.; Fernández-Fierro, A.; Covián, C.; Bueno, S.M.; Riedel, C.A.; Mackern-Oberti, J.P.; Kalergis, A.M. Naturally Derived Heme-Oxygenase 1 Inducers and Their Therapeutic Application to Immune-Mediated Diseases. Front. Immunol. 2020, 11, 1467. [Google Scholar] [CrossRef]
- Kapturczak, M.H.; Wasserfall, C.; Brusko, T.; Campbell-Thompson, M.; Ellis, T.M.; Atkinson, M.A.; Agarwal, A. Heme Oxygenase-1 Modulates Early Inflammatory Responses: Evidence from the Heme Oxygenase-1-Deficient Mouse. Am. J. Pathol. 2004, 165, 1045–1053. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Bianchi, R.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T Cell Apoptosis by Tryptophan Catabolism. Cell Death Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef]
- Fuchs, D.; Malkovsky, M.; Reibnegger, G.; Werner, E.R.; Forni, G.; Wachter, H. Endogenous Release of Interferon-Gamma and Diminished Response of Peripheral Blood Mononuclear Cells to Antigenic Stimulation. Immunol. Lett. 1989, 23, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Feng, G.S. Relationship between Interferon-Gamma, Indoleamine 2,3-Dioxygenase, and Tryptophan Catabolism. FASEB J. 1991, 5, 2516–2522. [Google Scholar] [CrossRef]
- Schröcksnadel, K.; Wirleitner, B.; Winkler, C.; Fuchs, D. Monitoring Tryptophan Metabolism in Chronic Immune Activation. Clin. Chim. Acta Int. J. Clin. Chem. 2006, 364, 82–90. [Google Scholar] [CrossRef]
- Murr, C.; Widner, B.; Wirleitner, B.; Fuchs, D. Neopterin as a Marker for Immune System Activation. Curr. Drug Metab. 2002, 3, 175–187. [Google Scholar] [CrossRef]
- Jenny, M.; Klieber, M.; Zaknun, D.; Schroecksnadel, S.; Kurz, K.; Ledochowski, M.; Schennach, H.; Fuchs, D. In Vitro Testing for Anti-Inflammatory Properties of Compounds Employing Peripheral Blood Mononuclear Cells Freshly Isolated from Healthy Donors. Inflamm. Res. 2011, 60, 127–135. [Google Scholar] [CrossRef]
- Becker, K.; Schwaiger, S.; Waltenberger, B.; Fuchs, D.; Pezzei, C.K.; Schennach, H.; Stuppner, H.; Gostner, J.M. Immunomodulatory Effects of Diterpene Quinone Derivatives from the Roots of Horminum Pyrenaicum in Human PBMC. Oxidative Med. Cell. Longev. 2018, 2018, 2980295. [Google Scholar] [CrossRef]
- Plaskova, A.; Mlcek, J. New Insights of the Application of Water or Ethanol-Water Plant Extract Rich in Active Compounds in Food. Front. Nutr. 2023, 10, 1118761. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Jenny, M.; Santer, E.; Klein, A.; Ledochowski, M.; Schennach, H.; Ueberall, F.; Fuchs, D. Cacao Extracts Suppress Tryptophan Degradation of Mitogen-Stimulated Peripheral Blood Mononuclear Cells. J. Ethnopharmacol. 2009, 122, 261–267. [Google Scholar] [CrossRef]
- Widner, B.; Werner, E.R.; Schennach, H.; Wachter, H.; Fuchs, D. Simultaneous Measurement of Serum Tryptophan and Kynurenine by HPLC. Clin. Chem. 1997, 43, 2424–2426. [Google Scholar] [CrossRef]
- Geisler, S.; Mayersbach, P.; Becker, K.; Schennach, H.; Fuchs, D.; Gostner, J.M. Serum Tryptophan, Kynurenine, Phenylalanine, Tyrosine and Neopterin Concentrations in 100 Healthy Blood Donors. Pteridines 2015, 26, 31–36. [Google Scholar] [CrossRef]
- Zlokarnik, G.; Negulescu, P.A.; Knapp, T.E.; Mere, L.; Burres, N.; Feng, L.; Whitney, M.; Roemer, K.; Tsien, R.Y. Quantitation of Transcription and Clonal Selection of Single Living Cells with β-Lactamase as Reporter. Science 1998, 279, 84–88. [Google Scholar] [CrossRef]
- Bass, D.A.; Parce, J.W.; Dechatelet, L.R.; Szejda, P.; Seeds, M.C.; Thomas, M. Flow Cytometric Studies of Oxidative Product Formation by Neutrophils: A Graded Response to Membrane Stimulation. J. Immunol. 1983, 130, 1910–1917. [Google Scholar] [CrossRef]
- Wang, H.; Joseph, J.A. Quantifying Cellular Oxidative Stress by Dichlorofluorescein Assay Using Microplate Reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Yu, G.; He, Q.-Y. ReactomePA: An R/Bioconductor Package for Reactome Pathway Analysis and Visualization. Mol. Biosyst. 2016, 12, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape Plug-in to Decipher Functionally Grouped Gene Ontology and Pathway Annotation Networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Schroecksnadel, S.; Jenny, M.; Fuchs, D. Myelomonocytic THP-1 Cells for In Vitro Testing of Immunomodulatory Properties of Nanoparticles. J. Biomed. Nanotechnol. 2011, 7, 209–210. [Google Scholar] [CrossRef] [PubMed]
- Schroecksnadel, S.; Jenny, M.; Kurz, K.; Klein, A.; Ledochowski, M.; Überall, F.; Fuchs, D. LPS-Induced NF-κB Expression in THP-1Blue Cells Correlates with Neopterin Production and Activity of Indoleamine 2,3-Dioxygenase. Biochem. Biophys. Res. Commun. 2010, 399, 642–646. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2-an update. Free Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef]
- Kalaiselvan, S.; Rasool, M.K. Triphala Herbal Extract Suppresses Inflammatory Responses in LPS-Stimulated RAW 264.7 Macrophages and Adjuvant-Induced Arthritic Rats via Inhibition of NF-κB Pathway. J. Immunotoxicol. 2016, 13, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, Y.; Schreiber, R.D.; Hamilton, T.A. Synergy between Interferon-Gamma and Tumor Necrosis Factor-Alpha in Transcriptional Activation Is Mediated by Cooperation between Signal Transducer and Activator of Transcription 1 and Nuclear Factor kappaB. J. Biol. Chem. 1997, 272, 14899–14907. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.J.; Lolis, E. Structure, Function, and Inhibition of Chemokines. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 469–499. [Google Scholar] [CrossRef]
- Tokunaga, R.; Zhang, W.; Naseem, M.; Puccini, A.; Berger, M.D.; Soni, S.; McSkane, M.; Baba, H.; Lenz, H.-J. CXCL9, CXCL10, CXCL11/CXCR3 Axis for Immune Activation—A Target for Novel Cancer Therapy. Cancer Treat. Rev. 2018, 63, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Saccani, A.; Saccani, S.; Orlando, S.; Sironi, M.; Bernasconi, S.; Ghezzi, P.; Mantovani, A.; Sica, A. Redox Regulation of Chemokine Receptor Expression. Proc. Natl. Acad. Sci. USA 2000, 97, 2761–2766. [Google Scholar] [CrossRef]
- Otsuji, M.; Kimura, Y.; Aoe, T.; Okamoto, Y.; Saito, T. Oxidative Stress by Tumor-Derived Macrophages Suppresses the Expression of CD3 Zeta Chain of T-Cell Receptor Complex and Antigen-Specific T-Cell Responses. Proc. Natl. Acad. Sci. USA 1996, 93, 13119–13124. [Google Scholar] [CrossRef]
- Thiel, G.; Cibelli, G. Regulation of Life and Death by the Zinc Finger Transcription Factor Egr-1. J. Cell. Physiol. 2002, 193, 287–292. [Google Scholar] [CrossRef]
- Suehiro, J.; Hamakubo, T.; Kodama, T.; Aird, W.C.; Minami, T. Vascular Endothelial Growth Factor Activation of Endothelial Cells Is Mediated by Early Growth Response-3. Blood 2010, 115, 2520–2532. [Google Scholar] [CrossRef]
- Dong, H.; Hao, L.; Zhang, W.; Zhong, W.; Guo, W.; Yue, R.; Sun, X.; Zhou, Z. Activation of AhR-NQO1 Signaling Pathway Protects Against Alcohol-Induced Liver Injury by Improving Redox Balance. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 793–811. [Google Scholar] [CrossRef]
- Asher, G.; Lotem, J.; Kama, R.; Sachs, L.; Shaul, Y. NQO1 Stabilizes P53 through a Distinct Pathway. Proc. Natl. Acad. Sci. USA 2002, 99, 3099–3104. [Google Scholar] [CrossRef]
- Terashima, J.; Habano, W.; Gamou, T.; Ozawa, S. Induction of CYP1 Family Members under Low-Glucose Conditions Requires AhR Expression and Occurs through the Nuclear Translocation of AhR. Drug Metab. Pharmacokinet. 2011, 26, 577–583. [Google Scholar] [CrossRef]
- Opitz, C.A.; Litzenburger, U.M.; Sahm, F.; Ott, M.; Tritschler, I.; Trump, S.; Schumacher, T.; Jestaedt, L.; Schrenk, D.; Weller, M.; et al. An Endogenous Tumour-Promoting Ligand of the Human Aryl Hydrocarbon Receptor. Nature 2011, 478, 197–203. [Google Scholar] [CrossRef] [PubMed]
- DiNatale, B.C.; Murray, I.A.; Schroeder, J.C.; Flaveny, C.A.; Lahoti, T.S.; Laurenzana, E.M.; Omiecinski, C.J.; Perdew, G.H. Kynurenic Acid Is a Potent Endogenous Aryl Hydrocarbon Receptor Ligand That Synergistically Induces Interleukin-6 in the Presence of Inflammatory Signaling. Toxicol. Sci. 2010, 115, 89–97. [Google Scholar] [CrossRef]
- Zhou, C.; Li, J.; Du, J.; Jiang, X.; Xu, X.; Liu, Y.; He, Q.; Liang, H.; Fang, P.; Zhan, H.; et al. HMGCS1 Drives Drug-Resistance in Acute Myeloid Leukemia through Endoplasmic Reticulum-UPR-Mitochondria Axis. Biomed. Pharmacother. 2021, 137, 111378. [Google Scholar] [CrossRef] [PubMed]
- Ghallab, A. In Vitro Test Systems and Their Limitations. EXCLI J. 2013, 12, 1024–1026. [Google Scholar]
- Arzumanian, V.A.; Kiseleva, O.I.; Poverennaya, E.V. The Curious Case of the HepG2 Cell Line: 40 Years of Expertise. Int. J. Mol. Sci. 2021, 22, 13135. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome P450 Research and The Journal of Biological Chemistry. J. Biol. Chem. 2019, 294, 1671–1680. [Google Scholar] [CrossRef]
- Pareek, A.; Godavarthi, A.; Issarani, R.; Nagori, B.P. Antioxidant and Hepatoprotective Activity of Fagonia schweinfurthii (Hadidi) Hadidi Extract in Carbon Tetrachloride Induced Hepatotoxicity in HepG2 Cell Line and Rats. J. Ethnopharmacol. 2013, 150, 973–981. [Google Scholar] [CrossRef]
- Baxter, E.W.; Graham, A.E.; Re, N.A.; Carr, I.M.; Robinson, J.I.; Mackie, S.L.; Morgan, A.W. Standardized Protocols for Differentiation of THP-1 Cells to Macrophages with Distinct M(IFNγ+LPS), M(IL-4) and M(IL-10) Phenotypes. J. Immunol. Methods 2020, 478, 112721. [Google Scholar] [CrossRef]
- Tice, R.R.; Austin, C.P.; Kavlock, R.J.; Bucher, J.R. Improving the Human Hazard Characterization of Chemicals: A Tox21 Update. Environ. Health Perspect. 2013, 121, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Bialonska, D.; Kasimsetty, S.G.; Khan, S.I.; Ferreira, D. Urolithins, Intestinal Microbial Metabolites of Pomegranate Ellagitannins, Exhibit Potent Antioxidant Activity in a Cell-Based Assay. J. Agric. Food Chem. 2009, 57, 10181–10186. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yan, C.C.; Zhang, X.; Zhang, X.; Dai, F.; Yin, J.; Zhang, Y. Drug-Target Interaction Prediction: Databases, Web Servers and Computational Models. Brief. Bioinform. 2016, 17, 696–712. [Google Scholar] [CrossRef] [PubMed]
- Cidem, A.; Bradbury, P.; Traini, D.; Ong, H.X. Modifying and Integrating in Vitro and Ex Vivo Respiratory Models for Inhalation Drug Screening. Front. Bioeng. Biotechnol. 2020, 8, 581995. [Google Scholar] [CrossRef]
- Kuenzi, B.M.; Park, J.; Fong, S.H.; Sanchez, K.S.; Lee, J.; Kreisberg, J.F.; Ma, J.; Ideker, T. Predicting Drug Response and Synergy Using a Deep Learning Model of Human Cancer Cells. Cancer Cell 2020, 38, 672–684.e6. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microarray (n = 1) | RT-qPCR (n = 4) | |||
|---|---|---|---|---|
| Gene | Gene Name | Log2FC | FC | p-Value |
| IDO1 | Indoleamine 2,3-dioxygenase 1 | −0.24 | 0.763 | 0.562 |
| CXCL11 | C-X-C motif chemokine ligand 11 | −1.71 | 0.233 | 0.001 |
| SQLE | Squalene epoxidase | 0.61 | 1.581 | 0.011 |
| NFKB1 | Nuclear factor kappa B subunit 1 | 0.05 | 0.935 | 0.858 |
| HMGCR | 3-hydroxy-3-methylglutaryl-CoA reductase | 0.63 | 1.377 | 0.478 |
| STAT1 | Signal transducer and activator of transcription 1 | −0.56 | 0.507 | 0.002 |
| GCH | GTP cyclohydrolase 1 | −0.23 | 0.738 | 0.087 |
| TNF | Tumor necrosis factor | −0.35 | 0.956 | 0.894 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofer, S.; Jenny, M.; Klein, A.; Becker, K.; Parráková, L.; Überall, F.; Ganzera, M.; Fuchs, D.; Hackl, H.; Monfort-Lanzas, P.; et al. Myrobalan Fruit Extracts Modulate Immunobiochemical Pathways In Vitro. Antioxidants 2025, 14, 350. https://doi.org/10.3390/antiox14030350
Hofer S, Jenny M, Klein A, Becker K, Parráková L, Überall F, Ganzera M, Fuchs D, Hackl H, Monfort-Lanzas P, et al. Myrobalan Fruit Extracts Modulate Immunobiochemical Pathways In Vitro. Antioxidants. 2025; 14(3):350. https://doi.org/10.3390/antiox14030350
Chicago/Turabian StyleHofer, Stefanie, Marcel Jenny, Angela Klein, Kathrin Becker, Lucia Parráková, Florian Überall, Markus Ganzera, Dietmar Fuchs, Hubert Hackl, Pablo Monfort-Lanzas, and et al. 2025. "Myrobalan Fruit Extracts Modulate Immunobiochemical Pathways In Vitro" Antioxidants 14, no. 3: 350. https://doi.org/10.3390/antiox14030350
APA StyleHofer, S., Jenny, M., Klein, A., Becker, K., Parráková, L., Überall, F., Ganzera, M., Fuchs, D., Hackl, H., Monfort-Lanzas, P., & Gostner, J. M. (2025). Myrobalan Fruit Extracts Modulate Immunobiochemical Pathways In Vitro. Antioxidants, 14(3), 350. https://doi.org/10.3390/antiox14030350