
Oral Administration of Lactobacillus gasseri and Lacticaseibacillus rhamnosus Ameliorates Amyloid Beta (Aβ)-Induced Cognitive Impairment by Improving Synaptic Function Through Regulation of TLR4/Akt Pathway

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Probiotics
2.2. Animal Design
2.3. Behavioral Test
2.3.1. Y-Maze Test
2.3.2. Passive Avoidance Test
2.3.3. Morris Water Maze (MWM) Test
2.4. Antioxidant System
2.5. Mitochondrial Function
2.6. Cholinergic System
2.7. Western Blot
2.8. 16S rRNA Sequencing
2.9. Statistical Analysis
3. Results
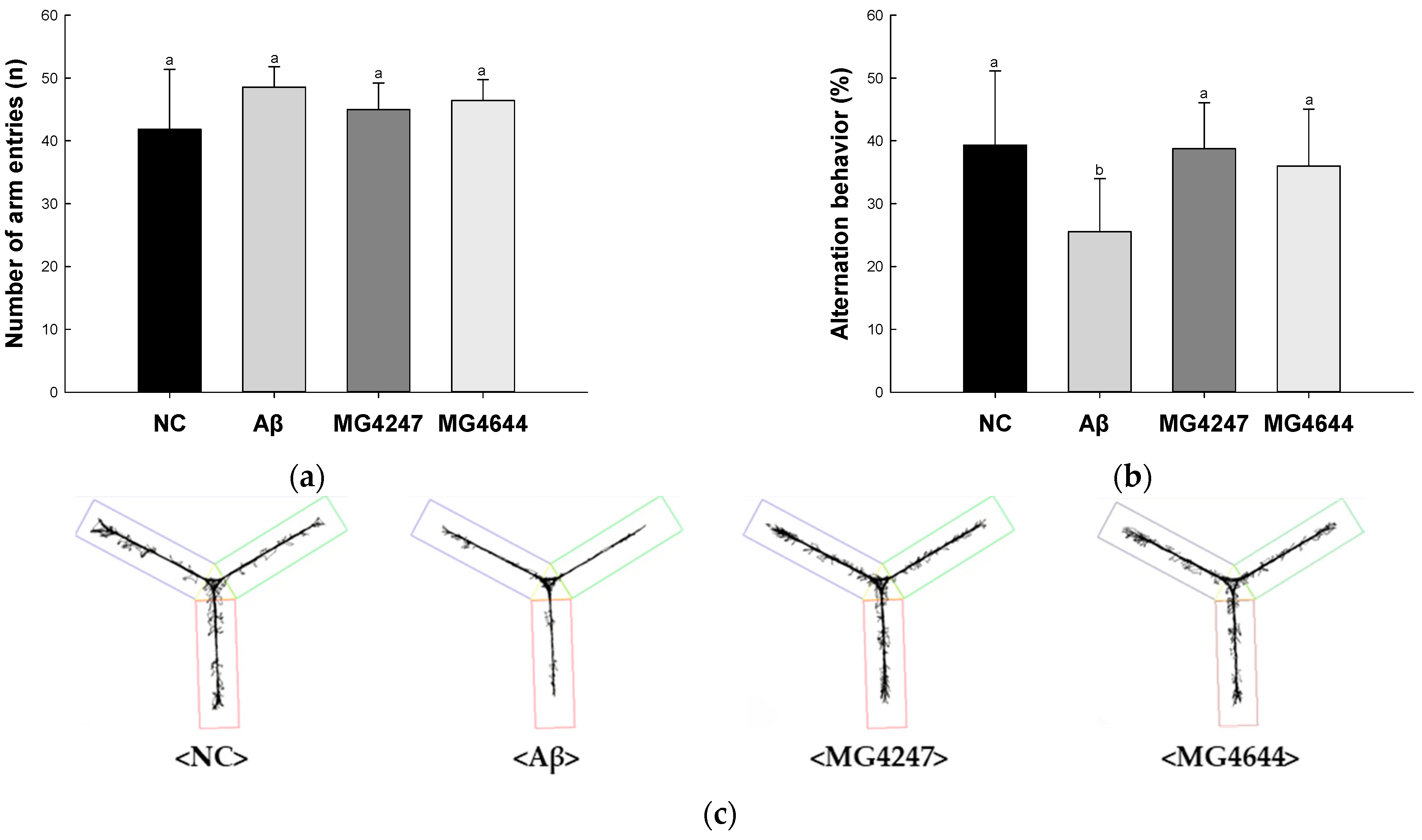
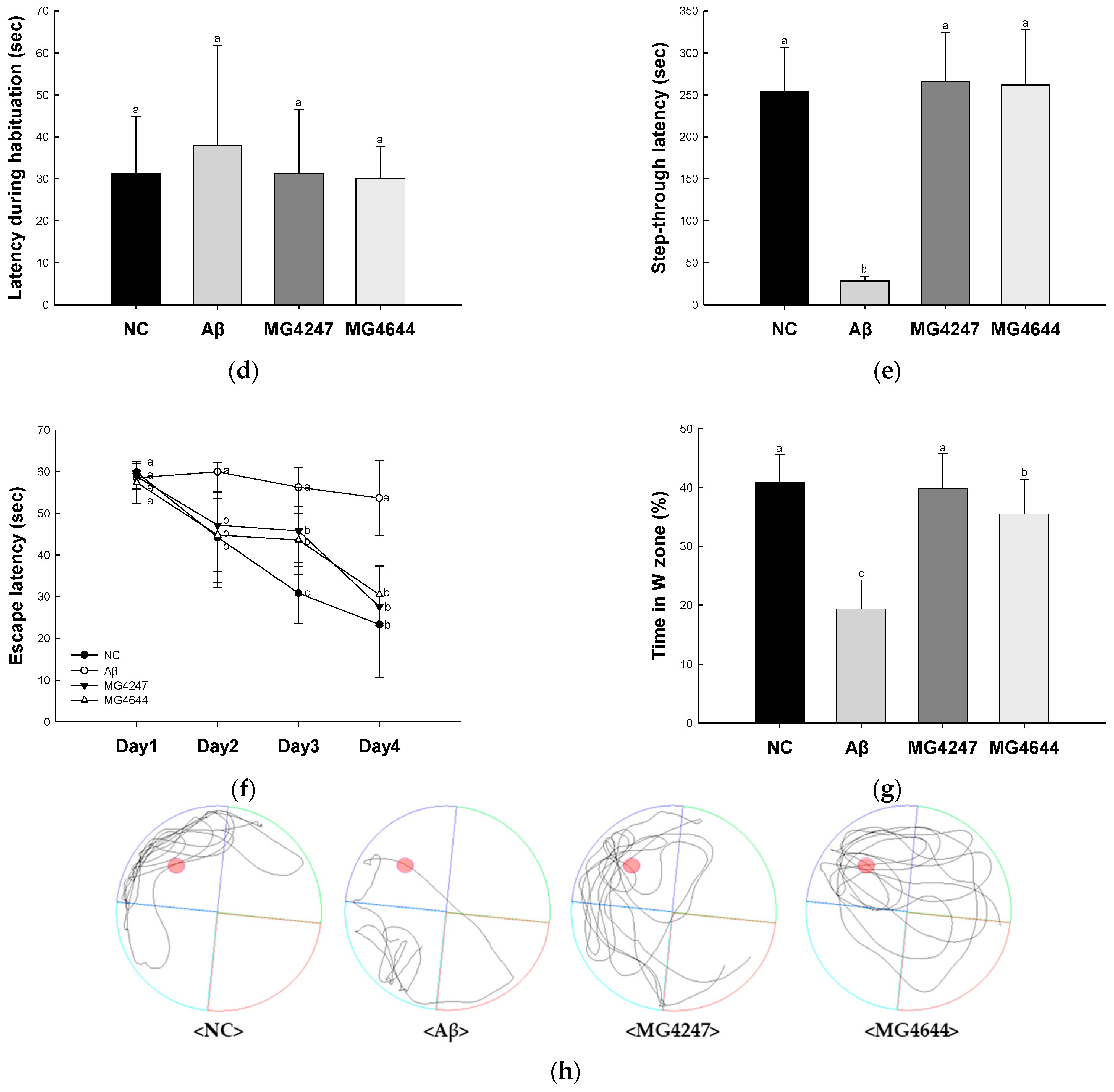
3.1. L. gasseri MG4247 and L. rhamnosus MG4644 Ameliorate Memory and Learning Ability
3.2. L. gasseri MG4247 and L. rhamnosus MG4644 Enhance Antioxidant Activity
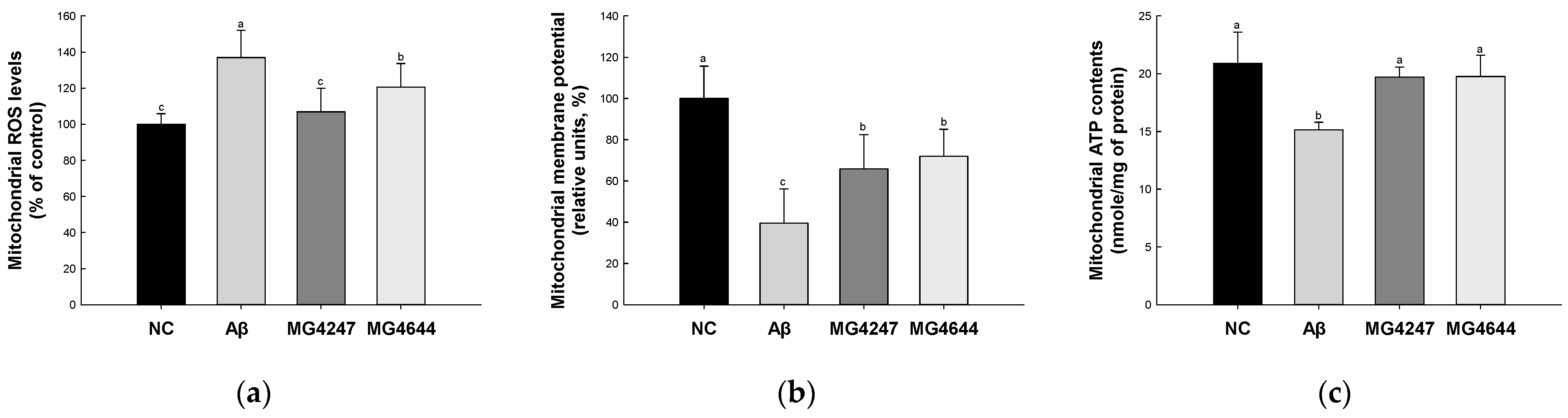
3.3. L. gasseri MG4247 and L. rhamnosus MG4644 Improve Mitochondrial Function
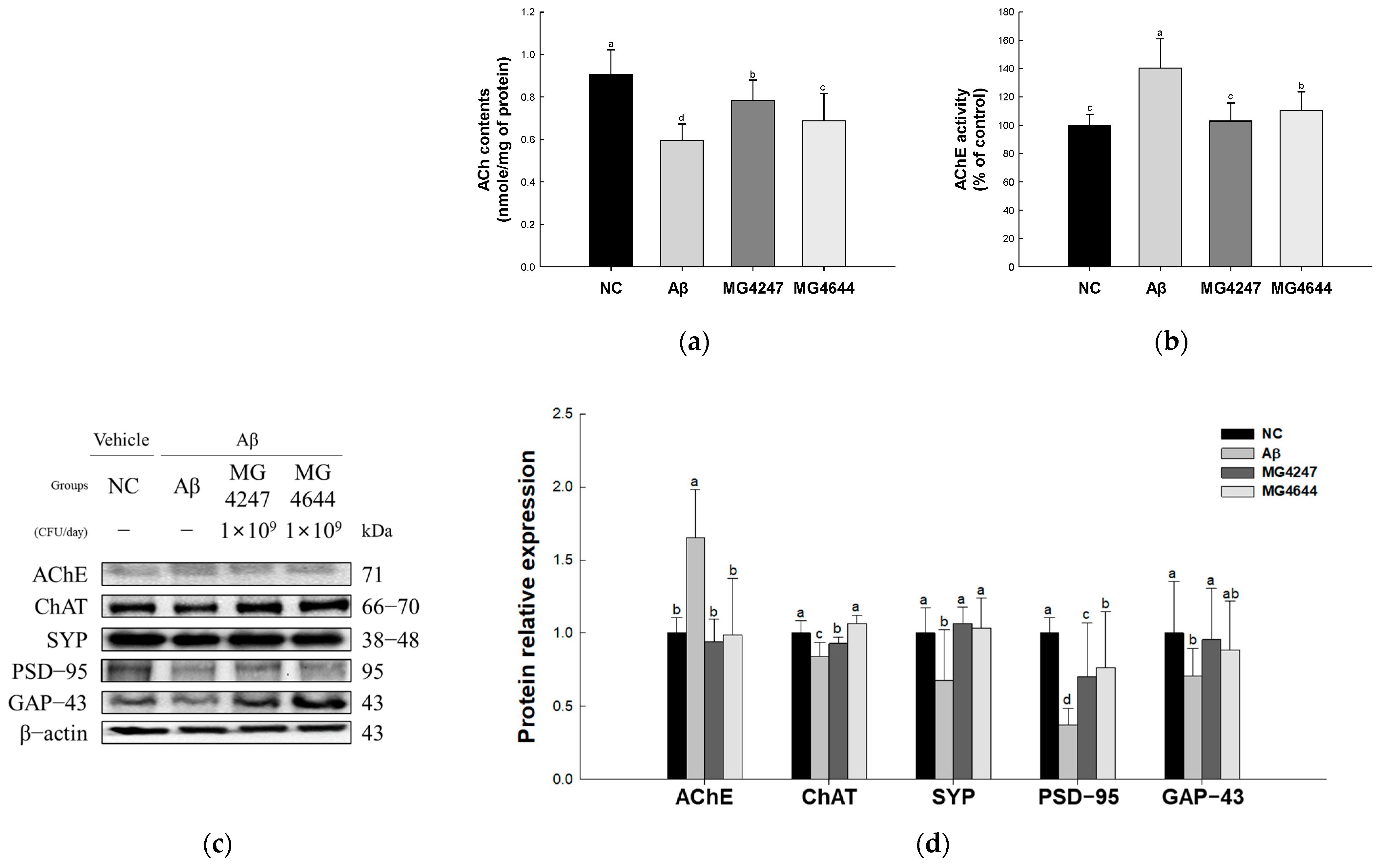
3.4. L. gasseri MG4247 and L. rhamnosus MG4644 Enhance Synaptic Plasticity
3.5. L. gasseri MG4247 and L. rhamnosus MG4644 Strengthen the BBB by Modulating Expression Levels of Tight Junction (TJ) Proteins
3.6. L. gasseri MG4247 and L. rhamnosus MG4644 Suppress Neuroinflammation Through Inhibition of the TLR4 Pathway
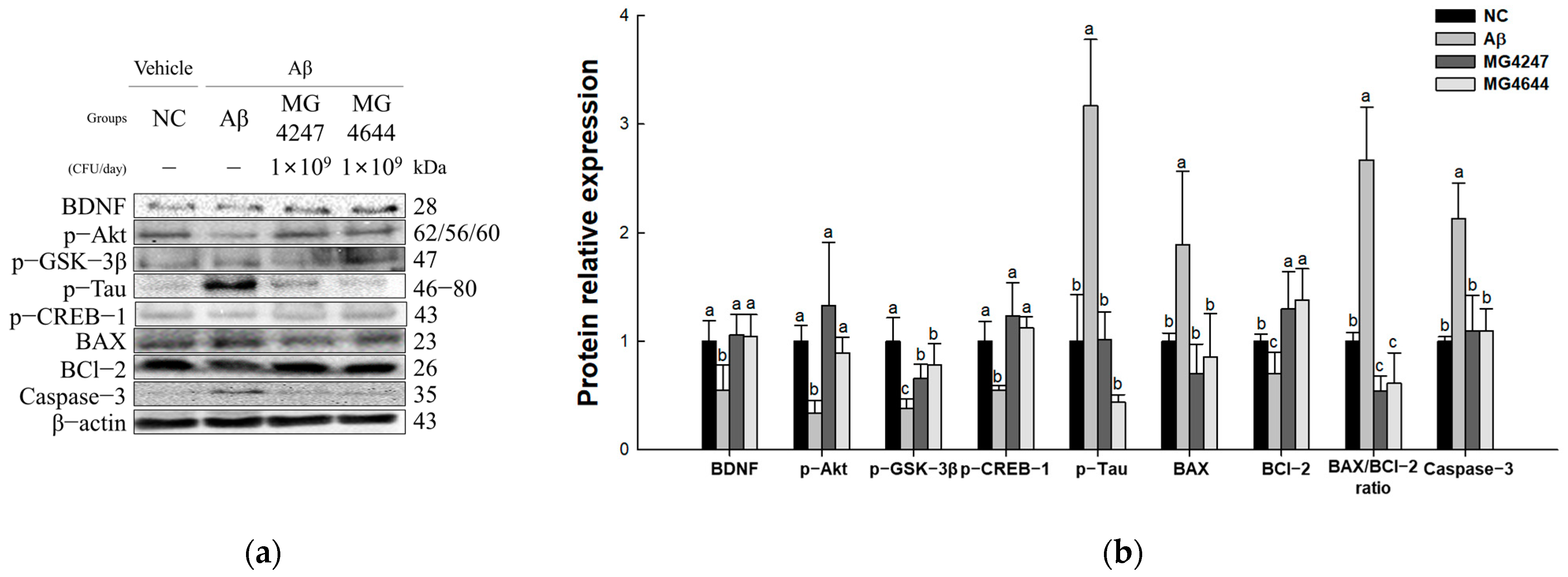
3.7. L. gasseri MG4247 and L. rhamnosus MG4644 Attenuate Neurotoxicity Through Regulation of the Akt Pathway
3.8. L. gasseri MG4247 and L. rhamnosus MG4644 Improve Dysbiosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mimura, Y.; Nishida, H.; Nakajima, S.; Tsugawa, S.; Morita, S.; Yoshida, K.; Tarumi, R.; Ogyu, K.; Wada, M.; Kurose, S. Neurophysiological biomarkers using transcranial magnetic stimulation in Alzheimer’s disease and mild cognitive impairment: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2021, 121, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. Probiotics for mild cognitive impairment and Alzheimer’s disease: A systematic review and meta-analysis. Foods 2021, 10, 1672. [Google Scholar] [CrossRef]
- Tahami Monfared, A.A.; Byrnes, M.J.; White, L.A.; Zhang, Q. Alzheimer’s disease: Epidemiology and clinical progression. Neurol. Ther. 2022, 11, 553–569. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Li, R.; Sterling, K.; Song, W. Amyloid β-based therapy for Alzheimer’s disease: Challenges, successes and future. Signal. Transduct. Target. Ther. 2023, 8, 248. [Google Scholar] [CrossRef]
- John, A.; Reddy, P.H. Synaptic basis of Alzheimer’s disease: Focus on synaptic amyloid beta, P-tau and mitochondria. Ageing Res. Rev. 2021, 65, 101208. [Google Scholar] [CrossRef]
- Chen, A.; Kang, Y.; Liu, J.; Wu, J.; Feng, X.; Wang, M.; Zhang, Y.; Wang, R.; Lai, X.; Shao, L. Improvement of synaptic plasticity by nanoparticles and the related mechanisms: Applications and prospects. J. Control. Release 2022, 347, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dai, Y.; Li, Q.; Chen, C.; Chen, H.; Song, Y.; Hua, F.; Zhang, Z. Beta-amyloid activates NLRP3 inflammasome via TLR4 in mouse microglia. Neurosci. Lett. 2020, 736, 135279. [Google Scholar] [CrossRef]
- Lin, L.; Li, C.; Zhang, Y.; Zhang, L.; Gao, L.; Jin, L.; Shu, Y.; Shen, Y. Effects of an Akt-activating peptide obtained from walnut protein degradation on the prevention of memory impairment in mice. Food. Funct. 2024, 15, 2115–2130. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, J.B.; Coradini, K.; Frozza, R.L.; Oliveira, C.M.; Meneghetti, A.B.; Bernardi, A.; Pires, E.S.; Beck, R.C.; Salbego, C.G. Free and nanoencapsulated curcumin suppress β-amyloid-induced cognitive impairments in rats: Involvement of BDNF and Akt/GSK-3β signaling pathway. Neurobiol. Learn. Mem. 2013, 106, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Chi, R.; Liu, G.; Tian, W.; Zhang, J.; Zhang, R. Aerobic exercise regulates apoptosis through the PI3K/Akt/GSK-3β signaling pathway to improve cognitive impairment in Alzheimer’s disease mice. Neural Plast. 2022, 2022, 1500710. [Google Scholar] [CrossRef]
- Ashleigh, T.; Swerdlow, R.H.; Beal, M.F. The role of mitochondrial dysfunction in Alzheimer’s disease pathogenesis. Alzheimers Dement. 2023, 19, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Sisodia, S.S.; Vassar, R.J. The gut microbiome in Alzheimer’s disease: What we know and what remains to be explored. Mol. Neurodegener. 2023, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- de JR De-Paula, V.; Forlenza, A.S.; Forlenza, O.V. Relevance of gutmicrobiota in cognition, behaviour and Alzheimer’s disease. Pharmacol. Res. 2018, 136, 29–34. [Google Scholar] [CrossRef]
- Verhaar, B.J.; Hendriksen, H.M.; de Leeuw, F.A.; Doorduijn, A.S.; van Leeuwenstijn, M.; Teunissen, C.E.; Barkhof, F.; Scheltens, P.; Kraaij, R.; van Duijn, C.M. Gut microbiota composition is related to AD pathology. Front. Immunol. 2022, 12, 794519. [Google Scholar] [CrossRef]
- Pistollato, F.; Sumalla Cano, S.; Elio, I.; Masias Vergara, M.; Giampieri, F.; Battino, M. Role of gut microbiota and nutrients in amyloid formation and pathogenesis of Alzheimer disease. Nutr. Rev. 2016, 74, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Mathipa-Mdakane, M.G.; Thantsha, M.S. Lacticaseibacillus rhamnosus: A suitable candidate for the construction of novel bioengineered probiotic strains for targeted pathogen control. Foods 2022, 11, 785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, K.; Cao, T.; Duan, Z. Characterization of a Lactobacillus gasseri strain as a probiotic for female vaginitis. Sci. Rep. 2024, 14, 14426. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.-W.; Kim, J.-K.; Lee, K.-E.; Oh, Y.J.; Choi, H.-J.; Han, M.J.; Kim, D.-H. A probiotic Lactobacillus gasseri alleviates Escherichia coli-induced cognitive impairment and depression in mice by regulating IL-1β expression and gut microbiota. Nutrients 2020, 12, 3441. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, J.-H.; Jung, Y.-R.; Kang, C.-H. Lactobacillus gasseri MG4247 and Lacticaseibacillus paracasei MG4272 and MG4577 modulate allergic inflammatory response in RAW 264.7 and RBL-2H3 cells. Probiotics Antimicrob. Proteins 2023, 15, 1092–1101. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Wu, X.; Lv, Y.-G.; Du, Y.-F.; Chen, F.; Reed, M.N.; Hu, M.; Suppiramaniam, V.; Tang, S.-S.; Hong, H. Neuroprotective effects of INT-777 against Aβ1–42-induced cognitive impairment, neuroinflammation, apoptosis, and synaptic dysfunction in mice. Brain Behav. Immun. 2018, 73, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, V.V.; Thandavarayan, R.A.; Arumugam, S.; Mizuno, M.; Nawa, H.; Suzuki, K.; Ko, K.M.; Krishnamurthy, P.; Watanabe, K.; Konishi, T. Schisandrin B ameliorates ICV-infused amyloid β induced oxidative stress and neuronal dysfunction through inhibiting RAGE/NF-κB/MAPK and up-regulating HSP/Beclin expression. PLoS ONE 2015, 10, e0142483. [Google Scholar] [CrossRef] [PubMed]
- Go, M.J.; Kim, J.M.; Kang, J.Y.; Park, S.K.; Lee, C.J.; Kim, M.J.; Lee, H.R.; Kim, T.Y.; Joo, S.G.; Kim, D.-O. Korean red pine (Pinus densiflora) bark extract attenuates Aβ-induced cognitive impairment by regulating cholinergic dysfunction and neuroinflammation. J. Microbiol. Biotechnol. 2022, 32, 1154. [Google Scholar] [CrossRef]
- Lee, H.L.; Kim, J.M.; Moon, J.H.; Kim, M.J.; Jeong, H.R.; Go, M.J.; Kim, H.-J.; Eo, H.J.; Lee, U.; Heo, H.J. Anti-amnesic effect of synbiotic supplementation containing Corni fructus and Limosilactobacillus reuteri in DSS-induced colitis mice. Int. J. Mol. Sci. 2022, 24, 90. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.B.; Baiseitova, A.; Zahoor, M.; Ahmad, I.; Ikram, M.; Bakhsh, A.; Shah, M.A.; Ali, I.; Idress, M.; Ullah, R. Probiotic significance of Lactobacillus strains: A comprehensive review on health impacts, research gaps, and future prospects. Gut Microbes 2024, 16, 2431643. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-q.; Zeng, S.-p.; Liang, M.-h.; Yousaf, M.; Wu, Y.-p.; Tang, J.; Xiong, J.; Liu, D.-m. Safety and metabolism characteristics of Lacticaseibacillus rhamnosus LR-ZB1107-01 based on complete genome and corresponding phenotype. LWT 2024, 204, 116443. [Google Scholar] [CrossRef]
- Cheignon, C.m.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hureau, C.; Collin, F. Oxidative stress and the amyloid beta peptide in Alzheimer’s disease. Redox Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Zhang, L.; Hu, C.; Zhou, L.; Cheng, Y.; Liu, Q. Ellagic acid (EA) ameliorates Alzheimer’s disease by reducing Aβ levels, oxidative stress and attenuating inflammation. Eur. J. Pharmacol. 2025, 986, 177099. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.Y.; Lee, S.H. Ameliorating activity of Ishige okamurae on the amyloid beta–induced cognitive deficits and neurotoxicity through regulating ERK, p38 MAPK, and JNK signaling in Alzheimer’s disease–like mice model. Mol. Nutr. Food Res. 2020, 64, 1901220. [Google Scholar] [CrossRef]
- Yun, S.-W.; Park, H.-S.; Shin, Y.-J.; Ma, X.; Han, M.J.; Kim, D.-H. Lactobacillus gasseri NK109 and its supplement alleviate cognitive impairment in mice by modulating NF-κB activation, BDNF expression, and gut microbiota composition. Nutrients 2023, 15, 790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, Z.; Liu, L.; Li, H.; Wang, T.; Zhu, X.; Wang, Y.; Zhu, D.; Ning, Y.; Xu, Y. Probiotic has prophylactic effect on spatial memory deficits by modulating gut microbiota characterized by the inhibitory growth of Escherichia coli. Front. Integr. Neurosci. 2023, 17, 1090294. [Google Scholar] [CrossRef]
- Ionescu-Tucker, A.; Cotman, C.W. Emerging roles of oxidative stress in brain aging and Alzheimer’s disease. Neurobiol. Aging 2021, 107, 86–95. [Google Scholar] [CrossRef]
- Kobatake, E.; Nakagawa, H.; Seki, T.; Miyazaki, T. Protective effects and functional mechanisms of Lactobacillus gasseri SBT2055 against oxidative stress. PLoS ONE 2017, 12, e0177106. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Ma, K.; Li, J.; Li, J.; Bi, C.; Shan, A. Deoxynivalenol exposure induces liver damage in mice: Inflammation and immune responses, oxidative stress, and protective effects of Lactobacillus rhamnosus GG. Food. Chem. Toxicol. 2021, 156, 112514. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Q.; Sun, J.; Wang, J.; Liu, X.; Gao, J. Mitochondrial protective mechanism of simvastatin protects against amyloid β peptide-induced injury in SH-SY5Y cells. Int. J. Mol. Med. 2018, 41, 2997–3005. [Google Scholar] [PubMed]
- Chen, L.-H.; Huang, S.-Y.; Huang, K.-C.; Hsu, C.-C.; Yang, K.-C.; Li, L.-A.; Chan, C.-H.; Huang, H.-Y. Lactobacillus paracasei PS23 decelerated age-related muscle loss by ensuring mitochondrial function in SAMP8 mice. Aging 2019, 11, 756. [Google Scholar] [CrossRef]
- González-Bosch, C.; Boorman, E.; Zunszain, P.A.; Mann, G.E. Short-chain fatty acids as modulators of redox signaling in health and disease. Redox Biol. 2021, 47, 102165. [Google Scholar] [CrossRef]
- Sharp, P.S.; Dekkers, J.; Dick, J.R.; Greensmith, L. Manipulating transmitter release at the neuromuscular junction of neonatal rats alters the expression of ChAT and GAP-43 in motoneurons. Brain Res. Dev. Brain Res. 2003, 146, 29–38. [Google Scholar] [CrossRef]
- Taqui, R.; Debnath, M.; Ahmed, S.; Ghosh, A. Advances on plant extracts and phytocompounds with acetylcholinesterase inhibition activity for possible treatment of Alzheimer’s disease. Phytomed. Plus. 2022, 2, 100184. [Google Scholar] [CrossRef]
- Moyano, P.; Flores, A.; García, J.; García, J.M.; Anadon, M.J.; Frejo, M.T.; Sola, E.; Pelayo, A.; Del Pino, J. Bisphenol A single and repeated treatment increases HDAC2, leading to cholinergic neurotransmission dysfunction and SN56 cholinergic apoptotic cell death through AChE variants overexpression and NGF/TrkA/P75NTR signaling disruption. Food. Chem. Toxicol. 2021, 157, 112614. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Guo, A.; Guan, K.; Chen, C.; Xu, S.; Tang, Y.; Li, X.; Huang, Z. Lactobacillus rhamnosus GG attenuates depression-like behaviour and cognitive deficits in chronic ethanol exposure mice by down-regulating systemic inflammatory factors. Addict. Biol. 2024, 29, e13445. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Dasriya, V.L.; Nataraj, B.H.; Nagpal, R.; Behare, P.V. Lacticaseibacillus rhamnosus-derived exopolysaccharide attenuates D-galactose-induced oxidative stress and inflammatory brain injury and modulates gut microbiota in a mouse model. Microorganisms 2022, 10, 2046. [Google Scholar] [CrossRef] [PubMed]
- Dang, Q.; Wu, D.; Li, Y.; Fang, L.; Liu, C.; Wang, X.; Liu, X.; Min, W. Walnut-derived peptides ameliorate d-galactose-induced memory impairments in a mouse model via inhibition of MMP-9-mediated blood–brain barrier disruption. Food Res. Int. 2022, 162, 112029. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Bai, K.; Liu, X.-H.; Zhang, L.-M.; Yu, G.-R. Hyperoside protects the blood-brain barrier from neurotoxicity of amyloid beta 1–42. Neural. Regen. Res. 2018, 13, 1974–1980. [Google Scholar]
- Caccamo, A.; Oddo, S.; Sugarman, M.C.; Akbari, Y.; LaFerla, F.M. Age-and region-dependent alterations in Aβ-degrading enzymes: Implications for Aβ-induced disorders. Neurobiol. Aging 2005, 26, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Li, H.; Huang, Y.; Su, Y.; Li, Y.; Jia, A.; Jiang, Y.; Zhang, Y.; Man, C. Lactobacillus gasseri JM1 isolated from infant feces alleviates colitis in mice via protecting the intestinal barrier. Nutrients 2022, 15, 139. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Sato, N.; Ikimura, K.; Nishino, H.; Rakugi, H.; Morishita, R. Increased blood–brain barrier vulnerability to systemic inflammation in an Alzheimer disease mouse model. Neurobiol. Aging 2013, 34, 2064–2070. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.-R.; Jiang, J.-X.; Hu, Y.; Pan, J.-P.; Mi, X.-N.; Gao, Q.; Xiao, F.; Zhang, W.; Luo, H.-M. The immune system drives synapse loss during lipopolysaccharide-induced learning and memory impairment in mice. Front. Aging Neurosci. 2019, 11, 279. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Xian, X.; Xu, G.; Tan, Z.; Dong, F.; Zhang, M.; Zhang, F. Toll-like receptor 4: A promising therapeutic target for Alzheimer’s disease. Mediat. Inflamm. 2022, 2022, 7924199. [Google Scholar] [CrossRef] [PubMed]
- Bairamian, D.; Sha, S.; Rolhion, N.; Sokol, H.; Dorothée, G.; Lemere, C.A.; Krantic, S. Microbiota in neuroinflammation and synaptic dysfunction: A focus on Alzheimer’s disease. Mol. Neurodegener. 2022, 17, 19. [Google Scholar] [CrossRef]
- Kim, Y.; Cho, S.-H. Lindera glauca Blume ameliorates amyloid-β1-42-induced memory impairment in mice with neuroprotection and activation of the CREB-BDNF pathway. Neurochem. Int. 2021, 147, 105071. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, H.; Zhang, X.; Chen, L.; Zhao, X.; Bai, X.; Zhang, J. Nobiletin protects against cerebral ischemia via activating the p-Akt, p-CREB, BDNF and Bcl-2 pathway and ameliorating BBB permeability in rat. Brain Res. Bull. 2013, 96, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Changizi, V.; Azadbakht, O.; Ghanavati, R.; Behrouj, H.; Motevaseli, E.; Khanzadeh, P. Effect of Lactobacillus species on apoptosis-related genes BCL2, BAX, and caspase 3 in the testes of gamma-irradiated rats. Rev. Assoc. Med. Bras. 2021, 67, 1581–1585. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, R.; Li, X.; Shao, P.; Xie, J.; Su, X.; Xu, S.; Huang, Y.; Hu, S. Lactobacillus rhamnosus GG improves cognitive impairments in mice with sepsis. PeerJ 2024, 12, e17427. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-s.; Li, X.-h.; Liu, P.; Li, J.; Liu, L. The relationship between Alzheimer’s disease and intestinal microflora structure and inflammatory factors. Front. Aging Neurosci. 2022, 14, 972982. [Google Scholar] [CrossRef]
- Shao, S.; Jia, R.; Zhao, L.; Zhang, Y.; Guan, Y.; Wen, H.; Liu, J.; Zhao, Y.; Feng, Y.; Zhang, Z. Xiao-Chai-Hu-Tang ameliorates tumor growth in cancer comorbid depressive symptoms via modulating gut microbiota-mediated TLR4/MyD88/NF-κB signaling pathway. Phytomedicine 2021, 88, 153606. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Cao, Y.; Zhang, T.; Xu, C.; Liu, Z.; Li, W.; Wang, Q. Multisensory fusion training and 7,8-dihydroxyflavone improve amyloid-β-induced cognitive impairment, anxiety, and depression-like behavior in mice through multiple mechanisms. Neuropsychiatr. Dis. Treat. 2024, 2024, 1247–1270. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Bi, N.; Wang, T.; Huang, C.; Wang, R.; Xu, Y.; Wang, H.-L. Probiotic Lactobacillus rhamnosus GR-1 supplementation attenuates Pb-induced learning and memory deficits by reshaping the gut microbiota. Front. Nutr. 2022, 9, 934118. [Google Scholar] [CrossRef] [PubMed]
- Su, I.-J.; Hsu, C.-Y.; Shen, S.; Chao, P.-K.; Hsu, J.T.-A.; Hsueh, J.-T.; Liang, J.-J.; Hsu, Y.-T.; Shie, F.-S. The beneficial effects of combining anti-Aβ antibody NP106 and curcumin analog TML-6 on the treatment of Alzheimer’s disease in APP/PS1 mice. Int. J. Mol. Sci. 2022, 23, 556. [Google Scholar] [CrossRef]
- Fu, J.; Qin, Y.; Xiao, L.; Dai, X. Causal relationship between gut microflora and dementia: A Mendelian randomization study. Front. Microbiol. 2024, 14, 1306048. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | NC | Aβ | MG4247 | MG4644 | |
|---|---|---|---|---|---|
| Phylum | Firmicutes | 43.62 ± 8.32 b | 29.60 ± 1.46 c | 55.39 ± 13.27 a | 60.10 ± 10.98 a |
| Bacteroidota | 49.56 ± 9.55 ab | 62.20 ± 2.08 a | 37.22 ± 16.35 b | 35.84 ± 12.47 b | |
| Firmicutes/Bacteroidota ratio | 85.26 ± 9.33 b | 54.23 ± 3.62 c | 115.18 ± 15.56 a | 120.68 ± 10.38 a | |
| Family | Lachnospiraceae | 31.93 ± 17.78 b | 12.76 ± 4.05 c | 39.41 ± 12.25 ab | 50.35 ± 12.26 a |
| Rikenellaceae | 7.77 ± 1.41 b | 12.74 ± 4.20 a | 7.09 ± 2.78 b | 8.21 ± 1.54 b | |
| Tannerellaceae | 2.21 ± 0.84 ab | 3.01 ± 1.15 a | 0.61 ± 0.07 c | 1.54 ± 1.03 bc | |
| Genus | Lachnospiraceae NK4A136 | 0.12 ± 0.05 ab | 0.04 ± 0.01 b | 0.22 ± 0.13 a | 0.16 ± 0.05 ab |
| Helicobacter | 0.01 ± 0.00 c | 0.03 ± 0.00 a | 0.01 ± 0.00 b | 0.01 ± 0.00 c | |
| Rikenellaceae RC9 | 0.02 ± 0.01 b | 0.05 ± 0.01 a | 0.01 ± 0.00 b | 0.02 ± 0.01 b | |
| Bacteroides | 0.03 ± 0.00 b | 0.05 ± 0.01 a | 0.01 ± 0.00 d | 0.02 ± 0.00 c | |
| Alistipes | 0.04 ± 0.01 b | 0.06 ± 0.02 a | 0.04 ± 0.02 b | 0.03 ± 0.01 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.J.; Lee, H.L.; Kim, I.Y.; Ju, Y.H.; Heo, Y.M.; Na, H.R.; Lee, J.Y.; Choi, S.-I.; Heo, H.J. Oral Administration of Lactobacillus gasseri and Lacticaseibacillus rhamnosus Ameliorates Amyloid Beta (Aβ)-Induced Cognitive Impairment by Improving Synaptic Function Through Regulation of TLR4/Akt Pathway. Antioxidants 2025, 14, 139. https://doi.org/10.3390/antiox14020139
Choi HJ, Lee HL, Kim IY, Ju YH, Heo YM, Na HR, Lee JY, Choi S-I, Heo HJ. Oral Administration of Lactobacillus gasseri and Lacticaseibacillus rhamnosus Ameliorates Amyloid Beta (Aβ)-Induced Cognitive Impairment by Improving Synaptic Function Through Regulation of TLR4/Akt Pathway. Antioxidants. 2025; 14(2):139. https://doi.org/10.3390/antiox14020139
Chicago/Turabian StyleChoi, Hye Ji, Hyo Lim Lee, In Young Kim, Yeong Hyeon Ju, Yu Mi Heo, Hwa Rang Na, Ji Yeon Lee, Soo-Im Choi, and Ho Jin Heo. 2025. "Oral Administration of Lactobacillus gasseri and Lacticaseibacillus rhamnosus Ameliorates Amyloid Beta (Aβ)-Induced Cognitive Impairment by Improving Synaptic Function Through Regulation of TLR4/Akt Pathway" Antioxidants 14, no. 2: 139. https://doi.org/10.3390/antiox14020139
APA StyleChoi, H. J., Lee, H. L., Kim, I. Y., Ju, Y. H., Heo, Y. M., Na, H. R., Lee, J. Y., Choi, S.-I., & Heo, H. J. (2025). Oral Administration of Lactobacillus gasseri and Lacticaseibacillus rhamnosus Ameliorates Amyloid Beta (Aβ)-Induced Cognitive Impairment by Improving Synaptic Function Through Regulation of TLR4/Akt Pathway. Antioxidants, 14(2), 139. https://doi.org/10.3390/antiox14020139