


The Antioxidant PAPLAL Protects against Allergic Contact Dermatitis in Experimental Models


Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Mice
2.3. Induction of ACD and Treatment of PAPLAL
2.4. 8-Isoprostane Contents
2.5. Histological Analyses
2.6. Quantitative Polymerase Chain Reaction (qPCR)
2.7. Treatment with PAPLAL in a Human Epidermal Skin Model
2.8. Statistical Analyses
3. Results
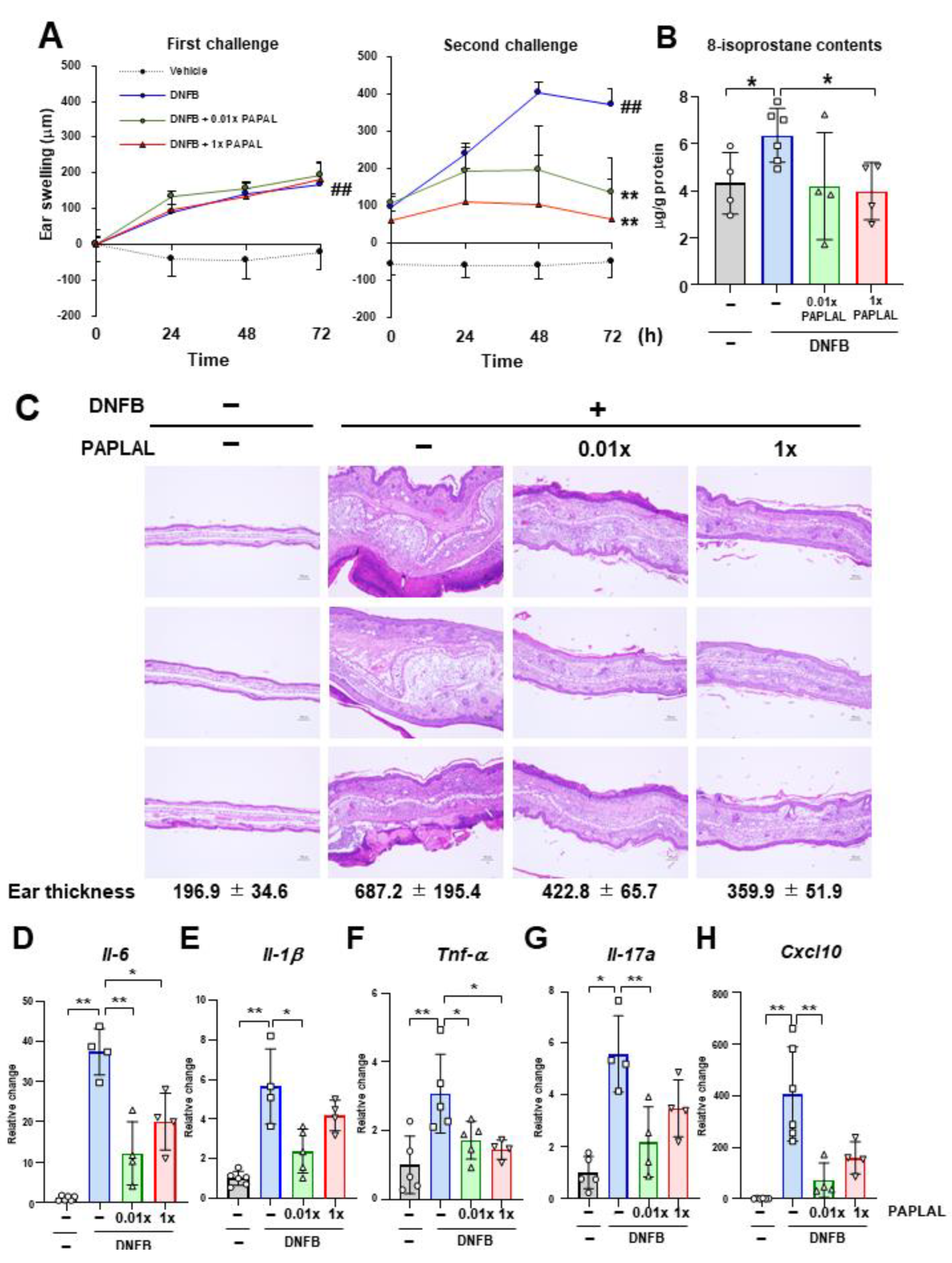
3.1. PAPLAL Improved Skin Inflammation in DNFB-Induced ACD
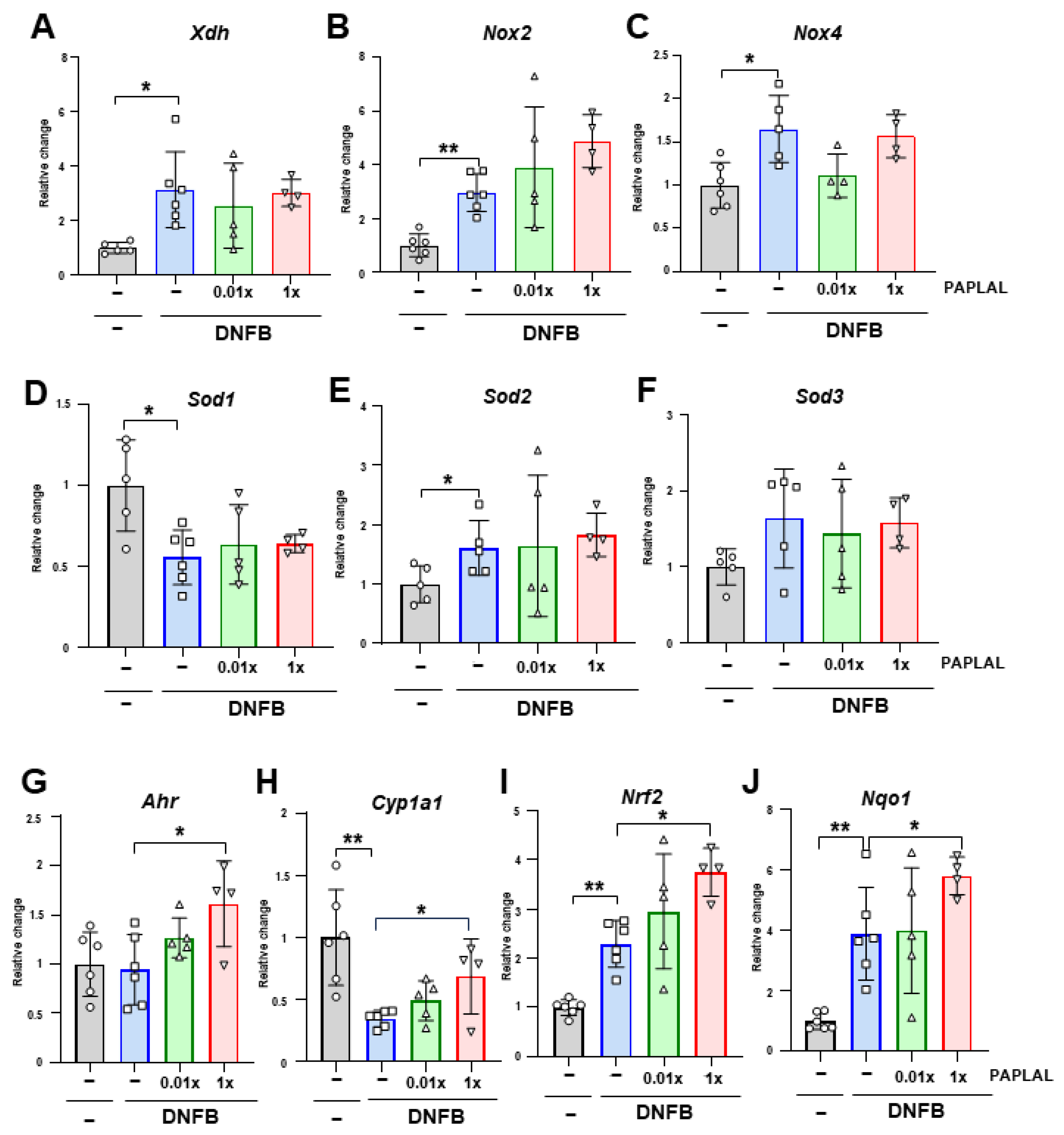
3.2. PAPLAL Did Not Change Redox-Related Gene Expression in the ACD Model
3.3. PAPLAL Increased the Expression of the Stress Response-Related Genes Ahr and Nrf2
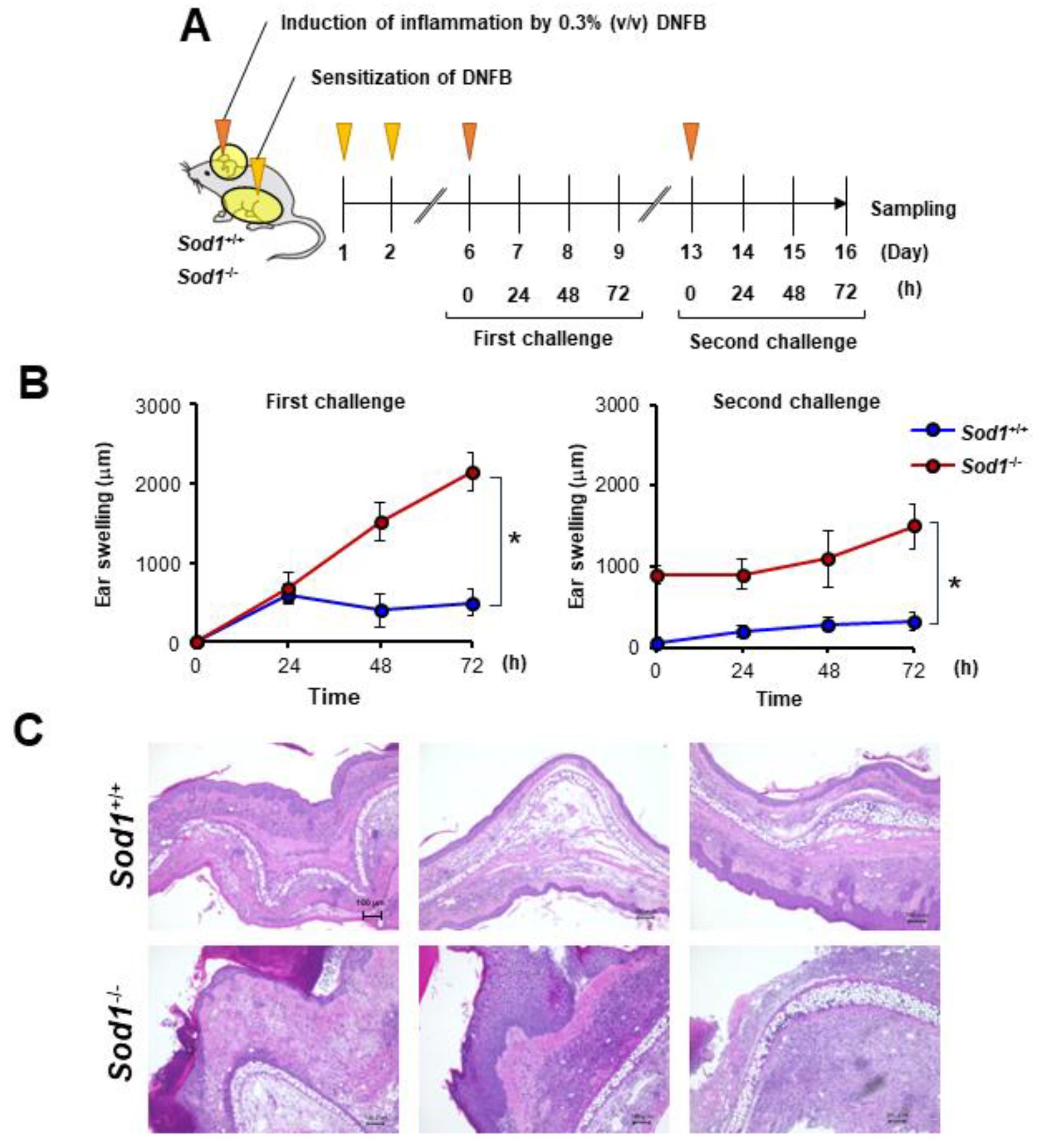
3.4. Sod1 Deficiency Exacerbated DNFB-Induced ACD
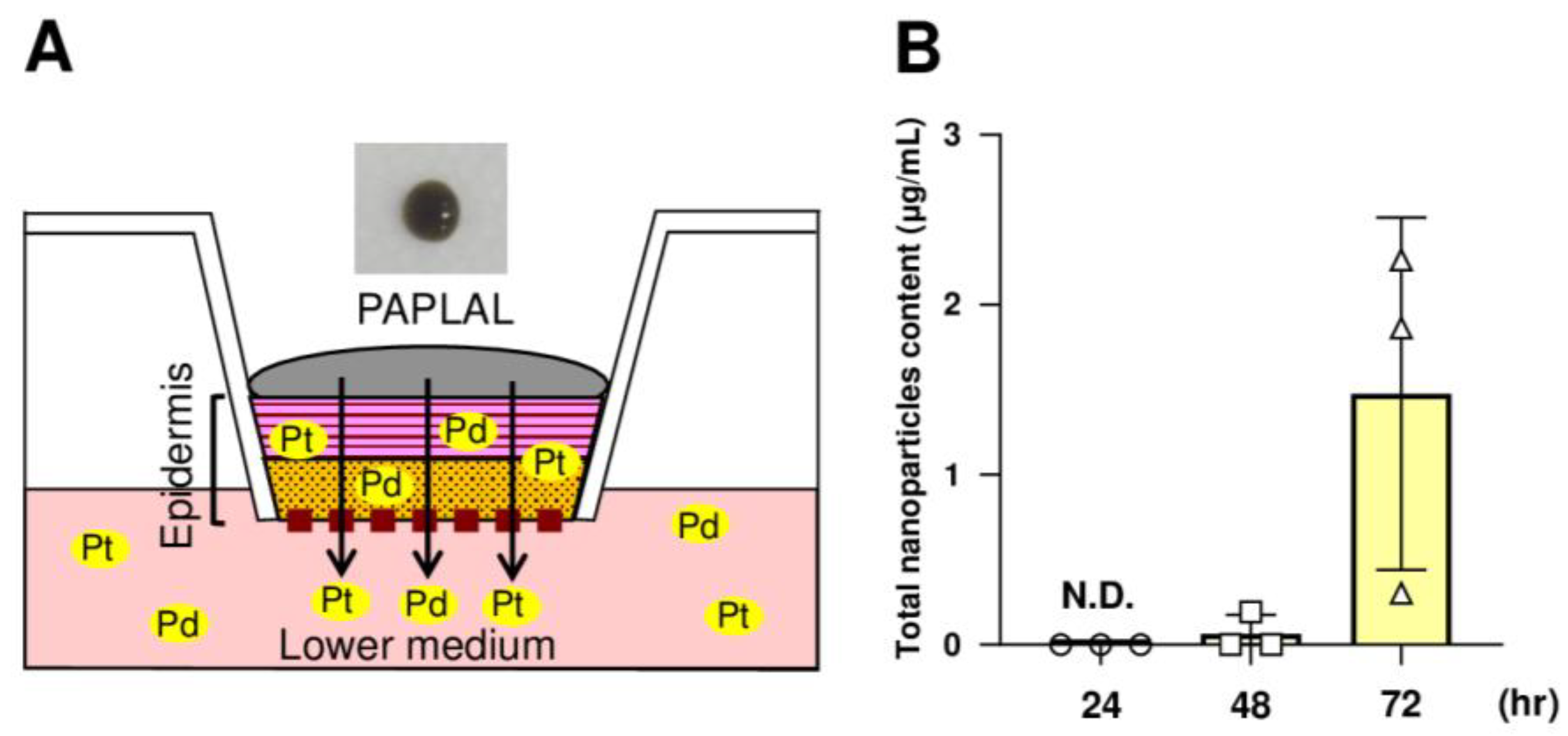
3.5. nPt and nPd Contained in PAPLAL Penetrated the Epidermal Layer in a Human Epidermal Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shibuya, S.; Watanabe, K.; Yokote, K.; Shimizu, T. Platinum and palladium nanoparticles regulate the redox balance and protect against age-related skin changes in mice. In Textbook of Aging Skin; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–11. [Google Scholar]
- Shibuya, S.; Ozawa, Y.; Watanabe, K.; Izuo, N.; Toda, T.; Yokote, K.; Shimizu, T. Palladium and platinum nanoparticles attenuate aging-like skin atrophy via antioxidant activity in mice. PLoS ONE 2014, 9, e109288. [Google Scholar] [CrossRef]
- Shibata, T.; Yoshikawa, R.; Ichihashi, M. The novel therapy for vitiligo vulgaris: Topical use of cosmetic cream of platinum nanoparticles and palladium nanoparticles which show strong catalase-like activity. J. Pigment. Disord. 2015, 2, 1000184. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Yoshida, M.; Ando, H.; Ichihashi, M. Establishment of photoaging in vitro by repetitive UVA irradiation: Induction of characteristic markers of senescence and its prevention by PAPLAL with potent catalase activity. Photochem. Photobiol. 2018, 94, 438–444. [Google Scholar] [CrossRef]
- Shibuya, S.; Watanabe, K.; Tsuji, G.; Ichihashi, M.; Shimizu, T. Platinum and palladium nanoparticle-containing mixture, PAPLAL, does not induce palladium allergy. Exp. Dermatol. 2019, 28, 1025–1028. [Google Scholar] [CrossRef]
- Coenraads, P.J.; Goncalo, M. Skin diseases with high public health impact. Contact dermatitis. Eur. J. Dermatol. 2007, 17, 564–565. [Google Scholar]
- Park, D.K.; Lee, Y.G.; Park, H.J. Extract of rhus verniciflua bark suppresses 2,4-dinitrofluorobenzene-induced allergic contact dermatitis. Evid. Based Complement. Alternat. Med. 2013, 2013, 879696. [Google Scholar] [CrossRef]
- Kwon, H.K.; Song, M.J.; Lee, H.J.; Park, T.S.; Kim, M.I.; Park, H.J. Pediococcus pentosaceus-Fermented Cordyceps militaris Inhibits Inflammatory Reactions and Alleviates Contact Dermatitis. Int. J. Mol. Sci. 2018, 19, 3504. [Google Scholar] [CrossRef]
- Johansson, S.G.; Bieber, T.; Dahl, R.; Friedmann, P.S.; Lanier, B.Q.; Lockey, R.F.; Motala, C.; Ortega Martell, J.A.; Platts-Mills, T.A.; Ring, J.; et al. Revised nomenclature for allergy for global use: Report of the Nomenclature Review Committee of the World Allergy Organization, October 2003. J. Allergy Clin. Immunol. 2004, 113, 832–836. [Google Scholar] [CrossRef]
- Mallabo, M.R.B.; Corpuz, M.J.T.; Salonga, R.B.; Vasquez, R.D. Inhibitory effect of sulfated polysaccharide from codium edule P.C. silva against 2,4-dinitrofluorobenzene (DNFB)-induced allergic contact dermatitis on female BALB/c mice. Adv. Pharm. Bull. 2022, 12, 410–418. [Google Scholar] [CrossRef]
- Welsh, E.; Goldenberg, A.; Welsh, O.; Jacob, S. Contact Dermatitis: Therapeutics when avoidance fails. J. Allergy Ther. 2014, 5, 1–4. [Google Scholar] [CrossRef]
- Murakami, K.; Inagaki, J.; Saito, M.; Ikeda, Y.; Tsuda, C.; Noda, Y.; Kawakami, S.; Shirasawa, T.; Shimizu, T. Skin atrophy in cytoplasmic SOD-deficient mice and its complete recovery using a vitamin C derivative. Biochem. Biophys. Res. Commun. 2009, 382, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, G.; Hashimoto-Hachiya, A.; Takemura, M.; Kanemaru, T.; Ichihashi, M.; Furue, M. Palladium and platinum nanoparticles activate AHR and NRF2 in human keratinocytes-implications in vitiligo therapy. J. Investig. Dermatol. 2017, 137, 1582–1586. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, S.; Ozawa, Y.; Toda, T.; Watanabe, K.; Tometsuka, C.; Ogura, T.; Koyama, Y.; Shimizu, T. Collagen peptide and vitamin C additively attenuate age-related skin atrophy in Sod1-deficient mice. Biosci. Biotechnol. Biochem. 2014, 78, 1212–1220. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, S.; Watanabe, K.; Ozawa, Y.; Shimizu, T. Xanthine oxidoreductase-mediated superoxide production is not involved in the age-related pathologies in Sod1-deficient mice. Int. J. Mol. Sci. 2021, 22, 3542. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ahn, Y.T.; Lee, G.S.; Cho, S.I.; Kim, J.M.; Lee, C.; Lim, B.K.; Ju, S.A.; An, W.G. Effects of astaxanthin on dinitrofluorobenzene-induced contact dermatitis in mice. Mol. Med. Rep. 2015, 12, 3632–3638. [Google Scholar] [CrossRef] [PubMed]
- Senaldi, G.; Pointaire, P.; Piguet, P.F.; Grau, G.E. Protective effect of N-acetylcysteine in hapten-induced irritant and contact hypersensitivity reactions. J. Investig. Dermatol. 1994, 102, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Ogino, H.; Sakazaki, F.; Okuno, T.; Arakawa, T.; Ueno, H. Oxidized dietary oils enhance immediate- and/or delayed-type allergic reactions in BALB/c mice. Allergol. Int. 2015, 64, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Li, X.K. Oxidative stress in atopic dermatitis. Oxid. Med. Cell. Longev. 2016, 2016, 2721469. [Google Scholar] [CrossRef] [PubMed]
- Eto, H.; Tsuji, G.; Chiba, T.; Furue, M.; Hyodo, F. Non-invasive evaluation of atopic dermatitis based on redox status using in vivo dynamic nuclear polarization magnetic resonance imaging. Free Radic. Biol. Med. 2017, 103, 209–215. [Google Scholar] [CrossRef]
- Hirakawa, S.; Saito, R.; Ohara, H.; Okuyama, R.; Aiba, S. Dual oxidase 1 induced by Th2 cytokines promotes STAT6 phosphorylation via oxidative inactivation of protein tyrosine phosphatase 1B in human epidermal keratinocytes. J. Immunol. 2011, 186, 4762–4770. [Google Scholar] [CrossRef]
- Ferreira, I.; Silva, A.; Martins, J.D.; Neves, B.M.; Cruz, M.T. Nature and kinetics of redox imbalance triggered by respiratory and skin chemical sensitizers on the human monocytic cell line THP-1. Redox Biol. 2018, 16, 75–86. [Google Scholar] [CrossRef]
- Kim, J.; Toda, T.; Watanabe, K.; Shibuya, S.; Ozawa, Y.; Izuo, N.; Cho, S.; Seo, D.B.; Yokote, K.; Shimizu, T. Syringaresinol reverses age-related skin atrophy by suppressing FoxO3a-mediated matrix metalloproteinase-2 activation in copper/zinc superoxide dismutase-deficient mice. J. Investig. Dermatol. 2019, 139, 648–655. [Google Scholar] [CrossRef]
- Helou, D.G.; Noel, B.; Gaudin, F.; Groux, H.; El Ali, Z.; Pallardy, M.; Chollet-Martin, S.; Kerdine-Romer, S. Cutting edge: Nrf2 regulates neutrophil recruitment and accumulation in skin during contact hypersensitivity. J. Immunol. 2019, 202, 2189–2194. [Google Scholar] [CrossRef]
- Furue, M.; Ulzii, D.; Vu, Y.H.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T. Pathogenesis of atopic dermatitis: Current paradigm. Iran. J. Immunol. 2019, 16, 97–107. [Google Scholar]
- Major, J.; Crotta, S.; Finsterbusch, K.; Chakravarty, P.; Shah, K.; Frederico, B.; D’Antuono, R.; Green, M.; Meader, L.; Suarez-Bonnet, A.; et al. Endothelial AHR activity prevents lung barrier disruption in viral infection. Nature 2023, 621, 813–820. [Google Scholar] [CrossRef]
- Wiggins, B.G.; Wang, Y.F.; Burke, A.; Grunberg, N.; Vlachaki Walker, J.M.; Dore, M.; Chahrour, C.; Pennycook, B.R.; Sanchez-Garrido, J.; Vernia, S.; et al. Endothelial sensing of AHR ligands regulates intestinal homeostasis. Nature 2023, 621, 821–829. [Google Scholar] [CrossRef]
- Furue, M. Regulation of Filaggrin, Loricrin, and Involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic implications in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef]
- Kennedy, L.H.; Sutter, C.H.; Leon Carrion, S.; Tran, Q.T.; Bodreddigari, S.; Kensicki, E.; Mohney, R.P.; Sutter, T.R. 2,3,7,8-Tetrachlorodibenzo-p-dioxin-mediated production of reactive oxygen species is an essential step in the mechanism of action to accelerate human keratinocyte differentiation. Toxicol. Sci. 2013, 132, 235–249. [Google Scholar] [CrossRef]
- van den Bogaard, E.H.; Podolsky, M.A.; Smits, J.P.; Cui, X.; John, C.; Gowda, K.; Desai, D.; Amin, S.G.; Schalkwijk, J.; Perdew, G.H.; et al. Genetic and pharmacological analysis identifies a physiological role for the AHR in epidermal differentiation. J. Investig. Dermatol. 2015, 135, 1320–1328. [Google Scholar] [CrossRef]
- Furue, M.; Tsuji, G.; Mitoma, C.; Nakahara, T.; Chiba, T.; Morino-Koga, S.; Uchi, H. Gene regulation of filaggrin and other skin barrier proteins via aryl hydrocarbon receptor. J. Dermatol. Sci. 2015, 80, 83–88. [Google Scholar] [CrossRef]
- Bruhs, A.; Haarmann-Stemmann, T.; Frauenstein, K.; Krutmann, J.; Schwarz, T.; Schwarz, A. Activation of the arylhydrocarbon receptor causes immunosuppression primarily by modulating dendritic cells. J. Investig. Dermatol. 2015, 135, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Haas, K.; Weighardt, H.; Deenen, R.; Kohrer, K.; Clausen, B.; Zahner, S.; Boukamp, P.; Bloch, W.; Krutmann, J.; Esser, C. Aryl hydrocarbon receptor in keratinocytes is essential for murine skin barrier integrity. J. Investig. Dermatol. 2016, 136, 2260–2269. [Google Scholar] [CrossRef] [PubMed]
- Kiyomatsu-Oda, M.; Uchi, H.; Morino-Koga, S.; Furue, M. Protective role of 6-formylindolo[3,2-b]carbazole (FICZ), an endogenous ligand for arylhydrocarbon receptor, in chronic mite-induced dermatitis. J. Dermatol. Sci. 2018, 90, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Navid, F.; Bruhs, A.; Schuller, W.; Fritsche, E.; Krutmann, J.; Schwarz, T.; Schwarz, A. The Aryl hydrocarbon receptor is involved in UVR-induced immunosuppression. J. Investig. Dermatol. 2013, 133, 2763–2770. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.H.; Jayawickreme, C.; Rickard, D.J.; Nicodeme, E.; Bui, T.; Simmons, C.; Coquery, C.M.; Neil, J.; Pryor, W.M.; Mayhew, D.; et al. Tapinarof is a natural AhR agonist that resolves skin inflammation in mice and humans. J. Investig. Dermatol. 2017, 137, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Uchi, H.; Takahara, M.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T.; Furue, M. Antioxidant soybean tar Glyteer rescues T-helper-mediated downregulation of filaggrin expression via aryl hydrocarbon receptor. J. Dermatol. 2015, 42, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, G.; Hashimoto-Hachiya, A.; Kiyomatsu-Oda, M.; Takemura, M.; Ohno, F.; Ito, T.; Morino-Koga, S.; Mitoma, C.; Nakahara, T.; Uchi, H.; et al. Aryl hydrocarbon receptor activation restores filaggrin expression via OVOL1 in atopic dermatitis. Cell Death. Dis. 2017, 8, e2931. [Google Scholar] [CrossRef] [PubMed]
- van den Bogaard, E.H.; Bergboer, J.G.; Vonk-Bergers, M.; van Vlijmen-Willems, I.M.; Hato, S.V.; van der Valk, P.G.; Schroder, J.M.; Joosten, I.; Zeeuwen, P.L.; Schalkwijk, J. Coal tar induces AHR-dependent skin barrier repair in atopic dermatitis. J. Clin. Investig. 2013, 123, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef]
- Jeong, H.; Shin, J.Y.; Kim, M.J.; Na, J.; Ju, B.G. Activation of aryl hydrocarbon receptor negatively regulates thymic stromal lymphopoietin gene expression via protein kinase cdelta-p300-NF-kappaB pathway in keratinocytes under inflammatory conditions. J. Investig. Dermatol. 2019, 139, 1098–1109. [Google Scholar] [CrossRef]
- Okamoto, H.; Horii, K.; Fujisawa, A.; Yamamoto, Y. Oxidative deterioration of platinum nanoparticle and its prevention by palladium. Exp. Dermatol. 2012, 21 (Suppl. S1), 5–7. [Google Scholar] [CrossRef] [PubMed]
- Larese Filon, F.; Mauro, M.; Adami, G.; Bovenzi, M.; Crosera, M. Nanoparticles skin absorption: New aspects for a safety profile evaluation. Regul. Toxicol. Pharmacol. 2015, 72, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Rose, L.; Schneider, C.; Stock, C.; Zollner, T.M.; Docke, W.D. Extended DNFB-induced contact hypersensitivity models display characteristics of chronic inflammatory dermatoses. Exp. Dermatol. 2012, 21, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Sultana, N.; Hayashi, I.; Fukamachi, M.; Sugiyama, M. Exopolysaccharide produced by Lactobacillus paracasei IJH-SONE68 prevents and improves the picryl chloride-induced contact dermatitis. Molecules 2019, 24, 2970. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.; Philippsen, R.; Schwarz, T. Mouse models of allergic contact dermatitis: Practical aspects. J. Investig. Dermatol. 2023, 143, 888–892. [Google Scholar] [CrossRef]
- Manabe, Y.; Yoshimura, M.; Sakamaki, K.; Inoue, A.; Kakinoki, A.; Hokari, S.; Sakanaka, M.; Aoki, J.; Miyachi, H.; Furuta, K.; et al. 1-Fluoro-2,4-dinitrobenzene and its derivatives act as secretagogues on rodent mast cells. Eur. J. Immunol. 2017, 47, 60–67. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Forward | Reverse |
|---|---|---|---|
| Ahr | NM_013464 | CTGGTTGTCACAGCAGATGCCT | CGGTCTTCTGTATGGATGAGCTC |
| Cxcl10 | NM_021274 | CACCATGAACCCAAGTGCTG | GGATAGGCTCGCAGGGATGA |
| Cyp1a1 | NM_001136059 | CATCACAGACAGCCTCATTGAGC | CTCCACGAGATAGCAGTTGTGAC |
| Il-1β | NM_008361 | ATGGCAACTGTTCCTGAACTCAACT | CAGGACAGGTATAGATTCTTTCCTTT |
| Il-6 | NM_031168 | GCTACCAAACGTGATATAATCAGGA | CCAGGTAGCTATGGTACTCCAGAA |
| Il-17a | NM_010552 | CAGACTACCTCAACCGTTCCAC | TCCAGCTTTCCCTCCGCATTGA |
| Nox2 | NM_007807 | TGGCGATCTCAGCAAAAGGTGG | GTACTGTCCCACCTCCATCTTG |
| Nox4 | NM_015760 | CGGGATTTGCTACTGCCTCCAT | GTGACTCCTCAAATGGGCTTCC |
| Nrf2 | NM_010902 | TTTTCCATTCCCGAATTACAGT | AGGAGATCGATGAGTAAAAATGGT |
| Nqo1 | NM_008706 | GCCGAACACAAGAAGCTGGAAG | GGCAAATCCTGCTACGAGCACT |
| Rps14 | NM_020600 | AGGAGTCTGGAGACGACGAT | CAGTCACTCGGCAGATGGTT |
| Sod1 | NM_011434 | AACCATCCACTTCGAGCAGAA | GCTGGCCTTCAGTTAATCCTGTA |
| Sod2 | NM_013671 | CTGGACAAACCTGAGCCCTAAG | AAGACCCAA AGTCACGCTTGA |
| Sod3 | NM_011435 | CTCTTGGGAGAGCCTGACA | GCCAGTAGCAAGCCGTAGAA |
| Tnf-α | NM_013693 | ATGAGCACAGAAAGCATGATCCGC | GCTTGGTGGTTTGCTACGAC |
| Xdh | NW_011723 | GCTCTTCGTGAGCACACAGAAC | CCACCCATTCTTTTCACTCGGAC |
| nPt | nPd | |||
|---|---|---|---|---|
| Migration | (μg) | (%) | (μg) | (%) |
| Epidermal tissue | 2.70 ± 0.56 | 6.77 ± 1.42 | 8.33 ± 0.58 | 11.87 ± 2.48 |
| Lower medium | 0.04 ± 0.06 | 0.28 ± 0.16 | 0.35 ± 0.13 | 0.59 ± 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibuya, S.; Watanabe, K.; Shimizu, T. The Antioxidant PAPLAL Protects against Allergic Contact Dermatitis in Experimental Models. Antioxidants 2024, 13, 748. https://doi.org/10.3390/antiox13060748
Shibuya S, Watanabe K, Shimizu T. The Antioxidant PAPLAL Protects against Allergic Contact Dermatitis in Experimental Models. Antioxidants. 2024; 13(6):748. https://doi.org/10.3390/antiox13060748
Chicago/Turabian StyleShibuya, Shuichi, Kenji Watanabe, and Takahiko Shimizu. 2024. "The Antioxidant PAPLAL Protects against Allergic Contact Dermatitis in Experimental Models" Antioxidants 13, no. 6: 748. https://doi.org/10.3390/antiox13060748
APA StyleShibuya, S., Watanabe, K., & Shimizu, T. (2024). The Antioxidant PAPLAL Protects against Allergic Contact Dermatitis in Experimental Models. Antioxidants, 13(6), 748. https://doi.org/10.3390/antiox13060748