


Oxidative Damage and Antioxidants as Markers for the Selection of Emersion Hardening Treatments in GreenshellTM Mussel Juveniles (Perna canaliculus)

 , and
, and
Abstract
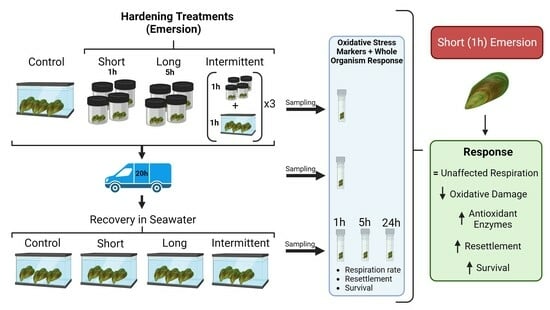
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Hardening, Transport and Recovery Phases
2.2. Resettlement Behaviour Test
2.3. Mussel Viability
2.4. Respiration Rate
2.5. Biomarker Analyses
2.5.1. Sampling for Biomarker Analyses
2.5.2. Macromolecule Extraction
2.5.3. Oxidative Damage Biomarkers
2.5.4. Enzymatic Antioxidant Biomarkers
2.6. Statistical Analyses
3. Results
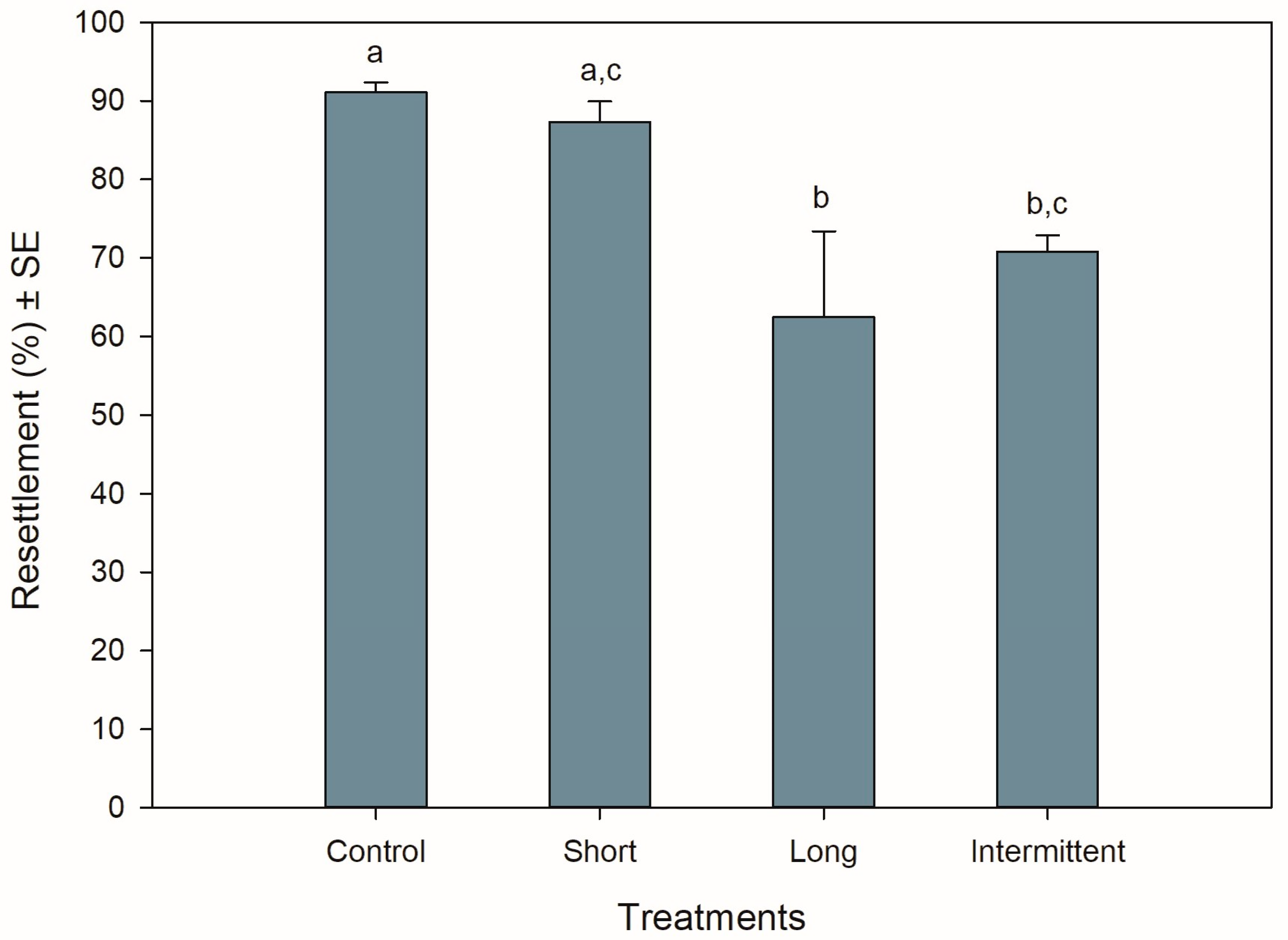
3.1. Resettlement Behaviour during Recovery
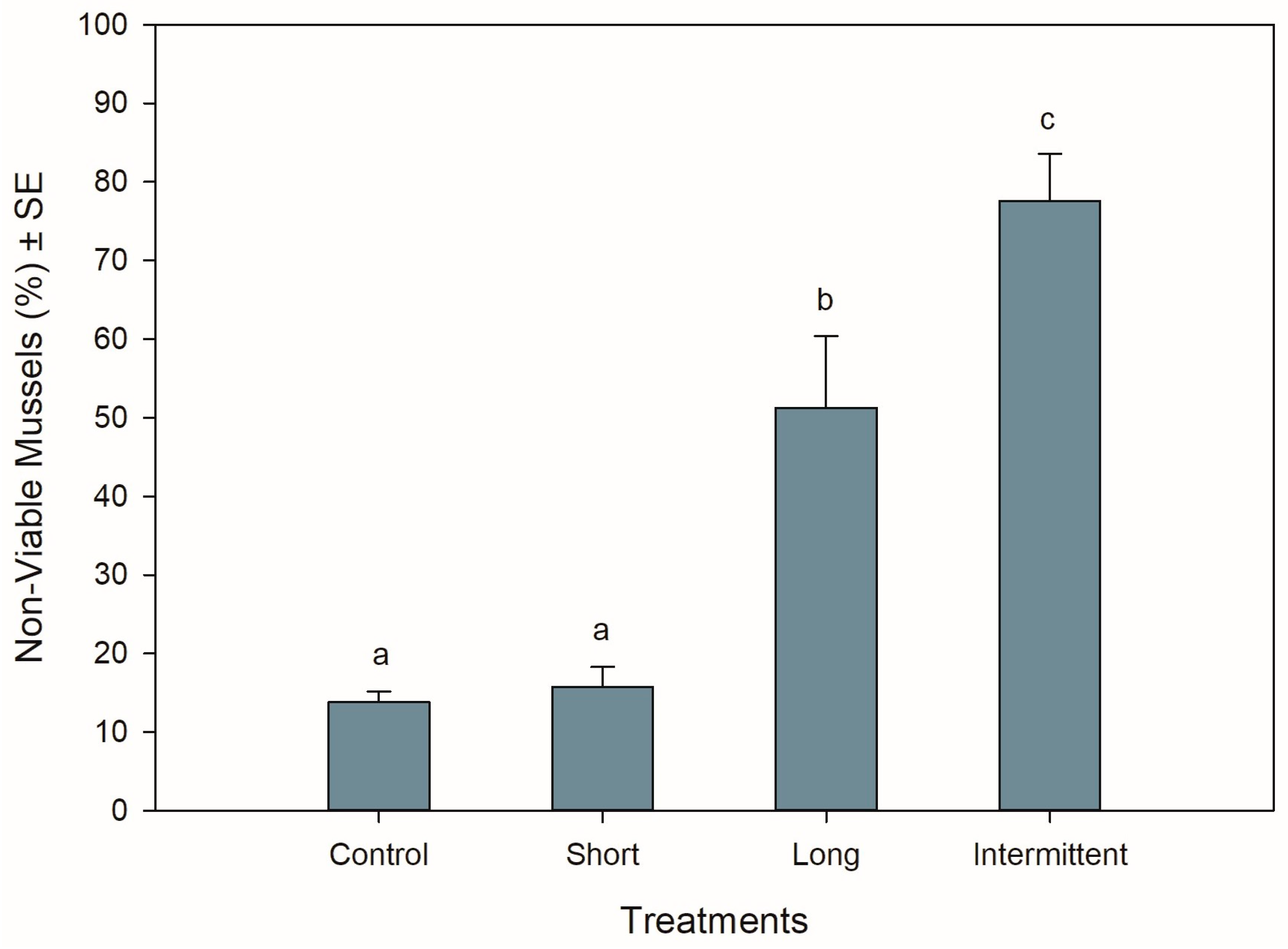
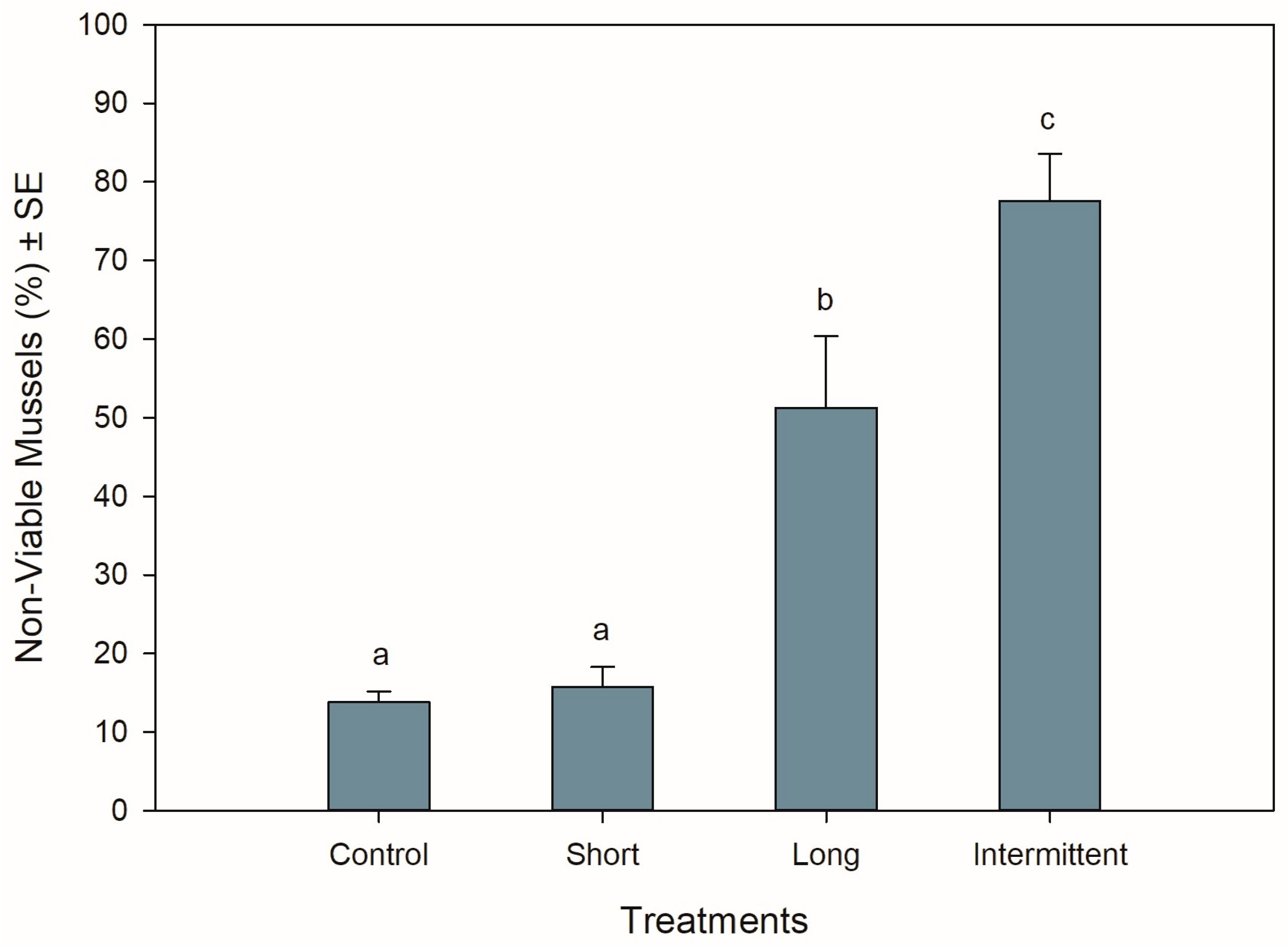
3.2. Mussel Viability at the End of Recovery
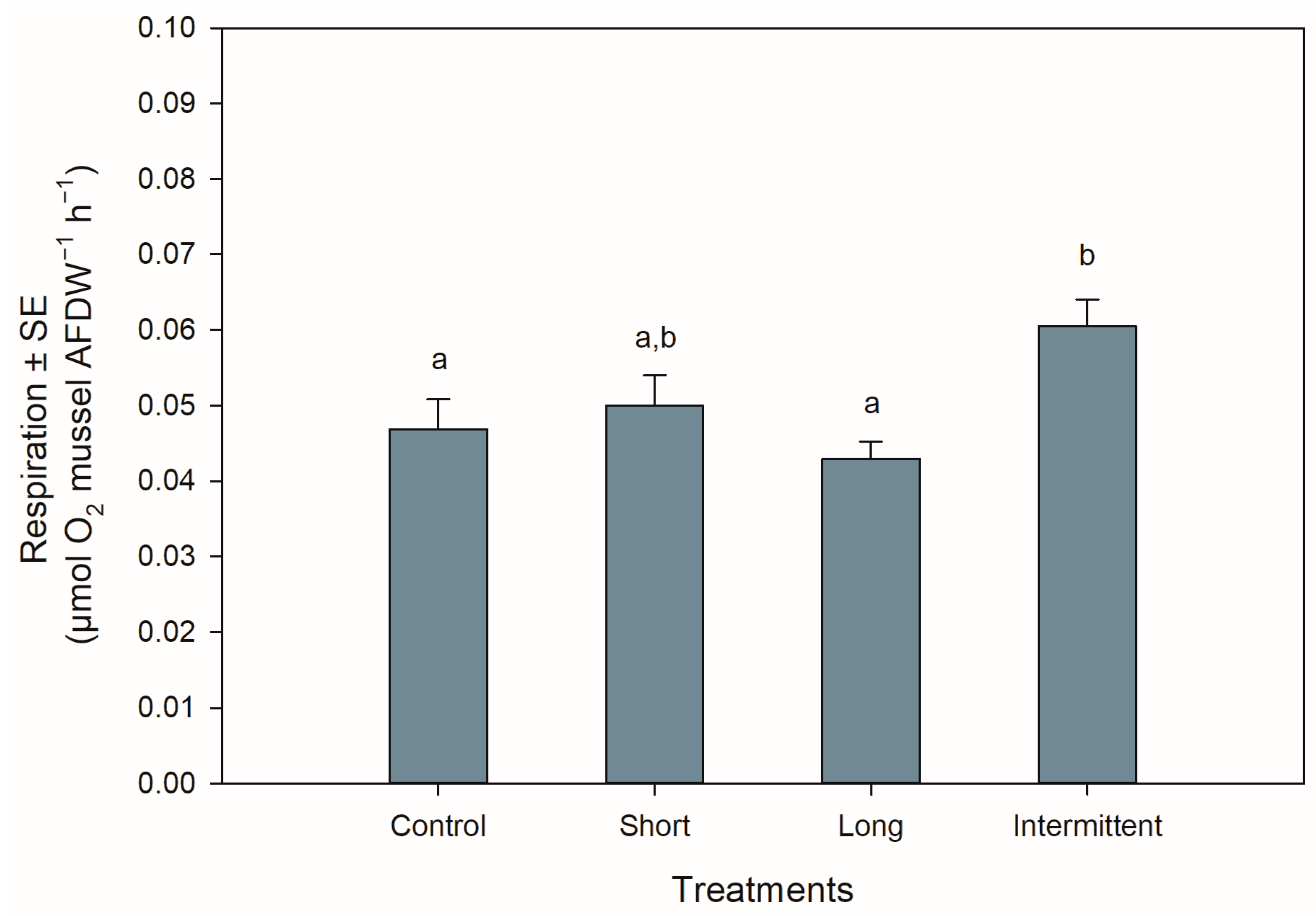
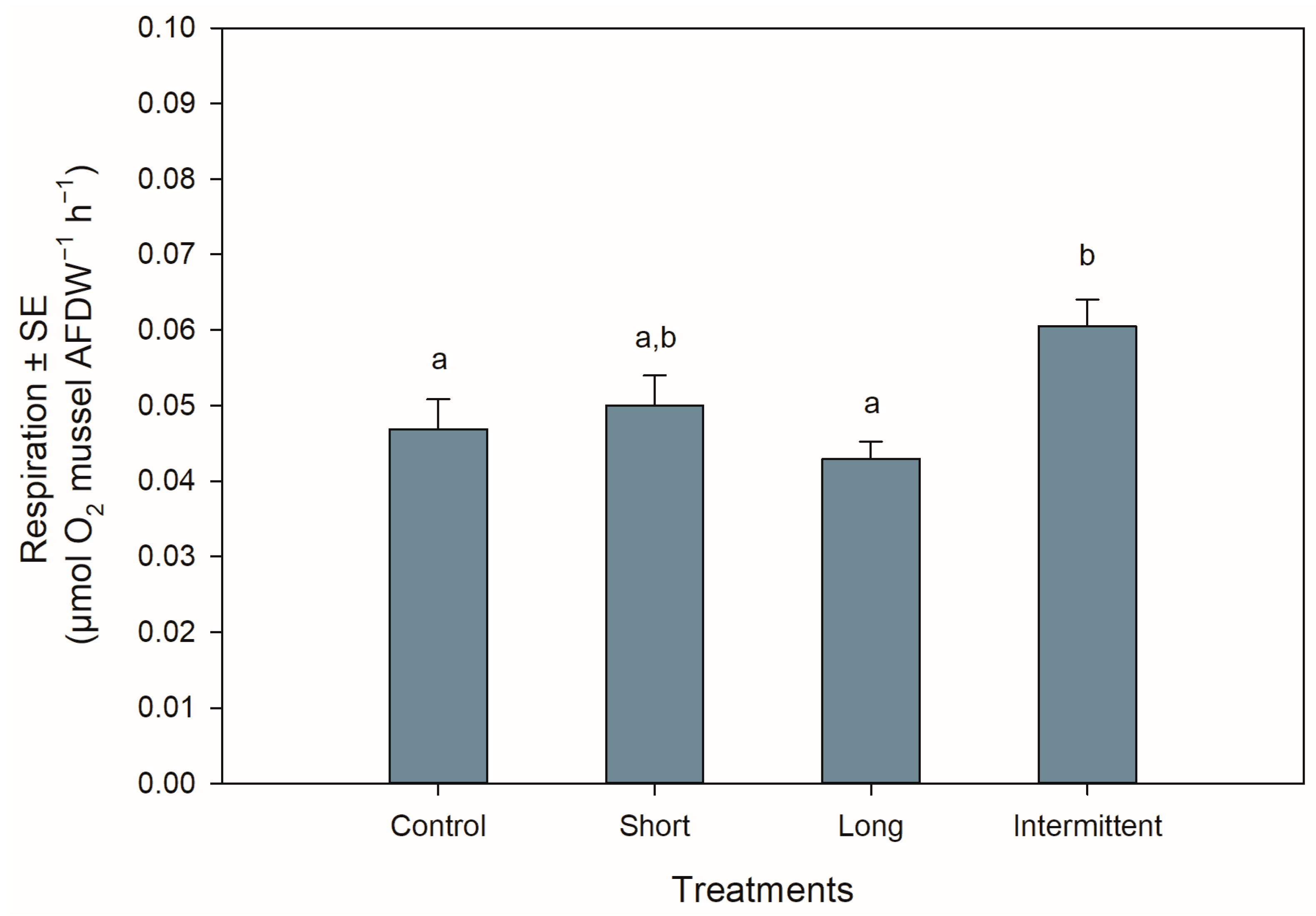
3.3. Respiration Rate during Recovery
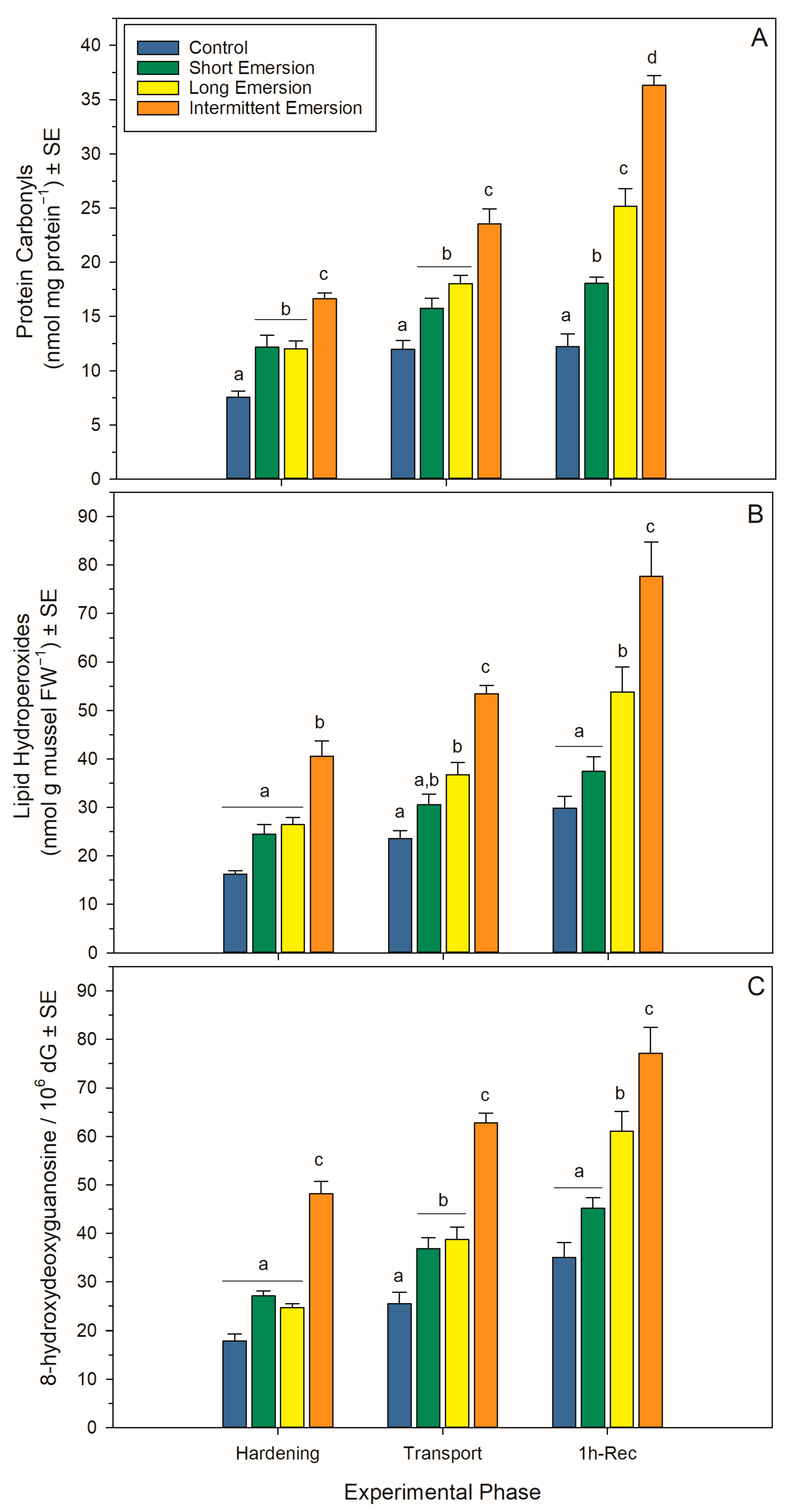
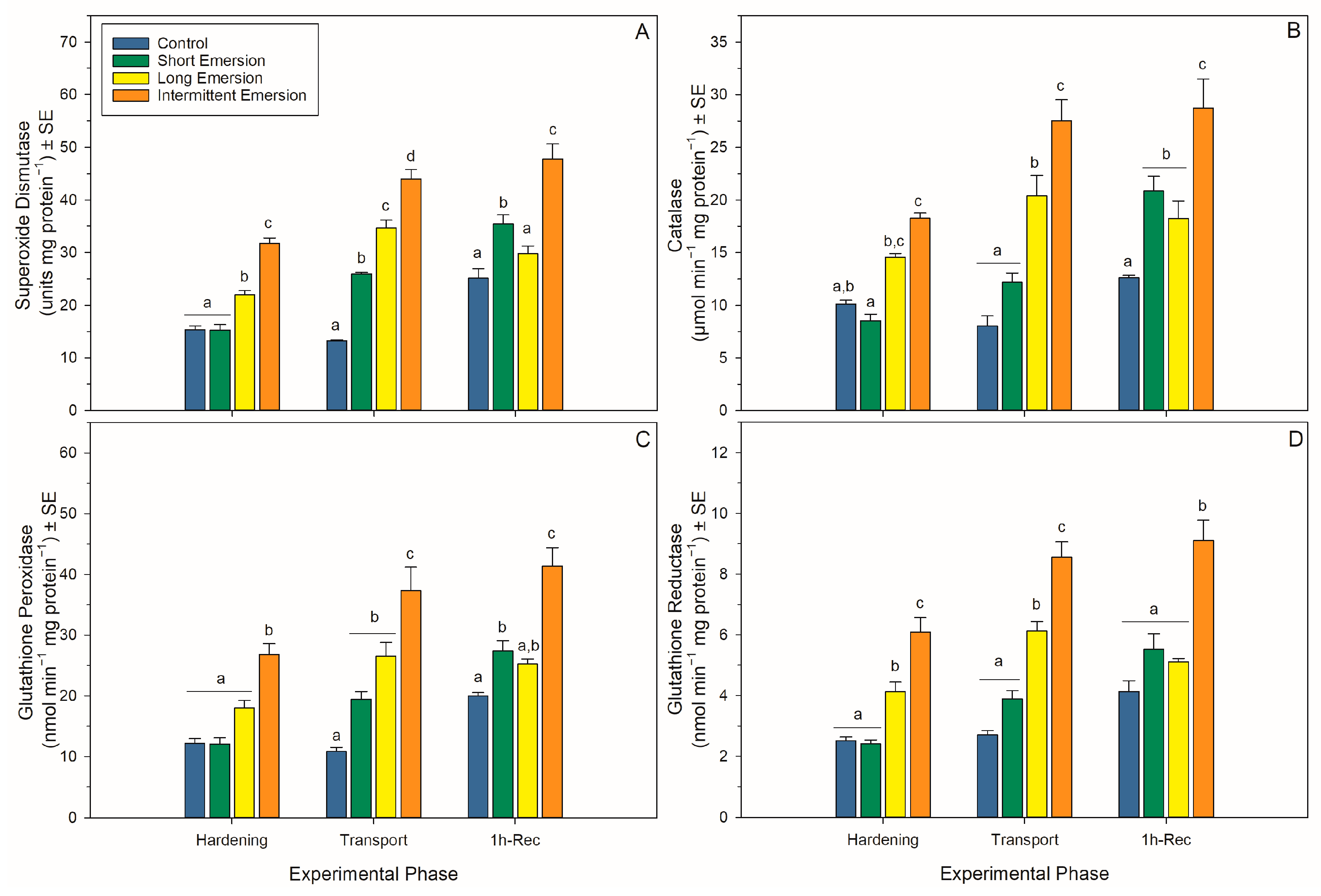
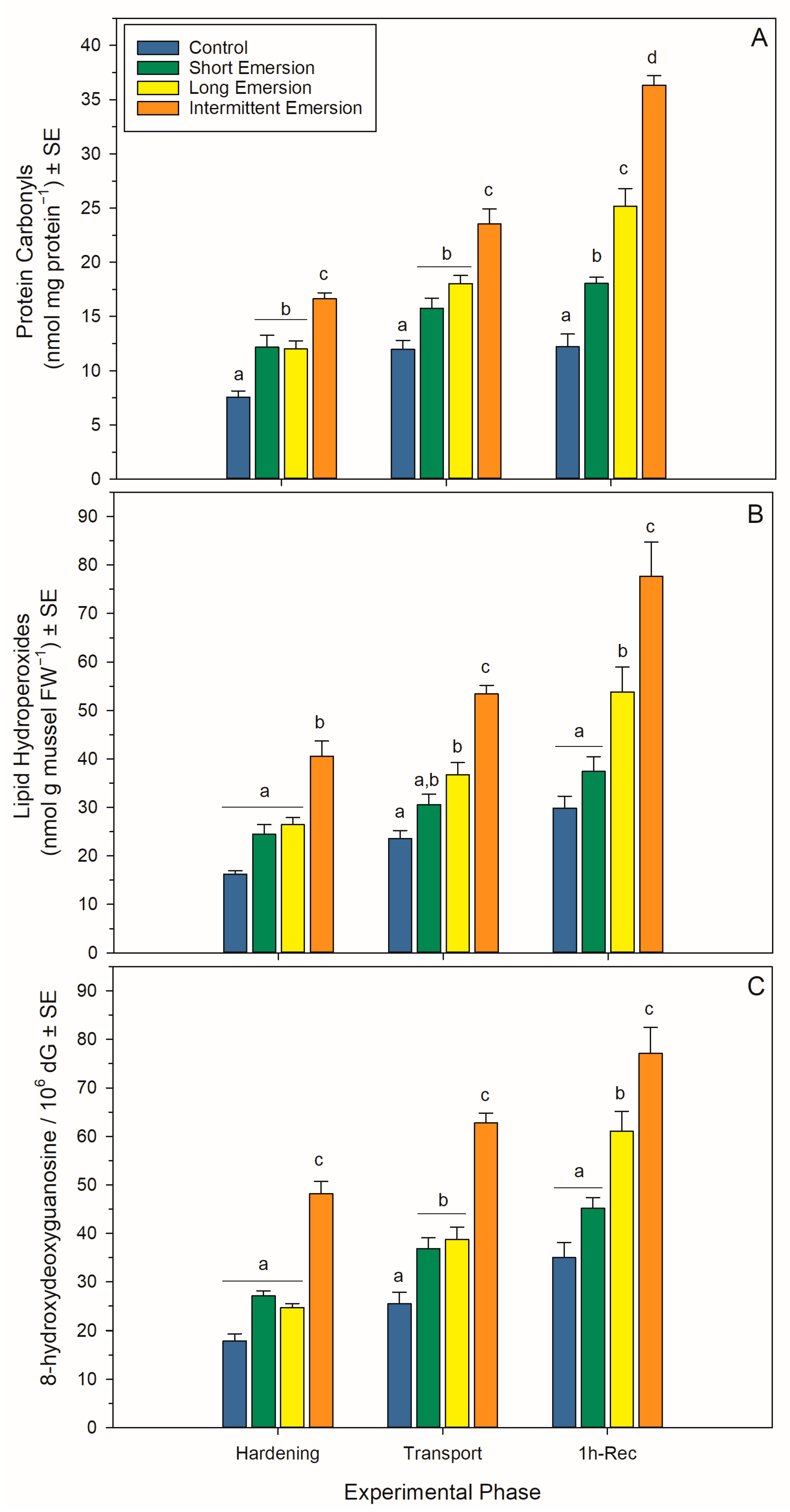
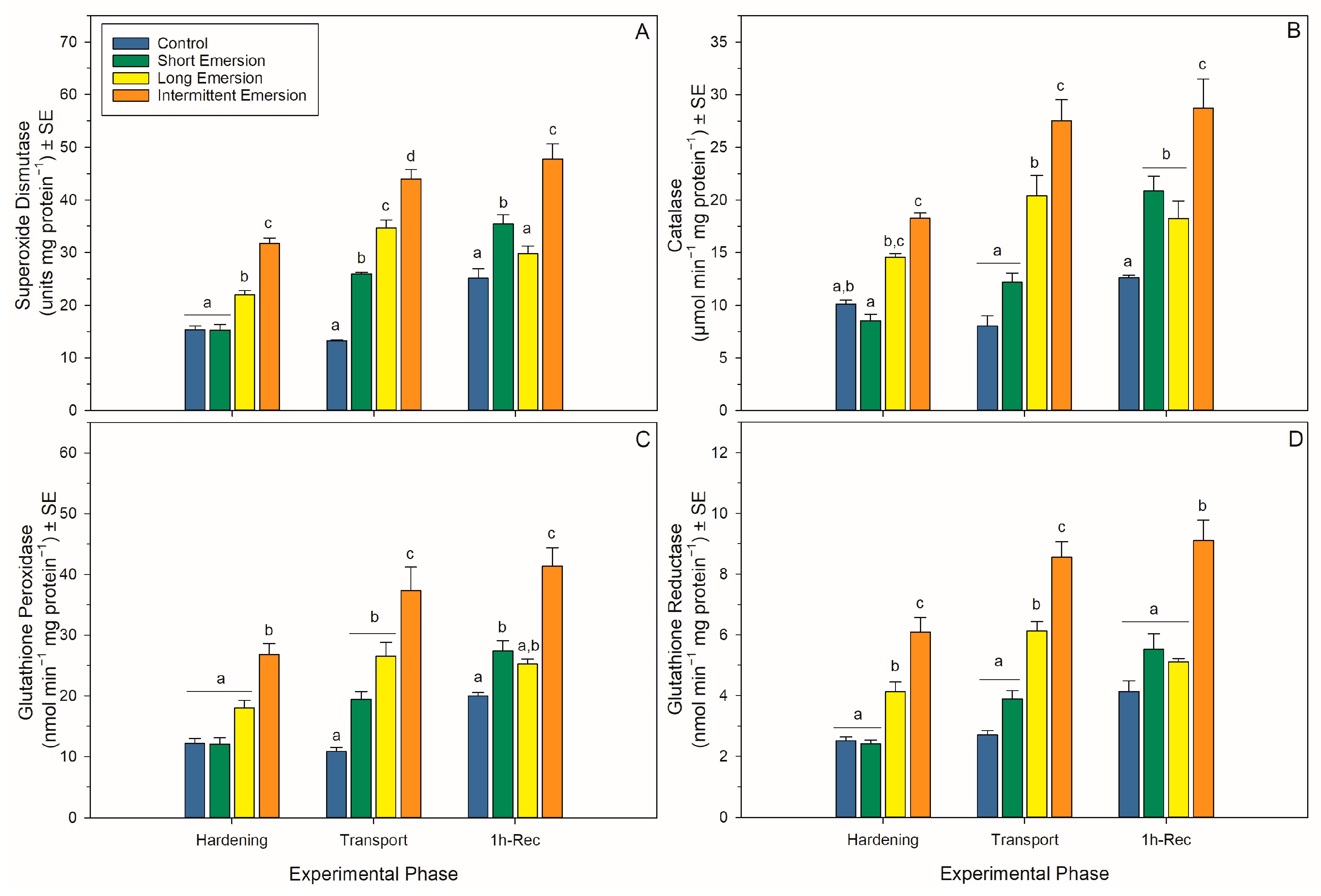
3.4. Oxidative Damage and Enzymatic Antioxidant Biomarkers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calderwood, J.; O’Connor, N.E.; Sigwart, J.D.; Roberts, D. Determining optimal duration of seed translocation periods for benthic mussel (Mytilus edulis) cultivation using physiological and behavioural measures of stress. Aquaculture 2014, 434, 288–295. [Google Scholar] [CrossRef]
- Sievers, M.; Fitridge, I.; Bui, S.; Dempster, T. To treat or not to treat: A quantitative review of the effect of biofouling and control methods in shellfish aquaculture to evaluate the necessity of removal. Biofouling 2017, 33, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Novaes, A.L.T.; de Andrade, G.J.P.O.; dos Santos Alonço, A.; Magalhães, A.R.M. Operational performance in aquaculture: A case study of the manual harvesting of cultivated mussels. Aquac. Eng. 2019, 84, 67–79. [Google Scholar] [CrossRef]
- Kamermans, P.; Capelle, J. Provisioning of mussel seed and its efficient use in culture. In Goods and Services of Marine Bivalves; Smaal, A.C., Ferreira, J.G., Grant, J., Petersen, J.K., Strand, O., Eds.; Springer: Cham, Switzerland, 2019; pp. 27–49. [Google Scholar]
- South, P.M.; Delorme, N.J.; Skelton, B.M.; Floerl, O.; Jeffs, A.G. The loss of seed mussels in longline aquaculture. Rev. Aquac. 2022, 14, 440–455. [Google Scholar] [CrossRef]
- Capelle, J.J.; Wijsman, J.W.; Schellekens, T.; van Stralen, M.R.; Herman, P.M.; Smaal, A.C. Spatial organisation and biomass development after relaying of mussel seed. J. Sea Res. 2014, 85, 395–403. [Google Scholar] [CrossRef]
- Carton, A.; Jeffs, A.; Foote, G.; Palmer, H.; Bilton, J. Evaluation of methods for assessing the retention of seed mussels (Perna canaliculus) prior to seeding for grow-out. Aquaculture 2007, 262, 521–527. [Google Scholar] [CrossRef]
- South, P.M.; Floerl, O.; Jeffs, A.G. Magnitude and timing of seed losses in mussel (Perna canaliculus) aquaculture. Aquaculture 2019, 515, 734528. [Google Scholar] [CrossRef]
- Hayden, B.J.; Woods, C.; Hatton, S. Handling and storage conditions to optimise survival of mussel spat. N. Z. Mussel Farmers Assoc. Newsl. 2007, 11–19. [Google Scholar]
- Hayden, B.J.; Woods, C.M.C. Effect of water velocity on growth and retention of cultured Greenshell™ mussel spat, Perna canaliculus (Gmelin, 1791). Aquac. Int. 2011, 19, 957–971. [Google Scholar] [CrossRef]
- Heasman, K. Temperature and Humidity of Kaitaia Weed during Harvesting, Storage and Transport and Suggested Influences on Spat viability. Prepared for Marine Farmers Association; Cawthron Report No. 2428; Cawthron Institute: Nelson, New Zealand, 2013; p. 18. [Google Scholar]
- Jenewein, B.T.; Gosselin, L.A. Ontogenetic shift in stress tolerance thresholds of Mytilus trossulus: Effects of desiccation and heat on juvenile mortality. Mar. Ecol. Prog. Ser. 2013, 481, 147–159. [Google Scholar] [CrossRef]
- Hamilton, H.; Gosselin, L. Ontogenetic shifts and interspecies variation in tolerance to desiccation and heat at the early benthic phase of six intertidal invertebrates. Mar. Ecol. Prog. Ser. 2020, 634, 15–28. [Google Scholar] [CrossRef]
- Rivera-Ingraham, G.A.; Rocchetta, I.; Meyer, S.; Abele, D. Oxygen radical formation in anoxic transgression and anoxia-reoxygenation: Foe or phantom? Experiments with a hypoxia tolerant bivalve. Mar. Environ. Res. 2013, 92, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Almeida, E.A.; Bainy, A.C.D.; Dafre, A.L.; Gomes, O.F.; Medeiros, M.H.; Di Mascio, P. Oxidative stress in digestive gland and gill of the brown mussel (Perna perna) exposed to air and re-submersed. J. Exp. Mar. Biol. Ecol. 2005, 318, 21–30. [Google Scholar] [CrossRef]
- Giannetto, A.; Maisano, M.; Cappello, T.; Oliva, S.; Parrino, V.; Natalotto, A.; De Marco, G.; Fasulo, S. Effects of oxygen availability on oxidative stress biomarkers in the mediterranean mussel Mytilus galloprovincialis. Mar. Biotechnol. 2017, 19, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Falfushynska, H.; Piontkivska, H.; Sokolova, I.M. Effects of intermittent hypoxia on cell survival and inflammatory responses in the intertidal marine bivalves Mytilus edulis and Crassostrea gigas. J. Exp. Biol. 2020, 223, 217026. [Google Scholar] [CrossRef] [PubMed]
- South, P.M.; Quirino, M.N.; LaDiega, C.; Delorme, N.J. Emersion and relative humidity control resettlement success of juvenile marine mussels. Aquaculture 2020, 529, 735675. [Google Scholar] [CrossRef]
- Delorme, N.J.; Burritt, D.J.; Ragg, N.L.C.; South, P.M. Emersion and Relative humidity modulate stress response and recovery dynamics in juvenile mussels (Perna canaliculus). Metabolites 2021, 11, 580. [Google Scholar] [CrossRef] [PubMed]
- Jeffs, A.G.; Holland, R.C.; Hooker, S.H.; Hayden, B.J. Overview and bibliography of research on the greenshell mussel, Perna canaliculus, from New Zealand waters. J. Shellfish. Res. 1999, 18, 347–360. [Google Scholar]
- Skelton, B.M.; Jeffs, A.G. The loss of spat following seeding onto coastal Greenshell™ mussel (Perna canaliculus) farms. Aquaculture 2021, 544, 737115. [Google Scholar] [CrossRef]
- Skelton, B.M.; South, P.M.; Jeffs, A.G. Inefficiency of conversion of seed into market-ready mussels in New Zealand’s Greenshell™ mussel (Perna canaliculus) industry. Aquaculture 2022, 560, 738584. [Google Scholar] [CrossRef]
- Delorme, N.; Biessy, L.; South, P.; Zamora, L.; Ragg, N.; Burritt, D. Stress-on-stress responses of a marine mussel, Perna canaliculus: Food limitation reduces the ability to cope with heat stress in juveniles. Mar. Ecol. Prog. Ser. 2020, 644, 105–117. [Google Scholar] [CrossRef]
- Supono, S.; Dunphy, B.; Jeffs, A. Retention of green-lipped mussel spat: The roles of body size and nutritional condition. Aquaculture 2020, 520, 735017. [Google Scholar] [CrossRef]
- Supono, S.; Yu, X.; Skelton, B.M.; McKay, W.J.; Jeffs, A. Effect of starvation on the nutritional condition of juvenile green-lipped mussels of different sizes. Aquaculture 2022, 560, 738580. [Google Scholar] [CrossRef]
- Bowler, K. Acclimation, heat shock and hardening. J. Therm. Biol. 2005, 30, 125–130. [Google Scholar] [CrossRef]
- Clegg, J.; Uhlinger, K.; Jackson, S.; Cherr, G.; Rifkin, E.; Friedman, C. Induced thermotolerance and the heat shock protein-70 family in the Pacific oyster Crassostrea gigas. Mol. Mar. Biol. Biotechnol. 1998, 7, 21–30. [Google Scholar]
- Dunphy, B.; Ruggiero, K.; Zamora, L.; Ragg, N. Metabolomic analysis of heat-hardening in adult green-lipped mussel (Perna canaliculus): A key role for succinic acid and the GABAergic synapse pathway. J. Therm. Biol. 2018, 74, 37–46. [Google Scholar] [CrossRef]
- Demers, A.; Guderley, H. Acclimatization to intertidal conditions modifies the physiological response to prolonged air exposure in Mytilus edulis. Mar. Biol. 1994, 118, 115–122. [Google Scholar] [CrossRef]
- Anestis, A.; Pörtner, H.O.; Michaelidis, B. Anaerobic metabolic patterns related to stress responses in hypoxia exposed mussels Mytilus galloprovincialis. J. Exp. Mar. Biol. Ecol. 2010, 394, 123–133. [Google Scholar] [CrossRef]
- Ragg, N.L.; King, N.; Watts, E.; Morrish, J. Optimising the delivery of the key dietary diatom Chaetoceros calcitrans to intensively cultured Greenshell™ mussel larvae, Perna canaliculus. Aquaculture 2010, 306, 270–280. [Google Scholar] [CrossRef]
- Webb, S.; Heasman, K. Evaluation of fast green uptake as a simple fitness test for spat of Perna canaliculus (Gmelin, 1791). Aquaculture 2006, 252, 305–316. [Google Scholar] [CrossRef]
- Fryer, H.J.; Davis, G.E.; Manthorpe, M.; Varon, S. Lowry protein assay using an automatic microtiter plate spectrophotometer. Anal. Biochem. 1986, 153, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Reznick, A.Z.; Packer, L. Oxidative damage to proteins: Spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Mihaljević, B.; Katušin-Ražem, B.; Ražem, D. The reevaluation of the ferric thiocyanate assay for lipid hydroperoxides with special considerations of the mechanistic aspects of the response. Free. Radic. Biol. Med. 1996, 21, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Maral, J.; Puget, K.; Michelson, A. Comparative study of superoxide dismutase, catalase and glutathione peroxidase levels in erythrocytes of different animals. Biochem. Biophys. Res. Commun. 1977, 77, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Janssens, B.J.; Childress, J.J.; Baguet, F.; Rees, J.-F. Reduced enzymatic antioxidative defense in deep-sea fish. J. Exp. Biol. 2000, 203, 3717–3725. [Google Scholar] [CrossRef] [PubMed]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Cribb, A.E.; Leeder, J.; Spielberg, S.P. Use of a microplate reader in an assay of glutathione reductase using 5,5′-dithiobis(2-nitrobenzoic acid). Anal. Biochem. 1989, 183, 195–196. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Freire, C.A.; Welker, A.F.; Storey, J.M.; Storey, K.B.; Hermes-Lima, M. Oxidative stress in estuarine and intertidal environments (temperate and tropical). In Oxidative Stress in Aquatic Ecosystems; Abele, D., Vázquez-Medina, J.P., Zenteno-Savín, T., Eds.; Blackwell Publishing Ltd.: West Sussex, UK, 2012; pp. 41–57. [Google Scholar]
- David, E.; Tanguy, A.; Pichavant, K.; Moraga, D. Response of the Pacific oyster Crassostrea gigas to hypoxia exposure under experimental conditions. FEBS J. 2005, 272, 5635–5652. [Google Scholar] [CrossRef]
- Delorme, N.J.; Venter, L.; Rolton, A.; Ericson, J.A. Integrating animal health and stress assessment tools using the green-lipped mussel Perna canaliculus as a case study. J. Shellfish. Res. 2021, 40, 93–112. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Sukhotin, A.A.; Lannig, G. Stress effects on metabolims and energy budgets in mollusks. In Oxidative Stress in Aquatic Ecosystems; Abele, D., Vázquez-Medina, J.P., Zenteno-Savín, T., Eds.; Blackwell Publishing Ltd.: West Sussex, UK, 2012; pp. 263–280. [Google Scholar]
- Istomina, A.; Yelovskaya, O.; Chelomin, V.; Karpenko, A.; Zvyagintsev, A. Antioxidant activity of Far Eastern bivalves in their natural habitat. Mar. Environ. Res. 2021, 169, 105383. [Google Scholar] [CrossRef]
- de Almeida, E.A.; Bainy, A.C.D. Effects of aerial exposure on antioxidant defenses in the brown mussel Perna perna. Braz. Arch. Biol. Technol. 2006, 49, 225–229. [Google Scholar] [CrossRef]
- Istomina, A.; Belcheva, N.; Chelomin, V. Antioxidant system of the intertidal mollusk Littorina kurila in its natural habitat. J. Environ. Sci. Eng. A 2013, 12A, 713–718. [Google Scholar]
- Welker, A.F.; Moreira, D.C.; Campos, É.G.; Hermes-Lima, M. Role of redox metabolism for adaptation of aquatic animals to drastic changes in oxygen availability. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 165, 384–404. [Google Scholar] [CrossRef]
- Giraud-Billoud, M.; Rivera-Ingraham, G.A.; Moreira, D.C.; Burmester, T.; Castro-Vazquez, A.; Carvajalino-Fernández, J.M.; Dafre, A.; Niu, C.; Tremblay, N.; Paital, B.; et al. Twenty years of the ‘Preparation for Oxidative Stress’(POS) theory: Ecophysiological advantages and molecular strategies. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 234, 36–49. [Google Scholar] [CrossRef]
- Sokolov, E.P.; Adzigbli, L.; Markert, S.; Bundgaard, A.; Fago, A.; Becher, D.; Hirschfeld, C.; Sokolova, I.M. Intrinsic mechanisms underlying hypoxia-tolerant mitochondrial phenotype during hypoxia-reoxygenation stress in a marine facultative anaerobe, the blue mussel Mytilus edulis. Front. Mar. Sci. 2021, 8, 773734. [Google Scholar] [CrossRef]
- Sokolov, E.P.; Markert, S.; Hinzke, T.; Hirschfeld, C.; Becher, D.; Ponsuksili, S.; Sokolova, I.M. Effects of hypoxia-reoxygenation stress on mitochondrial proteome and bioenergetics of the hypoxia-tolerant marine bivalve Crassostrea gigas. J. Proteom. 2019, 194, 99–111. [Google Scholar] [CrossRef]
- Adzigbli, L.; Sokolov, E.P.; Ponsuksili, S.; Sokolova, I.M. Tissue- and substrate-dependent mitochondrial responses to acute hypoxia–reoxygenation stress in a marine bivalve (Crassostrea gigas). J. Exp. Biol. 2022, 225, 243304. [Google Scholar] [CrossRef] [PubMed]
- Haider, F.; Falfushynska, H.I.; Timm, S.; Sokolova, I.M. Effects of hypoxia and reoxygenation on intermediary metabolite homeostasis of marine bivalves Mytilus edulis and Crassostrea gigas. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 242, 110657. [Google Scholar] [CrossRef] [PubMed]
- Georgoulis, I.; Feidantsis, K.; Giantsis, I.A.; Kakale, A.; Bock, C.; Pörtner, H.O.; Sokolova, I.M.; Michaelidis, B. Heat hardening enhances mitochondrial potential for respiration and oxidative defence capacity in the mantle of thermally stressed Mytilus galloprovincialis. Sci. Rep. 2021, 11, 17098. [Google Scholar] [CrossRef]
- Nicastro, K.R.; Zardi, G.I.; McQuaid, C.D.; Pearson, G.A.; Serrão, E.A. Love Thy Neighbour: Group Properties of gaping behaviour in mussel aggregations. PLoS ONE 2012, 7, e47382. [Google Scholar] [CrossRef]
- Zamora, L.N.; Ragg, N.L.; Hilton, Z.; Webb, S.C.; King, N.; Adams, S. Emersion survival manipulation in Greenshell™ mussels (Perna canaliculus): Implications for the extension of live mussels’ shelf-life. Aquaculture 2019, 500, 597–606. [Google Scholar] [CrossRef]
- Powell, J.; Ragg, N.; Dunphy, B. Phenotypic biomarkers in selectively-bred families of the Greenshell™ mussel (Perna canaliculus): Anaerobic enzyme and shell gape behaviour as biomarkers of prolonged emersion tolerance. Aquaculture 2017, 479, 601–608. [Google Scholar] [CrossRef]
- Andreyeva, A.; Gostyukhina, O.L.; Kladchenko, E.S.; Afonnikov, D.A.; Rasskazov, D.A.; Lantushenko, A.O.; Vodiasova, E.A. Hypoxia exerts oxidative stress and changes in expression of antioxidant enzyme genes in gills of Mytilus galloprovincialis (Lamarck, 1819). Mar. Biol. Res. 2021, 17, 369–379. [Google Scholar] [CrossRef]
- Gostyukhina, O.; Yu, A.A.; Chelebieva, E.; Vodiasova, E.; Lantushenko, A.; Kladchenko, E. Adaptive potential of the Mediterranean mussel Mytilus galloprovincialis to short-term environmental hypoxia. Fish Shellfish. Immunol. 2022, 131, 654–661. [Google Scholar] [CrossRef]
- Foschi, J.; Mancini, G.; Fabbri, M.; Rosmini, R.; Serrazanetti, G.; Monari, M. Antioxidant defences role during post anoxic recovery in bivalve mollusc Scapharca inaequivalvis. J. Biol. Res.-Boll. Della Soc. Ital. Di Biol. Sper. 2011, 84. [Google Scholar] [CrossRef]
- Ravishankar, S.; Ragg, N.; Delorme, N.; Dunphy, B. Thermotolerance of Greenshell™ mussel spat (Perna canaliculus) improved by prickly pear (Opuntia ficus indica) treatments. Aquaculture 2023, 562, 738738. [Google Scholar] [CrossRef]
- Zhang, W.; Dong, Y. Membrane lipid metabolism, heat shock response and energy costs mediate the interaction between acclimatization and heat-hardening response in the razor clam Sinonovacula constricta. J. Exp. Biol. 2021, 224, jeb243031. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delorme, N.J.; Burritt, D.J.; Zamora, L.N.; Welford, M.R.V.; South, P.M. Oxidative Damage and Antioxidants as Markers for the Selection of Emersion Hardening Treatments in GreenshellTM Mussel Juveniles (Perna canaliculus). Antioxidants 2024, 13, 198. https://doi.org/10.3390/antiox13020198
Delorme NJ, Burritt DJ, Zamora LN, Welford MRV, South PM. Oxidative Damage and Antioxidants as Markers for the Selection of Emersion Hardening Treatments in GreenshellTM Mussel Juveniles (Perna canaliculus). Antioxidants. 2024; 13(2):198. https://doi.org/10.3390/antiox13020198
Chicago/Turabian StyleDelorme, Natalí J., David J. Burritt, Leonardo N. Zamora, Mena R. V. Welford, and Paul M. South. 2024. "Oxidative Damage and Antioxidants as Markers for the Selection of Emersion Hardening Treatments in GreenshellTM Mussel Juveniles (Perna canaliculus)" Antioxidants 13, no. 2: 198. https://doi.org/10.3390/antiox13020198
APA StyleDelorme, N. J., Burritt, D. J., Zamora, L. N., Welford, M. R. V., & South, P. M. (2024). Oxidative Damage and Antioxidants as Markers for the Selection of Emersion Hardening Treatments in GreenshellTM Mussel Juveniles (Perna canaliculus). Antioxidants, 13(2), 198. https://doi.org/10.3390/antiox13020198