Abstract
Neurodegenerative diseases (NDs) are a large category of progressive neurological disorders with diverse clinical and pathological characteristics. Among the NDs, Alzheimer’s disease (AD) is the most widespread disease, which affects more than 400 million people globally. Oxidative stress is evident in the pathophysiology of nearly all NDs by affecting several pathways in neurodegeneration. No single drug can manage multi-faceted diseases like NDs. Therefore, an alternative therapeutic strategy is required, which can affect several pathophysiological pathways at a time. To achieve this aim, hexane and ethyl acetate extract from Trachyspermum ammi (Carom) were prepared, and GC/MS identified the bioactive compounds. For the cell-based assays, oxidative stress was induced in SH-SY5Y neuroblastoma cells using hydrogen peroxide to evaluate the neuroprotective potential of the Carom extracts/bioactives. The extracts/bioactives provided neuroprotection in the cells by modulating multiple pathways involved in neurodegeneration, such as alleviating oxidative stress and mitochondrial membrane potential. They were potent inhibitors of acetylcholine esterase enzymes and displayed competitive/mixed-type inhibition. Additionally, anti-Aβ1-42 fibrilization/oligomerization and anti-glycation activities were also analyzed. The multi-faceted neuroprotection shown via Carom/Carvacrol makes it a prospective contender in drug development for NDs.
1. Introduction
Neurodegenerative diseases (NDs) (neuro: brain/neuron; degenerative: deterioration) have a progressive and devastating effect on the neuronal communication network. NDs are a group of complex diseases that arise in different brain regions and affect cognition, memory, speech, body movement, balance, and much more. The central pathological developments linked with NDs are the accumulation of specific proteins, synapse dysfunction, oxidative stress, apoptosis, inflammation, and anomalies in the ubiquitin–proteasomal, autophagosomal, or lysosomal systems [1]. NDs affect millions of people worldwide, with the most common being Alzheimer’s disease (AD) and Parkinson’s disease (PD). Globally, 416 million people are affected by AD [2], and it is the fourth leading reason for disability-adjusted life-years (DALYs) lost in the elderly population [3]. As per the recent Alzheimer’s Disease Association report, the number of deaths due to AD increased by 145% between the year 2000 and 2019 [4].
Till now, there is no single drug that can manage multi-faceted diseases like AD. The present drugs are target specific viz. Donepezil, Galantamine, and Rivastigmine are acetylcholine esterase inhibitors, Aduhelm is an amyloid antibody, and Memantine acts as an NMDA receptor antagonist. Secondly, these drugs have various side effects extending from moderate dizziness to anomalies observed in brain scans [amyloid-related imaging abnormalities (ARIA)]. Hence, a multi-faceted neuroprotective strategy is necessary to improve the disease symptoms, for which the search for novel compounds with better efficacy and minimum side effects is highly desirable. In this context, the neuroprotective potential of numerous plants has been recognized [5].
Carom (Trachyspermum ammi) is an annual herb from the Apiaceae family. The small light-brown seed-like fruits of Carom are highly fragrant and are used as a flavoring agent in cuisines owing to their pungent and aromatic taste. Besides being used as a spice, Carom has been used in the traditional system of Indian (Ayurveda) and Chinese medicines in treating conditions like peptic ulcers, hypertension, hyperlipidemia, diabetes, diarrhea, arthritis, viral infections, asthma, and indigestion [6]. Carom essential oil (EO) has been known for its antibacterial, anti-fungal, insecticidal, and antioxidant properties [7]. GC-MS analysis of the EO identified Thymol and Carvacrol as the principal components, while γ-terpinene, camphene, ρ-cymene, δ-3-carene, β-pinene, myrcene, limonene, and sabinene as the minor constituents [8]. The extract and the key component provided neuroprotection and amended learning and memory in the scopolamine-induced memory deficit mice model [9]. Thymol also alleviated the symptoms of diabetic–neuropathic pain in the Streptozotocin (STZ)-induced rat model [10]. The neuroprotective role of Carvacrol has been studied in animal models of traumatic brain injury (TBI) [11], PD [12,13], and AD [14]. Additionally, the lethal dose (LD50) of Thymol and Carvacrol (565.7 mg/kg and 471.2 mg/kg, respectively) is higher compared to the therapeutic doses (Thymol 0.5 mg/kg, Carvacrol 1 mg/kg) which indicate efficacy and safety of these phytocompounds in improving cognitive impairments [15].
The reactive oxygen species (ROS) are highly active species with unpaired electrons which are generated as a by-product of oxygen metabolism. The elevated ROS levels have a detrimental effect on the cellular components, and as the neuronal cells are more sensitive to oxidative stress compared to others, the damage is greater. The generated ROS further accelerates mitochondrial impairment, inflammation, tau phosphorylation, neurofibrillary tangles formation, and apoptosis. Hence, shielding against oxidative stress is a significant approach in the treatment of NDs. The plants are a rich source of antioxidants and can scavenge free radicals. Consequently, the present work was planned to assess the neuroprotective properties of Carom fruit extract (hexane and ethyl acetate) and the main bioactive component, Carvacrol, on H2O2-induced oxidative stress in SH-SY5Y neuroblastoma cell lines. Moreover, additional neuroprotection mechanisms were also inspected by reviewing acetylcholine esterase (AChE) inhibition, Aβ1-42-oligomerization, and fibrillation inhibition activity, and anti-advanced glycation end products (AGEs) potential to explain the neuroprotective mechanism exerted by the Carom extract and bioactive compounds.
2. Materials and Methods
2.1. Chemicals
Most of the chemicals and standards for biochemical and cell-based studies were purchased from Sigma-Aldrich (St. Louis, MO, USA), namely acetyl thiocholine chloride, Acetylcholinesterase (Electrophorus electricus, Type VI-S), 6,6′-dinitro-3,3′-dithiodibenzoic acid, bis(3-carboxy-4-nitrophenyl) disulfide (DTNB), galantamine, bovine serum albumin (BSA), sodium azide, aminoguanidine, dextrose, 2′,7′-dichlorofluorescin diacetate (DCFDA), tetramethylrhodamine, ethyl ester (TMRE), hydrogen peroxide (H2O2), thioflavin T (ThT), gallic acid, ascorbic acid, 2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS), 2,2-diphenyl-1-picrylhydrazyl (DPPH), 2,4,6-tripyridyl-s-triazine (TPTZ), and Folin–Ciocalteu reagent (FCR).
Thermo Fisher Scientific (Waltham, MA, USA) was the source for fetal bovine serum (FBS), kanamycin, penicillin, and phosphate-buffered saline (PBST). Dulbecco’s modified Eagle’s medium (DMEM) was supplied by Gibco (Thermo Fisher, Seoul, Republic of Korea). Carvacrol and Thymol were purchased from TCI (Chuo-ku, Tokyo, Japan). The suppliers for others: Aβ1-42 (AnaSpec, Fremont, CA, USA), WST-8 kit (Roche Diagnostics GmbH, Mannheim, Germany), Aβ1–42 for MDS (GenicBio Inc., Shanghai, China), purified anti-Aβ1–16 antibody (Biolegend, San Diego, CA, USA), and the horseradish peroxidase (HRP)-conjugated W0-2 monoclonal antibody (Peoplebio Inc., Seongnam, Republic of Korea). The HPLC-grade organic solvents were bought from Sigma-Aldrich.
2.2. Plant Material and Extraction
The Carom seeds (Expat Mart, Seoul, Republic of Korea) were weighed, powdered, and extracted sequentially with n-hexane and ethyl acetate. The extracted fractions were dried, weighed, and stored at 4 °C for the experiments.
2.3. Gas Chromatography–Mass Spectrometry (GC-MS) Method
The hexane and ethyl acetate fractions (1 μL, 1 mg/mL) were analyzed on a fused-silica capillary column (DB-5 ms UI, 30 m × 0.25 mm i.d., film thickness 0.25 μm, Agilent, Santa Clara, CA, USA) connected to GCMS-QP2020 (Shimadzu, Kyoto, Japan). The oven temperature was set (60 °C/2 min, 100 °C at 4 °C/min, 290 °C at 10 °C/min, isothermic for 10 min). The carrier gas, helium, was at a constant flow rate of 1 mL/min. The temperature was set for the injection port (280 °C), ion source (280 °C), and interface (150 °C). The ionization energy was set at 70 eV. The full scan mode (40–700 AMU) was used to obtain the mass spectra. The phytocompounds in the fractions were identified by comparing them with known compounds in the National Institute of Standards and Technology (NIST) library.
2.4. Determination of Total Phenolic Content
The Folin–Ciocalteu method [16] with modifications was used to evaluate the phenolic content. The extracts were incubated for 5 min at room temperature (RT) with 1 N FCR, after which a sodium carbonate solution (10%) was added. After incubating the 96-well plate for 2 h in the dark at RT, absorbance was measured at 765 nm using a multi-mode reader, Synergy-H1 BioTek, Agilent (Santa Clara, CA, USA). Calibration was performed using Gallic acid (10–200 mg/mL) as a standard, and the results were represented as a mg gallic acid equivalent (GAE)/g of the extract.
2.5. Determination of Total Flavonoids Content
The total flavonoids were estimated by a previously described protocol [17]. Briefly, aluminum chloride (10%), ethanol (96%), and sodium acetate (1 M) were added to the extracts. After mixing the reagents in a 96-well plate, incubation for 40 min at RT in the dark was performed, after which the absorbance at 415 nm was measured using a microplate reader (Synergy-H1 BioTek, Agilent, Santa Clara, CA, USA). Quercetin (10–100 μg/mL) was used as the standard to calculate the total flavonoids in the extracts, and the results were expressed in terms of mg quercetin equivalents per gram of sample (QE/g).
2.6. Determination of Antioxidant Capacity
2.6.1. 2,2′-Azino-bis (3-Ethylbenzothiazoline-6-Sulfonic Acid) [ABTS] Radical Scavenging Assay
The extracts were assessed for free radical scavenging activity using a previously reported method [18]. To produce ABTS radicals, ABTS (0.7 mM) and potassium persulfate (2.45 mM) were mixed in equal ratios. The mixture was incubated at RT in the dark for 30 min. The extract and ABTS radical solution were mixed and incubated in the dark for 30 min at RT. The absorbance was monitored at 734 nm using the microplate reader (Synergy-H1 BioTek, Agilent, USA). Ascorbic acid (100 μg/mL) served as a standard. The percentage ABTS+. scavenging activity was calculated using the formula:
where AB = absorbance of the blank and AE = absorbance of the extract.
% RSA = (AB − AE/AB) × 100
2.6.2. Free Radical Scavenging using a 2,2-Diphenyl-1-picrylhydrazylhydrate (DPPH) Radical Assay
The DPPH radical scavenging activity was assessed [19] by incubating the extract with ethanolic DPPH (120 μM) in the dark at RT for 30 min, and the absorbance was read at 515 nm (Multi-mode reader, Synergy-H1 BioTek, Agilent, Santa Clara, CA, USA). Ascorbic acid (0.1–10 μg/mL) was used as a positive control. Radical scavenging activity (RSA) was calculated using the following formula:
where AB = absorbance of the blank and AE = absorbance of the extract.
% RSA = (AB − AE/AB) × 100
2.6.3. Ferric-Reducing Antioxidant Potential (FRAP) Assay
The metal-chelating ability of the extracts was evaluated using the FRAP assay [20]. The extract was incubated with the FRAP reagent at RT for 30 min, and the absorbance was read at 593 nm (Multi-mode reader, Synergy-H1 BioTek, Agilent, USA). For the standard, FeSO4·7H2O (1 mM) was used, and the FRAP values were expressed as μM Fe2+/g.
2.7. Anti-Acetylcholinesterase Activity
The anti-AChE activity of the extracts/bioactives was measured [21] by incubating the extracts with AChE and 10 mM ATCC at 37 °C for 15 min. The absorbance was read at 412 nm (Multi-mode plate reader, Synergy-H1 BioTek, Agilent, USA) after adding a stopping reagent (DTNB). Galantamine was used as a positive control. The percent inhibition was calculated as follows:
where A1 is the absorbance without the inhibitor; A2 is the negative control without the inhibitor; B1 is the absorbance with the inhibitor; and B2 is the negative control with the inhibitor. The IC50 values were calculated using GraphPad Prism 10.0.
Percent Inhibition (I%) = [(A1 − A2) − (B1 − B2)]/(A1 − A2) × 100
2.8. Anti-Advanced Glycation End-Product (AGE) Activity
For the glycation reaction [22], the extract was incubated for 2 weeks at 37 °C with 100 mM and a pH 7.4 phosphate buffer [containing 50 mg/mL BSA, 0.5 M dextrose monohydrate, and 5 mM sodium azide]. Aminoguanidine served as a positive control in the assay. The fluorescence was read at Ex 370 nm/Ems 440 nm (Synergy-H1 BioTek, Agilent, Santa Clara, CA, USA), and the percent glycation inhibition was calculated as:
where FC and FT are fluorescence intensity in the absence and presence of the sample, respectively; the IC50 values were calculated using GraphPad Prism 10.0.
Inhibition (%) = [(FC − FT)/FC × 100]
2.9. Anti-Aβ1–42-Fibrilization Activity
The anti-Aβ1–42-fibrilization activity of the extracts/bioactives was monitored using a ThT assay [23]. The samples were incubated in the presence/absence of Aβ1–42 at 37 °C for 24 h. The samples were incubated with 100 μM ThT at 37 °C for 15 min. The fluorescence was monitored at Ex 450 nm/Ems 490 nm (Synergy-H1 BioTek, Agilent, Santa Clara, CA, USA). For the control, Phenol red (50 μM) was used. The Aβ1–42 aggregation inhibition was calculated as follows:
where FI and FC are the fluorescence intensity with and without the inhibitors, respectively.
Percent Inhibition (%) = [(1 − FI/FC) × 100]
2.10. Anti-Aβ1–42-Oligomerization Activity
The anti-Aβ1–42-oligomerization activity of the extracts/bioactives was monitored using a Multiple Detection System (MDS) as described previously [24]. Briefly, the extracts and Aβ1–42 were incubated at RT for different time points (0 h, 2 h, and 4 h). The samples were incubated on an anti-β-amyloid pre-coated plate for 1 h at RT. A HRP-conjugated W0-2 monoclonal antibody was added, and the plate was kept at RT for 30 min. Later, TMB was added, and the plate was incubated for 15 min at RT. The absorbance was read at 450 nm using a microplate reader (Victor3, PerkinElmer, CT, USA).
2.11. Cell Culture
SH-SY5Y human neuroblastoma cells (ATCC CRL-2266, Manassas, VA, USA) were maintained in DMEM media containing FBS (10%), kanamycin (1%), and penicillin (1%) with 5% CO2, and a 95% humidified atmosphere in the incubator set at 37 °C. The cells were passaged twice per week and used at 80–90% confluency.
2.11.1. Cell Viability Assay
The cell viability assay was conducted as previously described [24]. In short, 1 × 104 cells/well were seeded in 96-well plates and incubated for 24 h with various concentrations of extracts/bioactives with the final volume of 100 µL/well. The extracts/bioactives were removed from the cells and then washed twice with 1X PBS. The cells were incubated in a fresh medium containing a 10% WST-8 reagent for 2 h. The absorbance was read at 450 nm using a multi-plate reader (Synergy-H1 BioTek, Agilent, USA). The percent cytotoxicity was calculated as:
where AC = absorbance of the control cells and AT = absorbance of the treated cells.
Cytotoxicity % = (AC − AT)/(AC) × 100
2.11.2. Neuroprotective Activity Assay
The neuroprotective effect of extracts/bioactives on H2O2-induced oxidative stress in SH-SY5Y was evaluated [24]. The cells were seeded 1 × 104 cells/well in a 96-well plate and incubated for 24 h with various concentrations of extracts/bioactives with a final volume of 100 µL/well. The extracts/bioactives were drawn out from the cells and incubated with 100 μM H2O2 for 6 h. The cell viability was evaluated using a WST-8 reagent (as mentioned in Section 2.11.1).
2.11.3. Measurement of Intracellular Reactive Oxygen Species
The ROS was measured using H2DCFDA dye as previously described [24]. H2DCFDA is a nonpolar dye that easily diffuses into cells and is hydrolyzed to 2′,7′-dichloro dihydrofluorescin (DCFH) via intracellular esterase. DCFH is oxidized to 2′,7′-dichlorofluorescein (DCF), a highly fluorescent compound, due to ROS production in the cells. Thus, the fluorescence intensity is proportional to the amount of hydrogen peroxide/ROS produced by the cells. The 1 × 104 cells/well were plated and incubated in a 96-well plate for 24 h with various concentrations of extracts/bioactives with a final volume of 100 µL/well. The extracts/bioactives were drawn out from the cells and incubated with 100 μM H2O2 for 4 h. The cells were then incubated with 25 μM H2DCFDA dye in the dark for another 2 h at 37 °C. The fluorescence intensity (Ex 495 nm/Ems 520 nm) was monitored using a multi-mode microplate reader (Synergy-H1 BioTek, Agilent, USA). The ROS was calculated as a percentage of the untreated control cells (100%) in triplicate measurements.
2.11.4. Mitochondrial Membrane Potential (ΔΨm) Assay
TMRE staining was used to evaluate the mitochondrial membrane potential [25]. The cells were seeded (1 × 104 cells/well) in a 96-well plate and pre-treated with the extracts/bioactives for 12 h. After removing extracts/bioactives from the wells, the cells were incubated with H2O2 (200 μM) for 2 h. After the incubation, H2O2 was removed, and the cells were treated with 1 μM TMRE for 1 h at 37 °C. The fluorescence intensity (Ex 549 nm/Ems 575 nm) was monitored using a multi-mode microplate reader (Synergy-H1 BioTek, Agilent, USA). The ΔΨm was expressed as a percentage of the untreated control cells (100%).
2.12. Statistical Analysis
Statistical investigation was performed using a one-way analysis of variance (ANOVA) followed by Dunnett’s post hoc test. Data were reported as the mean ± SD of three experiments. Data were considered to be significant ####/**** p < 0.0001; ###/*** p < 0.001; ##/** p < 0.01; and #/* p < 0.05. The symbol # indicates significance compared to the H2O2 control, whereas * indicates significance compared to the untreated control. The IC50 values were calculated using non-linear regression. The Michaelis–Menten plot using a non-linear fit via GraphPad Prism 10.0 was used to calculate Vmax and Km values. Lineweaver–Burk plots were designed via linear regression on GraphPad Prism 10.0.
3. Results and Discussion
3.1. Phytochemical Evaluation and Antioxidant Ability of Carom Extract
The plant’s secondary metabolites (phenols and flavonoids) have vital roles in plant growth and communication besides having versatile medicinal benefits for human health. Lately, the contribution of total phenolic (TPC) and flavonoids (TFC) in promoting antioxidant activity has been highlighted [26]. In the present study, colorimetric assays were carried out to estimate TPC and TFC in the extracts. The TPC in Carom-H (Hexane) and Carom-EA (Ethyl acetate) extracts was assessed as 11.70 ± 0.32 mg GAE/g and 5.61 ± 0.04 mg GAE/g, respectively. The TFC in the Carom-H (9.79 ± 0.06 mg QE/g) was lower than the Carom-EA (14.3 ± 1.09 mg QE/g).
Various assays (DPPH, ABTS, and FRAP) were conducted to estimate the antioxidant potential of the extracts. The percent radical scavenging activity using the DPPH assay was 46.01 ± 1.07% and 32.58 ± 0.89% for Carom-H and Carom-EA, respectively. A similar trend was observed in the radical scavenging ABTS assay with 55.89 ± 0.45% (Carom-H) and 40.15 ± 0.34% (Carom-EA). The FRAP assay measures the reduction in Fe3+ to Fe2+ in the presence of antioxidants. Similar FRAP values were obtained for Carom-H (1.56 ± 0.06 mM Fe2+/g) and Carom-EA (1.25 ± 0.03 mM Fe2+/g). A positive correlation between phenolic content and the antioxidant potential of Carom-H was observed in this study.
TPC values varied from 16.52 to 43.2 mg GAE/g and TFC from 3.89 to 8.03 mg QE/g in different varieties of methanolic Carom extracts [27]. The antioxidant activities quantified in the same study displayed 6.23–10.31 mM Fe2+ in the FRAP assay and 65–80% radical scavenging activity in the DPPH assay [28]. The FRAP values varied from 0.67, 0.71, and 2.27 mM Fe2+/L for aqueous, methanolic, and acetone extract, respectively [28]. Thus, the existence of phenolic monoterpenes (Thymol and Carvacrol) in the extracts could be the reason for antioxidant activity [29]. These compounds possess good reducing potential and free radical scavenging capacity besides preventing hydroperoxydiene formation in the initial steps of lipid degradation [30].
3.2. GC–MS Analysis of Carom Extract
To identify the phytochemicals in dried Carom fruit extracts (Hexane and Ethyl acetate), GC–MS analysis was carried out. The chromatogram identified Phenol, 2-methyl-5-(1-methylethyl)-(Syn. Carvacrol) as the major peak in both hexane (87.17%) and ethyl acetate (80.15%) extracts at 11.5 min (Supplementary Figure S1) while the other compounds were present in less than 2% (hexane) and 4% (ethyl acetate) extracts.
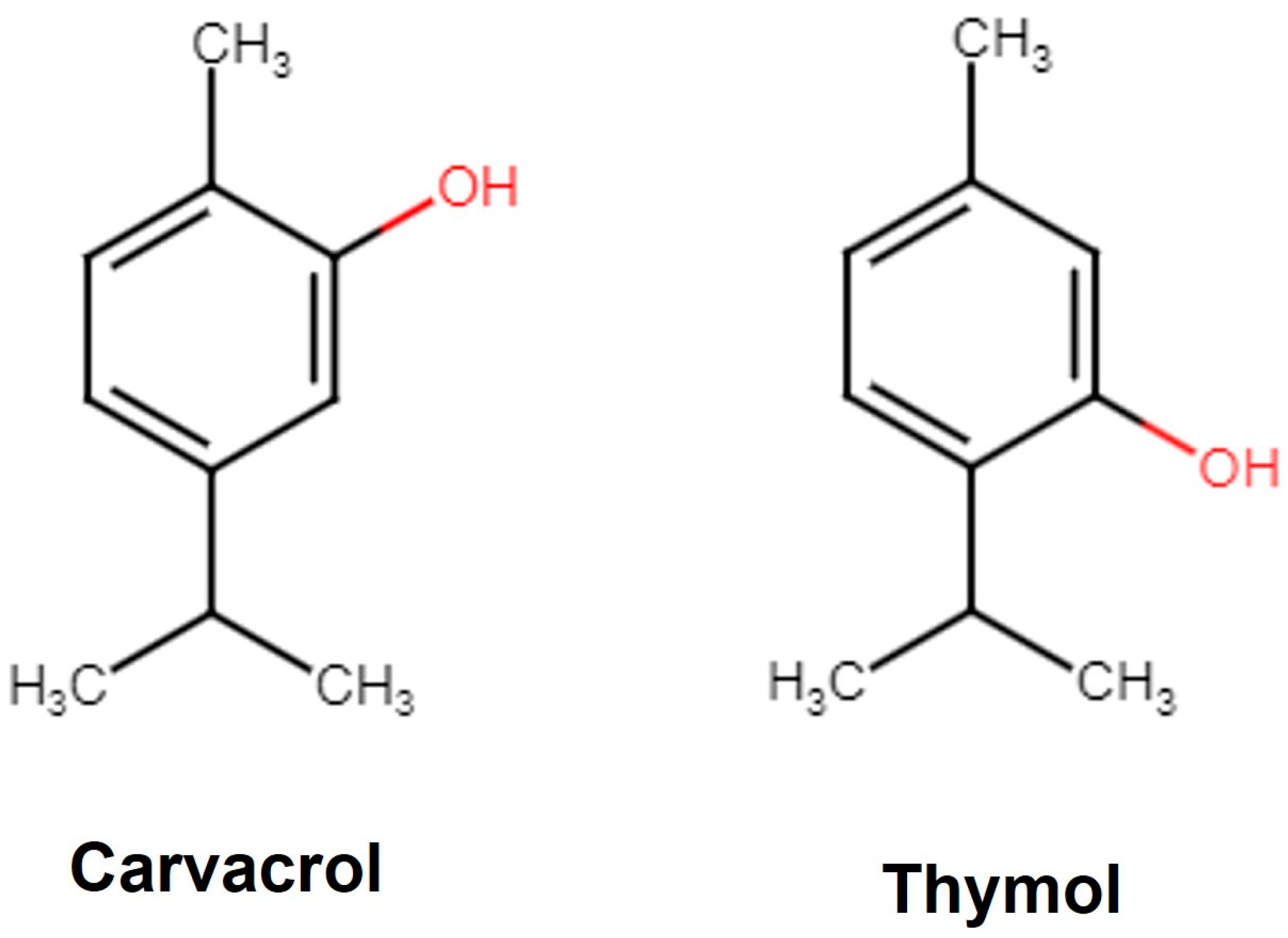
Carvacrol is a phenolic monoterpene derivative of Cymene (Figure 1) with a characteristic pungent odor and is a component of herbs like thyme, sage, and oregano. In plants, Carvacrol is synthesized by the mevalonate pathway, where the mevalonic acid is converted to γ-terpinene and forms Carvacrol via p-Cymene hydroxylation in the following steps [31]. Carvacrol is reported to exhibit anticancer [32], antidiabetic [33], anti-obesity [34], anti-inflammatory [35], antimicrobial [36], anti-asthmatic [37], and antiaging [38] properties. Carvacrol has a drug-like favorable pharmacokinetic and physiochemical profile (MW 150.22; logarithm of partition coefficient [log P 3.81]; hydrogen bond acceptor 1 [HBA 1]; hydrogen bond donor 1 [HBD 1]) including gastrointestinal (GI) absorption, blood–brain barrier (BBB) penetration [39], and inhibition of the Cytochrome (CYP450) complex [40]. The Food and Drug Administration (FDA) and the European Council have approved Carvacrol as a food flavoring agent [41]. Thymol [2-isopropyl-5-methylphenol] is an isomer of Carvacrol (Figure 1) with several therapeutic properties like anti-inflammatory, anti-fungal, and antioxidant [42]. It is also recognized as a safe food flavoring agent by the FDA [43].
Figure 1.
Structure of Carvacrol and Thymol. Carvacrol is the phenol isomer of Thymol.
3.3. In Vitro Anti-Acetylcholinesterase Activity
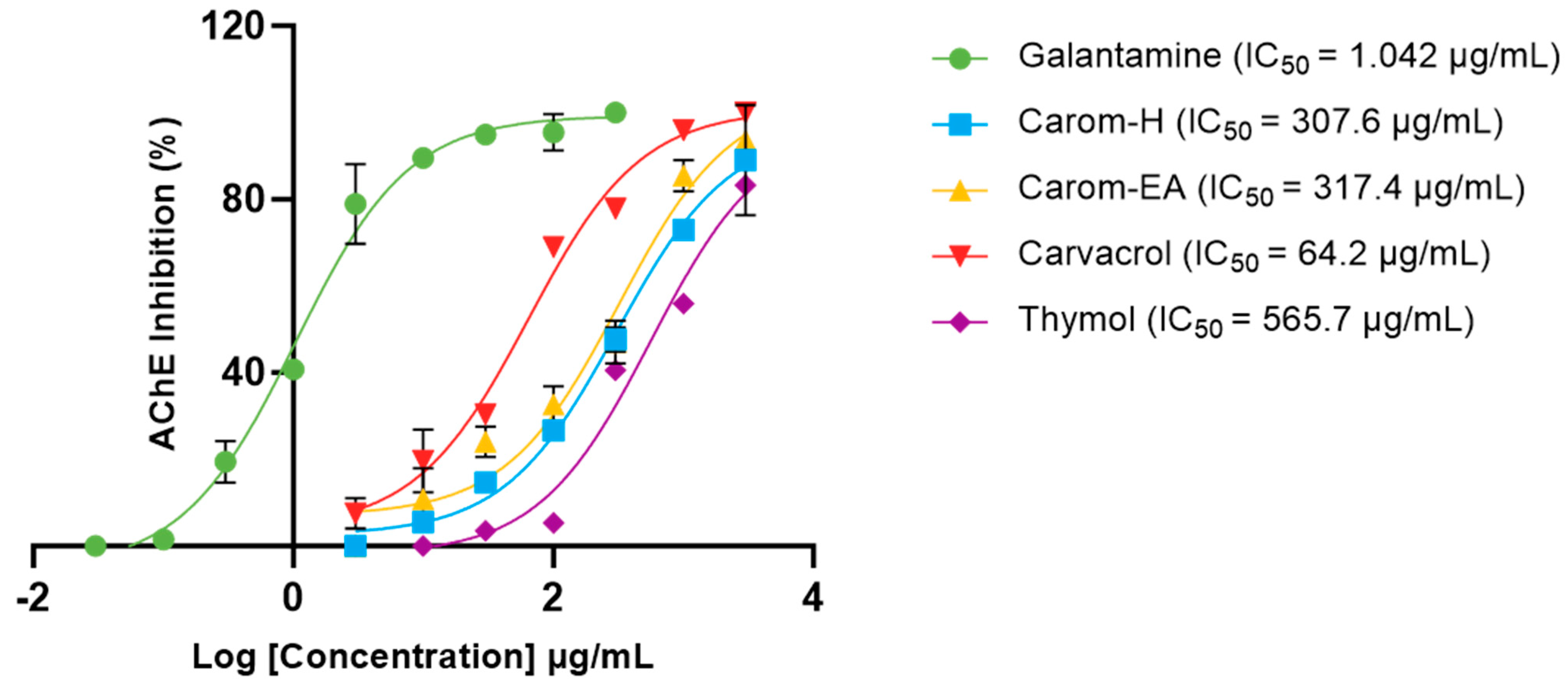
Acetylcholinesterase (AChE; E.C.3.1.1.7) is a cholinergic enzyme generally present at the postsynaptic neuromuscular junctions. It hydrolyses acetylcholine (ACh), an important neurotransmitter that plays an essential role in the cholinergic signaling pathway. This hydrolysis brings cholinergic neurons to return to the resting state [44]. In AD, the ACh level declines in the synaptic junction; hence, inhibition of AChE is desirable to maintain normal ACh levels. Therefore, the extracts and Carvacrol were examined for anti-AChE activity, and the IC50 values (half-maximal inhibitory concentration) were calculated using Galantamine hydrobromide as the inhibitor control. The IC50 values of extracts/bioactives were 307.6 μg/mL (Carom-H), 317.4 μg/mL (Carom-EA), and 64.21 μg/mL (Carvacrol) (Figure 2). We have also calculated IC50 of Thymol (565.7 μg/mL), an isomer of Carvacrol (Figure 1). The IC50 value of Galantamine was 1.042 μg/mL, similar to an earlier value of 1.78 μg/mL [45]. The IC50 value for Carvacrol obtained in our experiment was similar to previously obtained values of 288.26 μM (43.29 μg/mL) [46] and 91.7 μg/mL [29]. However, a recent study reported a lower IC50 value (3.8 μg/mL) for Carvacrol and Galantamine (0.6 μg/mL) [47].
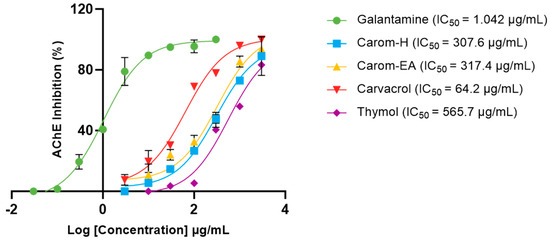
Figure 2.
IC50 curves of Carvacrol, Thymol, Carom-H, and Carom-EA extracts against Acetylcholinesterase (AChE) from Electrophorus. Galantamine was used as a standard inhibitor control. The IC50 values were calculated using GraphPad Prism 10.0.
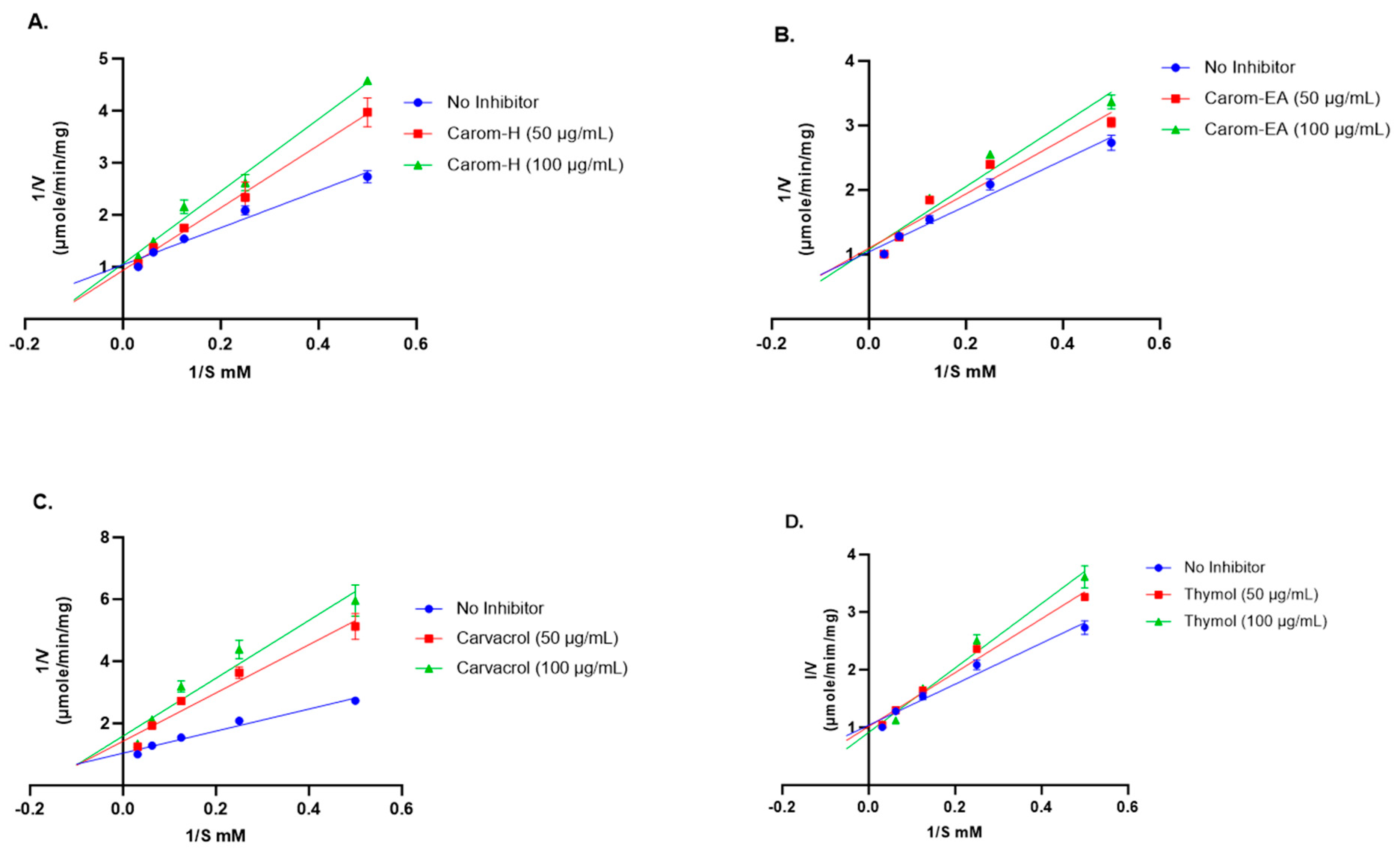
The mode of inhibition of the Carom extracts, Carvacrol, and Thymol was analyzed using the Lineweaver–Burk plot. The Vmax (the maximum reaction rate when the enzyme is saturated with substrate), Km (the substrate concentration that enables the enzyme to reach half Vmax), and inhibition pattern were summarized in Table 1.

Table 1.
AChE inhibition kinetic parameters for Carom extracts/bioactives.
Competitive inhibition was observed via the Carom extracts, Carvacrol, and Thymol (Figure 3). From the results obtained (Table 1), the value of Vmax with extracts/Carvacrol did not deviate much from the Vmax value of the no-inhibitor (1.096 μmol/min/mg). On the other hand, the Km value increased in the presence of extracts, Carvacrol, and Thymol as compared to no-inhibitor (5.02 mM). This trend indicates a competitive inhibition where the substrate molecule and inhibitor compete for binding with the enzyme’s active site. In the presence of an inhibitor, amplified Km decreases the enzyme’s affinity for the substrate; hence, a greater substrate concentration is needed to attain Vmax. However, in the Lineweaver–Burk plot, Carvacrol exhibited mixed inhibition (combination of competitive and uncompetitive inhibition) (Figure 3), where it can bind to both a free enzyme (E) as well as a substrate-bound enzyme (ES). The higher Km value obtained in the presence of Carvacrol indicates that it occupies the enzyme active site for a more extended period, reducing the enzyme’s affinity for the substrate, which makes it the best inhibitor.
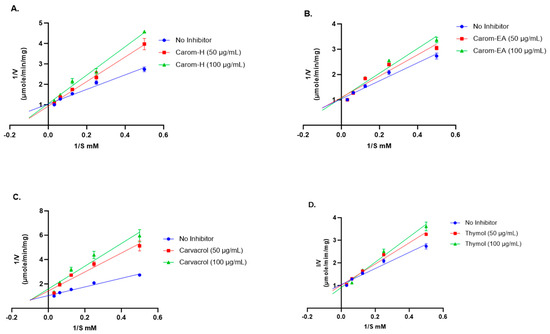
Figure 3.
Lineweaver-Burk plots of Acetylcholinesterase in the presence of 50 μg/mL and 100 μg/mL of Carom-H (A), Carom-EA (B), Carvacrol (C), and Thymol (D). The graphs were plotted using GraphPad Prism 10.0. Abbreviations: V: velocity of enzyme-catalyzed reaction; S: substrate.
The IC50 values obtained were similar for the hexane and ethyl acetate extracts of Carom, suggesting the involvement of Carvacrol, the major component, in anti-AChE activity. The IC50 of Carvacrol was better than the extracts because it was a single purified compound. However, a significant difference was observed in the IC50 values of Carvacrol and Thymol, even though they were isomers. Previously, Carvacrol exhibited 10 times stronger anti-AChE activity compared to its isomer, Thymol [48,49]. This indicates the importance of the hydroxyl group at the para-position (as in Carvacrol) for effective binding to the AChE active site. The molecular docking studies showed that Carvacrol binds to the active site of human AChE through hydrogen bonding between the para-hydroxyl group and Asp74 side chain besides the π–π interaction with the side chain of Tyr341 [50]. In yet another in silico study, Carvacrol showed a docking score of −6.060 Kcal/mol and −6.986 Kcal/mol against AChE in Standard Precision (SP) and Extra Precision (XP) Glide docking, respectively. Additionally, Carvacrol interacted with Tyr334, Phe330, Trp432, Tyr442, Trp84, and His440 at the enzyme’s active site [50]. On the other hand, Thymol interacted with the active site’s Ser203 and His447 with a binding free energy (ΔGbind) of −18.49 kcal/mol [51].
3.4. In Vitro Anti-Glycation Potential of Carom
We monitored the in vitro anti-glycation potential of the Carom extracts, Carvacrol, and Thymol using a BSA-AGE fluorescence assay. At a concentration of 1 mg/mL, a weak anti-glycation effect via Carom-H (19.14 ± 0.75%), Carom-EA (21.22 ± 0.99%), Carvacrol (20.76 ± 1.47%), and Thymol (12.92 ± 0.6%) was seen. AGEs are known to be involved in the pathogenesis of several diseases. Plant extracts exert anti-glycation potential by reducing the interaction of proteins and sugars by interfering with the lysine residues in the protein. A previous study reported 13.53% glycation inhibition via the aqueous extract [52], while the hydro-ethanolic extract exerted 80% inhibition at 1 mg/mL [53]. On the contrary, Carvacrol and Thymol displayed a frail anti-AGE activity compared to the methanolic plant extract [54]. Interestingly, the synergistic action of the Thymol and p-Cymene (2:1) mixture effectively reduced the glycation as compared to Thymol alone (5 mg/mL) [55]. The antiglycation activity might have resulted either from its antioxidant nature and/or by masking the sugar-binding site in the protein [55]. However, we observed a feeble anti-glycation effect of Carom extracts and pure compounds at 1 mg/mL, indicating that a higher concentration might be effective in exerting the potent effect.
3.5. Effect of Carom on Aβ-Fibrilization and Oligomerization
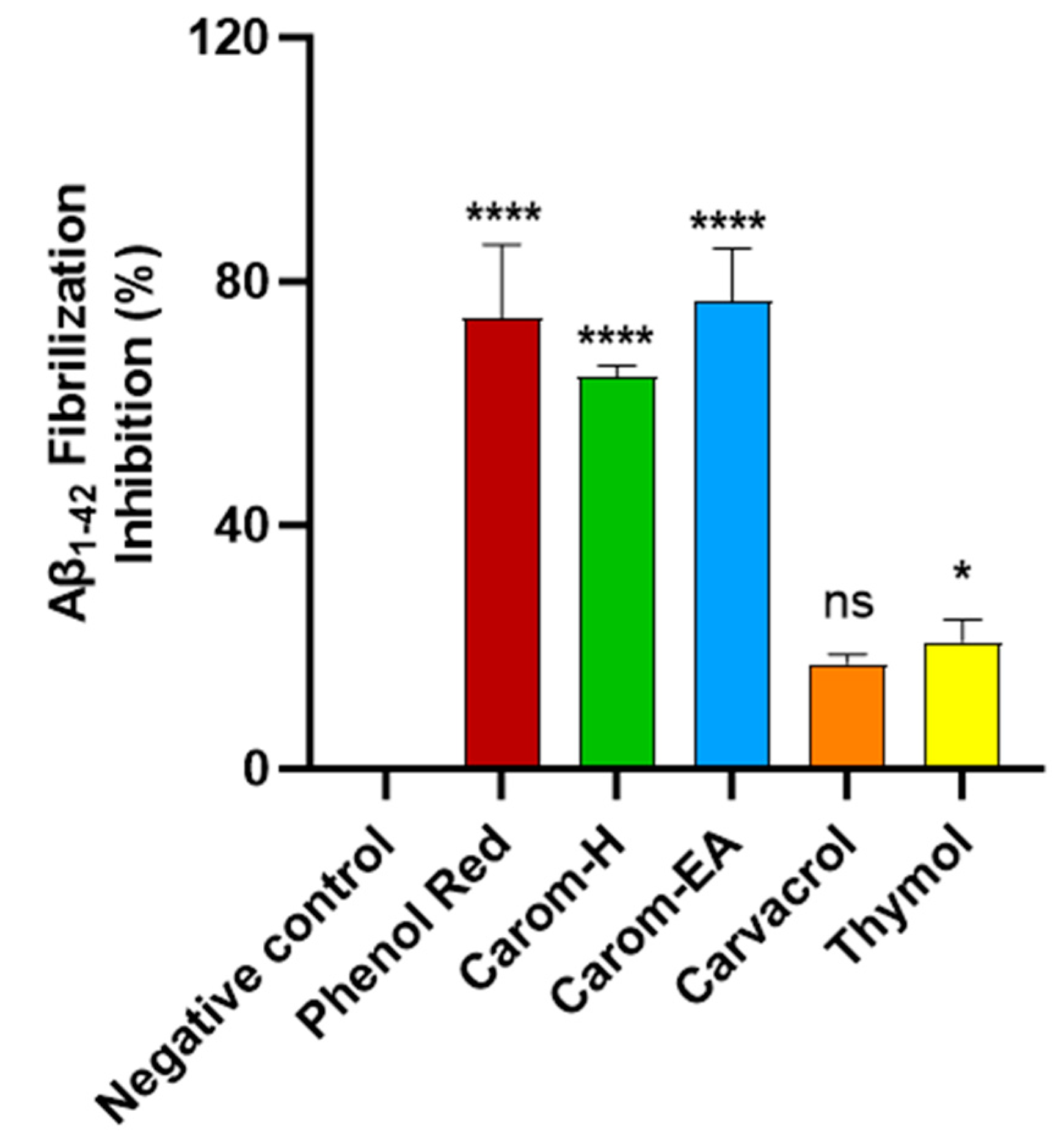
Protein aggregation to form amyloid fibrils is a general characteristic of diseases like AD, PD, and type 2 diabetes. Thioflavin T (ThT) is the commonly used probe to perceive in vitro amyloid fibrilization. The fluorescence of ThT increases upon binding to amyloid fibrils due to the rotational immobilization of the C–C bond linking the benzothiazole and aniline rings [56]. We screened the samples at 500 μg/mL where Carom-EA exerted the best Aβ-fibrilization inhibition (76.78 ± 8.63%; **** p < 0.0001), followed by Carom-H (64.38 ± 1.74%; **** p < 0.0001), Carvacrol (17.33 ± 1.55%; ns), and Thymol (20.91 ± 3.61%; * p < 0.05) (Figure 4). The positive control (Phenol red) exhibited 76.42 ± 7.98% at 50 μM, similar to a previously reported value [23].
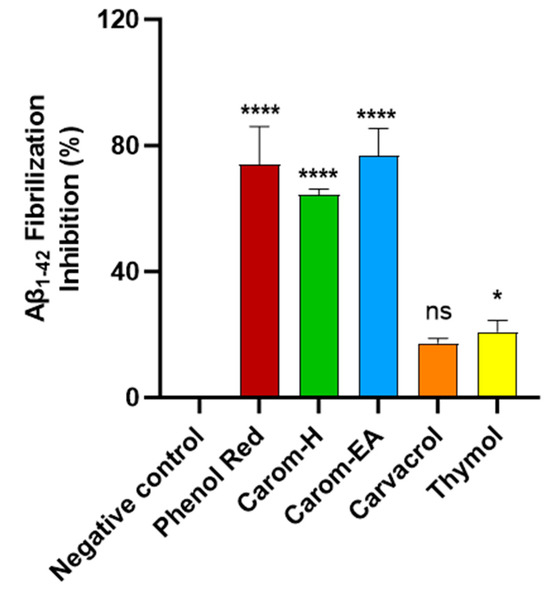
Figure 4.
Aβ-fibrilization inhibition in the presence of Carom extracts, Carvacrol, and Thymol at 500 μg/mL. Phenol Red (50 μM) served as a positive control. The values were expressed as the mean ± SD (n = 3). A significant difference (* p < 0.05) and (**** p < 0.0001) using one-way ANOVA followed by Dunnett’s post hoc was observed in the reduction in oligomerization vs. the negative control (buffer + Aβ). Statistically non-significant data were represented as ns.
In an earlier report, Carom seed aqueous extract exhibited a 30.39% reduction in β-amyloid aggregation in a ThT assay [52]. The π-stacking between Aβ aromatic side chains (especially diphenylalanine at the 19–20 position) is accountable for amyloid aggregation. The aromatic rings in the compounds might interact with diphenylalanine’s π stacking to exert anti-amyloidogenic activity [57]. The aromatic compounds are known to compete with polypeptide monomers for interaction with the extending fibrils, leading to fibrillization inhibition. Additionally, the OH groups probably perform an auxiliary role for effective interaction with β-sheet fibrils [58]. Therefore, it is reasoned that aromatic compounds in the extract stabilize the protein β structure through π-stacking or hydrophobic interaction [59]. However, groups other than the phenolic group are required for interactions with the protein for potent activity [60]. We suggest that in the present study, the mild fibrilization inhibition exhibited by Carvacrol (17.33%) and Thymol (20.91%) is due to the interaction of the phenolic ring with the growing Aβ. The lower inhibitory activity of pure compounds compared to the extract signifies the synergistic or additive involvement of other phytocompounds in the extracts for the anti-amyloidogenic property. Among the compounds identified by GC-MS (Figure S1), cis-9-Octadecenoic acid and carbamate derivative have been reported to modulate Aβ aggregation [61,62] and also possess antioxidant and anti-inflammatory activities [63]. However, the literature on the effect of other compounds on Aβ aggregation is lacking. Therefore, it can be speculated that their presence exerted a synergist effect to improve anti-fibrilization activity in the extracts compared to the pure compounds.
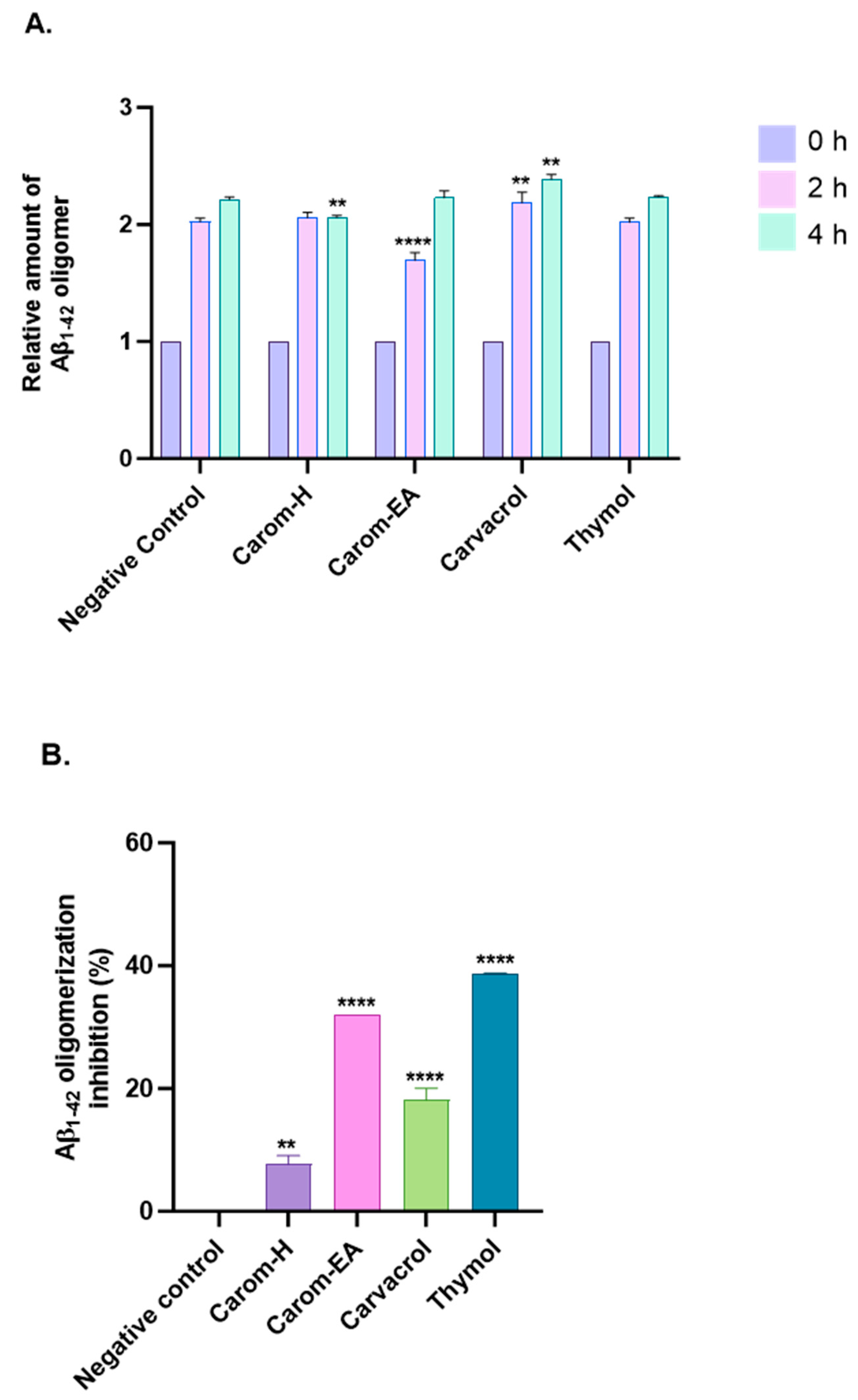
Aβ oligomerization inhibition via Carom extract and the bioactives was investigated using MDS. To evaluate the anti-Aβ oligomerization potential, the extracts/bioactives were treated with Aβ1–42 for different time intervals (Figure 5A and Supplementary Figure S2). The results were evaluated based on the 0 h value, which is why all sample signals were set at 1.0. The oligomerization reduction at 2 h was statistically significant in Carom-EA (**** p < 0.0001) and Carvacrol (** p < 0.01). Though a reduction in oligomerization was observed with Thymol, the p-value was insignificant. After 4 h of incubation, a statistically significant oligomerization reduction (** p < 0.01) was observed via Carom-H and Carvacrol. The compounds may inhibit Aβ oligomerization at different times due to structural differences affecting their interaction with the protein (Aβ). Moreover, degradation of compounds may occur with time, which this assay cannot observe.
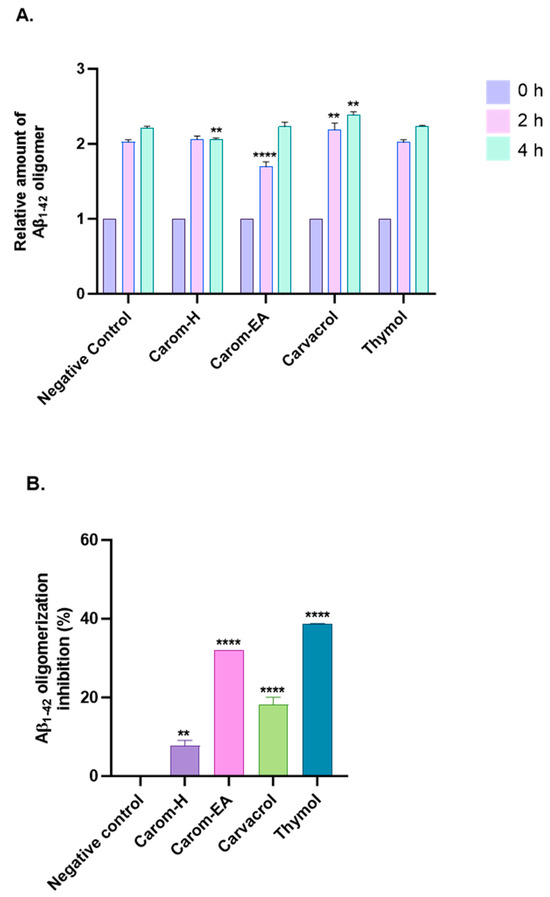
Figure 5.
Aβ-oligomerization inhibition in the presence of Carom extracts, Carvacrol, and Thymol. (A) The relative amount of Aβ oligomers at 0, 2, and 4 h. (B) value of Aβ oligomerization inhibition. All data are expressed as mean ± SEM (n = 3). A significant difference ** (p < 0.01), and **** (p < 0.0001) using the two-way ANOVA (A) and one-way ANOVA (B) followed by Dunnett’s post hoc was observed in the percent oligomerization reduction vs. the negative control (Buffer + Aβ).
The percentage inhibition of oligomerization is displayed in Figure 5B. Carom-EA and Carvacrol significantly inhibited Aβ oligomerization: 32.03% (**** p < 0.0001) and 18.12 ± 1.92% (**** p < 0.0001), respectively. However, Carom-H had a nominal effect (6.67 ± 0.02%; ** p < 0.01) on Aβ oligomerization inhibition (Figure 5B). The lower inhibition displayed via Carom-H could be due to higher concentrations of other metabolites in Carom-EA, which might synergistically enhance inhibition in Carom-EA. Interestingly, Thymol was more effective (38.67 ± 0.05%; **** p < 0.0001) than its isomer, Carvacrol, indicating the importance of the meta-hydroxyl group in oligomerization inhibition.
Amyloid aggregation is a crucial event in the pathogenesis of many neurodegenerative diseases. Therefore, inhibiting amyloid aggregation is an effective approach to treating/preventing such diseases. A recent study [64] investigated the role of central (ϕ1), C-terminal hydrophobic (ϕ2), and C-terminal end (ϕ3) of Aβ42 in the aggregation process. It was hypothesized that the ϕ1 domain drives Aβ42 amyloid aggregation, so compounds interacting with this domain suppress amyloid aggregation. The phenolic compounds are known to prevent Aβ oligomerization through the direct interaction of the phenolic hydroxyl group with the protein’s histidine or lysine side chain [65]. The compounds that inhibit Aβ assembly are classified into three categories: Class I (inhibit oligomerization but no effect on fibrilization), Class II (inhibit both), and Class III (inhibit fibrilization and no effect on oligomerization) [66]. According to the above classification, our compounds are Class II inhibitors, which stabilize Aβ conformations that do not facilitate either oligomers or fibril formation. Thymol’s better activity than Carvacrol indicates the advantage of the meta-substituted hydroxyl group in enhancing activity, as reported previously [67].
3.6. Non-Toxic Effect of Carom in SH-SY5Y Cells
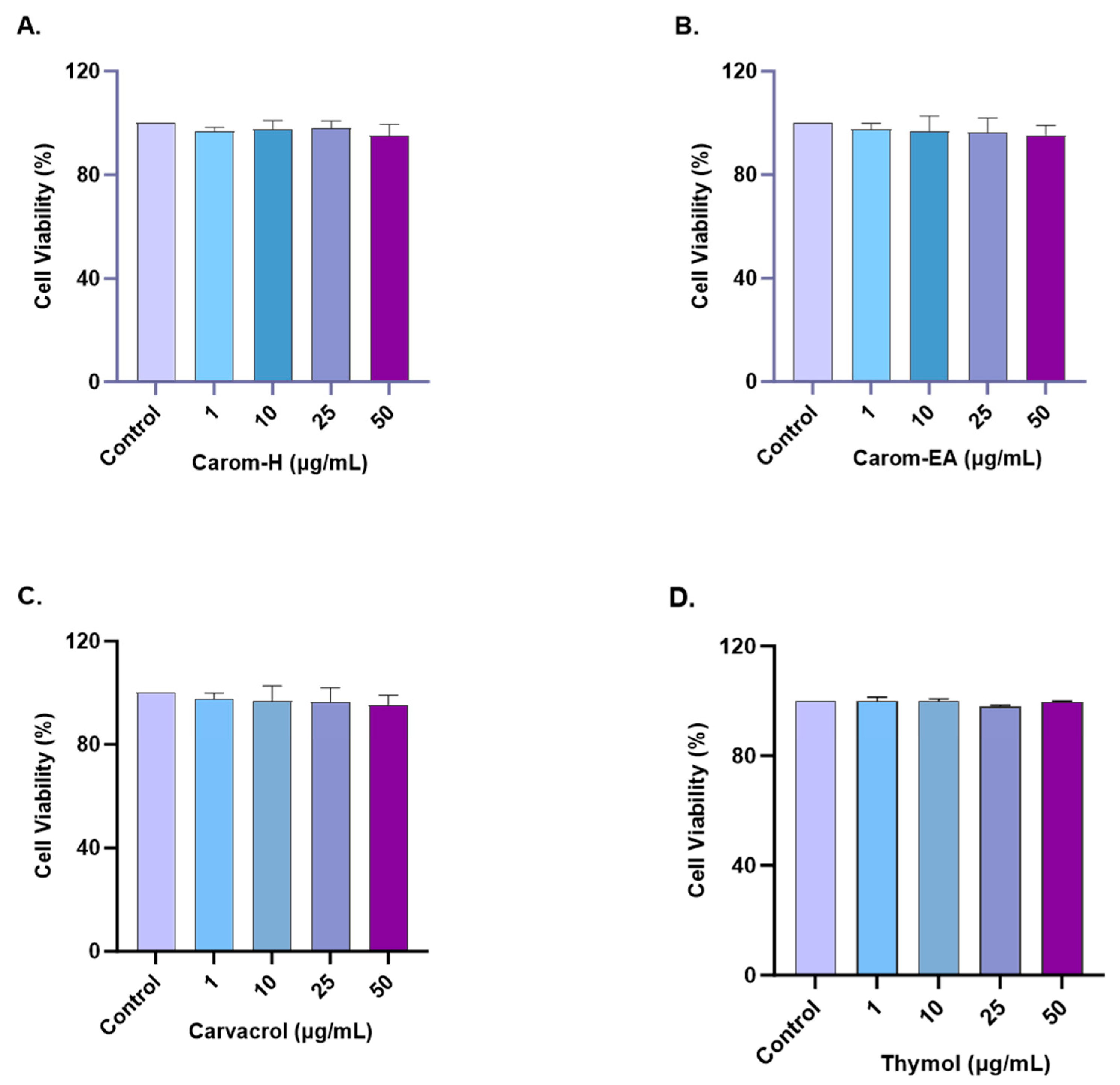
The cytotoxic effect of Carom on SH-SY5Y cells was monitored by treating cells with different concentrations (1, 10, 25, and 50 μg/mL) of the extracts and pure compounds for 24 h. The cell viability was measured using WST-8 dye. No statistically significant cytotoxicity was examined up to 50 μg/mL for both extracts and the pure compounds (Figure 6 and Supplementary Figure S3). Our results are supported by the previous findings where Carvacrol (200 μM) and Thymol (100 μM) produced no cytotoxicity in PC12 cells [68]. Carvacrol was non-cytotoxic, up to 100 mg/L in rat neurons and N2a neuroblastoma cells [69] and up to 333 μmol/L in SH-SY5Y cells [70]. Based on our results, further experiments were conducted up to 50 μg/mL concentrations of extracts/pure compounds.
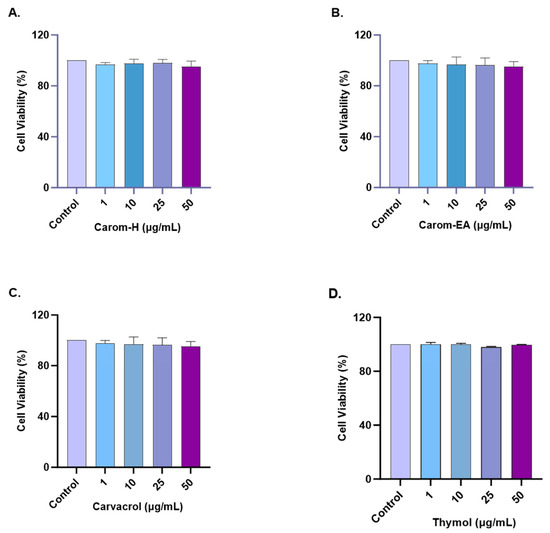
Figure 6.
Cytotoxicity assay of Carom-H (A), Carom-EA (B), Carvacrol (C), and Thymol (D) on the SH-SY5Y cells. The cells were treated with varying concentrations (1, 10, 25, and 50 μg/mL) of extract/bioactive for 24 h. The cell viability was calculated as the percentage of the control group (100%). The data were expressed as mean ± SD (n = 3). No significant difference was observed using a one-way ANOVA followed by Dunnett’s post hoc in the % cell viability vs. the control group (no treatment).
3.7. Neuroprotective Effect of Carom against H2O2-Induced Oxidative Stress in SH-SY5Y Cells
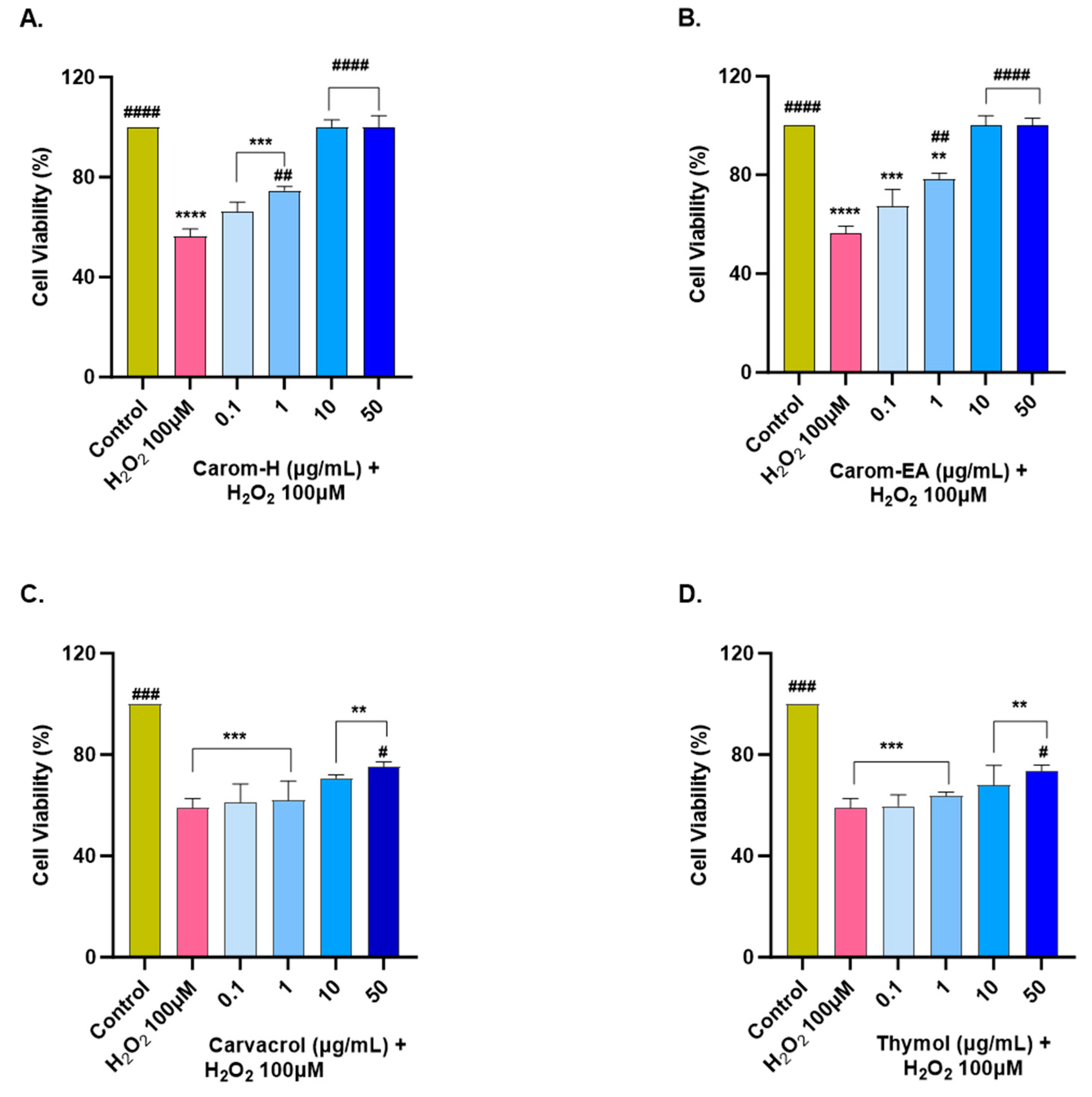
The extracts and pure compounds were assessed for neuroprotective potential in H2O2-induced oxidative stress in the SH-SY5Y cells. The concentration of H2O2 was optimized as 100 μM (that resulted in approx. 50% cell viability after 6 h treatment). The cells were pre-incubated with the extracts/bioactives at different concentrations (0.1, 1, 10, and 50 μg/mL) for 12 h, after which oxidative stress was induced by incubating the cells with 100 μM H2O2 for 6 h. A similar dose-dependent neuroprotection was seen in both extracts from 0.1 μg/mL to 10 μg/mL, after which the response was constant (Figure 7 and Supplementary Figure S4). The results were non-significant at the lowest concentration (0.1 μg/mL) but displayed significant neuroprotection (74.6 ± 1.61%, ## p < 0.01) in Carom-H, and (78.31 ± 2.41%, ## p < 0.01) in Carom-EA, at 1 μg/mL. The neuroprotective effect was more significant at 10 and 50 μg/mL (100%, #### p < 0.0001).
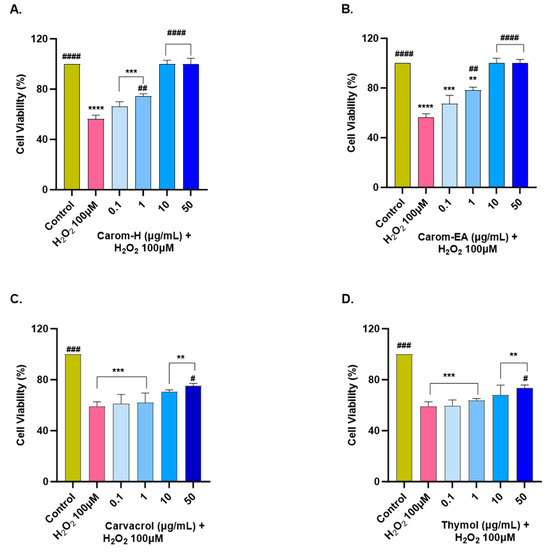
Figure 7.
Neuroprotective effect of Carom-H (A), Carom-EA (B), Carvacrol (C), and Thymol (D) in H2O2-induced oxidative stress in neuroblastoma SH-SY5Y cells. The SH-SY5Y cells were pre-treated with various concentrations of the extract/bioactives (0.1, 1, 10, and 50 μg/mL) for 12 h followed by 6 h of H2O2 (100 μM) treatment. The data were expressed as mean ± SD (n = 3). The results were displayed in terms of % cell viability vs. the control cells. A significant difference # (p < 0.05), **/## (p < 0.01), ***/### (p < 0.001), and ****/#### (p < 0.0001) using one-way ANOVA followed by Dunnett’s test was observed in the % cell viability vs. untreated cells (*) and H2O2 treated cells (#).
From the above results, both the extracts exerted the best neuroprotection at 10 μg/mL, after which the effect remains almost constant. A similar neuroprotective trend exerted by both extracts could be due to the main bioactive compound, Carvacrol. On the other hand, Carvacrol and Thymol presented statistically significant neuroprotection (# p < 0.05) at 50 μg/mL. The reason for the better neuroprotective effect of the extracts compared to the pure compounds could be the synergetic/additive effect of other phytocompounds present in the extract. Our results are supported by a previous finding where 10–50 μM of Carvacrol and Thymol significantly protected PC12 cells against Aβ25-35-induced oxidative stress [68]. Thus, the neuroprotective action of Carom/bioactives might be due to the attenuation of oxidative stress. The better neuroprotection by extracts compared to the pure compounds could be the result of synergistic/additive action of other minor phytocompounds in the extracts. Among the identified phyto-compounds by GC-MS (Figure S1), γ-terpinene [71], vinyltriphenylphosphonium bromide (VTPB) [72], and cis-9-Octadecenoic acid [63] displayed potent antioxidant activity in various in vitro and in vivo conditions, which might be responsible for better neuroprotection by the extracts.
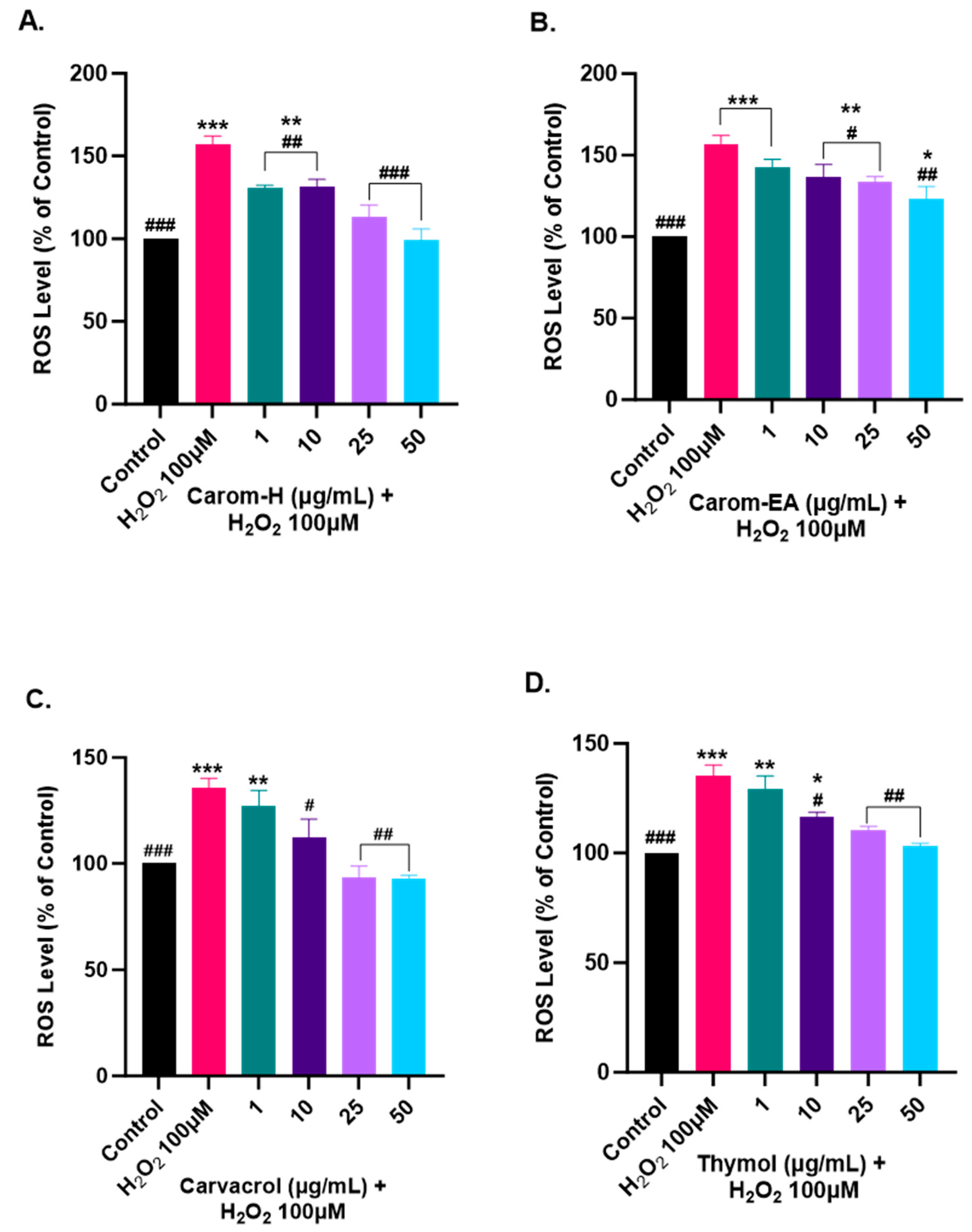
3.8. Alleviation of ROS Levels in SH-SY5Y Cells via Carom
Oxidative stress is generated due to the improper functioning of the antioxidant system and is a key modulator in aging and neurodegeneration. The ROS generation has adverse effects on the biomolecules, especially the neurons more prone to oxidative damage. Therefore, to study the protective effect of Carom extracts, Carvacrol, and Thymol, the SH-SY5Y cells were pre-incubated for 12 h with different concentrations of the extract/pure compounds followed by 4 h of H2O2 (100 μM) exposure. The ROS generation was monitored using H2DCFDA dye, which becomes oxidized in the presence of ROS to DCF, and the fluorescence was monitored at Ex 495 nm/Ems 520 nm.
Oxidative stress occurs when the cellular defense system cannot compensate for ROS production. The bioactive components are usually good antioxidants that help quench the ROS and maintain homeostasis. In our experiment, we used H2O2 to induce ROS in the cells as it can diffuse freely through cellular membranes due to its solubility in lipid and aqueous environments, affecting key cellular activities (growth, proliferation, and differentiation). We observed ~150% for the ROS level in H2O2 alone treated cells, while a dose-dependent decrease in the ROS level was observed in the cells pre-treated with the extracts/pure compounds. Carom-H is slightly more effective compared to Carom-EA (Figure 8 and Supplementary Figure S5). A statistically significant (130.48 ± 1.71%; ## p < 0.01) effect in reducing ROS was observed even at the lowest tested concentration (1 μg/mL) of Carom-H and almost complete alleviation of ROS level at 25 μg/mL (113.02 ± 7.41%; ### p < 0.001) and 50 μg/mL (99.12 ± 6.78%; #### p < 0.0001). On the other hand, with Carom-EA pre-treatment the ROS levels were 142.78 ± 4.63% (statistically non-significant) at 1 μg/mL, 136.77 ± 6.07% (# p < 0.05) at 10 μg/mL, 133.99 ± 2.39% (# p < 0.05) at 25 μg/mL, and 123.04 ± 7.68% (## p < 0.01) at 50 μg/mL. Both Carvacrol and Thymol displayed similar neuroprotective trends at 25 and 50 μg/mL (### p < 0.001), but Thymol displayed a better significant effect (## p < 0.01) than Carvacrol (# p < 0.05) at 1 μg/mL. The possible mechanism of a dose-dependent decrease in ROS levels signifies the extracts’ antioxidant nature, which activates the endogenous defense systems either by scavenging the free radicals or indirectly shielding them from oxidative stress [73]. The effective ROS alleviation by Carom-H could be related to better antioxidant activity compared to Carom-EA.
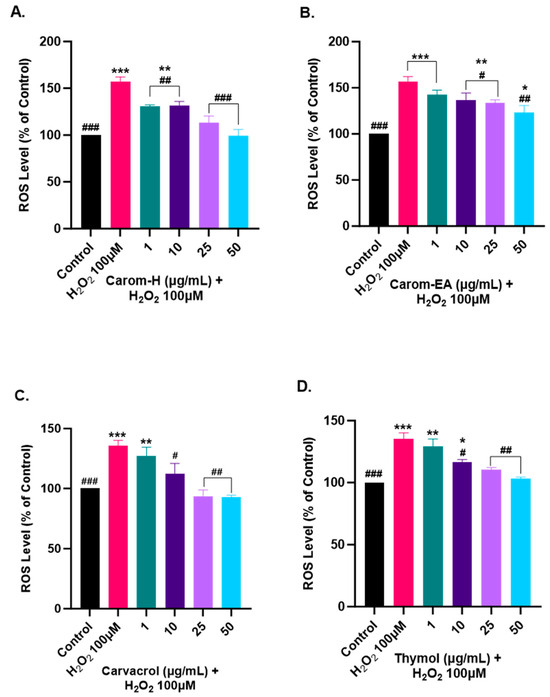
Figure 8.
Effect of Carom-H (A), Carom-EA (B), Carvacrol (C), and Thymol (D) on H2O2-induced ROS production in SH-SY5Y cells. The SH-SY5Y cells were pre-incubated for 12 h with varying concentrations (1, 10, 25, and 50 μg/mL) of the extracts/bioactives followed by 4 h H2O2 (100 μM) exposure. The data were expressed as mean ± SD (n = 3). The results were displayed in terms of % ROS level vs. the control cells (untreated cells). The data analysis was performed usin a one-way ANOVA followed by Dunnett’s test. A significant difference */# (p < 0.05), **/## (p < 0.01), and ***/### (p < 0.001), was observed in the % ROS vs. untreated cells (*) and H2O2 treated cells (#).
H2O2 and superoxide (O2•−) are the main ROS involved in the signaling pathways. The exogenous H2O2 is lethal to cells as it can easily be transformed into toxic ROS through the Fenton reaction. Additionally, the generation of hydroxyl radicals is enhanced in phosphate buffers in the Fenton reaction [74]. Oxidative stress occurs when ROS burdens the endogenous antioxidant defense and eventually results in various pathological conditions. Plants are a rich source of natural antioxidants that protect against the harmful effects of free radicals by strengthening the endogenous antioxidant defense and restoring the ideal equilibrium by counteracting free radicals [75]. Our results showed that pre-treatment of extracts/pure compounds provided protection against H2O2-induced oxidative stress in the cells via the enhancement of both non-enzymatic (GSH) and enzymatic (SOD, CAT, and GPX) endogenous antioxidant systems, which eventually protected the cells by mitigating the oxidative stress.
Alcoholic Carom extract has been reported to reduce ROS, boost antioxidant defense, and prevent apoptosis in PC12 cells [76]. In addition, extract and Thymol supplementation reduced oxidative stress, promoted neurogenesis, and reduced Aβ deposition in scopolamine-induced AD mouse model [9]. Thymol and Carvacrol are potential antioxidants that significantly reduce oxidative stress and ROS production in vitro [77]. Moreover, Thymol improves symptoms of neuropathic pain by reducing oxidative stress and cytokine release [10].
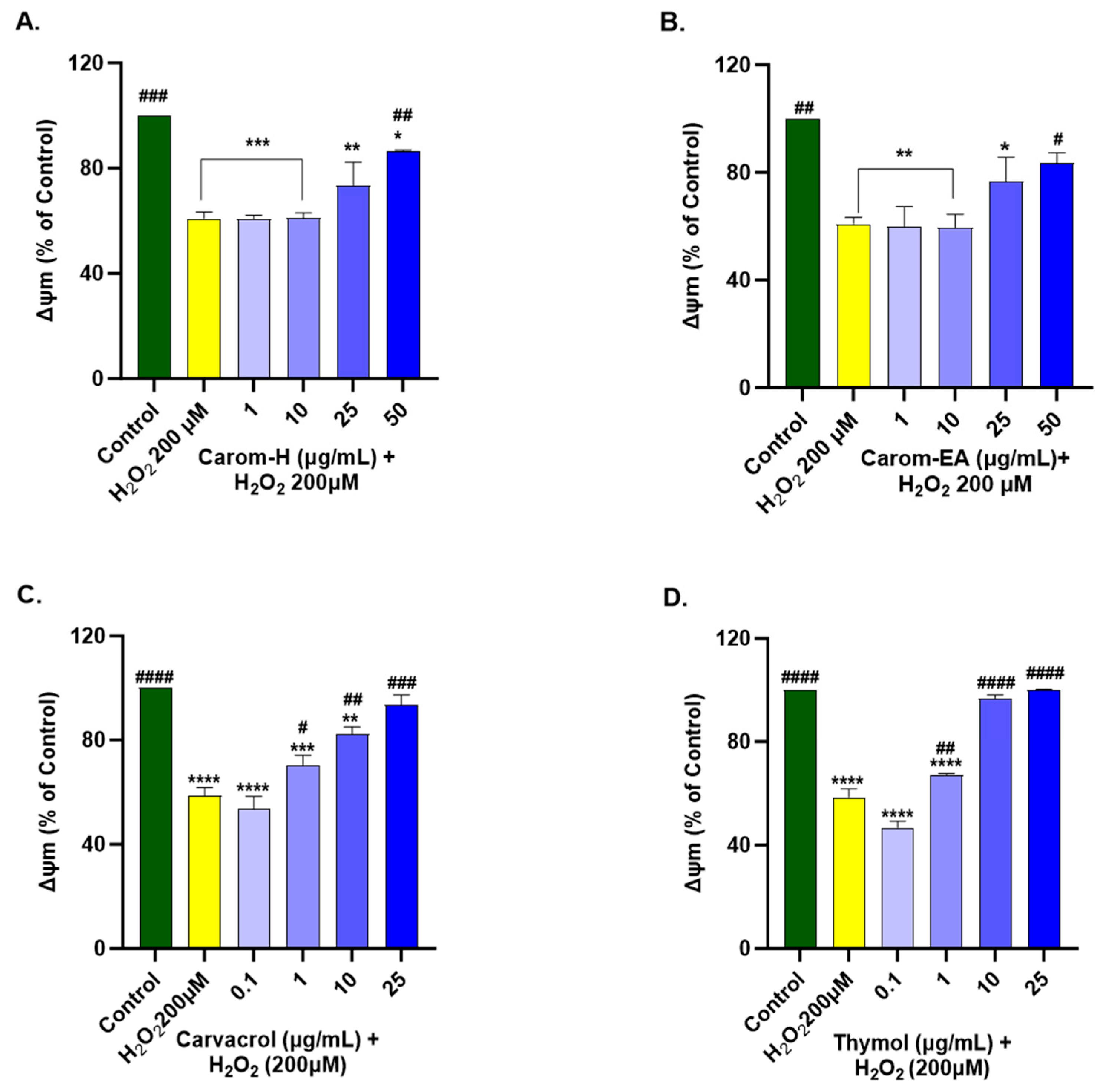
3.9. Protective Effect of Carom Extract on Mitochondrial Membrane Potential of SH-SY5Y Cells
ROS is produced in the mitochondria through electron transport, further stimulating proinflammatory cytokine production. Mitochondrial membrane potential (∆Ψm) is affected by oxidative damage to the cell, which disturbs membrane permeability by releasing Cytochrome C and/or pro-apoptotic factors in the cytoplasm. Hence, a decrease in MMP is viewed as a primary marker in events such as apoptosis and NDs.
We optimized the concentration of H2O2 (200 μM) and time of induction (2 h), which resulted in approx. 50% of cell death for the experiment. The H2O2 treatment decreased ∆Ψm by depolarizing the mitochondrial membrane. SH-SY5Y cells were treated for 12 h with Carom extracts, Carvacrol, and Thymol, after which the extracts were removed and incubated for 2 h with H2O2 (200 μM). Both the extracts exhibited a significant dose-dependent increase in MMP at 50 μg/mL in Carom-H (86.63 ± 0.28%; # p < 0.05) and Carom-EA (83.41 ± 3.95%; # p < 0.05). No significant effect was seen at the lower concentrations (1 and 10 μg/mL); however, a statistically non-significant increase (73.57 ± 8.8% in Carom-H; 76.68 ± 8.92% in Carom-EA) in MMP was seen at 25 μg/mL (Figure 9 and Supplementary Figure S6). In the case of pure compounds, statistically significant improvement in MMP was observed at 1 μg/mL, so a concentration lower (0.1 μg/mL) than 1 μg/mL was also studied. Carvacrol and Thymol exhibited a statistically significant dose-dependent increase in MMP, with Thymol displaying a better effect in restoring MMP.
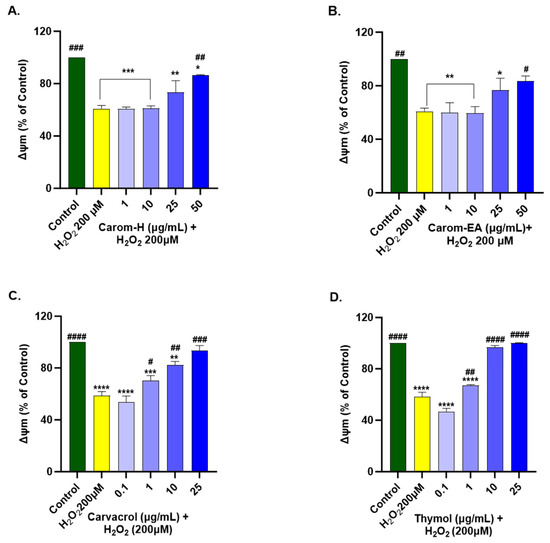
Figure 9.
Mitochondrial membrane potential in SH-SY5Y cells exposed to 200 μM H2O2 for 2 h after 12 h pre-treatment with Carom-H (A), Carom-EA (B) extracts at 1, 10, 25, and 50 μg/mL and Carvacrol (C), and Thymol (D) at 0.1, 1, 10, and 25 μg/mL. The results were expressed as mean ± SD (n = 3) in terms of % ∆Ψm vs. the control cells (untreated cells). The data analysis was performed using the one-way ANOVA followed by Dunnett’s test. A significant difference */# (p < 0.05), **/## (p < 0.01), ***/### (p < 0.001), and ****/#### (p < 0.0001), was observed in the % cell viability vs. untreated cells (*) and H2O2 treated cells (#). Abbreviations: ∆Ψm: mitochondrial membrane potential.
Mitochondrial dysfunction is central to several NDs as it increases ROS production, alters mitochondrial structure and permeability, decreases mitochondrial membrane potential, increases the release of Cytochrome C, increases Caspase (−3 and −9) expression, and finally, inflammation and apoptosis. Hence, protecting mitochondrial function using natural compounds is beneficial in preventing ROS/inflammation generation in NDs. Carvacrol exerted antioxidant and anti-inflammatory action to protect against H2O2-induced mitochondrial dysfunction in SH-SY5Y cells by modulating heme oxygenase-1/carbon monoxide/nuclear factor kappa B (HO-1/CO/NF-kB) signaling [78]. The heme degradation via the HO-1 enzyme generates CO, which in turn inhibits NF-kB, thus linking mitochondrial dysfunction and inflammation. Previously, the mitochondrial protective action of Carvacrol was observed in Fe2+ [70] and Cd2+ [79] induced toxicity in SH-SY5Y and PC12 cell lines, respectively. In addition, it also improved the mitochondrial homeostasis in 6-Hydroxydopamine hydrobromide (6-OHDA) - induced rat model and cultured SH-SY5Y cells by preventing superoxide formation [80]. Thymol also revoked mitochondrial dysfunction in myocardial infarction rat models [81] and mercuric chloride toxicity in hepatocellular carcinoma (HepG2) cells [82] through its antioxidant activity. However, detailed studies of Thymol in neuroblastoma cell lines are limited.
4. Conclusions
In the present work, different extracts of Carom were prepared, and major bioactive components were identified using GC-MS. Different cell-based and biochemical investigations were performed to assess the neuroprotective activity of the Carom extracts and Carvacrol. Additionally, Thymol (an isomer of Carvacrol) was also examined to study the role of the hydroxyl group in affecting various neuroprotective mechanisms. Oxidative stress, via H2O2 exposure, was generated in the neuroblastoma cell lines as it is an important generator of ROS. Oxidative stress is one of the key factors responsible for AD pathogenesis, as it accelerates abnormal protein aggregation, neuroinflammation, and neuronal death. Carom extracts provided effective neuroprotection by reducing ROS and restoring MMP in cell-based experiments, which could be attributed to their antioxidant behavior. Additionally, the extracts displayed anti-AChE, anti-oligomerization, and fibrilization activities. Comparing the two pure compounds, both displayed similar effects in Aβ-fibrilization inhibition, an effect on ROS, and neuroprotection. However, Carvacrol was found to be a stronger (~10 times) inhibitor of AChE and inhibited the enzyme through a mixed-type inhibition compared to Thymol (competitive inhibition). These results suggested the role of the para-substituted hydroxy group compared to the meta-substitute in AChE inhibition.
On the other hand, Thymol efficiently inhibited Aβ-oligomerization compared to Carvacrol, indicating the importance of the meta-substituted hydroxyl group. The extracts and pure compounds can be categorized as Class II inhibitors (inhibit both oligomerization and fibrilization), stabilizing Aβ conformations that do not facilitate oligomers or fibril formation. Thus, the present study demonstrates the potential of Carom as a multifunctional curative remedy for AD treatment, but further in vitro and in vivo experiments are required to elucidate the detailed pathways.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/antiox13010009/s1, Figure S1: Phytoconstituents identified in (A) Carom-H and (B) Carom-EA extracts using gas chromatography-mass spectrometry. Figure S2: Aβ-Oligomerization inhibition in the presence of Carom extracts, Carvacrol, and Thymol. Figure S3: Cytotoxicity assay of Carom extracts, Carvacrol, and Thymol on the SH-SY5Y cells. Figure S4: Neuroprotective effect of Carom extracts, Carvacrol, and Thymol in H2O2-induced oxidative stress in neuroblastoma SH-SY5Y cells. Figure S5: Effect of Carom extracts, Carvacrol, and Thymol on H2O2-induced ROS production in SH-SY5Y cells. Figure S6: Mitochondrial membrane potential in SH-SY5Y cells exposed to H2O2 after pre-treatment with Carom extracts and the pure compounds.
Author Contributions
Conceptualization, N.S. and S.S.A.A.; experimentation and data analysis, H.S., H.Y., and N.S.; writing—original draft preparation, N.S.; writing—review and editing, N.S., H.S., and S.S.A.A.; funding acquisition, S.S.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (grant number RS-2023-00251396 and 2021R1A6A1A03038996).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is provided in the Supplementary materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dugger, B.N.; Dickson, D.W. Pathology of Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, A.; Norton, N.; Fast, T.; Frölich, L.; Georges, J.; Holzapfel, D.; Kirabali, T.; Krolak-Salmon, P.; Rossini, P.M.; Ferretti, M.T. Global estimates on the number of persons across the Alzheimer’s disease continuum. Alzheimer’s Dement. 2023, 19, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Vos, T.; Lim, S.S.; Abbafati, C.; Abbas, K.M.; Abbasi, M.; Abbasifard, M.; Abbasi-Kangevari, M.; Abbastabar, H.; Abd-Allah, F.; Abdelalim, A. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association. 2023 Alzheimer’s Disease Facts and Figures. Alzheimer’s Dement 2023, 19, 1598–1695. [Google Scholar] [CrossRef] [PubMed]
- Mohd Sairazi, N.S.; Sirajudeen, K.N.S. Natural Products and Their Bioactive Compounds: Neuroprotective Potentials against Neurodegenerative Diseases. Evid.-Based Complement. Altern. Med. 2020, 2020, 6565396. [Google Scholar] [CrossRef] [PubMed]
- Zarshenas, M.M.; Moein, M.; Samani, S.M.; Petramfar, P. An overview on ajwain (Trachyspermum ammi) pharmacological effects; modern and traditional. J. Nat. Remedies 2013, 14, 98–105. [Google Scholar]
- Preedy, V.R.; Watson, R.R. Nuts and Seeds in Health and Disease Prevention, 2nd ed.; Academic Press: Cambridge, MA, USA, 2020; p. 598. ISBN 9780128185537. [Google Scholar]
- Singh, H.; Meghwal, M. Ajwain a potential source of phytochemical for better health. Pharma Innov 2019, 8, 599–603. [Google Scholar]
- Timalsina, B.; Haque, M.N.; Choi, H.J.; Dash, R.; Moon, I.S. Thymol in Trachyspermum ammi seed extract exhibits neuroprotection, learning, and memory enhancement in scopolamine-induced Alzheimer’s disease mouse model. Phytother. Res. 2023, 37, 2811–2826. [Google Scholar] [CrossRef]
- Sachan, N.; Saraswat, N.; Chandra, P.; Khalid, M.; Kabra, A. Isolation of Thymol from Trachyspermum ammi Fruits for Treatment of Diabetes and Diabetic Neuropathy in STZ-Induced Rats. Biomed. Res. Int. 2022, 2022, 8263999. [Google Scholar] [CrossRef]
- Lee, M.; Lee, S.H.; Choi, S.; Choi, B.Y.; Suh, S.W. Carvacrol Inhibits Expression of Transient Receptor Potential Melastatin 7 Channels and Alleviates Zinc Neurotoxicity Induced by Traumatic Brain Injury. Int. J. Mol. Sci. 2022, 23, 13840. [Google Scholar] [CrossRef]
- Haddadi, H.; Rajaei, Z.; Alaei, H.; Shahidani, S. Chronic treatment with carvacrol improves passive avoidance memory in a rat model of Parkinson’s disease. Arq. De Neuro-Psiquiatr. 2018, 76, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Manouchehrabadi, M.; Farhadi, M.; Azizi, Z.; Torkaman-Boutorabi, A. Carvacrol protects against 6-hydroxydopamine-induced neurotoxicity in in vivo and in vitro models of Parkinson’s disease. Neurotox. Res. 2020, 37, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Azizi, Z.; Majlessi, N.; Choopani, S.; Naghdi, N. Neuroprotective effects of carvacrol against Alzheimer’s disease and other neurodegenerative diseases: A review. Avicenna J. Phytomedicine 2022, 12, 371–387. [Google Scholar]
- Azizi, Z.; Ebrahimi, S.; Saadatfar, E.; Kamalinejad, M.; Majlessi, N. Cognitive-enhancing activity of thymol and carvacrol in two rat models of dementia. Behav. Pharmacol. 2012, 23, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Packer, L., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar] [CrossRef]
- Ribarova, F.; Atanassova, M.; Marinova, D.; Ribarova, F.; Atanassova, M. Total phenolics and flavonoids in Bulgarian fruits and vegetables. JU Chem. Met. 2005, 40, 255–260. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Koleva, I.I.; Van Beek, T.A.; Linssen, J.P.; Groot, A.d.; Evstatieva, L.N. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem. Anal. Int. J. Plant Chem. Biochem. Technol. 2002, 13, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Aktumsek, A.; Zengin, G.; Guler, G.O.; Cakmak, Y.S.; Duran, A. Antioxidant potentials and anticholinesterase activities of methanolic and aqueous extracts of three endemic Centaurea L. species. Food Chem. Toxicol. 2013, 55, 290–296. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Singh, P.; Jayaramaiah, R.H.; Agawane, S.B.; Vannuruswamy, G.; Korwar, A.M.; Anand, A.; Dhaygude, V.S.; Shaikh, M.L.; Joshi, R.S.; Boppana, R.; et al. Potential Dual Role of Eugenol in Inhibiting Advanced Glycation End Products in Diabetes: Proteomic and Mechanistic Insights. Sci. Rep. 2016, 6, 18798. [Google Scholar] [CrossRef]
- Tan, M.A.; Tan, B.L.U.; Nonato, M.G.; An, S.S.A. Neuroprotective effects on amyloid-beta induced cytotoxicity of Pandanus clementis Merr. 3 Biotech 2021, 11, 330. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Kim, D.Y.; Shim, K.H.; Sharma, N.; An, S.S.A. Multi-Targeting Neuroprotective Effects of Syzygium aromaticum Bud Extracts and Their Key Phytocompounds against Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 8148. [Google Scholar] [CrossRef] [PubMed]
- Alvariño, R.; Alonso, E.; Lacret, R.; Oves-Costales, D.; Genilloud, O.; Reyes, F.; Alfonso, A.; Botana, L.M. Caniferolide A, a macrolide from Streptomyces caniferus, attenuates neuroinflammation, oxidative stress, amyloid-beta, and tau pathology in vitro. Mol. Pharm. 2019, 16, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D. Girija. Evaluation of in vitro antioxidant and antimicrobial activities of various spices of Indian origin. Int. J. Pharm. Pharm. Sci. 2015, 8, 137–141. [Google Scholar]
- Goswami, N.; Chatterjee, S. Assessment of free radical scavenging potential and oxidative DNA damage preventive activity of Trachyspermum ammi L. (carom) and Foeniculum vulgare Mill. (fennel) seed extracts. Biomed. Res. Int. 2014, 2014, 582767. [Google Scholar] [CrossRef] [PubMed]
- Aazza, S.; Lyoussi, B.; Miguel, M.G. Antioxidant and antiacetylcholinesterase activities of some commercial essential oils and their major compounds. Molecules 2011, 16, 7672–7690. [Google Scholar] [CrossRef] [PubMed]
- Ruberto, G.; Baratta, M.T. Antioxidant activity of selected essential oil components in two lipid model systems. Food Chem. 2000, 69, 167–174. [Google Scholar] [CrossRef]
- Imran, M.; Aslam, M.; Alsagaby, S.A.; Saeed, F.; Ahmad, I.; Afzaal, M.; Arshad, M.U.; Abdelgawad, M.A.; El-Ghorab, A.H.; Khames, A. Therapeutic application of carvacrol: A comprehensive review. Food Sci. Nutr. 2022, 10, 3544–3561. [Google Scholar] [CrossRef]
- Güneş Bayır, A.; Kızıltan, H.Ş.; Koçyiğit, A. Dietary Interventions in Gastrointestinal Diseases: Foods, Nutrients, and Dietary Supplements, 1st ed.; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; p. 358. ISBN 9780128144688. [Google Scholar]
- Liu, Y.; Wei, J.; Ma, K.-T.; Li, C.-L.; Mai, Y.-P.; Qiu, X.-X.; Wei, H.; Hou, N.; Luo, J.-D. Carvacrol protects against diabetes-induced hypercontractility in the aorta through activation of the PI3K/Akt pathway. Biomed. Pharmacother. 2020, 125, 109825. [Google Scholar] [CrossRef]
- Spalletta, S.; Flati, V.; Toniato, E.; Di Gregorio, J.; Marino, A.; Pierdomenico, L.; Marchisio, M.; D’Orazi, G.; Cacciatore, I.; Robuffo, I. Carvacrol reduces adipogenic differentiation by modulating autophagy and ChREBP expression. PLoS ONE 2018, 13, e0206894. [Google Scholar] [CrossRef] [PubMed]
- Rolim, M.d.O.P.; de Almeida, A.R.; da Rocha Pitta, M.G.; de Melo Rêgo, M.J.B.; Quintans-Júnior, L.J.; Quintans, J.d.S.S.; Heimfarth, L.; Scotti, L.; Scotti, M.T.; da Cruz, R.M.D. Design, synthesis and pharmacological evaluation of CVIB, a codrug of carvacrol and ibuprofen as a novel anti-inflammatory agent. Int. Immunopharmacol. 2019, 76, 105856. [Google Scholar] [CrossRef] [PubMed]
- Scaffaro, R.; Maio, A.; Nostro, A. Poly (lactic acid)/carvacrol-based materials: Preparation, physicochemical properties, and antimicrobial activity. Appl. Microbiol. Biotechnol. 2020, 104, 1823–1835. [Google Scholar] [CrossRef] [PubMed]
- Ezz-Eldin, Y.M.; Aboseif, A.A.; Khalaf, M.M. Potential anti-inflammatory and immunomodulatory effects of carvacrol against ovalbumin-induced asthma in rats. Life Sci. 2020, 242, 117222. [Google Scholar] [CrossRef] [PubMed]
- El-Far, A.H.; Mohamed, H.H.; Elsabagh, D.A.; Mohamed, S.A.; Noreldin, A.E.; Al Jaouni, S.K.; Alsenosy, A.A. Eugenol and carvacrol attenuate brain d-galactose-induced aging-related oxidative alterations in rats. Environ. Sci. Pollut. Res. 2022, 29, 47436–47447. [Google Scholar] [CrossRef] [PubMed]
- Sisti, F.M.; Dos Santos, N.A.G.; do Amaral, L.; Dos Santos, A.C. The neurotrophic-like effect of carvacrol: Perspective for axonal and synaptic regeneration. Neurotox. Res. 2021, 39, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Bhardwaj, M.; Shukla, S.; Min, S.H.; Choi, D.K.; Bajpai, V.K.; Huh, Y.S.; Kang, S.C. Carvacrol inhibits cytochrome P450 and protects against binge alcohol-induced liver toxicity. Food Chem. Toxicol. 2019, 131, 110582. [Google Scholar] [CrossRef]
- A Food Additive Database. Center for Food Safety and Applied Nutrition, Office of Premarket Approva. 2004. Available online: https://www.fda.gov/food/food-additives-petitions/substances-added-food-formerly-eafus (accessed on 5 September 2023).
- Nagoor Meeran, M.F.; Javed, H.; Al Taee, H.; Azimullah, S.; Ojha, S.K. Pharmacological Properties and Molecular Mechanisms of Thymol: Prospects for Its Therapeutic Potential and Pharmaceutical Development. Front. Pharmacol. 2017, 8, 380. [Google Scholar] [CrossRef]
- FDA. Substances Added to Food. 2023. Available online: https://www.accessdata.fda.gov/scripts/fdcc/?set=FoodSubstances&sort=Sortterm&order=ASC&startrow=1&type=basic&search=thymol (accessed on 8 September 2023).
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef]
- Hung, N.H.; Quan, P.M.; Satyal, P.; Dai, D.N.; Hoa, V.V.; Huy, N.G.; Giang, L.D.; Ha, N.T.; Huong, L.T.; Hien, V.T. Acetylcholinesterase Inhibitory Activities of Essential Oils from Vietnamese Traditional Medicinal Plants. Molecules 2022, 27, 7092. [Google Scholar] [CrossRef]
- Kurt, B.Z.; Gazioglu, I.; Dag, A.; Salmas, R.E.; Kayık, G.; Durdagi, S.; Sonmez, F. Synthesis, anticholinesterase activity and molecular modeling study of novel carbamate-substituted thymol/carvacrol derivatives. Bioorganic Med. Chem. 2017, 25, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Caputo, L.; Amato, G.; De Martino, L.; De Feo, V.; Nazzaro, F. Anti-Cholinesterase and Anti-α-Amylase Activities and Neuroprotective Effects of Carvacrol and p-Cymene and Their Effects on Hydrogen Peroxide Induced Stress in SH-SY5Y Cells. Int. J. Mol. Sci. 2023, 24, 6073. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.; Kartal, M.; Kan, Y.; Şener, B. Activity of essential oils and individual components against acetyland butyrylcholinesterase. Z. Fuer Naturforschung C 2008, 63, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Jukic, M.; Politeo, O.; Maksimovic, M.; Milos, M.; Milos, M. In vitro acetylcholinesterase inhibitory properties of thymol, carvacrol and their derivatives thymoquinone and thymohydroquinone. Phytother. Res. 2007, 21, 259–261. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.M.; Andreolla, M.C.; Ribeiro, T.C.; Gonçalves, A.E.; Medeiros, A.R.; de Souza, A.S.; Ferreira, L.L.G.; Andricopulo, A.D.; Yunes, R.A.; de Oliveira, A.S. Structure-activity relationships of sulfonamides derived from carvacrol and their potential for the treatment of Alzheimer’s disease. RSC Med. Chem. 2020, 11, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.G.; da Costa, R.A.; de Oliveira, M.S.; da Cruz, J.N.; Figueiredo, P.L.B.; Brasil, D.d.S.B.; Nascimento, L.D.; Chaves Neto, A.M.d.J.; de Carvalho Junior, R.N.; Andrade, E.H.d.A. Chemical profile of Lippia thymoides, evaluation of the acetylcholinesterase inhibitory activity of its essential oil, and molecular docking and molecular dynamics simulations. PLoS ONE 2019, 14, e0213393. [Google Scholar] [CrossRef] [PubMed]
- Tupe, R.S.; Sankhe, N.M.; Shaikh, S.A.; Kemse, N.G.; Khaire, A.A.; Phatak, D.V.; Parikh, J.U. Nutraceutical properties of dietary plants extracts: Prevention of diabetic nephropathy through inhibition of glycation and toxicity to erythrocytes and HEK293 cells. Pharm. Biol. 2015, 53, 40–50. [Google Scholar] [CrossRef]
- Ahmed, H. Cumin Seeds: The Functional Food with an Ability to Inhibit the Deleterious Phenomenon of Glycation. Zia. J. Pharm. 2021, 2, 62–66. [Google Scholar]
- Morimitsu, Y.; Yoshida, K.; Esaki, S.; Hirota, A. Protein glycation inhibitors from thyme (Thymus vulgaris). Biosci. Biotechnol. Biochem. 1995, 59, 2018–2021. [Google Scholar] [CrossRef]
- Abbasi, S.; Gharaghani, S.; Benvidi, A.; Rezaeinasab, M. New insights into the efficiency of thymol synergistic effect with p-cymene in inhibiting advanced glycation end products: A multi-way analysis based on spectroscopic and electrochemical methods in combination with molecular docking study. J. Pharm. Biomed. Anal. 2018, 150, 436–451. [Google Scholar] [CrossRef]
- Voropai, E.; Samtsov, M.; Kaplevskii, K.; Maskevich, A.; Stepuro, V.; Povarova, O.; Kuznetsova, I.; Turoverov, K.; Fink, A.; Uverskii, V. Spectral properties of thioflavin T and its complexes with amyloid fibrils. J. Appl. Spectrosc. 2003, 70, 868–874. [Google Scholar] [CrossRef]
- Gour, N.; Koshti, B.; Kshtriya, V.S. A Chemical Perspective to the Anti-Amyloid Action of Compounds and a Nanoparticle Based Assay for Screening Amyloid Inhibitors. ChemRxiv 2019. [Google Scholar]
- Sharoar, M.G.; Thapa, A.; Shahnawaz, M.; Ramasamy, V.S.; Woo, E.-R.; Shin, S.Y.; Park, I.-S. Keampferol-3-O-rhamnoside abrogates amyloid beta toxicity by modulating monomers and remodeling oligomers and fibrils to non-toxic aggregates. J. Biomed. Sci. 2012, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Gazit, E. Mechanisms of amyloid fibril self-assembly and inhibition: Model short peptides as a key research tool. FEBS J. 2005, 272, 5971–5978. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, Y.; Nishimura, S.; Murasugi, T.; Kaneko, I.; Meguro, M.; Marumoto, S.; Kogen, H.; Koyama, K.; Oda, T. A novel β-sheet breaker, RS-0406, reverses amyloid β-induced cytotoxicity and impairment of long-term potentiation in vitro. Br. J. Pharmacol. 2002, 137, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Amtul, Z.; Westaway, D.; Cechetto, D.F.; Rozmahel, R.F. Oleic acid ameliorates amyloidosis in cellular and mouse models of Alzheimer’s disease. Brain Pathol. 2011, 21, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.P.; Kumar, N.; Chauhan, B.S.; Garg, P. Carbamate as a potential anti-Alzheimer’s pharmacophore: A review. Drug Dev. Res. 2023, 84, 1624–1651. [Google Scholar] [CrossRef]
- Song, J.; Kim, Y.-S.; Lee, D.H.; Lee, S.H.; Park, H.J.; Lee, D.; Kim, H. Neuroprotective effects of oleic acid in rodent models of cerebral ischaemia. Sci. Rep. 2019, 9, 10732. [Google Scholar] [CrossRef]
- Im, D.; Kim, S.; Yoon, G.; Hyun, D.G.; Eom, Y.-G.; Lee, Y.E.; Sohn, C.H.; Choi, J.-M.; Kim, H.I. Decoding the Roles of Amyloid-β (1–42)’s Key Oligomerization Domains toward Designing Epitope-Specific Aggregation Inhibitors. JACS Au 2023, 3, 1065–1075. [Google Scholar] [CrossRef]
- Kobayashi, H.; Murata, M.; Kawanishi, S.; Oikawa, S. Polyphenols with Anti-Amyloid β Aggregation Show Potential Risk of Toxicity Via Pro-Oxidant Properties. Int. J. Mol. Sci. 2020, 21, 3561. [Google Scholar] [CrossRef]
- Necula, M.; Kayed, R.; Milton, S.; Glabe, C.G. Small Molecule Inhibitors of Aggregation Indicate That Amyloid β Oligomerization and Fibrillization Pathways Are Independent and Distinct. J. Biol. Chem. 2007, 282, 10311–10324. [Google Scholar] [CrossRef] [PubMed]
- Orlando, R.A.; Gonzales, A.M.; Royer, R.E.; Deck, L.M.; Vander Jagt, D.L. A chemical analog of curcumin as an improved inhibitor of amyloid Abeta oligomerization. PLoS ONE 2012, 7, e31869. [Google Scholar] [CrossRef] [PubMed]
- Azizi, Z.; Salimi, M.; Amanzadeh, A.; Majelssi, N.; Naghdi, N. Carvacrol and thymol attenuate cytotoxicity induced by amyloid β25-35 via activating protein kinase C and inhibiting oxidative stress in PC12 cells. Iran. Biomed. J. 2020, 24, 243. [Google Scholar] [CrossRef]
- Aydin, E.; Turkez, H.; Keles, M.S. The effect of carvacrol on healthy neurons and N2a cancer cells: Some biochemical, anticancerogenicity and genotoxicity studies. Cytotechnology 2014, 66, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.-w.; Xie, Z.-x.; Wang, B.-f.; Zhong, Z.-h.; Chen, X.-y.; Sun, Y.-h.; Sun, Q.-f.; Yang, G.-y.; Bian, L.-g. Carvacrol protects neuroblastoma SH-SY5Y cells against Fe2+-induced apoptosis by suppressing activation of MAPK/JNK-NF-κB signaling pathway. Acta Pharmacol. Sin. 2015, 36, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Wojtunik-Kulesza, K.A.; Targowska-Duda, K.; Klimek, K.; Ginalska, G.; Jóźwiak, K.; Waksmundzka-Hajnos, M.; Cieśla, Ł. Volatile terpenoids as potential drug leads in Alzheimer’s disease. Open Chem. 2017, 15, 332–343. [Google Scholar] [CrossRef]
- Smolecule. Vinyltriphenylphosphonium Bromide. 2023. Available online: https://www.smolecule.com/products/s714471 (accessed on 10 September 2023).
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Chen, H.Y. Why the Reactive Oxygen Species of the Fenton Reaction Switches from Oxoiron(IV) Species to Hydroxyl Radical in Phosphate Buffer Solutions? A Computational Rationale. ACS Omega 2019, 4, 14105–14113. [Google Scholar] [CrossRef]
- Pradeep, K.; Raj Mohan, C.V.; Gobianand, K.; Karthikeyan, S. Protective effect of Cassia fistula Linn. on diethylnitrosamine induced hepatocellular damage and oxidative stress in ethanol pretreated rats. Biol. Res. 2010, 43, 113–125. [Google Scholar] [CrossRef]
- Banik, S.; Rahman, M.M.; Sikder, M.T.; Saito, T.; Kurasaki, M. Protective effects of ajwain (Trachyspermum ammi L.) extract against cadmium-induced cytotoxicity and apoptosis in PC12 cells. J. Herb. Med. 2021, 26, 100423. [Google Scholar] [CrossRef]
- Wei, H.K.; Xue, H.X.; Zhou, Z.X.; Peng, J. A carvacrol–thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Chenet, A.L.; Duarte, A.R.; de Almeida, F.J.S.; Andrade, C.M.B.; de Oliveira, M.R. Carvacrol Depends on Heme Oxygenase-1 (HO-1) to Exert Antioxidant, Anti-inflammatory, and Mitochondria-Related Protection in the Human Neuroblastoma SH-SY5Y Cells Line Exposed to Hydrogen Peroxide. Neurochem. Res. 2019, 44, 884–896. [Google Scholar] [CrossRef]
- Banik, S.; Akter, M.; Corpus Bondad, S.E.; Saito, T.; Hosokawa, T.; Kurasaki, M. Carvacrol inhibits cadmium toxicity through combating against caspase dependent/independent apoptosis in PC12 cells. Food Chem. Toxicol. 2019, 134, 110835. [Google Scholar] [CrossRef]
- Tiefensee Ribeiro, C.; Gasparotto, J.; Petiz, L.L.; Brum, P.O.; Peixoto, D.O.; Kunzler, A.; da Rosa Silva, H.T.; Bortolin, R.C.; Almeida, R.F.; Quintans-Junior, L.J.; et al. Oral administration of carvacrol/β-cyclodextrin complex protects against 6-hydroxydopamine-induced dopaminergic denervation. Neurochem. Int. 2019, 126, 27–35. [Google Scholar] [CrossRef]
- Meeran, M.N.; Jagadeesh, G.; Selvaraj, P. Thymol, a dietary monoterpene phenol abrogates mitochondrial dysfunction in β-adrenergic agonist induced myocardial infarcted rats by inhibiting oxidative stress. Chem.-Biol. Interact. 2016, 244, 159–168. [Google Scholar] [CrossRef]
- Shettigar, N.B.; Das, S.; Rao, N.B.; Rao, S.B. Thymol, a monoterpene phenolic derivative of cymene, abrogates mercury-induced oxidative stress resultant cytotoxicity and genotoxicity in hepatocarcinoma cells. Environ. Toxicol. 2015, 30, 968–980. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).