
Effect of Media Composition and Oxygen Tension on Cellular Stress Response and Nrf2 Activation in HepG2ARE Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Nrf2 Activation Assay
2.3. MTT Assay
2.4. GSH and GSSG Assay
2.5. Cytosolic MDA Measurement
2.6. Data Analysis
3. Results
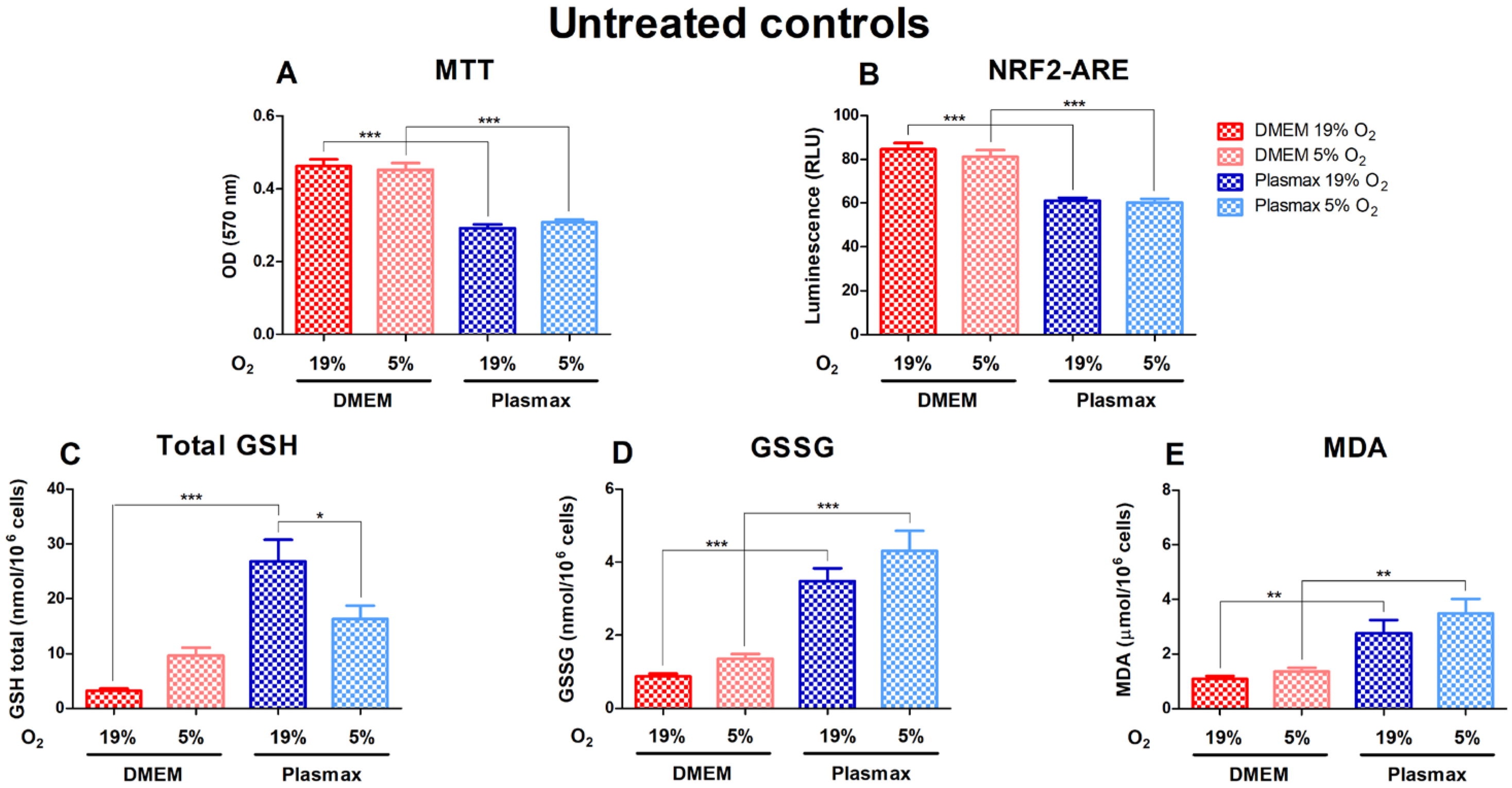
3.1. HepG2ARE Cell Growth and Antioxidant Status Depended on Media Composition and Oxygen Tension
3.2. The Effectiveness of Stress-Inducing Compounds on HepG2ARE Cells in Different Growth Conditions
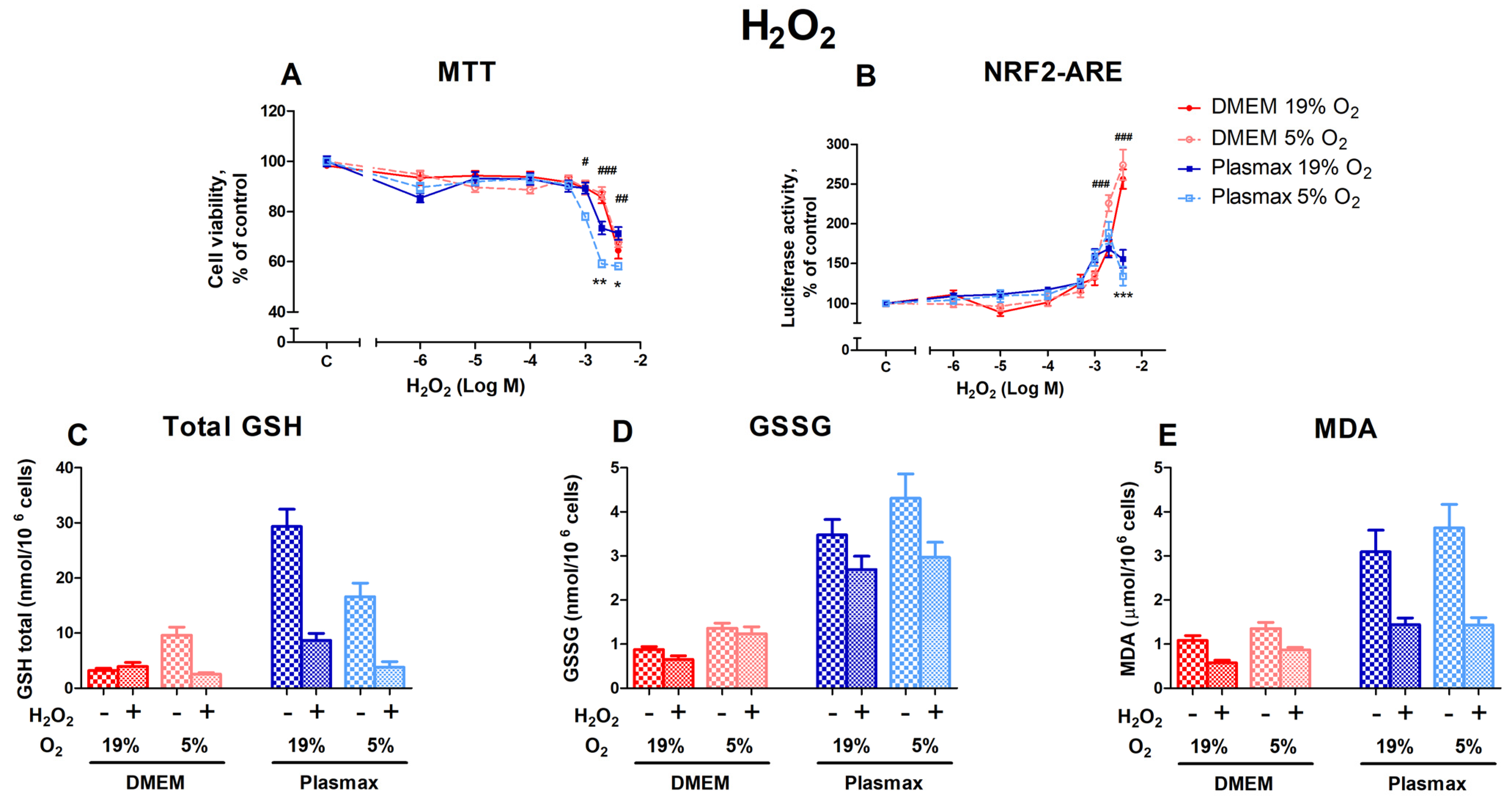
3.2.1. Cell Viability, Nrf2 Activation, and Redox Status of HepG2ARE Cells in Response to Hydrogen Peroxide Were Dependent on Media
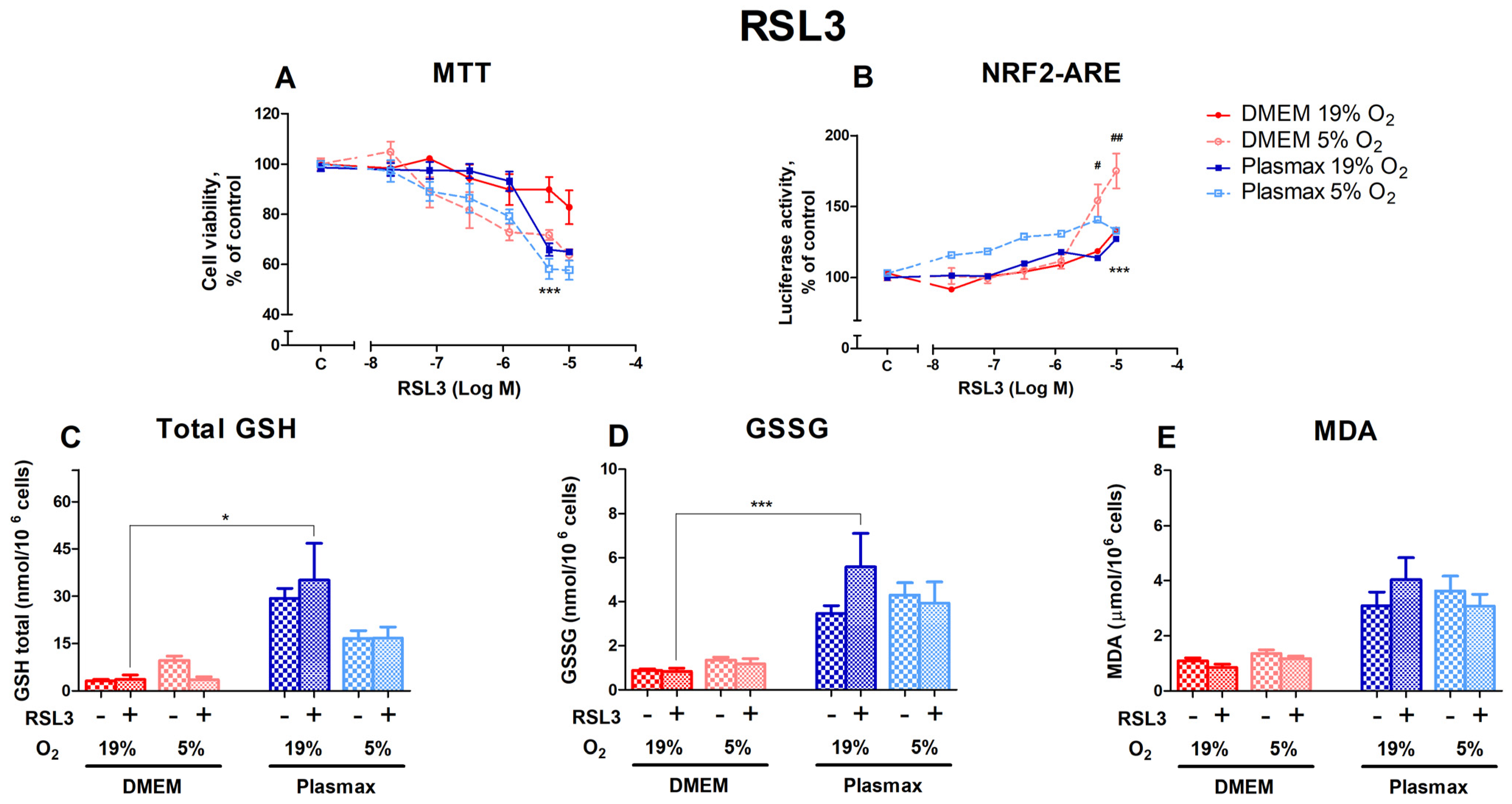
3.2.2. HepG2ARE Cells’ Response to Glutathione Peroxidase 4 Inhibitor RSL3 Was More Pronounced in 5% Oxygen
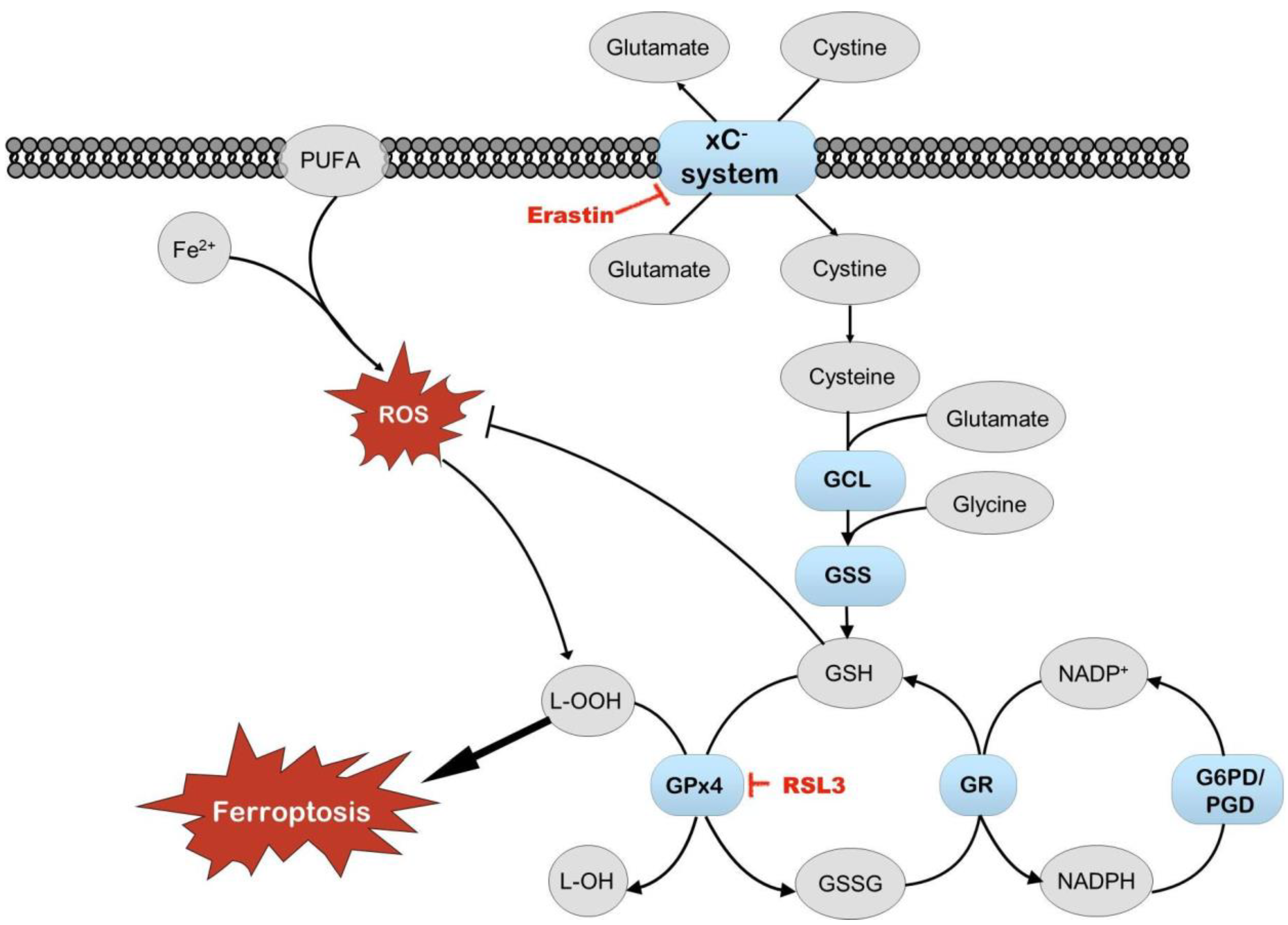
3.2.3. The Impact of the xC− System Inhibitor Erastin on HepG2ARE Cells Depended on Both Media and Oxygen
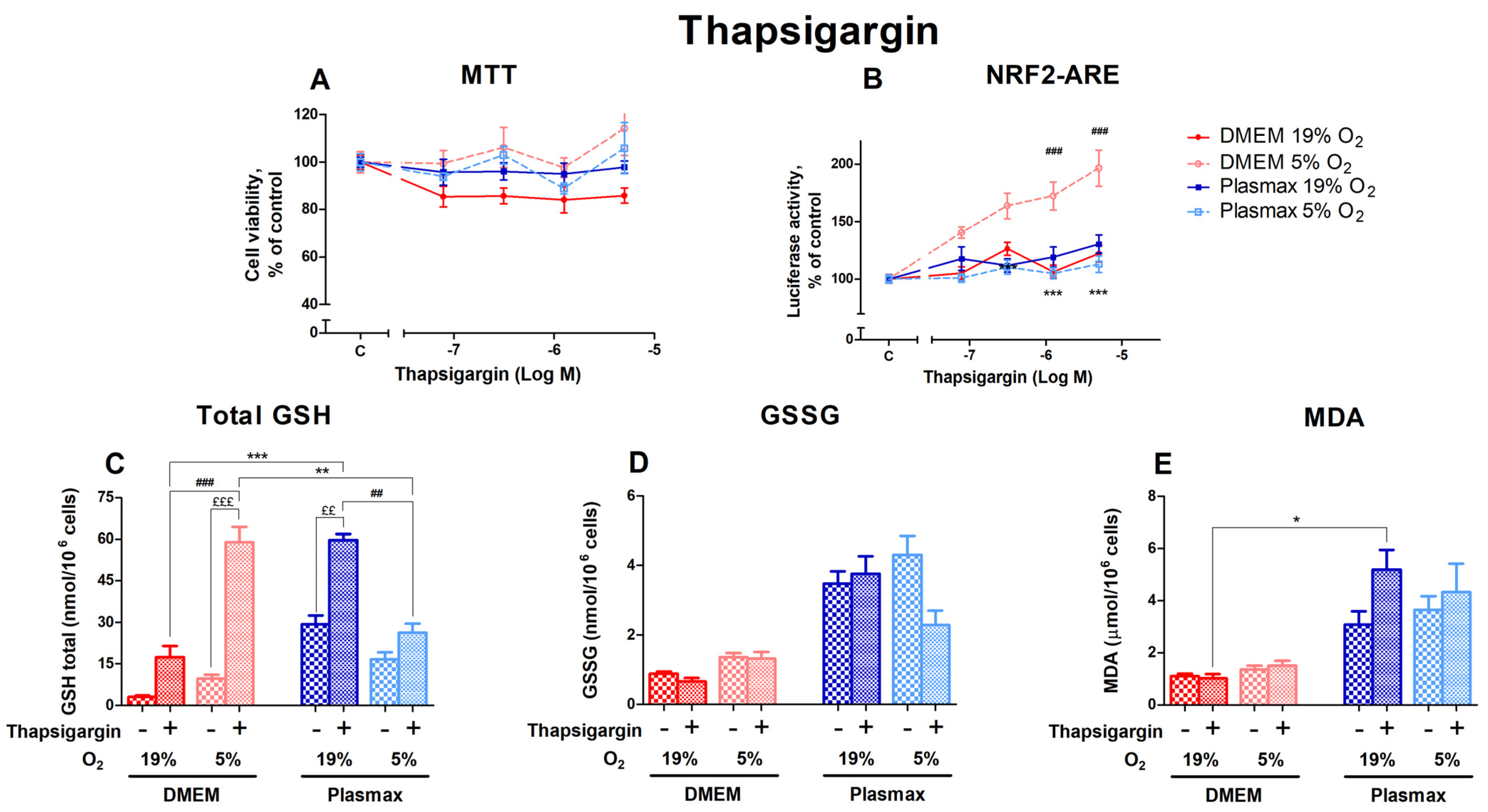
3.2.4. Oxygen Sensitivity of Thapsigargin Effect on Nrf2 Activation and Redox Status in HepG2ARE Cells Was Determined by Media
3.3. The Effect of Nrf2 Activators on HepG2ARE Cells in Different Growth Conditions
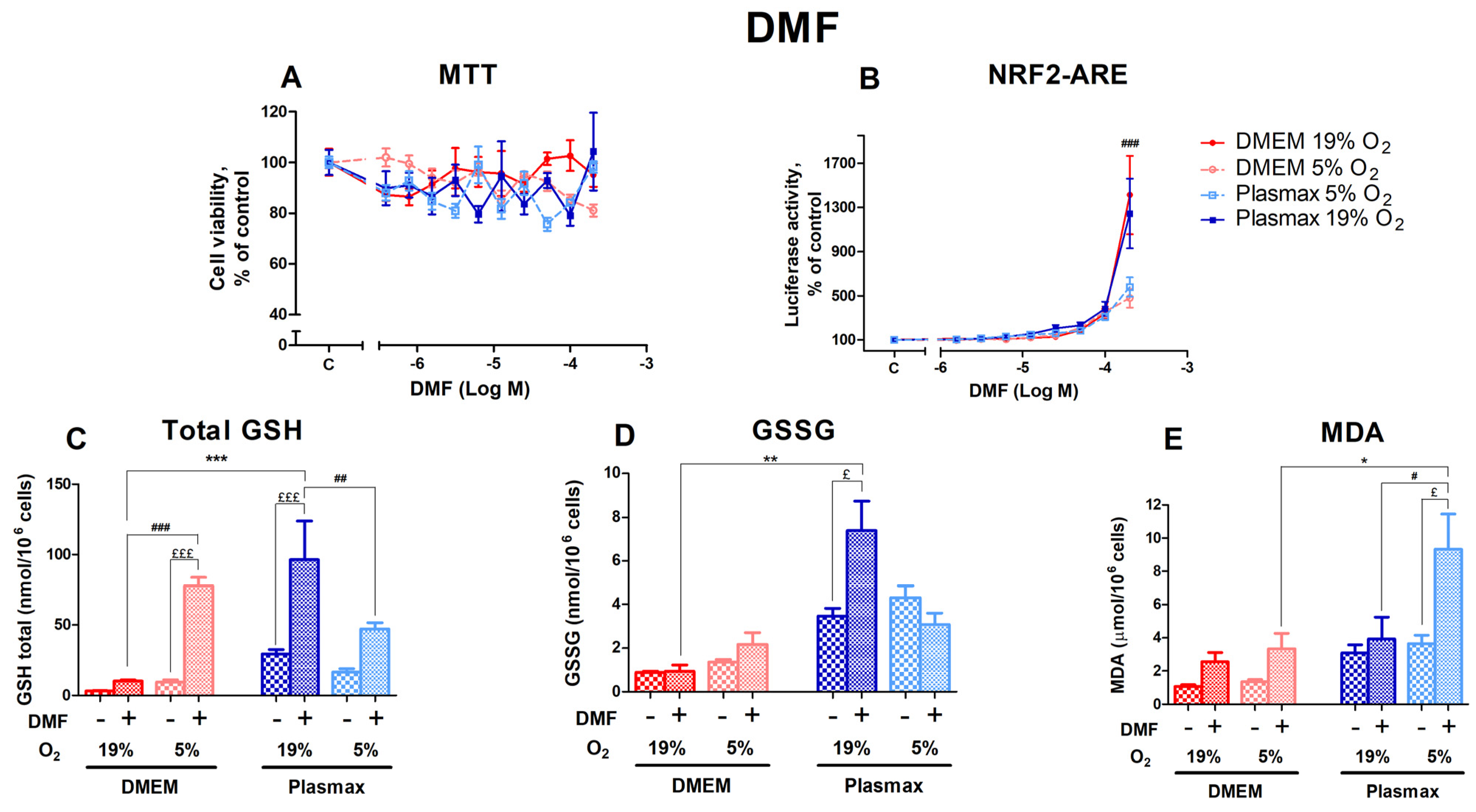
3.3.1. DMF’s Effect on the Redox Status of HepG2ARE Cells Depended on Both Media and Oxygen
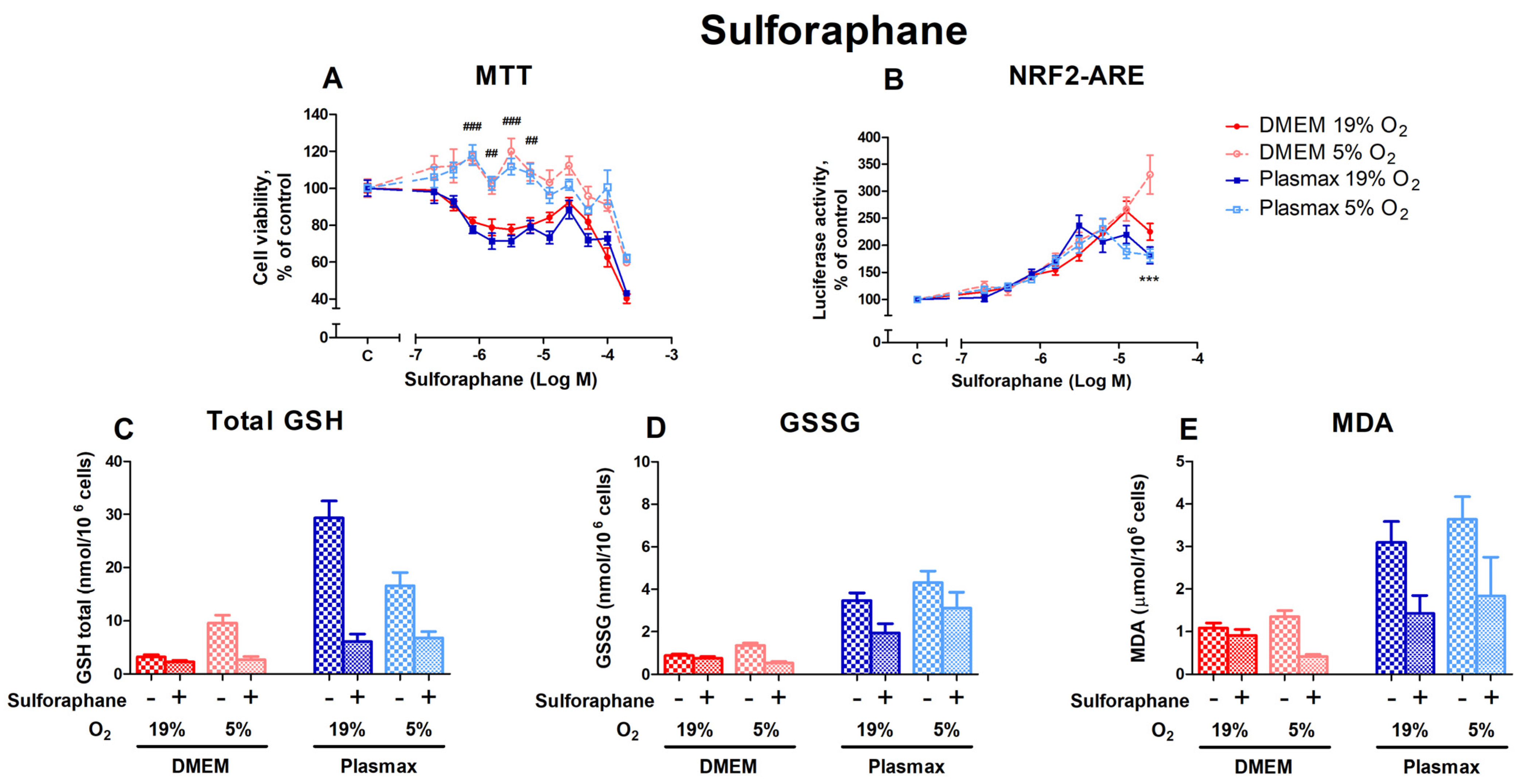
3.3.2. HepG2ARE Cells’ Response to Sulforaphane Is Most Evident at 5% Oxygen and Physiological Media
3.3.3. The Impact of Bardoxolone on HepG2ARE Cells’ Nrf2 Activation and Modulation of Redox Status Was Revealed in 5% Oxygen or Physiological Media
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eagle, H. Amino acid metabolism in mammalian cell cultures. Science 1959, 130, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H. The specific amino acid requirements of a human carcinoma cell (stain hela) in tissue culture. J. Exp. Med. 1955, 102, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H. The specific amino acid requirements of a mammalian cell (strain l) in tissue culture. J. Biol. Chem. 1955, 214, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H. Nutrition needs of mammalian cells in tissue culture. Science 1955, 122, 501–514. [Google Scholar] [CrossRef]
- Eagle, H. Amino acid metabolism in human cell cultures. Fed. Proc. 1958, 17, 985–986. [Google Scholar] [PubMed]
- Vande Voorde, J.; Ackermann, T.; Pfetzer, N.; Sumpton, D.; Mackay, G.; Kalna, G.; Nixon, C.; Blyth, K.; Gottlieb, E.; Tardito, S. Improving the metabolic fidelity of cancer models with a physiological cell culture medium. Sci. Adv. 2019, 5, eaau7314. [Google Scholar] [CrossRef] [PubMed]
- Dulbecco, R.; Freeman, G. Plaque production by the polyoma virus. Virology 1959, 8, 396–397. [Google Scholar] [CrossRef]
- Cantor, J.R.; Abu-Remaileh, M.; Kanarek, N.; Freinkman, E.; Gao, X.; Louissaint, A., Jr.; Lewis, C.A.; Sabatini, D.M. Physiologic medium rewires cellular metabolism and reveals uric acid as an endogenous inhibitor of ump synthase. Cell 2017, 169, 258–272.e217. [Google Scholar] [CrossRef]
- Ast, T.; Mootha, V.K. Oxygen and mammalian cell culture: Are we repeating the experiment of dr. Ox? Nat. Metab. 2019, 1, 858–860. [Google Scholar] [CrossRef] [PubMed]
- Alva, R.; Gardner, G.L.; Liang, P.; Stuart, J.A. Supraphysiological oxygen levels in mammalian cell culture: Current state and future perspectives. Cells 2022, 11, 3123. [Google Scholar] [CrossRef] [PubMed]
- Keeley, T.P.; Mann, G.E. Defining physiological normoxia for improved translation of cell physiology to animal models and humans. Physiol. Rev. 2019, 99, 161–234. [Google Scholar] [CrossRef] [PubMed]
- Warpsinski, G.; Smith, M.J.; Srivastava, S.; Keeley, T.P.; Siow, R.C.M.; Fraser, P.A.; Mann, G.E. Nrf2-regulated redox signaling in brain endothelial cells adapted to physiological oxygen levels: Consequences for sulforaphane mediated protection against hypoxia-reoxygenation. Redox Biol. 2020, 37, 101708. [Google Scholar] [CrossRef] [PubMed]
- Chettouh-Hammas, N.; Fasani, F.; Boileau, A.; Gosset, D.; Busco, G.; Grillon, C. Improvement of antioxidant defences in keratinocytes grown in physioxia: Comparison of 2d and 3d models. Oxid. Med. Cell. Longev. 2023, 2023, 6829931. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.L.; Kensler, T.W.; et al. Therapeutic targeting of the nrf2 and keap1 partnership in chronic diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef] [PubMed]
- El Alami, M.; Vina-Almunia, J.; Gambini, J.; Mas-Bargues, C.; Siow, R.C.; Penarrocha, M.; Mann, G.E.; Borras, C.; Vina, J. Activation of p38, p21, and nrf-2 mediates decreased proliferation of human dental pulp stem cells cultured under 21% O2. Stem Cell Rep. 2014, 3, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.; Chrusciel, S.; Fayad-Kobeissi, S.; Dubois-Rande, J.L.; Azuaje, F.; Boczkowski, J.; Motterlini, R.; Foresti, R. Permanent culture of macrophages at physiological oxygen attenuates the antioxidant and immunomodulatory properties of dimethyl fumarate. J. Cell. Physiol. 2015, 230, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Mari, M.; de Gregorio, E.; de Dios, C.; Roca-Agujetas, V.; Cucarull, B.; Tutusaus, A.; Morales, A.; Colell, A. Mitochondrial glutathione: Recent insights and role in disease. Antioxidants 2020, 9, 909. [Google Scholar] [CrossRef]
- Li, S.; Huang, Y. Ferroptosis: An iron-dependent cell death form linking metabolism, diseases, immune cell and targeted therapy. Clin. Transl. Oncol. 2022, 24, 1–12. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (mda) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Robledinos-Anton, N.; Fernandez-Gines, R.; Manda, G.; Cuadrado, A. Activators and inhibitors of nrf2: A review of their potential for clinical development. Oxid. Med. Cell. Longev. 2019, 2019, 9372182. [Google Scholar] [CrossRef] [PubMed]
- Lynch, D.R.; Perlman, S.; Schadt, K. Omaveloxolone for the treatment of friedreich ataxia: Clinical trial results and practical considerations. Expert Rev. Neurother. 2024, 24, 251–258. [Google Scholar] [CrossRef]
- Lynch, D.R.; Goldsberry, A.; Rummey, C.; Farmer, J.; Boesch, S.; Delatycki, M.B.; Giunti, P.; Hoyle, J.C.; Mariotti, C.; Mathews, K.D.; et al. Propensity matched comparison of omaveloxolone treatment to friedreich ataxia natural history data. Ann. Clin. Transl. Neurol. 2024, 11, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Subramony, S.H.; Lynch, D.L. A milestone in the treatment of ataxias: Approval of omaveloxolone for friedreich ataxia. Cerebellum 2024, 23, 775–777. [Google Scholar] [CrossRef]
- Profeta, V.; McIntyre, K.; Wells, M.; Park, C.; Lynch, D.R. Omaveloxolone: An activator of nrf2 for the treatment of friedreich ataxia. Expert Opin. Investig. Drugs 2023, 32, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Ghanekar, S.D.; Miller, W.W.; Meyer, C.J.; Fenelon, K.J.; Lacdao, A.; Zesiewicz, T.A. Orphan drugs in development for the treatment of friedreich’s ataxia: Focus on omaveloxolone. Degener. Neurol. Neuromuscul. Dis. 2019, 9, 103–107. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Escudero-Ferruz, P.; Ontiveros, N.; Cano-Estrada, C.; Sutcliffe, D.J.; Jinnah, H.A.; Torres, R.J.; Lopez, J.M. A new physiological medium uncovers biochemical and cellular alterations in lesch-nyhan disease fibroblasts. Mol. Med. 2024, 30, 3. [Google Scholar] [CrossRef] [PubMed]
- Antico Arciuch, V.G.; Elguero, M.E.; Poderoso, J.J.; Carreras, M.C. Mitochondrial regulation of cell cycle and proliferation. Antioxid. Redox Signal. 2012, 16, 1150–1180. [Google Scholar] [CrossRef]
- Avellino, G.; Deshmukh, R.; Rogers, S.N.; Charnock-Jones, D.S.; Smith, G.C.S.; Tardito, S.; Aye, I. Physiologically relevant culture medium plasmax improves human placental trophoblast stem cell function. Am. J. Physiology. Cell Physiol. 2023, 324, C878–C885. [Google Scholar] [CrossRef] [PubMed]
- Golikov, M.V.; Karpenko, I.L.; Lipatova, A.V.; Ivanova, O.N.; Fedyakina, I.T.; Larichev, V.F.; Zakirova, N.F.; Leonova, O.G.; Popenko, V.I.; Bartosch, B.; et al. Cultivation of cells in a physiological plasmax medium increases mitochondrial respiratory capacity and reduces replication levels of rna viruses. Antioxidants 2021, 11, 97. [Google Scholar] [CrossRef] [PubMed]
- Golikov, M.V.; Valuev-Elliston, V.T.; Smirnova, O.A.; Ivanov, A.V. Physiological media in studies of cell metabolism. Mol. Biol. 2022, 56, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Chetsawang, J.; Govitrapong, P.; Chetsawang, B. Hydrogen peroxide toxicity induces ras signaling in human neuroblastoma sh-sy5y cultured cells. J. Biomed. Biotechnol. 2010, 2010, 803815. [Google Scholar] [CrossRef]
- Covas, G.; Marinho, H.S.; Cyrne, L.; Antunes, F. Activation of nrf2 by H2O2: De novo synthesis versus nuclear translocation. Methods Enzymol. 2013, 528, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Chen, Q.M. Fresh medium or l-cystine as an effective nrf2 inducer for cytoprotection in cell culture. Cells 2023, 12, 291. [Google Scholar] [CrossRef] [PubMed]
- Babich, H.; Liebling, E.J.; Burger, R.F.; Zuckerbraun, H.L.; Schuck, A.G. Choice of dmem, formulated with or without pyruvate, plays an important role in assessing the in vitro cytotoxicity of oxidants and prooxidant nutraceuticals. Vitr. Cell. Dev. Biol. Anim. 2009, 45, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Bailey-Downs, L.; Gautam, T.; Jimenez, R.; Losonczy, G.; Zhang, C.; Ballabh, P.; Recchia, F.A.; Wilkerson, D.C.; Sonntag, W.E.; et al. Adaptive induction of nf-e2-related factor-2-driven antioxidant genes in endothelial cells in response to hyperglycemia. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1133–H1140. [Google Scholar] [CrossRef] [PubMed]
- Abdalkader, M.; Lampinen, R.; Kanninen, K.M.; Malm, T.M.; Liddell, J.R. Targeting nrf2 to suppress ferroptosis and mitochondrial dysfunction in neurodegeneration. Front. Neurosci. 2018, 12, 466. [Google Scholar] [CrossRef]
- Keeley, T.P.; Siow, R.C.M.; Jacob, R.; Mann, G.E. Reduced serca activity underlies dysregulation of Ca2+ homeostasis under atmospheric O2 levels. FASEB J. 2018, 32, 2531–2538. [Google Scholar] [CrossRef]
- Moradi, F.; Moffatt, C.; Stuart, J.A. The effect of oxygen and micronutrient composition of cell growth media on cancer cell bioenergetics and mitochondrial networks. Biomolecules 2021, 11, 1177. [Google Scholar] [CrossRef] [PubMed]
- Moradi, F.; Fiocchetti, M.; Marino, M.; Moffatt, C.; Stuart, J.A. Media composition and O2 levels determine effects of 17beta-estradiol and selective estrogen receptor modulators on mitochondrial bioenergetics and cellular reactive oxygen species. Am. J. Physiol. Cell Physiol. 2021, 321, C72–C81. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taba, R.; Põlluaed, M.; Tein, K.; Puurand, M.; Käämbre, T.; Terasmaa, A. Effect of Media Composition and Oxygen Tension on Cellular Stress Response and Nrf2 Activation in HepG2ARE Cells. Antioxidants 2025, 14, 137. https://doi.org/10.3390/antiox14020137
Taba R, Põlluaed M, Tein K, Puurand M, Käämbre T, Terasmaa A. Effect of Media Composition and Oxygen Tension on Cellular Stress Response and Nrf2 Activation in HepG2ARE Cells. Antioxidants. 2025; 14(2):137. https://doi.org/10.3390/antiox14020137
Chicago/Turabian StyleTaba, Rutt, Marie Põlluaed, Karin Tein, Marju Puurand, Tuuli Käämbre, and Anton Terasmaa. 2025. "Effect of Media Composition and Oxygen Tension on Cellular Stress Response and Nrf2 Activation in HepG2ARE Cells" Antioxidants 14, no. 2: 137. https://doi.org/10.3390/antiox14020137
APA StyleTaba, R., Põlluaed, M., Tein, K., Puurand, M., Käämbre, T., & Terasmaa, A. (2025). Effect of Media Composition and Oxygen Tension on Cellular Stress Response and Nrf2 Activation in HepG2ARE Cells. Antioxidants, 14(2), 137. https://doi.org/10.3390/antiox14020137