
Health Benefits of Antioxidant Bioactive Compounds in Ginger (Zingiber officinale) Leaves by Network Pharmacology Analysis Combined with Experimental Validation


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Proximate Components and Minerals
2.3. Amino Acids
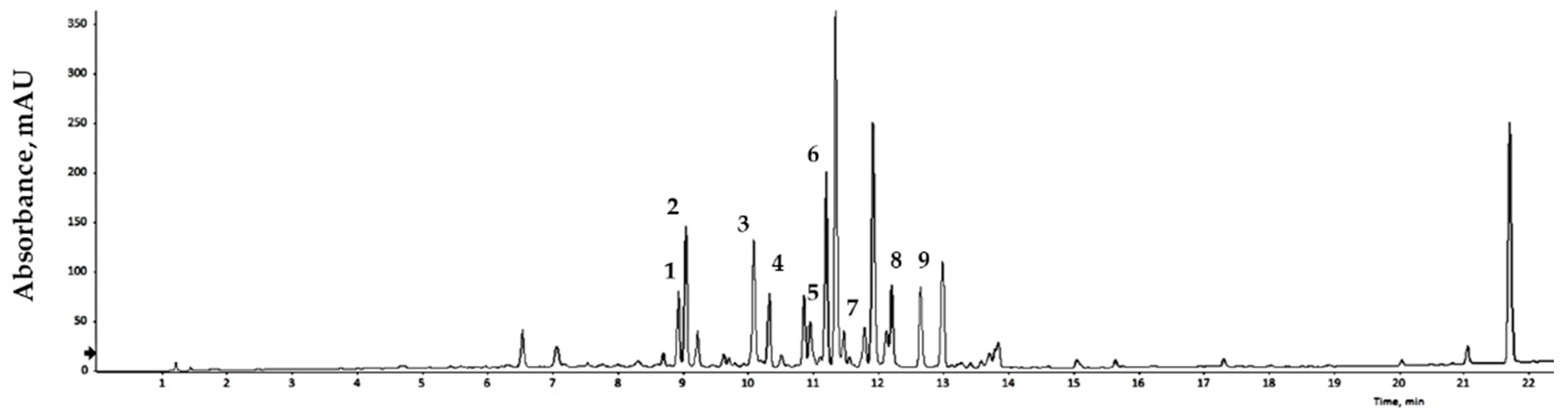
2.4. Identification of Phytochemicals Using UPLC-DAD-QTOF/MS
2.5. Antioxidant Properties and Radical Scavenging Activity
2.6. Antioxidant-Related Target Screening
2.7. Protein-Protein Interactions (PPI)
2.8. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment
2.9. Validation of Prognostic Power of Key Genes
2.10. Molecular Docking
3. Results and Discussion
3.1. Proximate Composition and Mineral Content of GLs
3.2. Amino Acid Contents of GLs
3.3. Flavonoid Identification Compound Content, Total Phenolic Content, Total Flavonoid Content, and RSA of GLs
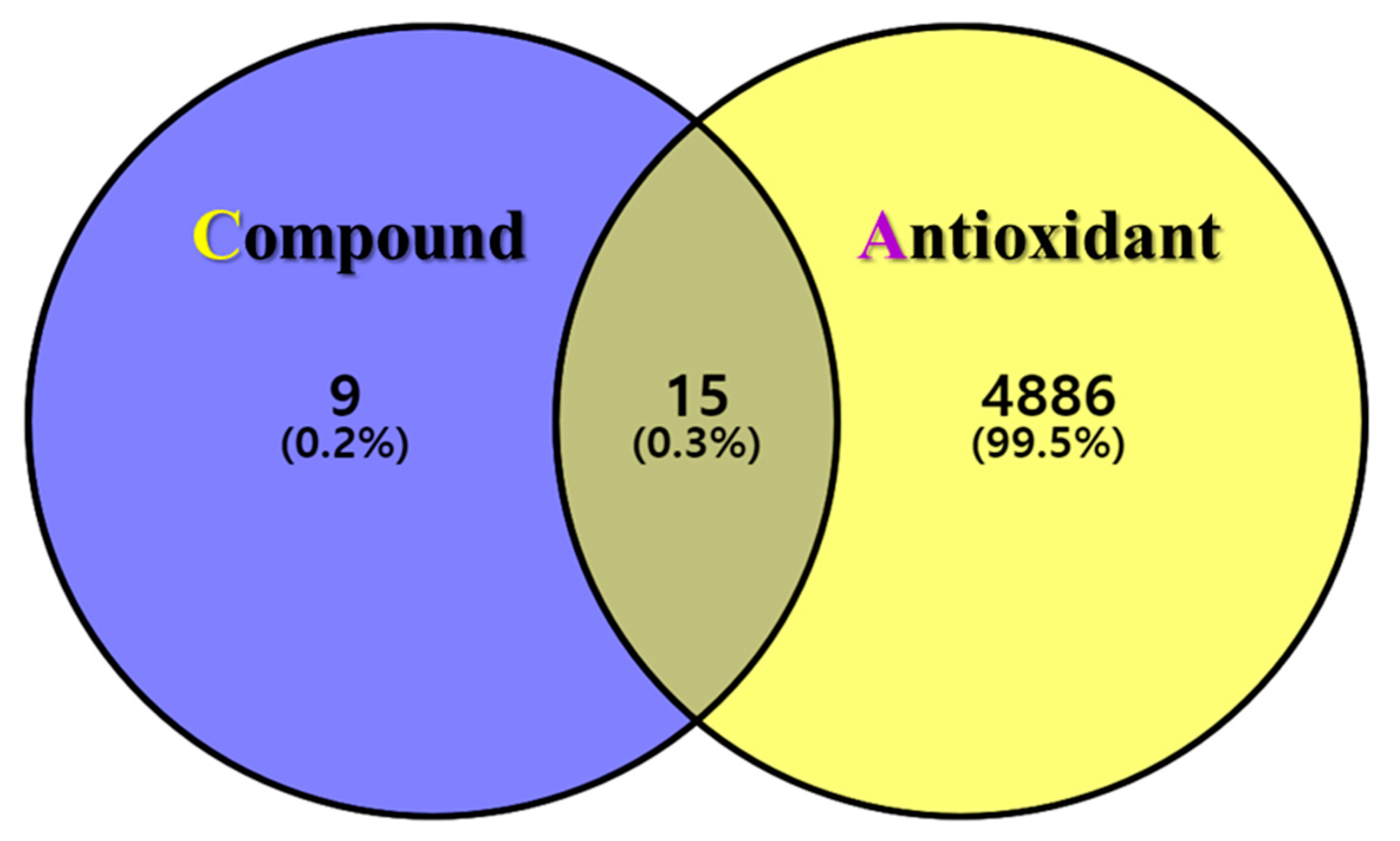
3.4. Antioxidant-Related Effective Target Prediction
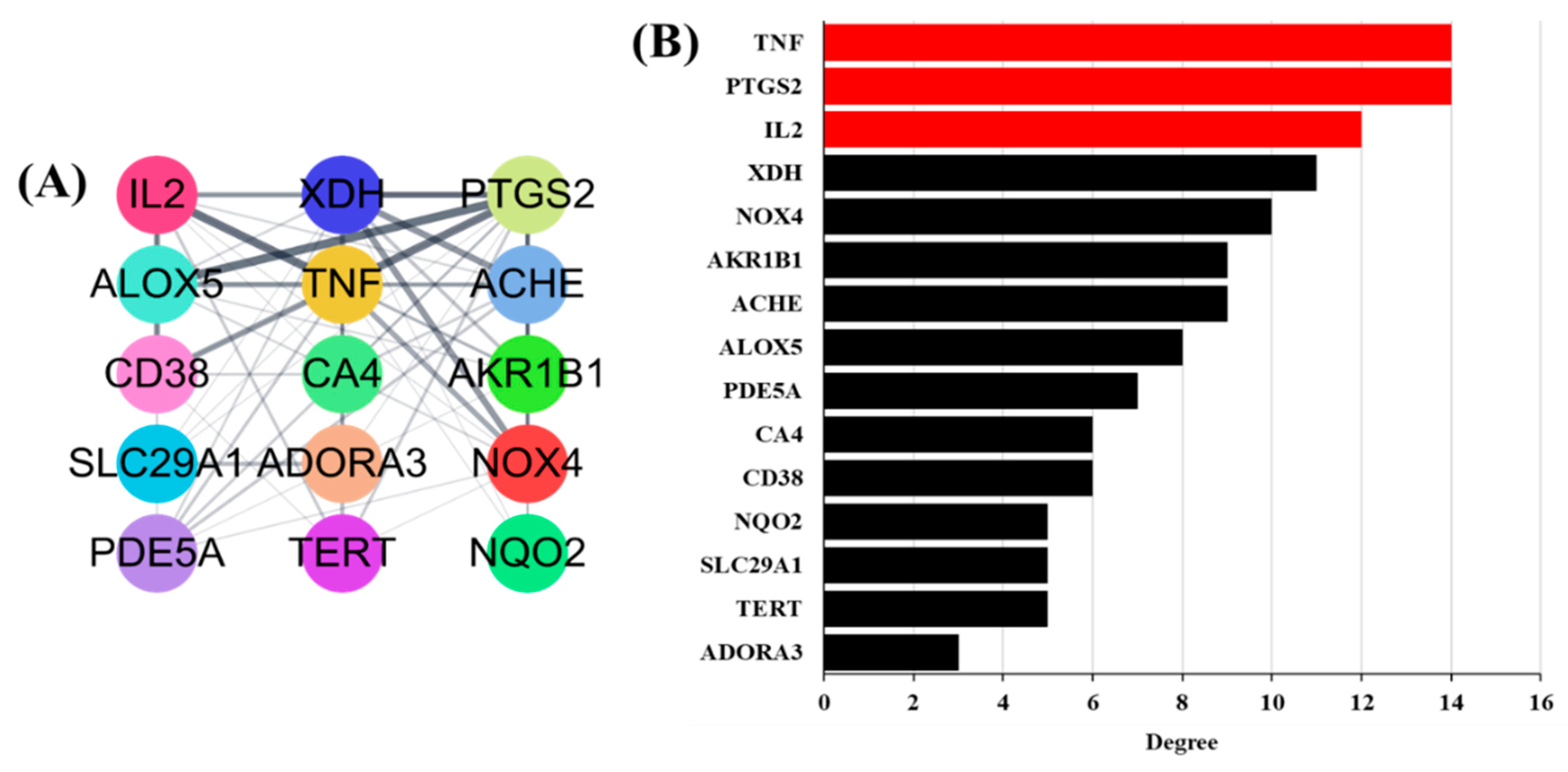
3.5. Antioxidant Target PPI Network of GLs
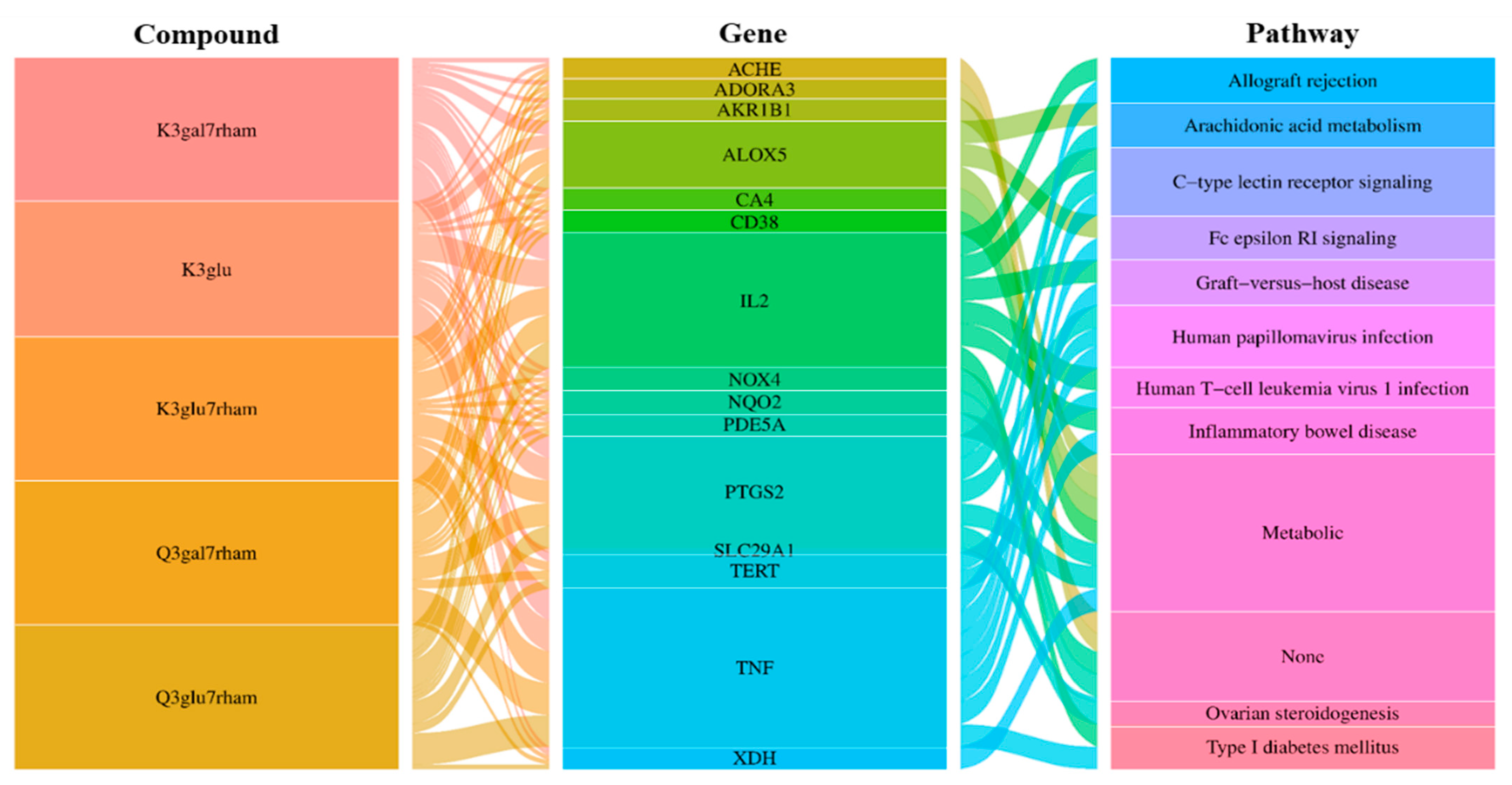
3.6. Build an Integrated Network
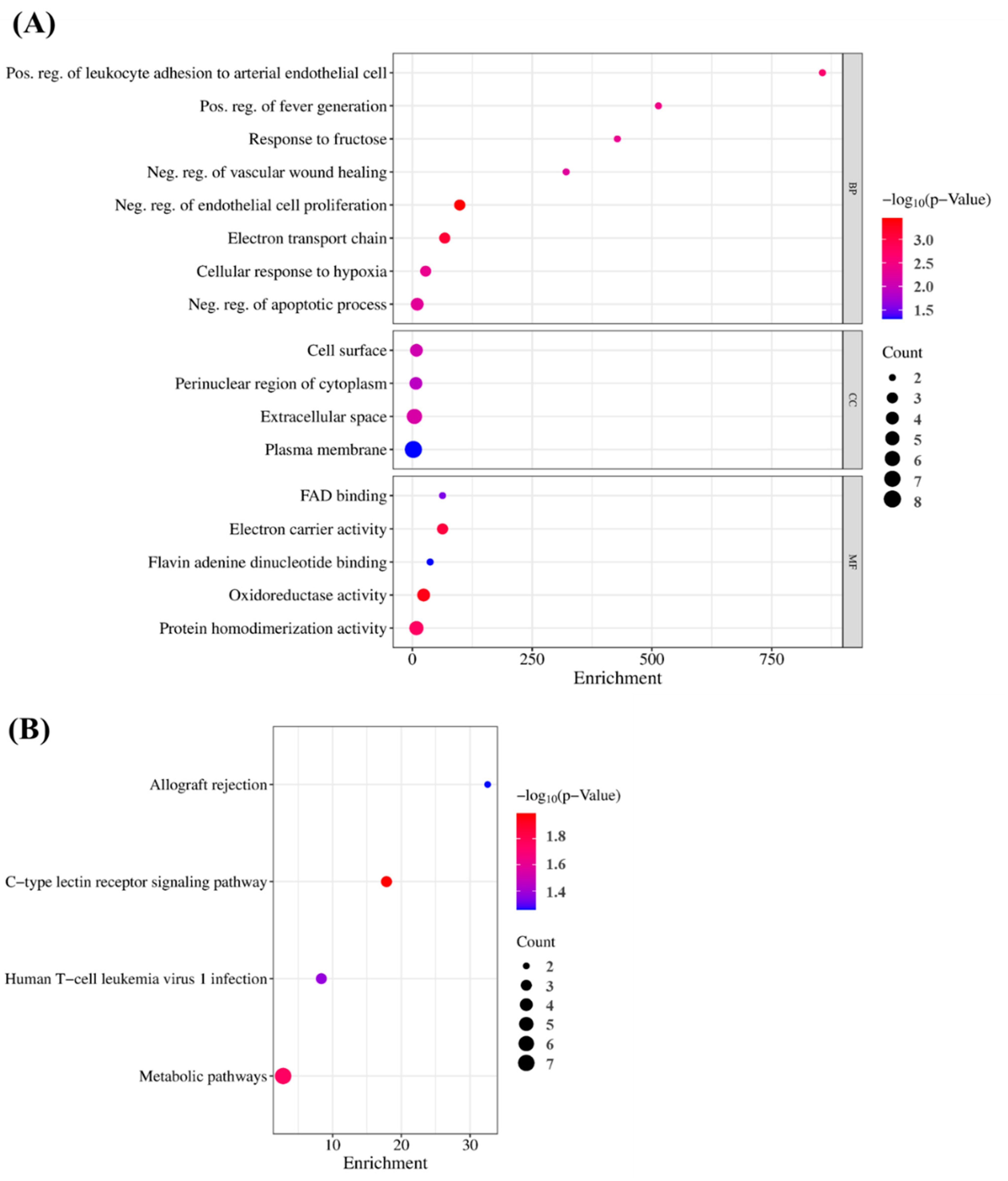
3.7. GO and KEGG Pathway Enrichment Analyses
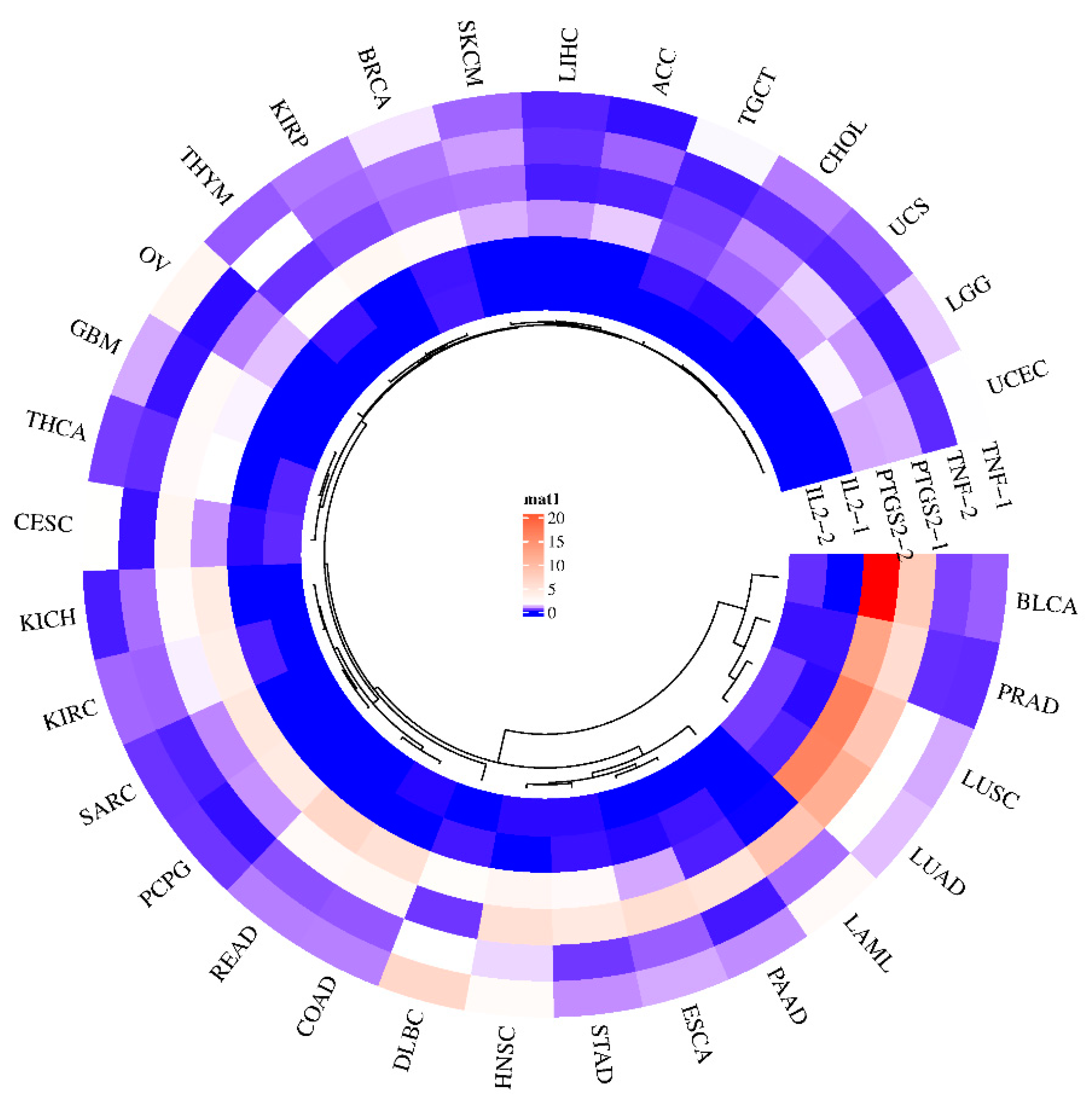
3.8. Predicting Gene Expression of GLs Target Genes in Various Cancer Types
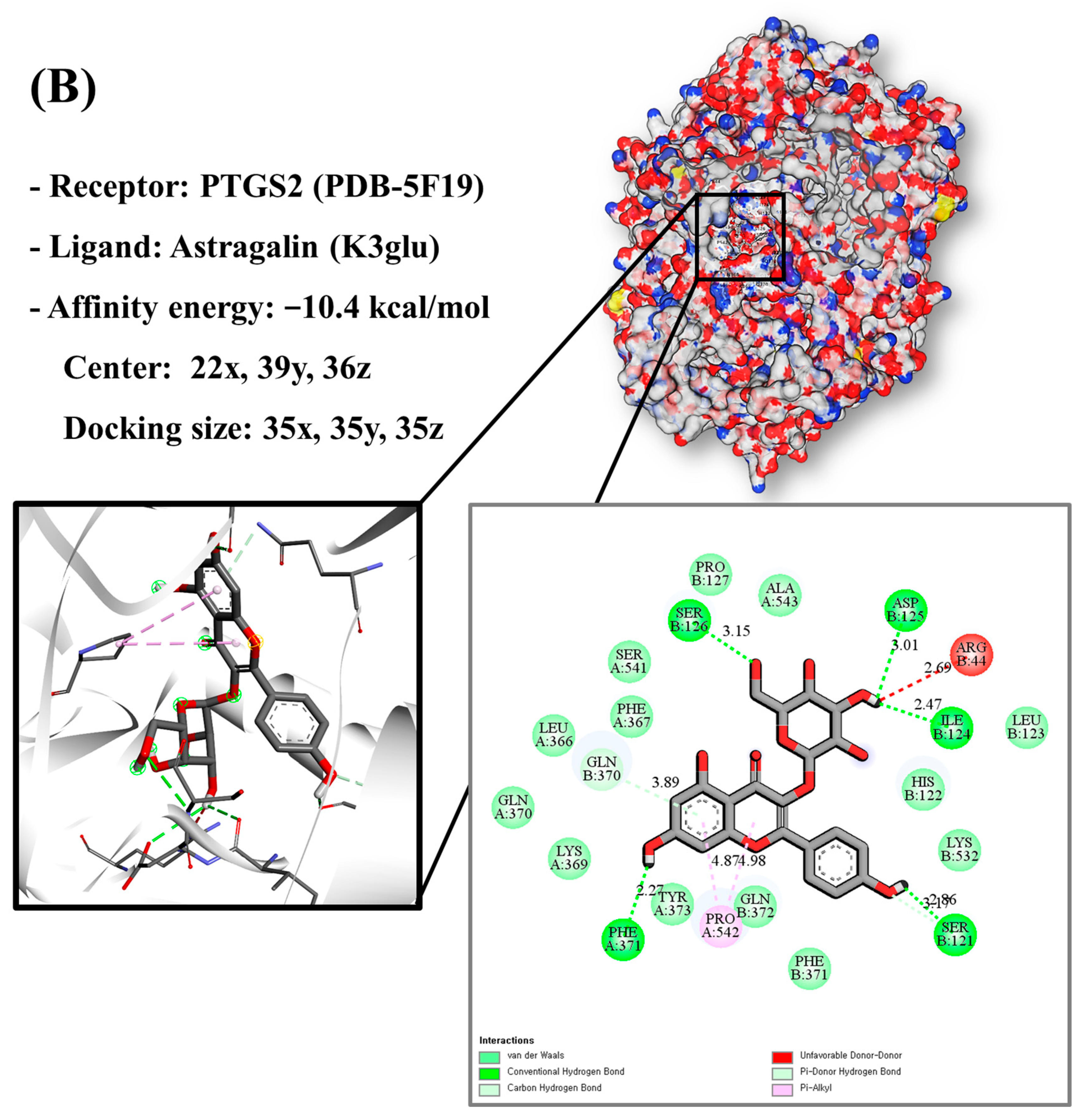
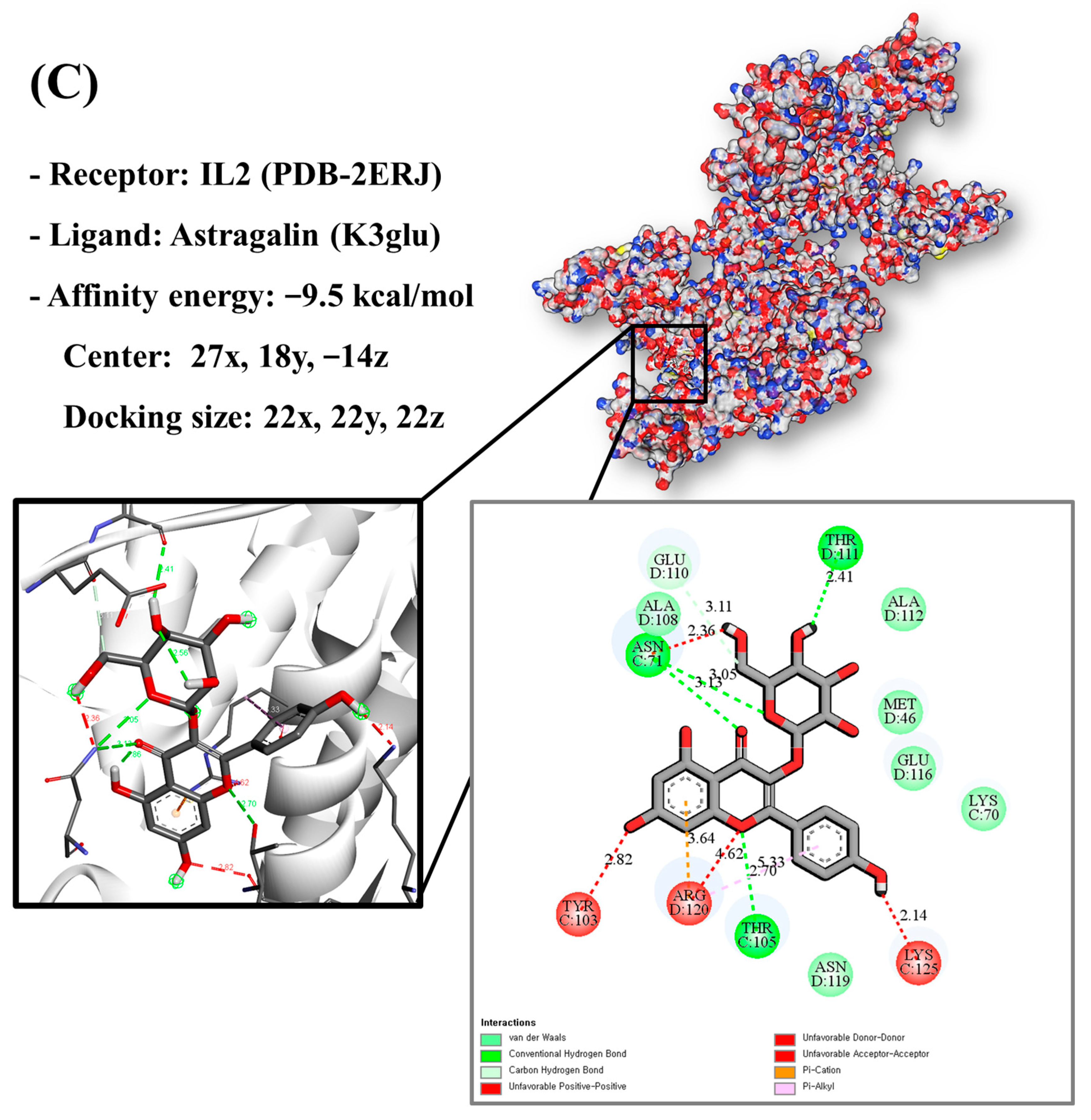
3.9. Molecular Docking of Bioactive Compounds and Key Targets
3.10. Potential Antioxidant Pathway
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, K.; Gaitsch, H.; Poon, H.; Cox, N.J.; Rzhetsky, A. Classification of common human diseases derived from shared genetic and environmental determinants. Nat. Genet. 2017, 49, 1319–1325. [Google Scholar] [CrossRef]
- Choudhury, S.P.; Arora, A.; Jain, N.; Dey, S.K. Climate change, urbanisation, and their impact on increased occurrence of cardiometabolic syndrome. Urban Ecol. Glob. Clim. Chang. 2022, 2022, 30–56. [Google Scholar] [CrossRef]
- Ilacqua, A.; Izzo, G.; Emerenziani, G.P.; Baldari, C.; Aversa, A. Lifestyle and fertility: The influence of stress and quality of life on male fertility. Reprod. Biol. Endocrinol. 2018, 16, 115. [Google Scholar] [CrossRef]
- Dong, G.; Qu, L.; Gong, X.; Pang, B.; Yan, W.; Wei, J. Effect of social factors and the natural environment on the etiology and pathogenesis of diabetes mellitus. Int. J. Endocrinol. 2019, 2019, 8749291. [Google Scholar] [CrossRef]
- Woods, J.A.; Hutchinson, N.T.; Powers, S.K.; Roberts, W.O.; Gomez-Cabrera, M.C.; Radak, Z.; Berkes, I.; Boros, A.; Boldogh, I.; Leeuwenburgh, C.; et al. The COVID-19 pandemic and physical activity. Sports Med. Health Sci. 2020, 2, 55–64. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Cianca, S.I.; Romero-Castillo, C.; Gálvez-Romero, J.L.; Juárez, Z.N.; Hernández, L.R. Antioxidant and anti-inflammatory compounds from edible plants with anti-cancer activity and their potential use as drugs. Molecules 2023, 28, 1488. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, F.; Capasso, R.; Aviello, G.; Pittler, M.H.; Izzo, A.A. Effectiveness and safety of ginger in the treatment of pregnancy-induced nausea and vomiting. Obstet. Gynecol. 2015, 105, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Barnard, N.D.; Katcher, H.I.; Jenkins, D.J.; Cohen, J.; Turner-McGrievy, G. Vegetarian and vegan diets in type 2 diabetes management. Nutr. Rev. 2009, 67, 255–263. [Google Scholar] [CrossRef]
- Lakhan, S.E.; Ford, C.T.; Tepper, D. Zingiberaceae extracts for pain: A systematic review and meta-analysis. Nutr. J. 2015, 14, 50. [Google Scholar] [CrossRef]
- Aryaeian, N.; Shahram, F.; Mahmoudi, M.; Tavakoli, H.; Yousefi, B.; Arablou, T.; Karegar, S.J. The effect of ginger supplementation on some immunity and inflammation intermediate genes expression in patients with active Rheumatoid Arthritis. Gene 2019, 698, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Koch, E. Complex interactions between phytochemicals. The multi-target therapeutic concept of phytotherapy. Curr. Drug Targets 2011, 12, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Richards, L.A.; Glassmire, A.E.; Ochsenrider, K.M.; Smilanich, A.M.; Dodson, C.D.; Jeffrey, C.S.; Dyer, L.A. Phytochemical diversity and synergistic effects on herbivores. Phytochem. Rev. 2016, 15, 1153–1166. [Google Scholar] [CrossRef]
- Pham, D.C.; Shibu, M.A.; Mahalakshmi, B.; Velmurugan, B.K. Effects of phytochemicals on cellular signaling: Reviewing their recent usage approaches. Crit. Rev. Food Sci. Nutr. 2020, 60, 3522–3546. [Google Scholar] [CrossRef]
- Santos, D.I.; Saraiva, J.M.A.; Vicente, A.A.; Moldão-Martins, M. Methods for determining bioavailability and bioaccessibility of bioactive compounds and nutrients. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Woodhead Publishing: Sawston, UK, 2019; pp. 23–54. [Google Scholar]
- Giampieri, F.; Quiles, J.L.; Cianciosi, D.; Forbes-Hernández, T.Y.; Orantes-Bermejo, F.J.; Alvarez-Suarez, J.M.; Battino, M. Bee products: An emblematic example of underutilized sources of bioactive compounds. J. Agric. Food Chem. 2022, 70, 6833–6848. [Google Scholar] [CrossRef]
- Guengerich, F.P. Human Cytochrome P450 Enzymes. Cytochrome P450: Structure, Mechanism, and Biochemistry; Springer: Berlin/Heidelberg, Germany, 2015; pp. 523–785. [Google Scholar]
- Motilva, M.J.; Serra, A.; Rubio, L. Nutrikinetic studies of food bioactive compounds: From in vitro to in vivo approaches. Int. J. Food Sci. Nutr. 2015, 66, S41–S52. [Google Scholar] [CrossRef]
- Ward, R.M.; Benjamin, D.; Barrett, J.S.; Allegaert, K.; Portman, R.; Davis, J.M.; Turner, M.A. Safety, dosing, and pharmaceutical quality for studies that evaluate medicinal products (including biological products) in neonates. Pediatr. Res. 2017, 81, 692–711. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Iyengar, R. Systems pharmacology: Network analysis to identify multiscale mechanisms of drug action. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 505–521. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Cui, H.; Liu, G.; Zhao, X.; Li, W.; Piao, G. How can synergism of traditional medicines benefit from network pharmacology? Molecules 2017, 22, 1135. [Google Scholar] [CrossRef]
- Block, K.I.; Gyllenhaal, C.; Lowe, L.; Amedei, A.; Amin, A.R.; Amin, A.; Aquilano, K.; Arbiser, J.; Arreola, A.; Arzumanyan, A.; et al. Designing a broad-spectrum integrative approach for cancer prevention and treatment. Semin. Cancer Biol. 2015, 35, S276–S304. [Google Scholar] [CrossRef] [PubMed]
- Guedes, I.A.; de Magalhães, C.S.; Dardenne, L.E. Receptor–ligand molecular docking. Biophys. Rev. 2014, 6, 75–87. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 18th ed.; Method 985.29; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Nam, D.G.; Kim, M.; Choe, J.S.; Choi, A.J. Effects of high-pressure, hydrothermal, and enzyme-assisted treatment on the taste and flavor profile of water-soluble ginger (Zingiber officinale) extract. Foods 2022, 11, 508. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.G.; Lee, S.; Im, J.Y.; Hwang, K.; Choe, J.S.; Im, P. Comparative Analysis of the Flavonoid Content of Korean Ginger (Zingiber officinale Rosc.) Leaves and Stem by Solvent Extraction using UPLC-DAD-QTOF/MS. Korean J. Food Cook Sci. 2021, 37, 81–92. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Li, C.X.; Wang, F.R.; Zhang, B.; Deng, Z.Y.; Li, H.Y. Stability and antioxidant activity of phenolic compounds during in vitro digestion. J. Food Sci. 2023, 88, 696–716. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards suite: From gene data mining to disease genome sequence analyses. Curr. Protoc. Bioinform. 2016, 54, 1–30. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.C.; Kwon, S.J.; Eom, S.H. Effect of thermal processing on color, phenolic compounds, and antioxidant activity of faba bean (Vicia faba L.) leaves and seeds. Antioxidants 2021, 10, 1207. [Google Scholar] [CrossRef]
- Yu, Y.; Li, H.; Zhang, B.; Wang, J.; Shi, X.; Huang, J.; Yang, J.; Zhang, Y.; Deng, Z. Nutritional and functional components of mulberry leaves from different varieties: Evaluation of their potential as food materials. Int. J. Food Prop. 2018, 21, 1495–1507. [Google Scholar] [CrossRef]
- Goulas, V.; Papoti, V.T.; Exarchou, V.; Tsimidou, M.Z.; Gerothanassis, I.P. Contribution of flavonoids to the overall radical scavenging activity of olive (Olea europaea L.) leaf polar extracts. J. Agric. Food Chem. 2010, 58, 3303–3308. [Google Scholar] [CrossRef] [PubMed]
- Yamassaki, F.T.; Campestrini, L.H.; Zawadzki-Baggio, S.F.; Maurer, J.B.B. Avocado leaves: Influence of drying process, thermal incubation, and storage conditions on preservation of polyphenolic compounds and antioxidant activity. Int. J. Food Prop. 2017, 20, 2280–2293. [Google Scholar] [CrossRef]
- Gong, J.; Xia, D.; Huang, J.; Ge, Q.; Mao, J.; Liu, S.; Zhang, Y. Functional components of bamboo shavings and bamboo leaf extracts and their antioxidant activities in vitro. J. Med. Food. 2014, 18, 453–459. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant activity/capacity measurement. 1. Classification, physicochemical principles, mechanisms, and electron transfer (ET)-based assays. J. Agric. Food Chem. 2016, 64, 997–1027. [Google Scholar] [CrossRef]
- Kunzmann, A.T.; Murray, L.J.; Cardwell, C.R.; McShane, C.M.; McMenamin, U.C.; Cantwell, M.M. PTGS2 (Cyclooxygenase-2) expression and survival among colorectal cancer patients: A systematic review. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1490–1497. [Google Scholar] [CrossRef]
- Markosyan, N.; Li, J.; Sun, Y.H.; Richman, L.P.; Lin, J.H.; Yan, F.; Quinones, L.; Sela, Y.; Yamazoe, T.; Gordon, N.; et al. Tumor cell–intrinsic EPHA2 suppresses antitumor immunity by regulating PTGS2 (COX-2). J. Clin. Investig. 2019, 129, 3594–3609. [Google Scholar] [CrossRef]
- Kobayashi, K.; Omori, K.; Murata, T. Role of prostaglandins in tumor microenvironment. Cancer Metastasis Rev. 2018, 37, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Travelli, C.; Ercoli, J.; Colombo, G.; Buoso, E.; Trabalzini, L. Prostaglandin E2 and cancer: Insight into tumor progression and immunity. Biology 2020, 9, 434. [Google Scholar] [CrossRef] [PubMed]
- Jo, B.G.; Park, N.J.; Jegal, J.; Choi, S.; Lee, S.W.; Yi, L.W.; Kim, S.N.; Yang, M.H. Stellera chamaejasme and its main compound luteolin 7-O-glucoside alleviates skin lesions in oxazolone-and 2, 4-dinitrochlorobenzene-stimulated murine models of atopic dermatitis. Planta Med. 2019, 85, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, K.S.; Afzal, O.; Kazmi, I.; Shaikh, M.A.J.; Thangavelu, L.; Gulati, M.; Singh, S.K.; Jha, N.K.; Gupta, P.K.; Chellappan, D.K.; et al. Nuclear factor-kappa B (NF-κB) inhibition as a therapeutic target for plant nutraceuticals in mitigating inflammatory lung diseases. Chem Biol. Interact. 2022, 354, 109842. [Google Scholar] [CrossRef] [PubMed]
- Soromou, L.W.; Chen, N.; Jiang, L.; Huo, M.; Wei, M.; Chu, X.; Millimouno, F.M.; Feng, H.; Sidime, Y.; Deng, X. Astragalin attenuates lipopolysaccharide-induced inflammatory responses by down-regulating NF-κB signaling pathway. Biochem. Biophys. Res. Commun. 2012, 419, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liang, D.; Yang, Z.; Wang, T.; Wang, W.; Song, X.; Guo, M.; Zhou, E.; Li, D.; Cao, Y.; et al. Astragalin suppresses inflammatory responses via down-regulation of NF-κB signaling pathway in lipopolysaccharide-induced mastitis in a murine model. Int. Immunopharmacol. 2013, 17, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine 2015, 74, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Asadi-Samani, M.; Bagheri, N.; Rafieian-Kopaei, M.; Shirzad, H. Inhibition of Th1 and Th17 cells by medicinal plants and their derivatives: A systematic review. Phytother. Res. 2017, 31, 1128–1139. [Google Scholar] [CrossRef]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharmacother. 2011, 133, 110917. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Values |
|---|---|
| Proximate composition (%) | |
| Moisture | 4.48 ± 0.25 (1) |
| Crude protein | 18.61 ± 0.03 |
| Crude fat | 4.51 ± 0.55 |
| Ashes | 10.05 ± 0.10 |
| Minerals (mg/100 g) | |
| Calcium | 866.00 ± 3.20 |
| Copper | 2.33 ± 0.03 |
| Iron | 30.40 ± 0.70 |
| Magnesium | 387.00 ± 2.30 |
| Manganese | 92.40 ± 0.60 |
| Phosphorus | 262.00 ± 1.50 |
| Potassium | 2548.00 ± 549.00 |
| Silicic acid | 2.22 ± 0.08 |
| Sodium | 33.90 ± 0.81 |
| Zinc | 3.32 ± 0.01 |
| Parameters | Values | Parameters | Values |
|---|---|---|---|
| Non-essential (mg/100 g) | Threonine | 30.59 ± 0.72 | |
| Alanine | 82.62 ± 0.54 (1) | Tryptophan | 27.09 ± 0.23 |
| Asparagine | - | Valine | 90.72 ± 0.45 |
| Aspartic acid | 29.55 ± 0.81 | Conditionally essential | |
| Glutamic acid | 112.81 ± 2.02 | Arginine | 1.24 ± 0.03 |
| Serine | 1.85 ± 0.23 | Citrulline | 11.71 ± 0.38 |
| Essential | Glutamine | - | |
| Histidine | 12.01 ± 0.28 | Glycine | 23.69 ± 1.22 |
| Isoleucine | 54.31 ± 0.16 | Proline | 2.12 ± 0.15 |
| Leucine | 67.03 ± 0.53 | Tyrosine | 1.34 ± 0.08 |
| Lysine | 29.31 ± 2.34 | Other amino acid | |
| Methionine | 5.09 ± 0.34 | GABA | 63.12 ± 3.12 |
| Phenylalanine | 35.78 ± 0.44 | Ornithine | 4.09 ± 0.49 |
| Symbol | Units, Full Name, Description | ME (ppm) (1) | m/z | Values |
|---|---|---|---|---|
| Flavonoid identification analysis | ||||
| Q3rob7rham | %, Quercetin 3-O-robinobioside-7-O-rhamnoside, Peak 1 | 0.4 | 757.2 | 15.97 ± 0.05 (2) |
| Q3rut7rham | %, Quercetin 3-O-rutinoside-7-O-rhamnoside, Peaks 2 | −0.2 | 757.2 | 20.46 ± 0.08 |
| Q3gal7rham | %, Quercetin 3-O-galactoside-7-O-rhamnoside, Peaks 5 | 0.7 | 611.2 | 13.02 ± 0.07 |
| Q3glu7rham | %, Quercetin 3-O-glucoside-7-O-rhamnoside, Peaks 6 | 1.7 | 611.2 | 6.80 ± 0.02 |
| K3rob7rham | %, Kaempferol 3-O-robinobioside-7-O-rhamnoside, Peaks 3 | 0.3 | 741.2 | 11.92 ± 0.12 |
| K3rut7rham | %, Kaempferol 3-O-rutinoside-7-O-rhamnoside, Peaks 4 | 0.7 | 741.2 | 4.65 ± 0.23 |
| K3glu | %, Kaempferol 3-O-glucoside, Peaks 7 | 2.1 | 449.1 | 0.71 ± 0.05 |
| K3gal7rham | %, Kaempferol 3-O-galactoside-7-O-rhamnoside, Peaks 8 | 0.4 | 595.1 | 12.71 ± 0.04 |
| K3glu7rham | %, Kaempferol 3-O-glucoside-7-O-rhamnoside, Peaks 9 | 0.8 | 595.1 | 13.77 ± 0.16 |
| Dose-dependence test and radical scavenging activity | ||||
| TP | mg GAE/g, total polyphenol | 20.44 ± 0.34 | ||
| TF | mg QE/g, total flavonoid | 16.46 ± 2.09 | ||
| DPPH-RSA | %, DPPH-radical scavenging activity | 66.93 ± 1.67 | ||
| ABTS-RSA | %, ABTS-radical scavenging activity | 61.90 ± 5.65 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, D.-G.; Kim, M.; Choi, A.-J.; Choe, J.-S. Health Benefits of Antioxidant Bioactive Compounds in Ginger (Zingiber officinale) Leaves by Network Pharmacology Analysis Combined with Experimental Validation. Antioxidants 2024, 13, 652. https://doi.org/10.3390/antiox13060652
Nam D-G, Kim M, Choi A-J, Choe J-S. Health Benefits of Antioxidant Bioactive Compounds in Ginger (Zingiber officinale) Leaves by Network Pharmacology Analysis Combined with Experimental Validation. Antioxidants. 2024; 13(6):652. https://doi.org/10.3390/antiox13060652
Chicago/Turabian StyleNam, Dong-Geon, Mina Kim, Ae-Jin Choi, and Jeong-Sook Choe. 2024. "Health Benefits of Antioxidant Bioactive Compounds in Ginger (Zingiber officinale) Leaves by Network Pharmacology Analysis Combined with Experimental Validation" Antioxidants 13, no. 6: 652. https://doi.org/10.3390/antiox13060652
APA StyleNam, D.-G., Kim, M., Choi, A.-J., & Choe, J.-S. (2024). Health Benefits of Antioxidant Bioactive Compounds in Ginger (Zingiber officinale) Leaves by Network Pharmacology Analysis Combined with Experimental Validation. Antioxidants, 13(6), 652. https://doi.org/10.3390/antiox13060652