
Enhanced In Vitro Efficacy of Verbascoside in Suppressing Hepatic Stellate Cell Activation via ROS Scavenging with Reverse Microemulsion

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Treatment
2.3. Determination of Cell Viability by MTT Assay
2.4. Reverse Transcription–Quantitative PCR (RT-qPCR)
2.5. Protein Extraction and Western Blot Analysis
2.6. Construction of Pseudo-Ternary Phase Diagrams
2.7. Preparation of VB-Loaded Microemulsion
2.8. Particle Size, Size Distribution, and Zeta Potential Analysis
2.9. Infrared Spectrum Analysis
2.10. High-Performance Liquid Chromatography
2.11. Cell Migration Assay
2.12. Ki67 Immunofluorescence Assay
2.13. Reactive Oxygen Species (ROS) Assay
2.14. Detecting the Activity of SOD
2.15. Statistical Analysis
3. Results
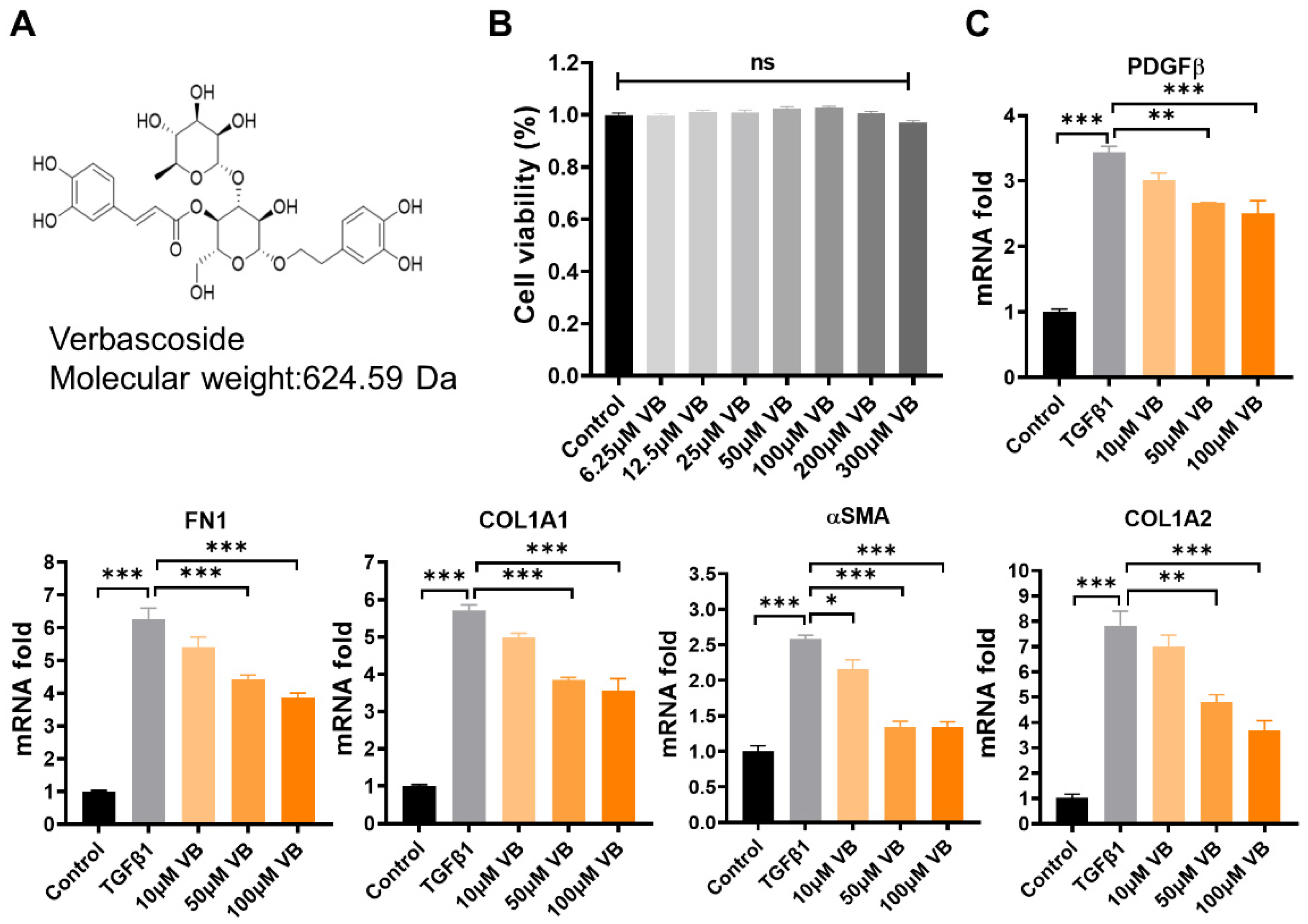
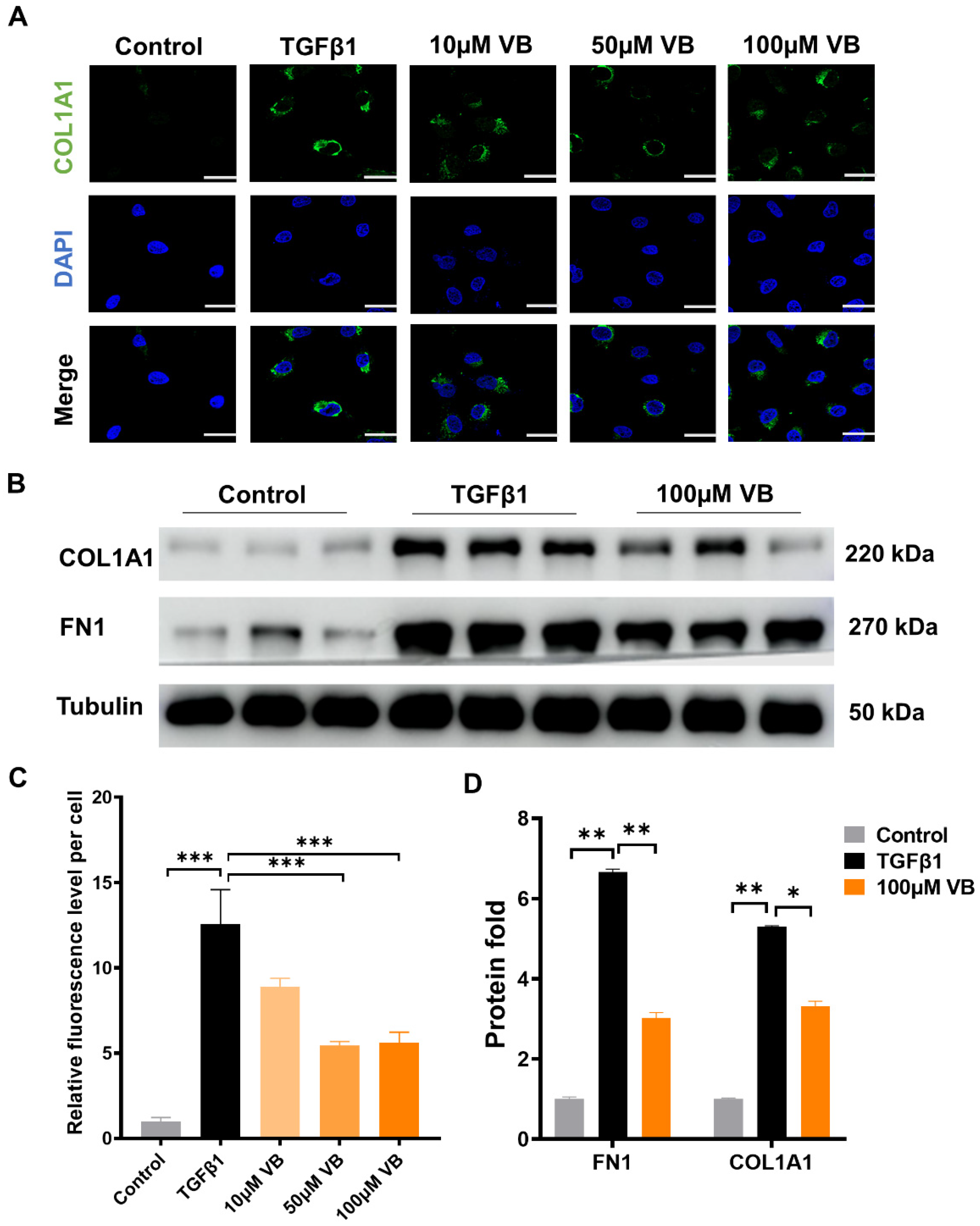
3.1. VB Suppresses the mRNA and Protein Levels of Fibrogenic Genes in Activated LX-2 Cells
3.2. VB Scavenges ROS in Activated LX-2 Cells and Downregulates MAPK Pathway
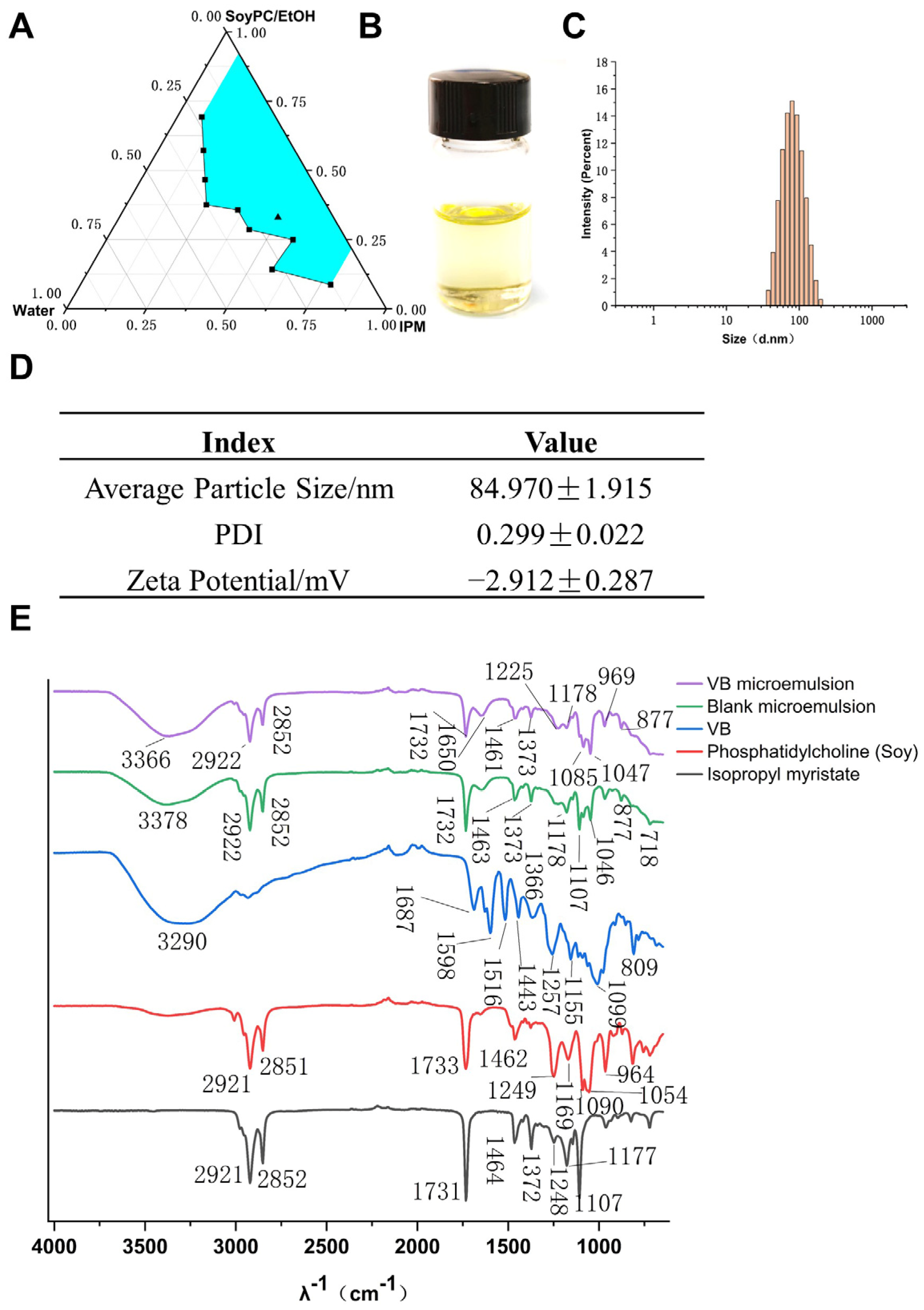
3.3. Development and Characterization of Verbascoside-Loaded Reverse Microemulsion
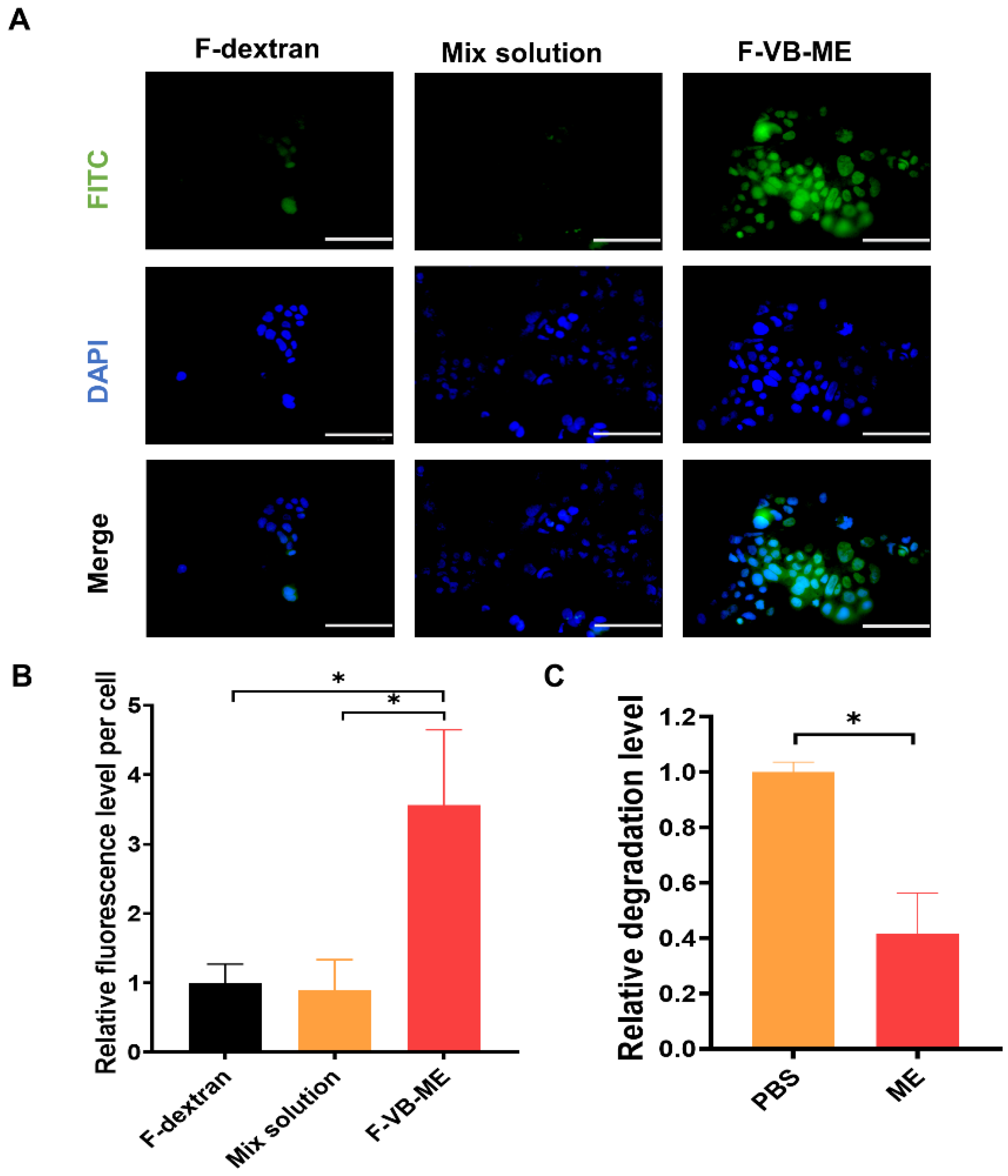
3.4. VB-ME Enhanced Stability and Cell Uptake of VB
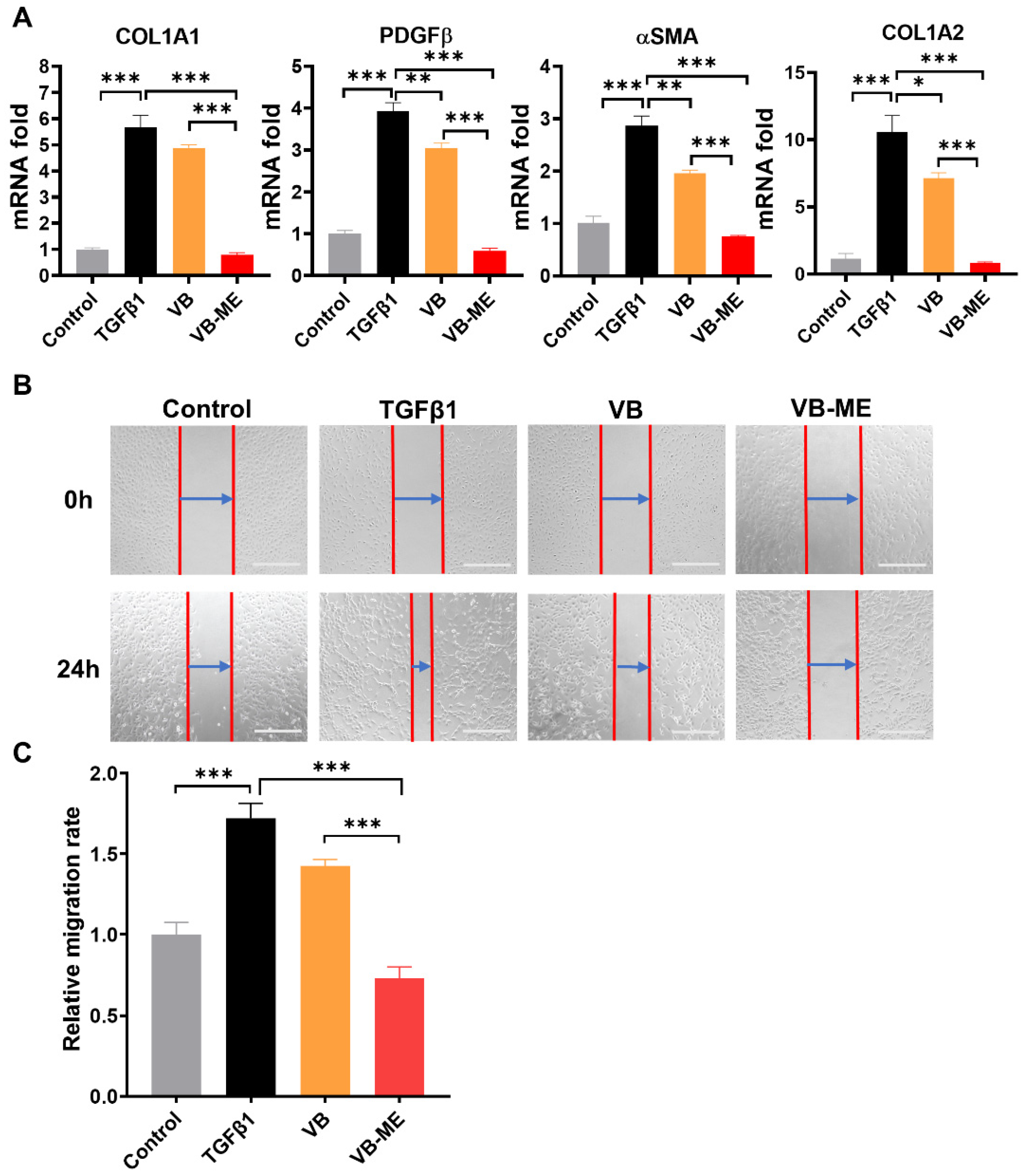
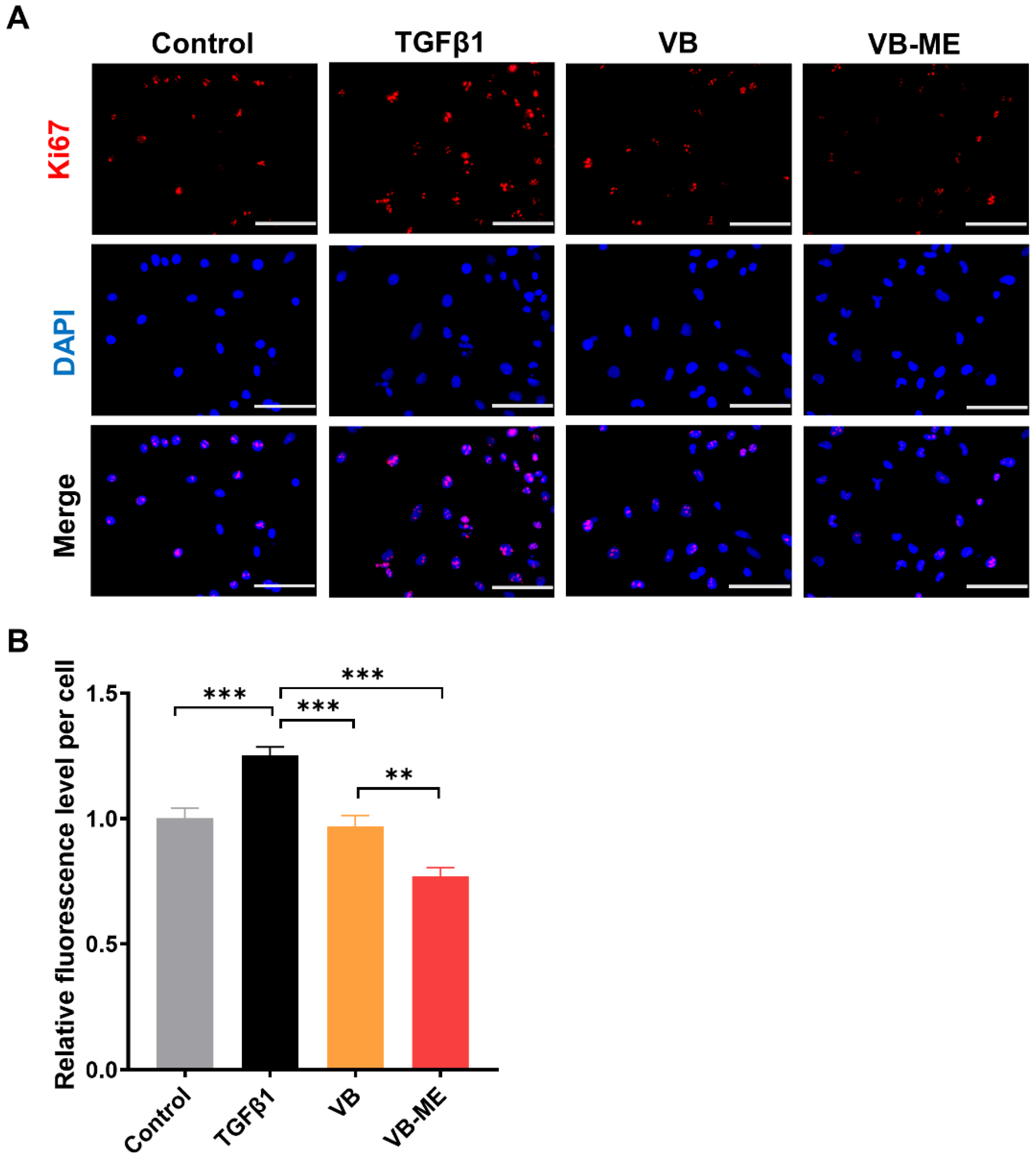
3.5. VB-ME Enhanced the Suppression of Fibrogenic Genes and Activation Features in LX-2 Cells
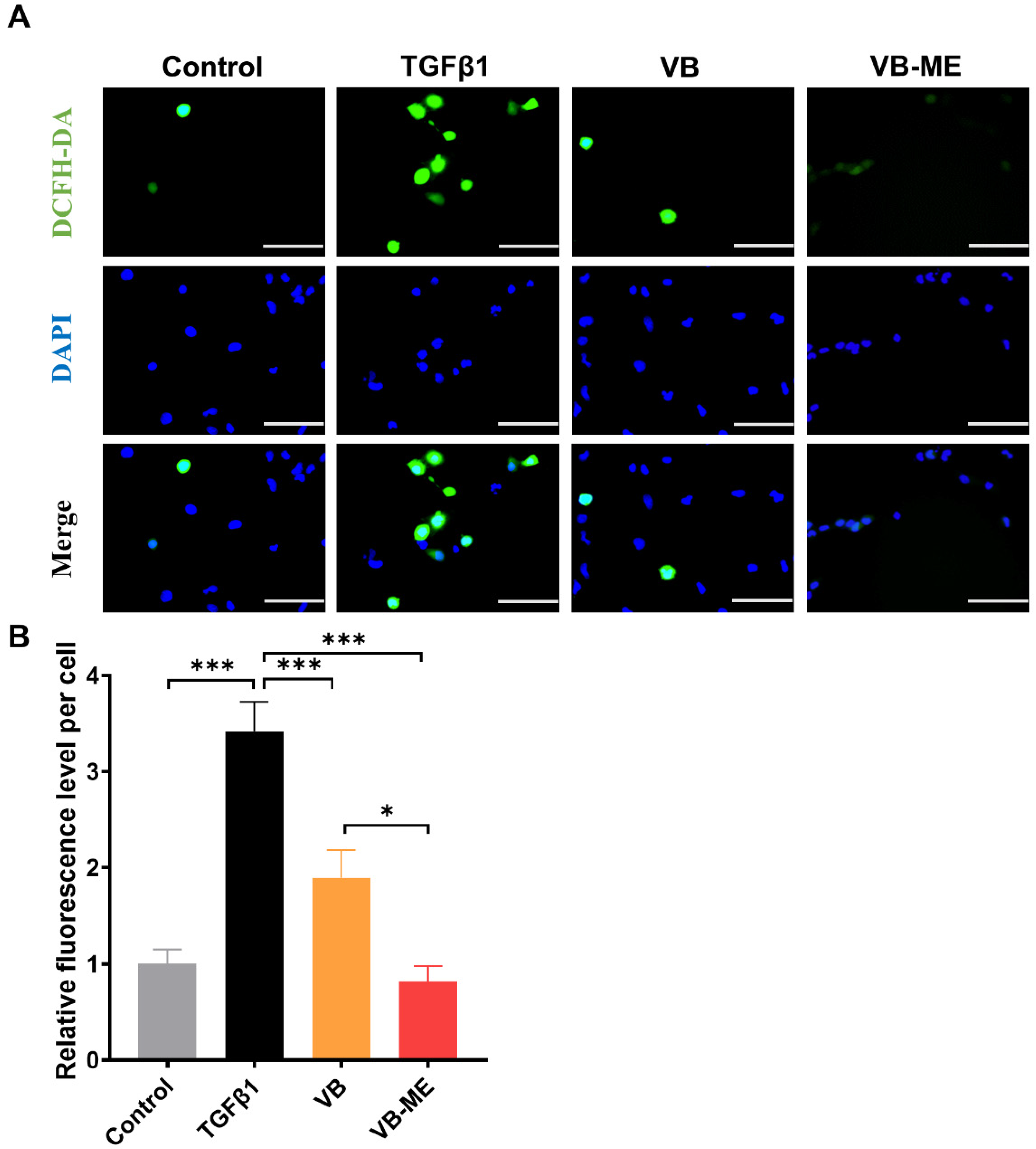
3.6. VB-ME Enhanced the Scavenging of ROS in LX-2 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global burden of liver disease: 2023 update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Orci, L.A.; Sanduzzi-Zamparelli, M.; Caballol, B.; Sapena, V.; Colucci, N.; Torres, F.; Bruix, J.; Reig, M.; Toso, C. Incidence of hepatocellular carcinoma in patients with nonalcoholic fatty liver disease: A systematic review, meta-analysis, and meta-regression. Clin. Gastroenterol. Hepatol. 2022, 20, 283–292.e10. [Google Scholar] [CrossRef]
- Fallowfield, J.; Hayes, P. Pathogenesis and treatment of hepatic fibrosis: Is cirrhosis reversible? Clin. Med. 2011, 11, 179–183. [Google Scholar] [CrossRef]
- Puche, J.E.; Saiman, Y.; Friedman, S.L. Hepatic stellate cells and liver fibrosis. Compr. Physiol. 2013, 3, 1473–1492. [Google Scholar] [CrossRef] [PubMed]
- Lee, U.E.; Friedman, S.L. Mechanisms of hepatic fibrogenesis. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 195–206. [Google Scholar] [CrossRef]
- Baghaei, K.; Mazhari, S.; Tokhanbigli, S.; Parsamanesh, G.; Alavifard, H.; Schaafsma, D.; Ghavami, S. Therapeutic potential of targeting regulatory mechanisms of hepatic stellate cell activation in liver fibrosis. Drug Discov. Today 2022, 27, 1044–1061. [Google Scholar] [CrossRef]
- Higashi, T.; Friedman, S.L.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Guo, J.S. Developing natural marine products for treating liver diseases. World J. Clin. Cases 2022, 10, 2369–2381. [Google Scholar] [CrossRef]
- Chan, Y.T.; Wang, N.; Tan, H.Y.; Li, S.; Feng, Y. Targeting hepatic stellate cells for the treatment of liver fibrosis by natural products: Is it the dawning of a new era? Front. Pharmacol. 2020, 11, 548. [Google Scholar] [CrossRef]
- Shan, L.; Liu, Z.; Ci, L.; Shuai, C.; Lv, X.; Li, J. Research progress on the anti-hepatic fibrosis action and mechanism of natural products. Int. Immunopharmacol. 2019, 75, 105765. [Google Scholar] [CrossRef]
- Rossi, R.; Mainardi, E.; Vizzarri, F.; Corino, C. Verbascoside-rich plant extracts in animal nutrition. Antioxidants 2023, 13, 39. [Google Scholar] [CrossRef]
- Sciandra, F.; Bottoni, P.; De Leo, M.; Braca, A.; Brancaccio, A.; Bozzi, M. Verbascoside elicits its beneficial effects by enhancing mitochondrial spare respiratory capacity and the nrf2/ho-1 mediated antioxidant system in a murine skeletal muscle cell line. Int. J. Mol. Sci. 2023, 24, 15276. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, S.; Pan, J.; Ma, K. Verbascoside: A neuroprotective phenylethanoid glycosides with anti-depressive properties. Phytomedicine 2023, 120, 155027. [Google Scholar] [CrossRef]
- Mao, Y.; Yu, J.; Da, J.; Yu, F.; Zha, Y. Acteoside alleviates uuo-induced inflammation and fibrosis by regulating the hmgn1/tlr4/trem1 signaling pathway. PeerJ 2023, 11, e14765. [Google Scholar] [CrossRef]
- Jiang, Y.; Lin, X.; Mao, Y.; Zhao, J.; Zhang, G.; Yu, J.; Dong, R.; Zha, Y. Acteoside alleviates renal fibrosis by inhibiting β-catenin/ctgf signaling pathway in uuo rats. Nat. Prod. Commun. 2022, 17, 1934578X221134880. [Google Scholar] [CrossRef]
- Chen, C.Y.; Tung, H.Y.; Tseng, Y.F.; Huang, J.S.; Shi, L.S.; Ye, Y.L. Verbascoside and isoverbascoside ameliorate transforming growth factor β1-induced collagen expression by lung fibroblasts through Smad/non-Smad signaling pathways. Life Sci. 2022, 308, 120950. [Google Scholar] [CrossRef]
- D’Imperio, M.; Cardinali, A.; D’Antuono, I.; Linsalata, V.; Minervini, F.; Redan, B.W.; Ferruzzi, M.G. Stability–activity of verbascoside, a known antioxidant compound, at different pH conditions. Food Res. Int. 2014, 66, 373–378. [Google Scholar] [CrossRef]
- Xiao, Y.; Ren, Q.; Wu, L. The pharmacokinetic property and pharmacological activity of acteoside: A review. Biomed. Pharmacother. 2022, 153, 113296. [Google Scholar] [CrossRef]
- Zhang, W.; Huo, S.X.; Wen, Y.L.; Xing, H.; Zhang, Q.; Li, N.; Zhao, D.; Sun, X.L.; Xu, J.; Yan, M.; et al. Pharmacokinetics of acteoside following single dose intragastric and intravenous administrations in dogs. Chin. J. Nat. Med. 2015, 13, 634–640. [Google Scholar] [CrossRef]
- Yang, X.; Li, W.; Li, S.; Chen, S.; Hu, Z.; He, Z.; Zhu, X.; Niu, X.; Zhou, X.; Li, H.; et al. Fish oil-based microemulsion can efficiently deliver oral peptide blocking pd-1/pd-l1 and simultaneously induce ferroptosis for cancer immunotherapy. J. Control. Release 2024, 365, 654–667. [Google Scholar] [CrossRef]
- Nikolaev, B.; Yakovleva, L.; Fedorov, V.; Li, H.; Gao, H.; Shevtsov, M. Nano- and microemulsions in biomedicine: From theory to practice. Pharmaceutics 2023, 15, 1989. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Yang, J.; Huang, Y.; Li, J.; Li, Y. Gut microbiota metabolite 3-indolepropionic acid directly activates hepatic stellate cells by ros/jnk/p38 signaling pathways. Biomolecules 2023, 13, 1464. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Chen, Y.; Miao, J.; Li, Y.; Liu, J.; Zhang, J.; Liang, J.; Chen, S.; Hou, S. Esculin inhibits hepatic stellate cell activation and ccl(4)-induced liver fibrosis by activating the nrf2/gpx4 signaling pathway. Phytomedicine 2024, 128, 155465. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Xu, G.; Ni, Q.; Wang, Y.; Gao, Q.; Zhang, Y. Microemulsion delivery system improves cellular uptake of genipin and its protective effect against aβ1-42-induced pc12 cell cytotoxicity. Pharmaceutics 2022, 14, 617. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Zender, L.; Miething, C.; Dickins, R.A.; Hernando, E.; Krizhanovsky, V.; Cordon-Cardo, C.; Lowe, S.W. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature 2007, 445, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Yang, X.; Dong, N.; Xie, X.; Bai, X.; Shi, Y. Generation and selection of immunized fab phage display library against human b cell lymphoma. Cell Res. 2007, 17, 650–660. [Google Scholar] [CrossRef]
- Zhu, S.; Zeng, J.; Pan, C.; Chai, Y.; Bai, M.; Li, J.; Chen, A. Reverse microemulsions as nano-carriers of tea polyphenols retard oxidation of eucommia ulmoides oliver seed oil. Colloids Surf. A Physicochem. Eng. Asp. 2023, 671, 131687. [Google Scholar] [CrossRef]
- He, E.; Li, H.; Li, X.; Wu, X.; Lei, K.; Diao, Y. Transdermal delivery of indirubin-loaded microemulsion gel: Preparation, characterization and anti-psoriatic activity. Int. J. Mol. Sci. 2022, 23, 3798. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhu, Q.; Dong, X.; Pan, H.; Chen, J.; Zheng, Z.P. Oxyresveratrol and ascorbic acid o/w microemulsion: Preparation, characterization, anti-isomerization and potential application as antibrowning agent on fresh-cut lotus root slices. Food Chem. 2017, 214, 269–276. [Google Scholar] [CrossRef]
- Cao, X.; Liang, Y.; Liu, R.; Zao, X.; Zhang, J.; Chen, G.; Liu, R.; Chen, H.; He, Y.; Zhang, J.; et al. Uncovering the pharmacological mechanisms of gexia-zhuyu formula (gxzy) in treating liver cirrhosis by an integrative pharmacology strategy. Front. Pharmacol. 2022, 13, 793888. [Google Scholar] [CrossRef]
- Gabbia, D.; Carpi, S.; Sarcognato, S.; Cannella, L.; Colognesi, M.; Scaffidi, M.; Polini, B.; Digiacomo, M.; Esposito Salsano, J.; Manera, C.; et al. The extra virgin olive oil polyphenol oleocanthal exerts antifibrotic effects in the liver. Front. Nutr. 2021, 8, 715183. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Xue, X. Detection of total reactive oxygen species in adherent cells by 2′,7′-dichlorodihydrofluorescein diacetate staining. J. Vis. Exp. 2020, 160, e60682. [Google Scholar] [CrossRef]
- Weng, Q.; Sun, H.; Fang, C.; Xia, F.; Liao, H.; Lee, J.; Wang, J.; Xie, A.; Ren, J.; Guo, X.; et al. Catalytic activity tunable ceria nanoparticles prevent chemotherapy-induced acute kidney injury without interference with chemotherapeutics. Nat. Commun. 2021, 12, 1436. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Hui, A.Y.; Albanis, E.; Arthur, M.J.; O’Byrne, S.M.; Blaner, W.S.; Mukherjee, P.; Friedman, S.L.; Eng, F.J. Human hepatic stellate cell lines, lx-1 and lx-2: New tools for analysis of hepatic fibrosis. Gut 2005, 54, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Tovar, E.; Muriel, P. Molecular mechanisms that link oxidative stress, inflammation, and fibrosis in the liver. Antioxidants 2020, 9, 1279. [Google Scholar] [CrossRef] [PubMed]
- Burgos, C.; Muñoz-Mingarro, D.; Navarro, I.; Martín-Cordero, C.; Acero, N. Neuroprotective potential of verbascoside isolated from acanthus mollis l. Leaves through its enzymatic inhibition and free radical scavenging ability. Antioxidants 2020, 9, 1207. [Google Scholar] [CrossRef] [PubMed]
- Papich, M.G.; Martinez, M.N. Applying biopharmaceutical classification system (bcs) criteria to predict oral absorption of drugs in dogs: Challenges and pitfalls. AAPS J. 2015, 17, 948–964. [Google Scholar] [CrossRef] [PubMed]
- Yameen, B.; Choi, W.I.; Vilos, C.; Swami, A.; Shi, J.; Farokhzad, O.C. Insight into nanoparticle cellular uptake and intracellular targeting. J. Control. Release 2014, 190, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Dinache, A.; Tozar, T.; Smarandache, A.; Andrei, I.R.; Nistorescu, S.; Nastasa, V.; Staicu, A.; Pascu, M.L.; Romanitan, M.O. Spectroscopic characterization of emulsions generated with a new laser-assisted device. Molecules 2020, 25, 1729. [Google Scholar] [CrossRef]
- Jiang, J.X.; Török, N.J. Liver injury and the activation of the hepatic myofibroblasts. Curr. Pathobiol. Rep. 2013, 1, 215–223. [Google Scholar] [CrossRef]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.H.; Qian, Z.J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.K. Hepatic fibrosis inhibitory effect of peptides isolated from navicula incerta on tgf-β1 induced activation of lx-2 human hepatic stellate cells. Prev. Nutr. Food Sci. 2013, 18, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Dewidar, B.; Meyer, C.; Dooley, S.; Meindl-Beinker, A.N. Tgf-β in hepatic stellate cell activation and liver fibrogenesis-updated 2019. Cells 2019, 8, 1419. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Huda, M.N.; Shin, Y.; Han, S.; Akter, S.; Kang, I.; Ha, J.; Choe, W.; Choi, T.G.; Kim, S.S. Correlation between oxidative stress and transforming growth factor-beta in cancers. Int. J. Mol. Sci. 2021, 22, 13181. [Google Scholar] [CrossRef] [PubMed]
- Foglia, B.; Cannito, S.; Bocca, C.; Parola, M.; Novo, E. Erk pathway in activated, myofibroblast-like, hepatic stellate cells: A critical signaling crossroad sustaining liver fibrosis. Int. J. Mol. Sci. 2019, 20, 2700. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, C.; Wang, H.; Li, L.; Wang, C. The function of mapk cascades in response to various stresses in horticultural plants. Front. Plant Sci. 2020, 11, 952. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Cheong, Y.K.; Kim, N.H.; Chung, H.T.; Kang, D.G.; Pae, H.O. Mitogen-activated protein kinases and reactive oxygen species: How can ros activate mapk pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef] [PubMed]
- Liebert, M.A. Final report on the safety assessment of myristyl myristate and isopropyl myristate. Int. J. Toxicol. 1982, 1, 55–80. [Google Scholar]
- Jin, S.; Li, S.; Wang, C.; Liu, J.; Yang, X.; Wang, P.C.; Zhang, X.; Liang, X.J. Biosafe nanoscale pharmaceutical adjuvant materials. J. Biomed. Nanotechnol. 2014, 10, 2393–2419. [Google Scholar] [CrossRef]
- Vertuani, S.; Beghelli, E.; Scalambra, E.; Malisardi, G.; Copetti, S.; Dal Toso, R.; Baldisserotto, A.; Manfredini, S. Activity and stability studies of verbascoside, a novel antioxidant, in dermo-cosmetic and pharmaceutical topical formulations. Molecules 2011, 16, 7068–7080. [Google Scholar] [CrossRef]
- Lopes, L.B. Overcoming the cutaneous barrier with microemulsions. Pharmaceutics 2014, 6, 52–77. [Google Scholar] [CrossRef] [PubMed]
- Alberti, I.; Kalia, Y.N.; Naik, A.; Bonny, J.; Guy, R.H. Effect of ethanol and isopropyl myristate on the availability of topical terbinafine in human stratum corneum, in vivo. Int. J. Pharm. 2001, 219, 11–19. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; Yang, F.; Huang, Y.; Liu, S.; Hu, Z.; Liao, S.; Li, Y. Enhanced In Vitro Efficacy of Verbascoside in Suppressing Hepatic Stellate Cell Activation via ROS Scavenging with Reverse Microemulsion. Antioxidants 2024, 13, 907. https://doi.org/10.3390/antiox13080907
Xiao X, Yang F, Huang Y, Liu S, Hu Z, Liao S, Li Y. Enhanced In Vitro Efficacy of Verbascoside in Suppressing Hepatic Stellate Cell Activation via ROS Scavenging with Reverse Microemulsion. Antioxidants. 2024; 13(8):907. https://doi.org/10.3390/antiox13080907
Chicago/Turabian StyleXiao, Xiao, Feiyu Yang, Yuling Huang, Shaohui Liu, Zhenhua Hu, Shanggao Liao, and Yuanyuan Li. 2024. "Enhanced In Vitro Efficacy of Verbascoside in Suppressing Hepatic Stellate Cell Activation via ROS Scavenging with Reverse Microemulsion" Antioxidants 13, no. 8: 907. https://doi.org/10.3390/antiox13080907
APA StyleXiao, X., Yang, F., Huang, Y., Liu, S., Hu, Z., Liao, S., & Li, Y. (2024). Enhanced In Vitro Efficacy of Verbascoside in Suppressing Hepatic Stellate Cell Activation via ROS Scavenging with Reverse Microemulsion. Antioxidants, 13(8), 907. https://doi.org/10.3390/antiox13080907