
The Effect of Ovariectomy and Estradiol Substitution on the Metabolic Parameters and Transcriptomic Profile of Adipose Tissue in a Prediabetic Model

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Biochemical Analysis
2.3. Tissue Insulin Sensitivity
2.4. Transcriptomic Analysis
2.5. Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
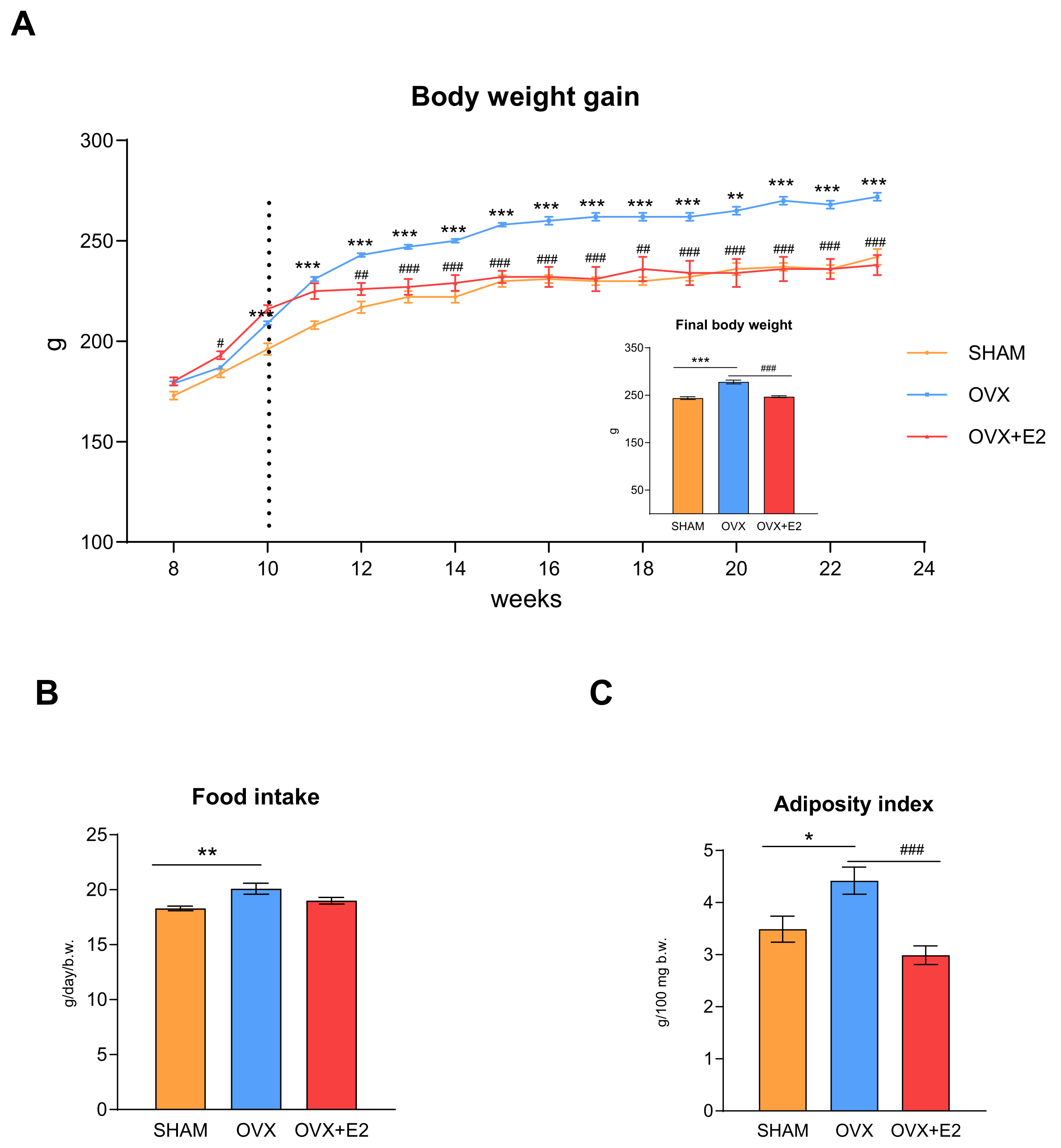
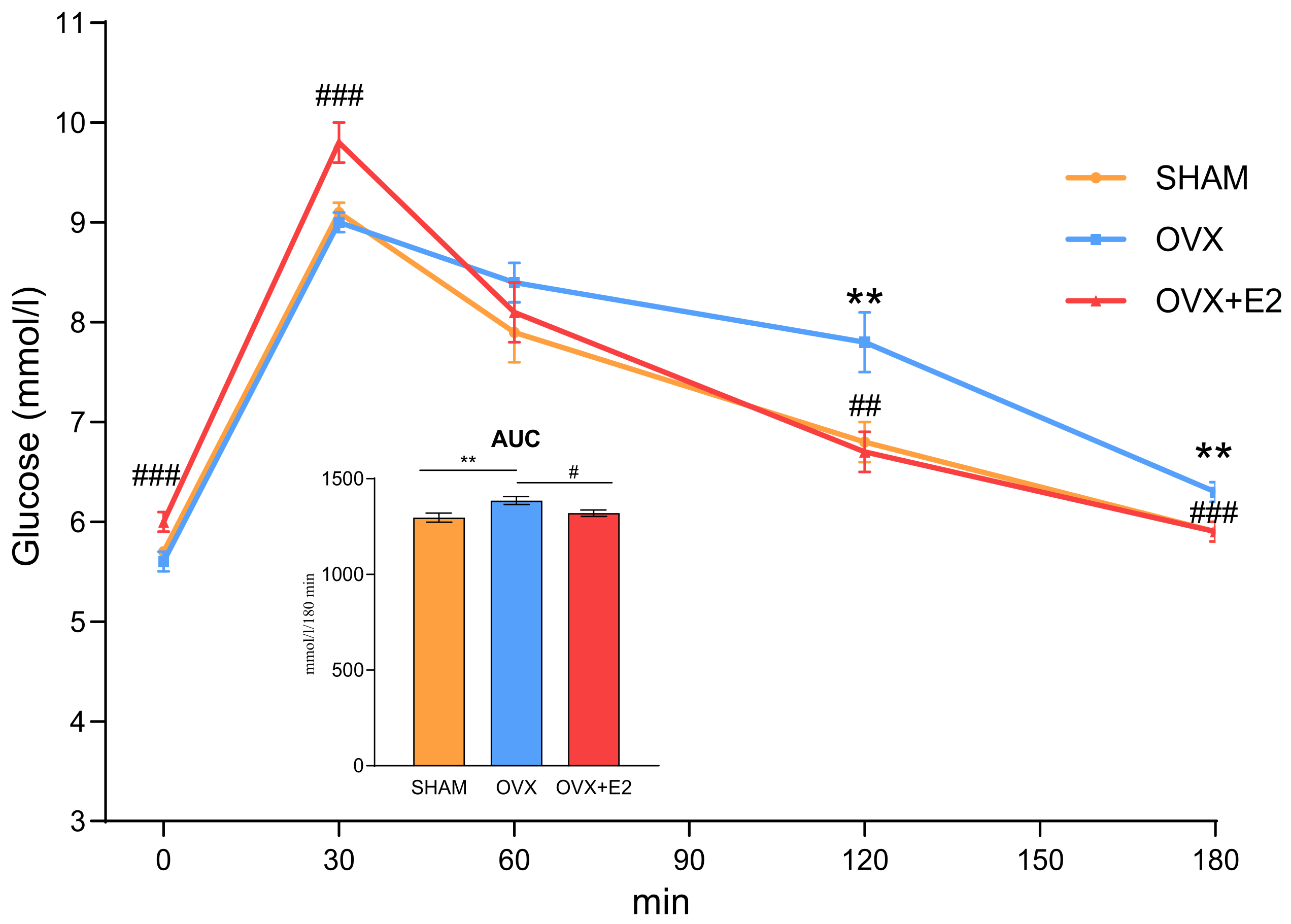
3.1. Effect of Ovariectomy and 17β-Estradiol Substitution on Body Weight, Food Intake, and Glucose Tolerance
3.2. Effect of Ovariectomy and 17β-Estradiol Substitution on Serum Metabolic Parameters, Inflammatory Markers, and Ectopic Lipid Accumulation
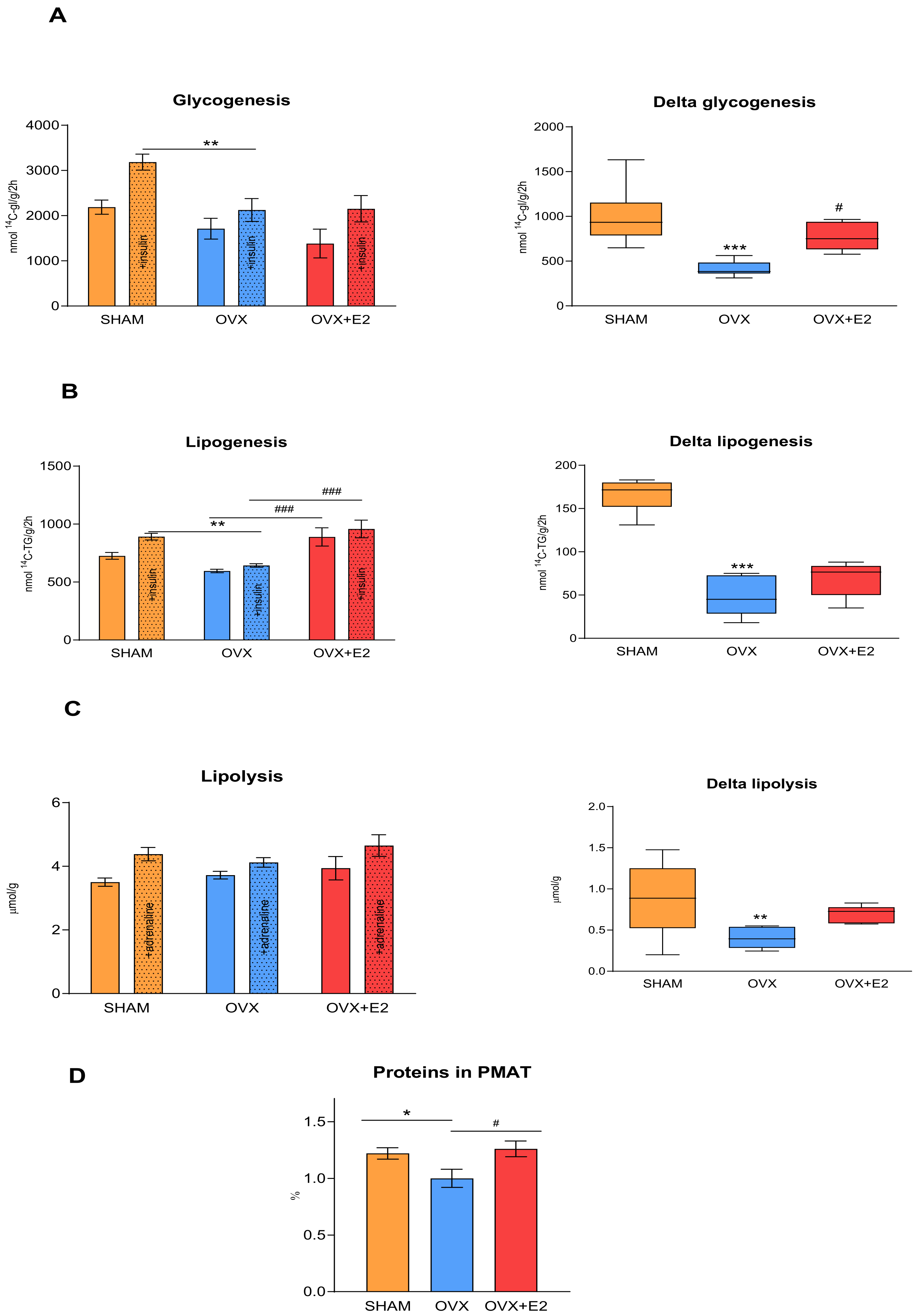
3.3. Effect of Ovariectomy and 17β-Estradiol Substitution on Insulin Sensitivity of Peripheral Tissues
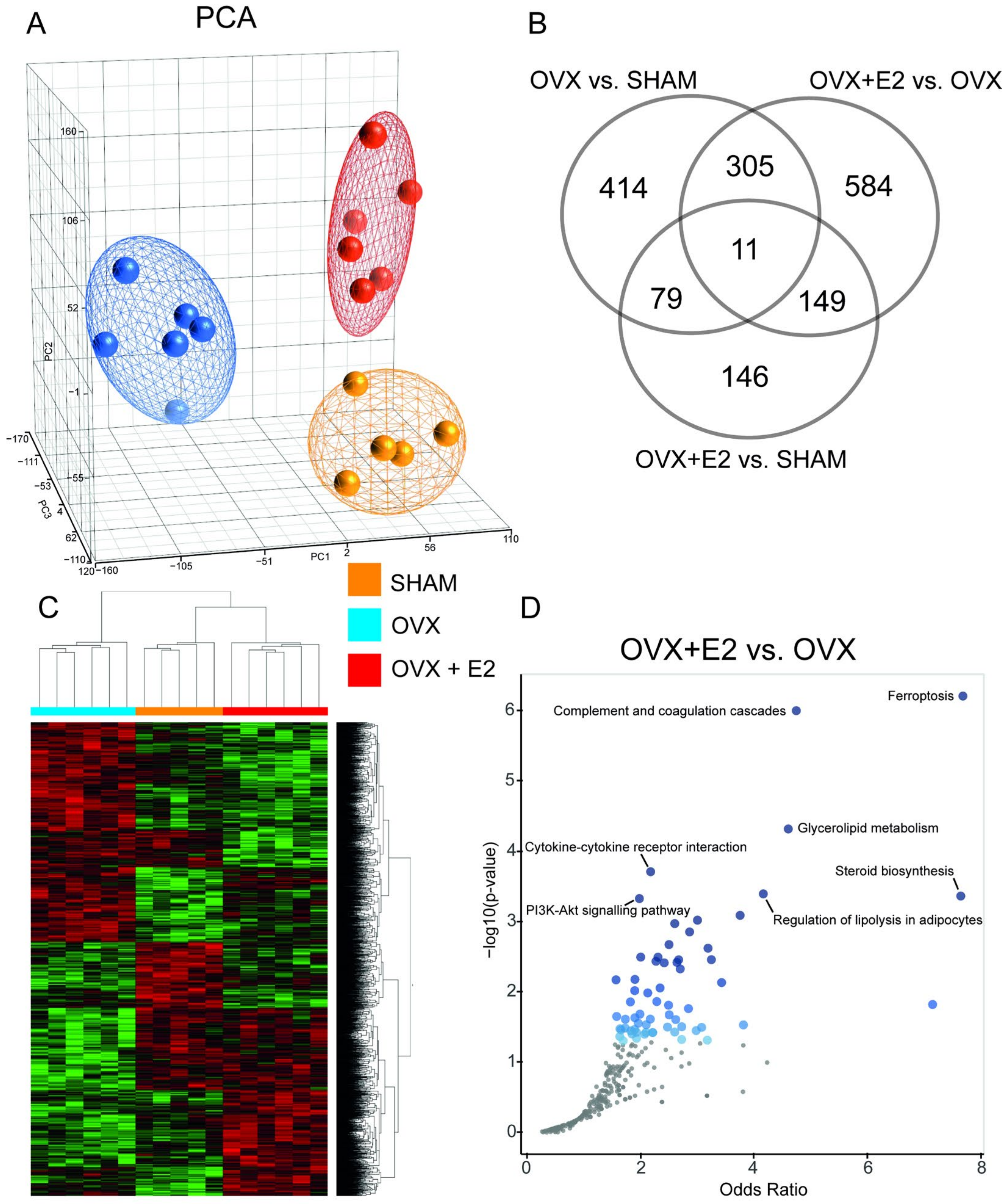
3.4. Effect of Ovariectomy and 17β-Estradiol Substitution on Gene Expression Profiles of Perimetrial Adipose Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | acetyl-CoA carboxylase |
| Acsl | acyl-CoA synthetase |
| AUC | area under the glycemic curve |
| B-H | Benjamini–Hochberg |
| Cish | cytokine-inducible SH2 domain containing protein |
| Cryab | crystallin alpha B |
| CVD | cardiovascular disease |
| DEG | differentially expressed genes |
| FDR | false discovery rate |
| GSH | glutathione |
| HMW adiponectin | high molecular weight adiponectin |
| hsCRP | high sensitivity C-reactive protein |
| HHTg | hereditary hypertriglyceridemic rat |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MCP-1 | monocyte chemoattractant protein-1 |
| MS | metabolic syndrome |
| NEFA | nonesterified fatty acids |
| OGTT | oral glucose tolerance test |
| OVX | ovariectomized HHTg rats |
| OVX+E2 | ovariectomized HHTg rats receiving 17β-estradiol |
| PCA | principal component analysis |
| PMAT | perimetrial adipose tissue |
| PPARγ | peroxisome proliferator-activated receptor-γ |
| Scd-1 | stearoyl-CoA desaturase-1 |
| SEM | standard error of the mean |
| Serpine1 | serpin family E member 1 |
| SHAM | sham-operated HHTg rats |
| qPCR | quantitative real-time PCR |
| TG | triglycerides |
References
- Jeong, J.H.; Park, H. Metabolic disorders in menopause. Metabolites 2022, 12, 954. [Google Scholar] [CrossRef] [PubMed]
- Hodis, H.N.; Mack, W.J. Menopausal Hormone Replacement Therapy and Reduction of All-Cause Mortality and Cardiovascular Disease: It’s About Time and Timing. Cancer J. 2022, 28, 208–223. [Google Scholar] [CrossRef] [PubMed]
- Pu, D.; Tan, R.; Yu, Q.; Wu, J. Metabolic syndrome in menopause and associated factors: A meta-analysis. Climacteric 2017, 20, 583–591. [Google Scholar] [CrossRef]
- Park, Y.W.; Zhu, S.; Palaniappan, L.; Heshka, S.; Carnethon, M.R.; Heymsfield, S.B. The metabolic syndrome: Prevalence and associated risk factor findings in the US population from the Third National Health and Nutrition Examination Survey, 1988–1994. Arch. Intern. Med. 2003, 163, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; Augustyn, M.N.; Ascott-Evans, B.H. The metabolic syndrome-pathogenesis, clinical features and management. Cardiovasc. J. S. Afr. 2002, 13, 181–186. [Google Scholar] [PubMed]
- Ribas, V.; Nguyen, M.T.A.; Henstridge, D.C.; Nguyen, A.K.; Beaven, S.W.; Watt, M.J.; Hevener, A.L. Impaired oxidative metabolism and inflammation are associated with insulin resistance in ERα-deficient mice. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E304–E319. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, S.H.; Mahmoud, B.F.; Saad, M.I.; Shehata, M.; Kamel, M.A.; Helmy, M.H. Similar and Additive Effects of Ovariectomy and Diabetes on Insulin Resistance and Lipid Metabolism. Biochem. Res. Int. 2015, 2015, 567945. [Google Scholar] [CrossRef] [PubMed]
- Eaton, S.A.; Sethi, J.K. Immunometabolic Links between Estroge, Adipose Tissue and Female Reproductive Metabolism. Biology 2019, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Kozakowski, J.; Gietka-Czernel, M.; Leszczyńska, D.; Majo, A. Obesity in menopause—our negligence or an unfortunate inevitability? Menopause Rev. 2017, 16, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, M.; Yokota-Nakagi, N.; Uji, M.; Yoshida, K.I.; Tazumi, S.; Takamata, A.; Uchida, Y.; Morimoto, K. Estrogen replacement enhances insulin-induced AS160 activation and improves insulin sensitivity in ovariectomized rats. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1296–E1304. [Google Scholar] [CrossRef] [PubMed]
- Yokota-Nakagi, N.; Takahashi, H.; Kawakami, M.; Takamata, A.; Uchida, Y.; Morimoto, K. Estradiol Replacement Improves High-Fat Diet-Induced Obesity by Suppressing the Action of Ghrelin in Ovariectomized Rats. Nutrients 2020, 12, 907. [Google Scholar] [CrossRef] [PubMed]
- Leeners, B.; Geary, N.; Tobler, P.N.; Asarian, L. Ovarian hormones and obesity. Human. Reproduction Update 2017, 23, 300–321. [Google Scholar] [CrossRef] [PubMed]
- Salpeter, S.R.; Walsh, J.M.E.; Ormiston, T.M.; Greyber, E.; Buckley, N.S.; Salpeter, E.E. Meta-analysis: Effect of hormone-replacement therapy on components of the metabolic syndrome in postmenopausal women. Diabetes Obes. Metab. 2006, 8, 538–554. [Google Scholar] [CrossRef]
- Mehta, J.M.; Chester, R.C.; Kling, J.M. The Timing Hypothesis: Hormone Therapy for Treating Symptomatic Women During Menopause and Its Relationship to Cardiovascular Disease. J. Womens Health 2019, 28, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Stuenkel, C.A. Menopause, hormone therapy and diabetes. Climacteric 2017, 20, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Malínská, H.; Hüttl, M.; Miklánková, D.; Trnovská, J.; Zapletalová, I.; Poruba, M.; Marková, I. Ovariectomy-Induced Hepatic Lipid and Cytochrome P450 Dysmetabolism Precedes Serum Dyslipidemia. Int. J. Mol. Sci. 2021, 22, 4527. [Google Scholar] [CrossRef] [PubMed]
- Boldarine, V.T.; Pedroso, A.P.; Brandão-Teles, C.; LoTurco, E.G.; Nascimento, C.M.O.; Oyama, L.M.; Bueno, A.A.; Martins-de-Souza, D.; Ribeiro, E.B. Ovariectomy modifies lipid metabolism of retroperitoneal white fat in rats: A proteomic aproach. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E427–E437. [Google Scholar] [CrossRef] [PubMed]
- Vrána, A.; Kazdová, L. The hereditary hypertriglyceridemic nonobese rat: An experimental model of human hypertriglyceridemia. Transplant. Proc. 1990, 22, 2579. [Google Scholar] [PubMed]
- Zicha, J.; Pechánová, O.; Cacányiová, S.; Cebová, M.; Kristek, F.; Török, J.; Simko, F.; Dobesová, Z.; Kunes, J. Hereditary hypertriglyceridemic rat: A suitable model of cardiovascular disease and metabolic syndrome? Physiol. Res. 2006, 55 (Suppl. 1), S49–S63. [Google Scholar] [CrossRef] [PubMed]
- Malinska, H.; Huttl, M.; Oliyarnyk, O.; Markova, I.; Poruba, M.; Racova, Z.; Kazdova, L.; Vecera, R. Beneficial effects of troxerutin on metabolic disorders in non-obese model of metabolic syndrome. PLoS ONE 2019, 14, e0220377. [Google Scholar] [CrossRef]
- Amengual-Cladera, E.; Capllonch-Amer, G.; Lladó, I.; Gianotti, M.; Proenza, A.M. Proteomic study of periovarian tissue in 17β-estradiol-treated and untreated ovariectomized rats. Biochem. Cell Biol. 2016, 94, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Després, J.-P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef]
- Qi, N.; Kazdova, L.; Zidek, V.; Landa, V.; Kren, V.; Pershadsingha, H.A.; Lezin, E.S.; Abumrad, N.A.; Pravenec, M.; Kurtz, T.W. Pharmacogenetic evidence that cd36 is a key determinant of the metabolic effects of pioglitazone. J. Biol. Chem. 2002, 277, 48501–48507. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R. Heart disease and stroke statistics-2019 update: A report from the American Heart Association. Circulation 2019, 139, e506–e532. [Google Scholar] [CrossRef] [PubMed]
- Sites, C.K.; Brochu, M.; Tchernov, A.; Poehlman, E.T. Relationship between hormone replacement therapy use with body fat distribution and insulin sensitivity in obese postmenopausal women. Metabolism 2001, 50, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cho, H.T.; Kim, Y.J. The role of estrogen in adipose tissue metabolism: Insights into glucose homeostasis regulation. Endocr. J. 2014, 61, 1055–1067. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.I.; Chou, P.; Tsai, S.T. The impact of years since menopause on the development of impaired glucose tolerance. J. Clin. Epidemiol. 2001, 54, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, S.; Holmäng, A.; Björntorp, P. The effects of oestrogen and progesterone on insulin sensitivity in female rats. Acta Physiol. Scand. 1993, 149, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kanaya, A.M.; Herrington, D.; Vittinghoff, E.; Lin, F.; Grady, D.; Bittner, V.; Cauley, J.A.; Barrett-Connor, E. Glycemic effects of postmenopausal hormone therapy: The heart and estrogen/progestin replacement study: A randomized, double-blind, placebo-controlled trial. Ann. Intern. Med. 2003, 138, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Menopause, estrogens and glucose homeostasis in women. Adv. Exp. Med. Biol. 2017, 1043, 217–225. [Google Scholar] [PubMed]
- Kim, J.H.; Meyers, M.S.; Khuder, S.S.; Abdallah, S.L.; Muturi, H.T.; Russo, L.; Tate, C.R.; Hevener, A.L.; Najjar, S.M.; Leloup, C.; et al. Tissue-selective estrogen complexes with bazedoxifene prevent metabolic dysfunction in female mice. Mol. Metab. 2014, 3, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Torosyan, N.; Visrodia, P.; Torbati, T.; Minissian, M.B.; Schufelt, C.L. Dyslipidemia in midlife women: Approach and considerations during the postmenopausal transition. Maturitas 2022, 166, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.L.; Xu, X.; Rang, W.Q.; Li, Y.J.; Song, H.P. Influence of ovariectomy and 17β-estradiol treatment on insulin sensitivity, lipid metabolism and post-ischemic cardiac function. Int. J. Cardiol. 2004, 97, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Babei, P.; Mehdizadeh, R.; Ansar, M.M.; Damirchi, A. Effects of ovariectomy and estrogen replacement therapy on visceral adipose tissue and serum adiponectin levels in rats. Menopause Int. 2010, 16, 100–104. [Google Scholar] [CrossRef]
- Hodis, H.N.; Mack, W.J.; Henderson, V.W.; Shoupe, D.; Budoff, M.J.; Hwang-Levine, J.; Li, Y.; Feng, M.; Dustin, L.; Kono, N.; et al. Vascular effects of early versus late postmenopausal treatment with estradiol. N. Engl. J. Med. 2016, 374, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Hsia, J.; Johnson, K.C.; Rossouw, J.E.; Assaf, A.R.; Lasser, N.L.; Trevisan, M.; Black, H.R.; Heckbert, S.R.; Detrano, R.; et al. Women’s Health Initiative, I. Estrogen plus progestin and the risk of coronary heart disease. N. Engl. J. Med. 2003, 349, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Hulley, S.; Grady, D.; Bush, T.; Furberg, C.; Herrington, D.; Riggs, B.; Vittinghoff, E. Randomized trial of estrogen plus progestin for secondary prevention of coronary heart disease in postmenopausal women. Heart and Estrogen/progestin Replacement Study (HERS) Research Group. JAMA 1998, 280, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Wiegratz, I.; Jung-Hoffmann, C.; Gross, W.; Kuhl, H. Effect of two oral contraceptives containing ethinyl estradiol and gestodene or norgestimate on different lipid and lipoprotein parameters. Contraception 1998, 58, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Walsh, B.W.; Shiff, I.; Rosner, B.; Greenberg, I.; Ravnikar, V.; Sacks, F.M. Effects of postmenopausal estrogen replacement on the concentrations and metabolism of plasma lipoproteins. N. Engl. J. Med. 1991, 325, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, E.J.; Foster, D.M.; Zech, L.A.; Lindgren, F.T.; Brewer, H.B., Jr.; Levy, R.I. The effects of estrogen administration on plasma lipoprotein metabolism in premenopausal females. J. Clin. Endocrinol. Metab. 1983, 572, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Pitha, J.; Huttl, M.; Malinska, H.; Miklankova, D.; Bartuskova, H.; Hlinka, T.; Markova, I. Cardiovascular, metabolic and inflammatory changes after ovariectomy and estradiol substitution in hereditary hypertriglyceridemic rats. Int. J. Mol. Sci. 2022, 23, 2825. [Google Scholar] [CrossRef] [PubMed]
- Geisler, C.E.; Renquist, B.J. Hepatic lipid accumulation: Cause and consequence of dysregulated glucoregulatory hormones. J. Endocrinol. 2017, 234, R1–R21. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.H.; Perfield, J.W.; Strissel, K.J.; Obin, M.S.; Greenberg, A.S. Reduced energy expenditure and increased inflammation are early events in the development of ovariectomy-induced obesity. Endocrinology 2009, 150, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Schadinger, S.E.; Bucher, L.R.; Schreiber, B.M.; Farmer, S.R. PPARγ2 regulates lipogenesis and lipid accumulation in steatotic hepatocytes. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E1195–E1205. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Estrogens in the regulation of liver lipid metabolism. Adv. Exp. Med. Biol. 2017, 1043, 227–256. [Google Scholar] [PubMed]
- Zhu, L.; Brown, W.C.; Cai, Q.; Krust, A.; Chambon, P.; McGuiness, O.P.; Stafford, J.M. Estrogen treatment after ovariectomy protects against fatty liver and may improve pathway-selective insulin resistance. Diabetes 2013, 62, 424–434. [Google Scholar] [CrossRef]
- Hevener, A.L.; Clegg, D.J.; Mauvais-Jarvis, F. Impaired estrogen receptor action in the pathogenesis of metabolic syndrome. Mol. Cell Endocrinol. 2015, 418 Pt 3, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Estrogen and androgen receptors: Regulators of fuel homeostasis and emerging targets for diabetes and obesity. Trends Endocrinol. Metab. 2011, 22, 24–33. [Google Scholar] [CrossRef]
- Markova, I.; Miklankova, D.; Hüttl, M.; Kacer, P.; Skibova, J.; Kucera, J.; Sedlacek, R.; Kacerova, T.; Kazdova, L.; Malinska, H. The effect of lipotoxicity on renal dysfunction in a nonobese rat model of metabolic syndrome: A urinary proteomic approach. J. Diabetes Res. 2019, 2019, 8712979. [Google Scholar] [CrossRef] [PubMed]
- Rabe, K.; Lehrke, M.; Parhofer, K.G.; Broedl, U.C. Adipokines and insulin resistance. Mol. Med. 2008, 14, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Sieminska, L.; Wojciechowska, C.; Niedziolka, D.; Marek, B.; Kos-Kudla, B.; Kajdaniuk, D.; Nowak, M. Effect of menopause and hormone replacement therapy on serum adiponectin levels. Metabolism 2005, 54, 1610–1614. [Google Scholar] [CrossRef] [PubMed]
- Mankowska, A.; Nowak, L.; Sypniewska, G. Adiponectin and metabolic syndrome in women at menopause. EJIFCC 2009, 19, 173–184. [Google Scholar] [PubMed]
- Lu, J.Y.; Huang, K.C.; Chang, L.C.; Huang, Y.S.; Chi, Y.C.; Su, T.C.; Cheng, C.L.; Yang, W.S. Adiponectin: A biomarker of obesity-induced insulin resistance in adipose tissue and beyond. J. Biomed. Sci. 2008, 15, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Vieira Potter, V.J.; Strissel, K.J.; Xie, C.; Chang, E.; Bennet, G.; Defuria, J.; Obin, M.S.; Greenberg, A.S. Adipose tissue inflammation and reduced insulin sensitivity in ovariectomized mice occurs in the absence of increased adiposity. Endocrinology 2012, 159, 4266–4277. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Zhang, S.; Weng, X.; Huang, J.; Zhao, H.; Dai, X.; Bai, X.; Bao, X.; Zhao, C.; Zeng, M.; et al. Estrogen deficiency accelerates postmenopausal atherosclerosis by inducing endothelial cell ferroptosis through inhibiting NRF2/GPX4 pathway. FASEB J. 2023, 37, e22992. [Google Scholar] [CrossRef] [PubMed]
- Pope, L.E. Dixon SJ. Regulation of ferroptosis by lipid metabolism. Trends Cell Biol. 2023, 33, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Lehr, S.; Hartwig, S.; Lamers, D.; Famulla, S.; Müller, S.; Hanisch, F.G.; Cuvelier, C.; Ruige, J.; Eckhardt, K.; Ouwens, D.M.; et al. Identification and validation of novel adipokines released from primary human adipocytes. Mol. Cell Proteomics 2012, 11, M111.010504. [Google Scholar] [CrossRef] [PubMed]
- Sárvári, A.K.; Van Hauwaert, E.L.; Markussen, L.K.; Gammelmark, E.; Marcher, A.B.; Ebbesen, M.F.; Nielsen, R.; Brewer, J.R.; Madsen, J.G.S.; Mandrup, S. Plasticity of Epididymal Adipose Tissue in Response to Diet-Induced Obesity at Single-Nucleus Resolution. Cell Metab. 2021, 33, 437–453.e5. [Google Scholar] [CrossRef]
- Lundholm, L.; Movérare, S.; Steffensen, K.R.; Nilsson, M.; Otsuki, M.; Ohlsson, C.; Gustafsson, J.A.; Dahlman-Wright, K. Gene expression profiling identifies liver X receptor alpha as an estrogen-regulated gene in mouse adipose tissue. J. Mol. Endocrinol. 2004, 32, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Yoshioka, M.; Gan, L.; St-Amand, J. Regulation of global gene expression by ovariectomy and estrogen in female adipose tissue. Obes. Res. 2005, 13, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, A.; Duguay, S.R.; Ouellette, R.J.; Surette, M.E. 17β-estradiol induces stearoyl-CoA desaturase-1 expression in estrogen receptor-positive breast cancer cells. BMC Cancer 2015, 15, 440. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Miyazaki, M.; Socci, N.D.; Hagge-Greenberg, A.; Liedtke, W.; Soukas, A.A.; Sharma, R.; Hudgins, L.C.; Ntambi, J.M.; Friedman, J.M. Role for stearoyl-CoA desaturase-1 in leptin-mediated weight loss. Science 2002, 297, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Xing, X.; Wang, H.; Wan, K.; Fan, R.; Liu, C.; Wang, Y.; Wu, W.; Wang, Y.; Wang, R. SCD1 is the critical signaling hub to mediate metabolic diseases: Mechanism and the development of its inhibitors. Biomed. Pharmacother. 2023, 170, 115586. [Google Scholar] [CrossRef] [PubMed]
- Naser, W.; Maymand, S.; Rivera, L.R.; Connor, T.; Liongue, C.; Smith, C.M.; Aston-Mourney, K.; McCulloch, D.R.; McGee, A.L.; Ward, A.C. Cytokine-inducible SH2 domain containing protein contributes to regulation of adiposity, food intake, and glucose metabolism. FASEB J. 2022, 36, e22320. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Deng, J.; Jiao, F.; Hu, X.; Jiang, H.; Yuan, F.; Chen, S.; Niu, Y.; Jiang, X.; Guo, F. Hepatic cytokine-inducible SH2-containing protein (CISH) regulates gluconeogenesis via cAMP-responsive element binding protein (CREB). FASEB J. 2022, 36, e22541. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SHAM | OVX | OVX+E2 | PANOVA | |
|---|---|---|---|---|
| Non-fasting glucose (mmol/L) | 7.867 ± 0.319 | 8.367 ± 0.154 | 8.067 ± 0.287 | n.s. |
| Insulin (nmol/L) | 0.217 ± 0.025 | 0.195 ± 0.014 | 0.167 ± 0.010 | n.s. |
| Triglycerides (mmol/L) | 3.557 ± 0.300 | 1.720 ± 0.105 *** | 3.298 ± 0.137 ### | <0.001 |
| Cholesterol (mmol/L) | 1.665 ± 0.104 | 1.849 ± 0.045 | 2.177 ± 0.064 ## | <0.001 |
| NEFA (mmol/L) | 0.611 ± 0.057 | 0.648 ± 0.032 | 0.654 ± 0.028 | n.s. |
| 17β-estradiol (pg/mL) | 28.951 ± 3.913 | 15.062 ± 1.156 | 283.598 ± 32.489 ### | <0.001 |
| Leptin (ng/mL) | 5.311 ± 0.229 | 7.322 ± 0.322 *** | 7.526 ± 0.360 | <0.001 |
| Ghrelin (pg/mL) | 248.167 ± 63.016 | 98.833 ± 32.149 | 566.000 ± 281.889 | n.s. |
| HMW adiponectin (μg/mL) | 2.702 ± 0.130 | 3.336 ± 0.387 | 3.144 ± 0.226 | n.s. |
| MCP-1 (ng/mL) | 5.297 ± 0.473 | 6.135 ± 0.270 | 7.383 ± 1.666 | n.s. |
| hsCRP (mg/mL) | 1.194 ± 0.034 | 1.278 ± 0.217 | 1.571 ± 0.098 | n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marková, I.; Hüttl, M.; Miklánková, D.; Šedová, L.; Šeda, O.; Malínská, H. The Effect of Ovariectomy and Estradiol Substitution on the Metabolic Parameters and Transcriptomic Profile of Adipose Tissue in a Prediabetic Model. Antioxidants 2024, 13, 627. https://doi.org/10.3390/antiox13060627
Marková I, Hüttl M, Miklánková D, Šedová L, Šeda O, Malínská H. The Effect of Ovariectomy and Estradiol Substitution on the Metabolic Parameters and Transcriptomic Profile of Adipose Tissue in a Prediabetic Model. Antioxidants. 2024; 13(6):627. https://doi.org/10.3390/antiox13060627
Chicago/Turabian StyleMarková, Irena, Martina Hüttl, Denisa Miklánková, Lucie Šedová, Ondřej Šeda, and Hana Malínská. 2024. "The Effect of Ovariectomy and Estradiol Substitution on the Metabolic Parameters and Transcriptomic Profile of Adipose Tissue in a Prediabetic Model" Antioxidants 13, no. 6: 627. https://doi.org/10.3390/antiox13060627
APA StyleMarková, I., Hüttl, M., Miklánková, D., Šedová, L., Šeda, O., & Malínská, H. (2024). The Effect of Ovariectomy and Estradiol Substitution on the Metabolic Parameters and Transcriptomic Profile of Adipose Tissue in a Prediabetic Model. Antioxidants, 13(6), 627. https://doi.org/10.3390/antiox13060627