
Sleep Deprivation-Induced Oxidative Stress in Rat Models: A Scoping Systematic Review

 ,
,  ,
,  and
and 
Abstract
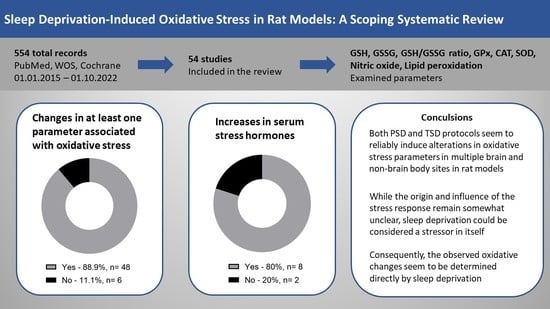
1. Introduction
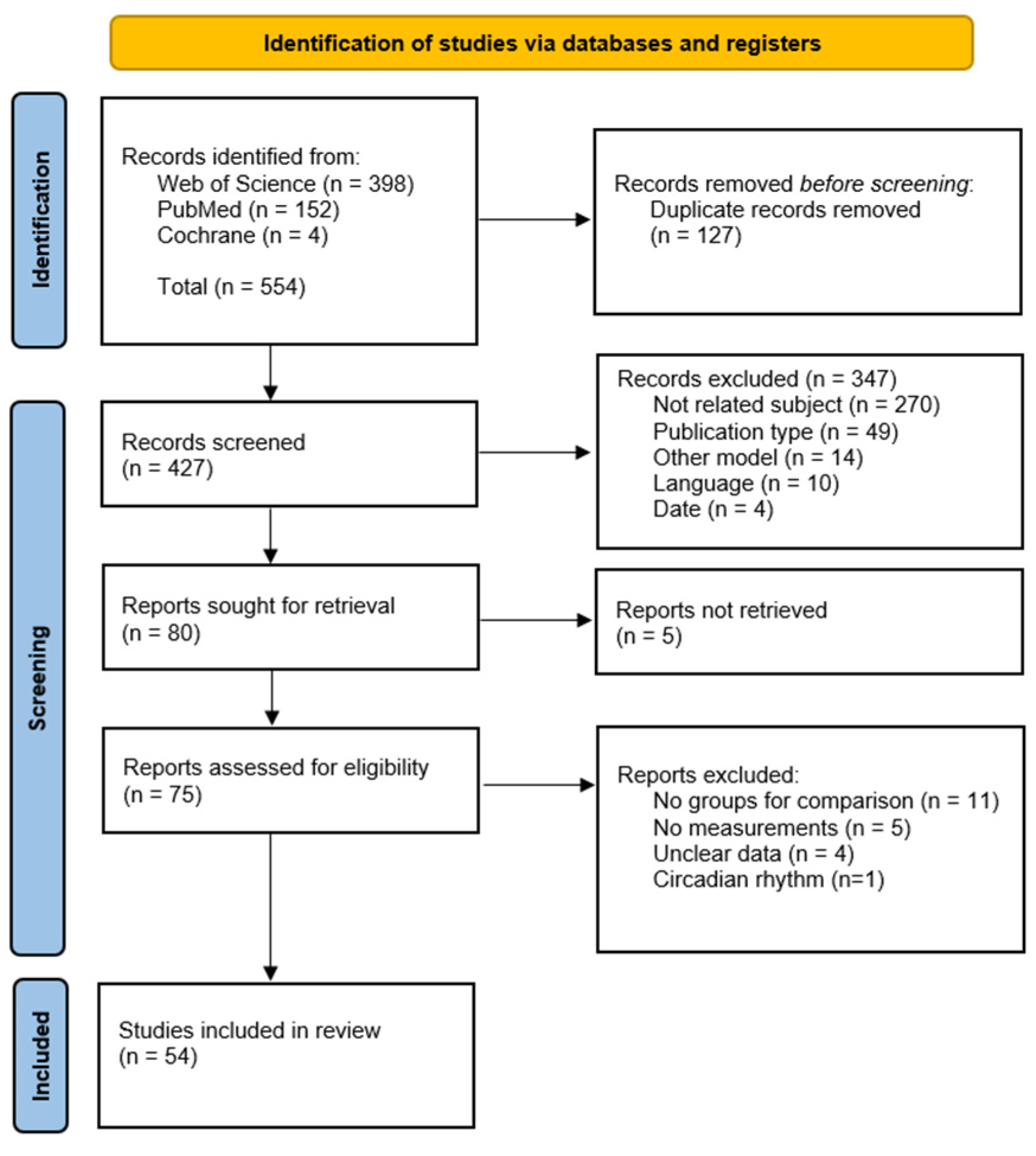
2. Materials and Methods
3. Results

{kind=link}
{kind=link}
| Reference | SD Protocol | SD Duration | Rat Breed, Sex, Age | Oxidative Stress Marker | Results |
|---|---|---|---|---|---|
| [27] | MSP | 48 h | Wistar, Male, PND 28 | LP | LP (-) |
| [44] | MSP | 48 h | Wistar, Male, Adult | GSH, NOx, LP | GSH (↓), NOx (↑), LP (-) |
| [26] | MSP | 72 h | Wistar, Male, Adult | GSH, LP | GSH (-), LP (-) |
| [45] | MSP | 72 h | Wistar, Male | LP | LP (↑) |
| [46] | MSP | 72 h | Wistar, Male, Adult | GSH, SOD, LP | GSH (↓), SOD (↓), LP (↑) |
| [47] | MSP | 72 h | Wistar, Male, Adult | LP | LP (↑) |
| [31] | MSP | 24, 48, 72 h | Wistar, Male, Adult | GSH, GPx, CAT, SOD, LP | GSH (-), GPx (-), CAT (-), SOD (-), LP (-) |
| [48] | CP | 4 days | Wistar, Male, 5 weeks | LP | LP (↑) * |
| [49] | MSP | 5 days | Sprague Dawley, Male and Female, 6 weeks | GSH/GSSG ratio, GPX, CAT, SOD, LP | GSH/GSSG ratio (↓), GPX (-), CAT (↓), SOD (↓), LP (↑) |
| [38] | MSP | 5 days | Sprague Dawley, Male, 48 weeks | NOx | NOx (↑) |
| [50] | CP | 6 days (48 h SD, 48 h Srec, 48 h SD, 48 h Srec, 48 h SD) | Wistar, Male, Adult | CAT, LP | CAT (-), LP (↑) |
| [43] | MSP | 21 days (18 h/day) 21 days (18 h/day) + 5 days Srec 21 days (18 h/day) + 21 days Srec | Sprague Dawley, Male, 12–16 weeks | GSH/GSSG ratio, LP | GSH/GSSG ratio (↓), LP (↑) GSH/GSSG ratio (↓), LP (↑) GSH/GSSG ratio (↓), LP (-) |
| [51] | MSP | 4 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, GPx, CAT | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓) |
| [52] | MSP | 4 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, GPx, CAT | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓) |
| [53] | MSP | 4 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, GPx, CAT, SOD, LP | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓), SOD (↓), LP (-) |
| [54] | MSP | 6 weeks (8 h/day) | Wistar, Male, 8–10 weeks | GSH, GSSG, GSH/GSSG ratio, GPx, CAT, SOD, LP | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓), SOD (↓), LP (-) |
| [55] | MSP | 6 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, GPx, CAT, SOD, LP | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓), SOD (↓), LP (-) |
| [56] | MSP | 6 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, GPx, CAT, SOD | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓), SOD (↓) |
| [57] | MSP | 6 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, CAT, SOD, LP | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), CAT (↓), SOD (↓), LP (-) |
| [58] | MSP | 8 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, GPx, CAT, SOD, LP | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓), SOD (-), LP (-) |
| [59] | MSP | 8 weeks (8 h/day) | Wistar, Male, Adult | GSH, GSSG, GSH/GSSG ratio, GPx, CAT | GSH (-), GSSG (↑), GSH/GSSG ratio (↓), GPx (↓), CAT (↓) |
| Reference | SD Protocol | SD Duration | Rat Breed, Sex, Age | Anatomical Site | Oxidative Stress Marker | Results |
|---|---|---|---|---|---|---|
| [28] | MSP | 48 h | Wistar, Male, Adult | Cortex | GSH, NOx, LP | GSH (-), NOx (-), LP (-) |
| [26] | MSP | 72 h | Wistar, Male, Adult | Prefrontal cortex | GSH, LP | GSH (-), LP (-) |
| [45] | MSP | 72 h | Wistar, Male | Forebrain cortex | LP | LP (↑) |
| [47] | MSP | 72 h | Wistar, Male | Forebrain cortex | LP | LP (↑) |
| [48] | CP | 4 days | Wistar, Male, 5 weeks | Cortex | LP | LP (↑) |
| [60] | MSP | 7 days | Wistar, Male, 8 weeks | Cortex | GSH, GPx, CAT, SOD, LP | GSH (↓), GPx (↓), CAT (↓), SOD (↓), LP (↑) |
| [26] | MSP | 72 h | Wistar, Male, Adult | Cerebellum, Brainstem | GSH, LP | GSH (-), LP (-) |
| [36] | MSP | 72 h | Sprague Dawley, Adult | Amygdala | SOD, LP | SOD (↓), LP (↑) |
| [61] | CP | 72 h | Sprague Dawley, Male, 8–10 weeks | Thalamus | GSH, CAT, SOD, LP | GSH (↓), CAT (↓), SOD (↓), LP (↑) |
| [62] | MSP | 5 days | Sprague Dawley, Male and Female, Adult | Whole brain | CAT, SOD, LP | CAT (↓), SOD (↓), LP (↑) |
| [63] | CP | 6 days | Wistar, Male, Adult | Locus coeruleus | GSH | GSH (↓) |
| Reference | SD Protocol | SD Duration | Rat Breed, Sex, Age | Oxidative Stress Markers | Results |
|---|---|---|---|---|---|
| [39] | MSP | 20 h | ns | LP | LP (↑) |
| [35] | MSP | 24 h | Long-Evans, Male, Old | LP | LP (↑) |
| [64] | CP | 24, 36, 48 h | Sprague Dawley, Male, 6 months | SOD, LP | SOD (↓), LP (↑) |
| [65] | CP | 48 h | Sprague Dawley, Male, 6 weeks | SOD, LP | SOD (↓), LP (↑) |
| [17] | MSP | 72 h | Wistar, Male, 10 weeks | NOx | NOx (↓) |
| [36] | MSP | 72 h | Sprague Dawley, Adult | SOD, LP | SOD (↓), LP (↑) |
| [40] | MSP | 5 days | Wistar, Male | LP | LP (↑) |
| [39] | MSP | 5 days (20 h/day) 5 days (20 h/day) + 5 days Srec | ns | LP | LP (↑) LP (-) |
| [50] | CP | 6 days (48 h SD, 48 h Srec, 48 h SD, 48 h Srec, 48 h SD) | Wistar, Male, Adult | CAT, LP | CAT (-), LP (-) |
| [66] | MSP | 7 days (20 h/day) and 7 days continuous | Wistar, Male | CAT, SOD, LP | CAT (-/↓), SOD (-), LP (↑) |
| [67] | MSP | 7 days | Wistar, Male | GPx, SOD | GPx (↓), SOD (↓) |
| [30] | MSP | 21 days (18 h/day) | Wistar, Male, Adult | eNOS | eNOS (-) |
| Reference | SD Protocol | SD Duration | Rat Breed, Sex, Age | Anatomical Site | Oxidative Stress Markers | Results |
|---|---|---|---|---|---|---|
| [17] | MSP | 72 h | Wistar, Male, 10 weeks | Testes, Epididymis | GSH, GPx, CAT, SOD, LP | GSH (↓), GPx (↓), CAT (↓), SOD (↓), LP (↑) |
| [40] | MSP | 5 days | Wistar, Male | Testes | GSH, GPx, LP | GSH (↓), GPx (↓), LP (↑) |
| [34] | MSP | 14 days (20 h/day) | Sprague Dawley, Male, 12 weeks | Testes | GSH, CAT, SOD, LP | GSH (↓), CAT (↓), SOD (↑), LP (↑) |
| [41] | MSP | 21 days (18 h/day) | Wistar, Male, Peripubertal | Testes | GSH, GSSG, GSH/GSSG ratio, LP | GSH (-), GSSG (-), GSH/GSSG ratio (-), LP (↑) |
| [32] | MSP | 21 days (18 h/day) | Wistar, Male, Peripubertal | Epididymis caput, cauda | GSH, LP | GSH (↑), LP (↑) |
| [33] | MSP | 4 days 8 days 8 days + 20 days Srec | Wistar, Male, 3 months | Liver, Pancreas | CAT, SOD, LP | CAT (-), SOD (-/↑), LP (-/↑) CAT (↓/-), SOD (↓/-), LP (↑/-) CAT (-), SOD (↓/-), LP (↑) |
| Wistar, Male, 14 months | Liver, Pancreas | CAT, SOD, LP | CAT (↓/-), SOD (-), LP (-) CAT (↓/-), SOD (-/↑), LP (-/↑) CAT (-), SOD (-/↑), LP (-) | |||
| [68] | MSP | 21 days (14 h/day) | Wistar, Male, Adult | Liver | SOD, LP | SOD (↓), LP (↑) |
| [69] | CP | 21 days (18 h/day) 21 days (22 h/day) | Wistar, Male, Adult | Liver | GPx, SOD, LP | GPx (-), SOD (-), LP (-) GPx (-), SOD (↓), LP (↑) |
| [70] | CP | 72 h 72 h + 72 h Srec | Sprague Dawley, Male, 8–10 weeks | Aorta | SOD, LP | SOD (↓), LP (↑) SOD (-), LP (-) |
| [71] | CP | 5 days | Sprague Dawley, Male, 24 weeks | Aorta | NOx, eNOS, p-eNOS | NOx (↓), eNOS (-), p-eNOS (↓) |
| [66] | MSP | 7 days (20 h/day) and 7 days continuous | Wistar, Male | Saliva Submandibular | CAT, SOD, LP | CAT (-), SOD (-), LP (-) CAT (-), SOD (↓), LP (-/↑) |
| [72] | CP | 21 days (18 h/day) | Wistar, Male | Thyroid | LP | LP (↑) |
| [45] | MSP | 72 h | Wistar, Male | Kidney Erythrocytes | LP | LP (-) LP (↑) |
| [37] | MSP | 4 days | Wistar, Male, 3 months | Plantar muscle Soleus muscle | LP | LP (-) LP (↑) |
| Reference | SD Protocol | SD Duration | Rat Breed, Sex, Age | Anatomical Site | Oxidative Stress Markers | Results |
|---|---|---|---|---|---|---|
| [29] | GH | 6 h | Wistar, Male, 10 weeks | Hippocampus | LP | LP (-) |
| [42] | GH | 12 h | Wistar, Female, 13–15 months | Hypothalamus | nNOS | nNOS (↓) |
| [73] | ASD | 14 days | Wistar, Male | Cortex and Hippocampus | GSH, CAT, SOD, LP | GSH (↓), CAT (↓), SOD (↓), LP (↑) |
| [74] | DOW | 5 days SD + 2 days Srec (3 total cycles) + 3 months Srec | Wistar, Male, Weanling | Hippocampus | GPx, CAT, SOD | GPx (↓), CAT (↓), SOD (↓/-) |
| Reference | SD Protocol | SD Duration | Rat Breed, Sex, Age | Anatomical Site | Oxidative Stress Markers | Results |
|---|---|---|---|---|---|---|
| [75] | DOW | 5 days | Wistar, Male | Liver | GPx, CAT, SOD, LP | GPx (↓), CAT (↓), SOD (↓), LP (↑) |
| [76] | DOW | 5 days SD + 2 days Srec (3 total cycles) | Wistar, Male, Adult | Liver | GPx, CAT, SOD, LP | GPx (↓), CAT (↓), SOD (↓), LP (↑) |
| [77] | ASD | 14 days 6 h/day–1st week 8 h/day–2nd week | Sprague Dawley, Male, PND 19 | Plasma | LP | PND 33: LP (↑) PND 90: LP (-) |
| Reference | SD Protocol | SD Duration | Rat Breed, Sex, Age | Anatomical Site | Oxidative Stress Measurements | Results |
|---|---|---|---|---|---|---|
| [78] | PSD–MSP | 96 h 21 days (18 h/day) | Wistar-Hannover, Male, Adult | Testes | iNOS, eNOS | iNOS (↑), eNOS (↓) iNOS (↑), eNOS (-) |
| [77] | TSD–ASD | 14 days 6 h/day–1st week 8 h/day–2nd week | Sprague Dawley, Male, PND 19 | Prefrontal cortex | GPx, CAT, SOD | PND 33: GPx (↑), CAT (-), SOD (↑) PND 90: GPx (-), CAT (-), SOD (-) |
| Reference | SD Type | SD Protocol | SD Duration | Rat Breed, Sex, Age | Cortisol/Corticosterone |
|---|---|---|---|---|---|
| [42] | TSD | GH | 12 h | Wistar, Female, 13–15 months | Yes - |
| [35] | PSD | MSP | 24 h | Long-Evans, Male, Old | Yes ↑ |
| [36] | PSD | MSP | 72 h | Sprague Dawley, Adult | Yes ↑ |
| [37] | PSD | MSP | 4 days | Wistar, Male, 3 months | Yes ↑ |
| [38] | PSD | MSP | 5 days | Sprague Dawley, Male, 48 weeks | Yes ↑ |
| [39] | PSD | MSP | 20 h 5 days (20 h/day) 5 days (20 h/day) + 5 days Srec | ns | Yes ↑ Yes ↑ Yes - |
| [40] | PSD | MSP | 5 days | Wistar, Male | Yes ↑ |
| [34] | PSD | MSP | 14 days (20 h/day) | Sprague Dawley, Male, 12 weeks | Yes ↑ |
| [41] | PSD | MSP | 21 days (18 h/day) | Wistar, Male, Peripubertal | Yes ↑ |
| [43] | PSD | MSP | 21 days (18 h/day) 21 days (18 h/day) + 5 days Srec 21 days (18 h/day) + 21 days Srec | Sprague Dawley, Male, 12–16 weeks | Yes - |
4. Discussion
4.1. Sleep Deprivation Determines Changes in Oxidative Stress Parameters
4.2. Sleep Recovery
4.3. Influence of Sex in Response to Sleep Deprivation
4.4. Sleep Deprivation and the Stress Response
4.5. Mechanisms of Sleep Deprivation-Induced Oxidative Stress
4.6. Negative Health Outcomes Associated with Sleep Deprivation
5. Limitations
5.1. Limitations Inherent to the Review Methodology and Results
5.2. Limitations Inherent to the Original Studies
5.3. Limitations Determined by the Existing Sleep Deprivation Protocols
6. Quality and Bias Evaluation of Included Studies
7. Conclusions
8. Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kocevska, D.; Lysen, T.S.; Dotinga, A.; Koopman-Verhoeff, M.E.; Luijk, M.P.C.M.; Antypa, N.; Biermasz, N.R.; Blokstra, A.; Brug, J.; Burk, W.J.; et al. Sleep Characteristics across the Lifespan in 1.1 Million People from the Netherlands, United Kingdom and United States: A Systematic Review and Meta-Analysis. Nat. Hum. Behav. 2021, 5, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Neculicioiu, V.S.; Colosi, I.A.; Costache, C.; Sevastre-Berghian, A.; Clichici, S. Time to Sleep?—A Review of the Impact of the COVID-19 Pandemic on Sleep and Mental Health. Int. J. Environ. Res. Public Health 2022, 19, 3497. [Google Scholar] [CrossRef]
- Colten, H.R.; Altevogt, B.M. Institute of Medicine (US) Committee on Sleep Medicine and Research. In Extent and Health Consequences of Chronic Sleep Loss and Sleep Disorders; National Academies Press (US): Washington, DC, USA, 2006. [Google Scholar]
- Garbarino, S.; Lanteri, P.; Bragazzi, N.L.; Magnavita, N.; Scoditti, E. Role of Sleep Deprivation in Immune-Related Disease Risk and Outcomes. Commun. Biol. 2021, 4, 1304. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.L. A Brief Report on Early Sleep Studies. Sleep Sci. 2020, 13, 1–2. [Google Scholar] [PubMed]
- Brinkman, J.E.; Reddy, V.; Sharma, S. Physiology of Sleep. In StatPearls [Internet]; StatPearls Publishing: St. Petersburg, FL, USA, 2022. [Google Scholar]
- Vyazovskiy, V.V. Sleep, Recovery, and Metaregulation: Explaining the Benefits of Sleep. Nat. Sci. Sleep 2015, 7, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Reimund, E. The Free Radical Flux Theory of Sleep. Med. Hypotheses 1994, 43, 231–233. [Google Scholar] [CrossRef]
- Villafuerte, G.; Miguel-Puga, A.; Rodríguez, E.M.; Machado, S.; Manjarrez, E.; Arias-Carrión, O. Sleep Deprivation and Oxidative Stress in Animal Models: A Systematic Review. Oxid. Med. Cell. Longev. 2015, 2015, 234952. [Google Scholar] [CrossRef]
- Hill, V.M.; O’Connor, R.M.; Sissoko, G.B.; Irobunda, I.S.; Leong, S.; Canman, J.C.; Stavropoulos, N.; Shirasu-Hiza, M. A Bidirectional Relationship between Sleep and Oxidative Stress in Drosophila. PLoS Biol. 2018, 16, e2005206. [Google Scholar] [CrossRef]
- Atrooz, F.; Salim, S. Sleep Deprivation, Oxidative Stress and Inflammation. Adv. Protein Chem. Struct. Biol. 2020, 119, 309–336. [Google Scholar]
- Mobbs, C. Oxidative Stress and Acidosis, Molecular Responses To. In Encyclopedia of Stress; Fink, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 3, p. 45. [Google Scholar]
- Sies, H. Chapter 13—Oxidative Stress: Eustress and Distress in Redox Homeostasis. In Stress: Physiology, Biochemistry, and Pathology; Fink, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 153–163. [Google Scholar]
- Katerji, M.; Filippova, M.; Duerksen-Hughes, P. Approaches and Methods to Measure Oxidative Stress in Clinical Samples: Research Applications in the Cancer Field. Oxid. Med. Cell. Longev. 2019, 2019, 1279250. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.A.; Akhigbe, T.M.; Akhigbe, R.E.; Aremu, A.O.; Oyedokun, P.A.; Gbadamosi, J.A.; Anifowose, P.E.; Adewole, M.A.; Aboyeji, O.O.; Yisau, H.O.; et al. Glutamine Restores Testicular Glutathione-Dependent Antioxidant Defense and Upregulates NO/CGMP Signaling in Sleep Deprivation-Induced Reproductive Dysfunction in Rats. Biomed. Pharmacother. 2022, 148, 112765. [Google Scholar] [CrossRef] [PubMed]
- Goshi, E.; Zhou, G.; He, Q. Nitric Oxide Detection Methods in Vitro and in Vivo. Med. Gas Res. 2019, 9, 192–207. [Google Scholar] [PubMed]
- Pierini, D.; Bryan, N. Nitric Oxide Availability as a Marker of Oxidative Stress. In Advanced Protocols in Oxidative Stress III; Armstrong, D., Ed.; Springer: New York, NY, USA, 2015; Volume 3. [Google Scholar]
- Wang, F.; Yuan, Q.; Chen, F.; Pang, J.; Pan, C.; Xu, F.; Chen, Y. Fundamental Mechanisms of the Cell Death Caused by Nitrosative Stress. Front. Cell Dev. Biol. 2021, 9, 742483. [Google Scholar] [CrossRef]
- Cialoni, D.; Brizzolari, A.; Samaja, M.; Bosco, G.; Paganini, M.; Pieri, M.; Lancellotti, V.; Marroni, A. Nitric Oxide and Oxidative Stress Changes at Depth in Breath-Hold Diving. Front. Physiol. 2020, 11, 609642. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Colavito, V.; Fabene, P.F.; Grassi-Zucconi, G.; Pifferi, F.; Lamberty, Y.; Bentivoglio, M.; Bertini, G. Experimental Sleep Deprivation as a Tool to Test Memory Deficits in Rodents. Front. Syst. Neurosci. 2013, 7, 106. [Google Scholar] [CrossRef]
- Wright, C.J.; Milosavljevic, S.; Pocivavsek, A. The Stress of Losing Sleep: Sex-Specific Neurobiological Outcomes. Neurobiol. Stress 2023, 24, 100543. [Google Scholar] [CrossRef]
- Turan, I.; Sayan Ozacmak, H.; Ozacmak, V.H.; Ergenc, M.; Bayraktaroğlu, T. The Effects of Glucagon-like Peptide 1 Receptor Agonist (Exenatide) on Memory Impairment, and Anxiety- and Depression-like Behavior Induced by REM Sleep Deprivation. Brain Res. Bull. 2021, 174, 194–202. [Google Scholar] [CrossRef]
- Sahin, L.; Cevik, O.S.; Cevik, K.; Guven, C.; Taskin, E.; Kocahan, S. Mild Regular Treadmill Exercise Ameliorated the Detrimental Effects of Acute Sleep Deprivation on Spatial Memory. Brain Res. 2021, 1759, 147367. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.S.; Khadrawy, Y.A. Electrophysiological and Neurochemical Evaluation of the Adverse Effects of REM Sleep Deprivation and Epileptic Seizures on Rat’s Brain. Life Sci. 2021, 273, 119303. [Google Scholar] [CrossRef]
- Forouzanfar, F.; Gholami, J.; Foroughnia, M.; Payvar, B.; Nemati, S.; Khodadadegan, M.A.; Saheb, M.; Hajali, V. The Beneficial Effects of Green Tea on Sleep Deprivation-Induced Cognitive Deficits in Rats: The Involvement of Hippocampal Antioxidant Defense. Heliyon 2021, 7, e08336. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, G.; Ozcan, K.M.; Keskin, O.; Tekeli, F.; Kaymaz, A.A. Effects of Chronic REM Sleep Deprivation on Lipocalin-2, Nitric Oxide Synthase-3, Interleukin-6 and Cardiotrophin-1 Levels: An Experimental Rat Model. Sleep Biol. Rhythm. 2019, 17, 305–310. [Google Scholar] [CrossRef]
- Nabaee, E.; Kesmati, M.; Shahriari, A.; Khajehpour, L.; Torabi, M. Cognitive and Hippocampus Biochemical Changes Following Sleep Deprivation in the Adult Male Rat. Biomed. Pharmacother. 2018, 104, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Siervo, G.E.M.L.; Ogo, F.M.; Valério, A.D.; Silva, T.N.X.; Staurengo-Ferrari, L.; Alvarenga, T.A.; Cecchini, R.; Verri, W.A.; Guarnier, F.A.; Andersen, M.L.; et al. Sleep Restriction in Wistar Rats Impairs Epididymal Postnatal Development and Sperm Motility in Association with Oxidative Stress. Reprod. Fertil. Dev. 2017, 29, 1813–1820. [Google Scholar] [CrossRef]
- Hernández Santiago, K.; López-López, A.L.; Sánchez-Muñoz, F.; Cortés Altamirano, J.L.; Alfaro-Rodríguez, A.; Bonilla-Jaime, H. Sleep Deprivation Induces Oxidative Stress in the Liver and Pancreas in Young and Aging Rats. Heliyon 2021, 7, e06466. [Google Scholar] [CrossRef]
- Medubi, L.J.; Nwosu, N.C.; Medubi, O.O.; Lawal, O.R.; Ama, C.; Kusemiju, T.O.; Osinubi, A.A. Increased de Novo Glutathione Production Enhances Sexual Dysfunctions in Rats Subjected to Paradoxical Sleep Deprivation. JBRA Assist. Reprod. 2021, 25, 215–222. [Google Scholar] [CrossRef]
- Solanki, N.; Atrooz, F.; Asghar, S.; Salim, S. Tempol Protects Sleep-Deprivation Induced Behavioral Deficits in Aggressive Male Long-Evans Rats. Neurosci. Lett. 2016, 612, 245–250. [Google Scholar] [CrossRef][Green Version]
- Zhang, L.; Guo, H.-L.; Zhang, H.-Q.; Xu, T.-Q.; He, B.; Wang, Z.-H.; Yang, Y.-P.; Tang, X.-D.; Zhang, P.; Liu, F.-E. Melatonin Prevents Sleep Deprivation-Associated Anxiety-like Behavior in Rats: Role of Oxidative Stress and Balance between GABAergic and Glutamatergic Transmission. Am. J. Transl. Res. 2017, 9, 2231–2242. [Google Scholar] [PubMed]
- Mônico-Neto, M.; Lee, K.S.; da Luz, M.H.M.; Pino, J.M.V.; Ribeiro, D.A.; Cardoso, C.M.; Sueur-Maluf, L.L.; Tufik, S.; Antunes, H.K.M. Histopathological Changes and Oxidative Damage in Type I and Type II Muscle Fibers in Rats Undergoing Paradoxical Sleep Deprivation. Cell. Signal. 2021, 81, 109939. [Google Scholar] [CrossRef] [PubMed]
- Gou, X.-J.; Cen, F.; Fan, Z.-Q.; Xu, Y.; Shen, H.-Y.; Zhou, M.-M. Serum and Brain Metabolomic Variations Reveal Perturbation of Sleep Deprivation on Rats and Ameliorate Effect of Total Ginsenoside Treatment. Int. J. Genom. Proteom. 2017, 2017, 5179271. [Google Scholar] [CrossRef] [PubMed]
- Olayaki, L.A.; Sulaiman, S.O.; Anoba, N.B. Vitamin C Prevents Sleep Deprivation-Induced Elevation in Cortisol and Lipid Peroxidation in the Rat Plasma. Niger. J. Physiol. Sci. 2015, 30, 5–9. [Google Scholar] [PubMed]
- Rizk, N.I.; Rizk, M.S.; Mohamed, A.S.; Naguib, Y.M. Attenuation of Sleep Deprivation Dependent Deterioration in Male Fertility Parameters by Vitamin C. Reprod. Biol. Endocrinol. 2020, 18, 2. [Google Scholar] [CrossRef] [PubMed]
- Siervo, G.E.M.L.; Ogo, F.M.; Staurengo-Ferrari, L.; Anselmo-Franci, J.A.; Cunha, F.Q.; Cecchini, R.; Guarnier, F.A.; Verri, W.A., Jr.; Fernandes, G.S.A. Sleep Restriction during Peripuberty Unbalances Sexual Hormones and Testicular Cytokines in Rats. Biol. Reprod. 2019, 100, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Kaur, G. Acute Sleep Deprivation-Induced Anxiety and Disruption of Hypothalamic Cell Survival and Plasticity: A Mechanistic Study of Protection by Butanol Extract of Tinospora Cordifolia. Neurochem. Res. 2022, 47, 1692–1706. [Google Scholar] [CrossRef]
- Konakanchi, S.; Raavi, V.; Ml, H.K.; Shankar Ms, V. Effect of Chronic Sleep Deprivation and Sleep Recovery on Hippocampal CA3 Neurons, Spatial Memory and Anxiety-like Behavior in Rats. Neurobiol. Learn. Mem. 2022, 187, 107559. [Google Scholar] [CrossRef]
- Aboul Ezz, H.S.; Noor, A.E.; Mourad, I.M.; Fahmy, H.; Khadrawy, Y.A. Neurochemical Effects of Sleep Deprivation in the Hippocampus of the Pilocarpine-Induced Rat Model of Epilepsy. Iran. J. Basic Med. Sci. 2021, 24, 85–91. [Google Scholar]
- Roubalová, L.; Vošahlíková, M.; Slaninová, J.; Kaufman, J.; Alda, M.; Svoboda, P. Tissue-Specific Protective Properties of Lithium: Comparison of Rat Kidney, Erythrocytes and Brain. Naunyn. Schmiedebergs. Arch. Pharmacol. 2021, 394, 955–965. [Google Scholar] [CrossRef]
- Zuo, J.-X.; Li, M.; Jiang, L.; Lan, F.; Tang, Y.-Y.; Kang, X.; Zou, W.; Wang, C.-Y.; Zhang, P.; Tang, X.-Q. Hydrogen Sulfide Prevents Sleep Deprivation-Induced Hippocampal Damage by Upregulation of Sirt1 in the Hippocampus. Front. Neurosci. 2020, 14, 169. [Google Scholar] [CrossRef]
- Vosahlikova, M.; Roubalova, L.; Cechova, K.; Kaufman, J.; Musil, S.; Miksik, I.; Alda, M.; Svoboda, P. Na+/K+-ATPase and Lipid Peroxidation in Forebrain Cortex and Hippocampus of Sleep-Deprived Rats Treated with Therapeutic Lithium Concentration for Different Periods of Time. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 102, 109953. [Google Scholar] [CrossRef]
- Kwon, K.J.; Lee, E.J.; Kim, M.K.; Jeon, S.J.; Choi, Y.Y.; Shin, C.Y.; Han, S.-H. The Potential Role of Melatonin on Sleep Deprivation-Induced Cognitive Impairments: Implication of FMRP on Cognitive Function. Neuroscience 2015, 301, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Su, G.; Zhao, T.; Wang, S.; Sun, B.; Zheng, L.; Zhao, M. The Memory Improving Effects of Round Scad (Decapterus Maruadsi) Hydrolysates on Sleep Deprivation-Induced Memory Deficits in Rats via Antioxidant and Neurotrophic Pathways. Food Funct. 2019, 10, 7733–7744. [Google Scholar] [CrossRef] [PubMed]
- Andrabi, M.; Andrabi, M.M.; Kunjunni, R.; Sriwastva, M.K.; Bose, S.; Sagar, R.; Srivastava, A.K.; Mathur, R.; Jain, S.; Subbiah, V. Lithium Acts to Modulate Abnormalities at Behavioral, Cellular, and Molecular Levels in Sleep Deprivation-Induced Mania-like Behavior. Bipolar Disord. 2020, 22, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Al-Jamal, F.F.; Mahasneh, A.F. Cerebrolysin Prevents Sleep Deprivation Induced Memory Impairment and Oxidative Stress. Physiol. Behav. 2020, 217, 112823. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Al Mosabih, H.S.; Mahasneh, A.F. The Protective Effect of Edaravone on Memory Impairment Induced by Chronic Sleep Deprivation. Psychiatry Res. 2019, 281, 112577. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Mayyas, F.A.; Khabour, O.F.; Bani Salama, F.M.; Alhashimi, F.H.; Mhaidat, N.M. Chronic Melatonin Treatment Prevents Memory Impairment Induced by Chronic Sleep Deprivation. Mol. Neurobiol. 2016, 53, 3439–3447. [Google Scholar] [CrossRef]
- Massadeh, A.M.; Alzoubi, K.H.; Milhem, A.M.; Rababa’h, A.M.; Khabour, O.F. Evaluating the Effect of Selenium on Spatial Memory Impairment Induced by Sleep Deprivation. Physiol. Behav. 2022, 244, 113669. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Rababa’h, A.M.; Owaisi, A.; Khabour, O.F. L-Carnitine Prevents Memory Impairment Induced by Chronic REM-Sleep Deprivation. Brain Res. Bull. 2017, 131, 176–182. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Khabour, O.F.; Albawaana, A.S.; Alhashimi, F.H.; Athamneh, R.Y. Tempol Prevents Chronic Sleep-Deprivation Induced Memory Impairment. Brain Res. Bull. 2016, 120, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Mhaidat, N.M.; Alzoubi, K.H.; Khabour, O.F.; Tashtoush, N.H.; Banihani, S.A.; Abdul-razzak, K.K. Exploring the Effect of Vitamin C on Sleep Deprivation Induced Memory Impairment. Brain Res. Bull. 2015, 113, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Mayyas, F.; Abu Zamzam, H.I. Omega-3 Fatty Acids Protects against Chronic Sleep-Deprivation Induced Memory Impairment. Life Sci. 2019, 227, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Malkawi, B.S.; Khabour, O.F.; El-Elimat, T.; Alali, F.Q. Arbutus andrachne L. Reverses Sleep Deprivation-Induced Memory Impairments in Rats. Mol. Neurobiol. 2018, 55, 1150–1156. [Google Scholar] [CrossRef]
- Suganya, K.; Kayalvizhi, E.; Yuvaraj, R.; Chandrasekar, M.; Kavitha, U.; Konakanchi Suresh, K. Effect of Withania Somnifera on the Antioxidant and Neurotransmitter Status in Sleep Deprivation Induced Wistar Rats. Bioinformation 2020, 16, 631–637. [Google Scholar] [CrossRef]
- Anis Syahirah, M.S.; Che Badariah, A.A.; Idris, L.; Rosfaiizah, S.; Liza, N. Impact of Rapid Eye Movement Sleep Deprivation on Pain Behaviour and Oxidative Stress in the Thalamus: Role of Tualang Honey Supplementation. Malays. J. Med. Sci. 2022, 29, 69–79. [Google Scholar]
- Ahmad, L.; Mujahid, M.; Mishra, A.; Rahman, M.A. Protective Role of Hydroalcoholic Extract of Cajanus Cajan Linn Leaves against Memory Impairment in Sleep Deprived Experimental Rats. J. Ayurveda Integr. Med. 2020, 11, 471–477. [Google Scholar] [CrossRef]
- Jameie, S.B.; Mesgar, S.; Aliaghaei, A.; Raoofi, A.; Amini, M.; Khodagholi, F.; Danyali, S.; Sorraya, M.; Sadeghi, Y. Neuroprotective Effect of Exogenous Melatonin on the Noradrenergic Neurons of Adult Male Rats’ Locus Coeruleus Nucleus Following REM Sleep Deprivation. J. Chem. Neuroanat. 2019, 100, 101656. [Google Scholar] [CrossRef]
- Hao, L.; Yang, R.; Xu, H. Effects of Exercise of Equal Intensity on Working Memory and BDNF Protein Expression in the Prefrontal Cortex in Rats with Different Degrees of Sleep Deprivation. Int. J. Clin. Exp. Med. 2019, 12, 11490–11496. [Google Scholar]
- Zhang, L.; Tang, M.; Wang, F.; Zhao, Y.; Liu, Z. The Impact of Different Exercise Intensities on Working Memory and BDNF Protein Expression in Prefrontal Cortex of Sleep Deprivation Rat. Int. J. Clin. Exp. Med. 2017, 10, 9265–9270. [Google Scholar]
- Lasisi, T.J.; Shittu, S.-T.T.; Abeje, J.I.; Ogunremi, K.J.; Shittu, S.A. Paradoxical Sleep Deprivation Induces Oxidative Stress in the Submandibular Glands of Wistar Rats. J. Basic Clin. Physiol. Pharmacol. 2021, 33, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Zhang, S.; Ma, W.; Wang, Q.; Li, Y.; Xia, C.; Xu, Y.; Zhang, T.; Yang, L.; Zhou, M. The Impact of Instant Coffee and Decaffeinated Coffee on the Gut Microbiota and Depression-Like Behaviors of Sleep-Deprived Rats. Front. Microbiol. 2022, 13, 778512. [Google Scholar] [CrossRef]
- Han, C.; Li, F.; Liu, Y.; Ma, J.; Yu, X.; Wu, X.; Zhang, W.; Li, D.; Chen, D.; Dai, N.; et al. Modified Si-Ni-San Decoction Ameliorates Central Fatigue by Improving Mitochondrial Biogenesis in the Rat Hippocampus. Evid. Based. Complement. Alternat. Med. 2018, 2018, 9452127. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Ji, G.; Shen, Y.; Zhao, N.; Liang, Y.; Wang, Z.; Liu, M.; Lin, L. Autophagy Triggered by Oxidative Stress Appears to Be Mediated by the AKT/MTOR Signaling Pathway in the Liver of Sleep-Deprived Rats. Oxid. Med. Cell. Longev. 2020, 2020, 6181630. [Google Scholar] [CrossRef] [PubMed]
- Nawi, A.; Eu, K.L.; Faris, A.N.A.; Wan Ahmad, W.A.N.; Noordin, L. Lipid Peroxidation in the Descending Thoracic Aorta of Rats Deprived of REM Sleep Using the Inverted Flowerpot Technique. Exp. Physiol. 2020, 105, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Gan, Z.; Li, Y.; Zhao, W.; Li, H.; Zheng, J.-P.; Ke, Y. REM Sleep Deprivation Induces Endothelial Dysfunction and Hypertension in Middle-Aged Rats: Roles of the ENOS/NO/CGMP Pathway and Supplementation with L-Arginine. PLoS ONE 2017, 12, e0182746. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, W.; Liu, M.; Zhang, Q.; Lin, Z.; Jia, M.; Liu, D.; Lin, L. Imbalance of Autophagy and Apoptosis Induced by Oxidative Stress May Be Involved in Thyroid Damage Caused by Sleep Deprivation in Rats. Oxid. Med. Cell. Longev. 2021, 2021, 5645090. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Li, K.; Dou, X.; Zhao, Y.; Huang, C.; Shu, F. The Neuroprotective Effect of Osthole against Chronic Sleep Deprivation (CSD)-Induced Memory Impairment in Rats. Life Sci. 2020, 263, 118524. [Google Scholar] [CrossRef]
- Chang, H.-M.; Lin, H.-C.; Cheng, H.-L.; Liao, C.-K.; Tseng, T.-J.; Renn, T.-Y.; Lan, C.-T.; Chen, L.-Y. Melatonin Successfully Rescues the Hippocampal Molecular Machinery and Enhances Anti-Oxidative Activity Following Early-Life Sleep Deprivation Injury. Antioxidants 2021, 10, 774. [Google Scholar] [CrossRef]
- Renn, T.-Y.; Yang, C.-P.; Wu, U.-I.; Chen, L.-Y.; Mai, F.-D.; Tikhonova, M.A.; Amstislavskaya, T.G.; Liao, W.-C.; Lin, C.-T.; Liu, Y.-C.; et al. Water Composed of Reduced Hydrogen Bonds Activated by Localized Surface Plasmon Resonance Effectively Enhances Anti-Viral and Anti-Oxidative Activities of Melatonin. Chem. Eng. J. 2022, 427, 131626. [Google Scholar] [CrossRef]
- Chen, H.-C.; Cheng, C.-Y.; Chen, L.-Y.; Chang, C.-C.; Yang, C.-P.; Mai, F.-D.; Liao, W.-C.; Chang, H.-M.; Liu, Y.-C. Plasmon-Activated Water Effectively Relieves Hepatic Oxidative Damage Resulting from Chronic Sleep Deprivation. RSC Adv. 2018, 8, 9618–9626. [Google Scholar] [CrossRef] [PubMed]
- Atrooz, F.; Liu, H.; Kochi, C.; Salim, S. Early Life Sleep Deprivation: Role of Oxido-Inflammatory Processes. Neuroscience 2019, 406, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, T.A.; Hirotsu, C.; Mazaro-Costa, R.; Tufik, S.; Andersen, M.L. Impairment of Male Reproductive Function after Sleep Deprivation. Fertil. Steril. 2015, 103, 1355–1362.e1. [Google Scholar] [CrossRef]
- Vollert, C.; Zagaar, M.; Hovatta, I.; Taneja, M.; Vu, A.; Dao, A.; Levine, A.; Alkadhi, K.; Salim, S. Exercise Prevents Sleep Deprivation-Associated Anxiety-like Behavior in Rats: Potential Role of Oxidative Stress Mechanisms. Behav. Brain Res. 2011, 224, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Martínez de Toda, I.; González-Sánchez, M.; Díaz-Del Cerro, E.; Valera, G.; Carracedo, J.; Guerra-Pérez, N. Sex Differences in Markers of Oxidation and Inflammation. Implications for Ageing. Mech. Ageing Dev. 2023, 211, 111797. [Google Scholar] [CrossRef] [PubMed]
- Hajali, V.; Andersen, M.L.; Negah, S.S.; Sheibani, V. Sex Differences in Sleep and Sleep Loss-Induced Cognitive Deficits: The Influence of Gonadal Hormones. Horm. Behav. 2019, 108, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Dib, R.; Gervais, N.J.; Mongrain, V. A Review of the Current State of Knowledge on Sex Differences in Sleep and Circadian Phenotypes in Rodents. Neurobiol. Sleep Circadian Rhythm. 2021, 11, 100068. [Google Scholar] [CrossRef]
- Wang, S.; Su, G.; Zhang, Q.; Zhao, T.; Liu, Y.; Zheng, L.; Zhao, M. Walnut (Juglans Regia) Peptides Reverse Sleep Deprivation-Induced Memory Impairment in Rat via Alleviating Oxidative Stress. J. Agric. Food Chem. 2018, 66, 10617–10627. [Google Scholar] [CrossRef]
- Hsu, J.-C.; Lee, Y.-S.; Chang, C.-N.; Chuang, H.-L.; Ling, E.-A.; Lan, C.-T. Sleep Deprivation Inhibits Expression of NADPH-d and NOS While Activating Microglia and Astroglia in the Rat Hippocampus. Cells Tissues Organs 2003, 173, 242–254. [Google Scholar] [CrossRef]
- Hirotsu, C.; Tufik, S.; Andersen, M.L. Interactions between Sleep, Stress, and Metabolism: From Physiological to Pathological Conditions. Sleep Sci. 2015, 8, 143–152. [Google Scholar] [CrossRef]
- Gong, S.; Miao, Y.-L.; Jiao, G.-Z.; Sun, M.-J.; Li, H.; Lin, J.; Luo, M.-J.; Tan, J.-H. Dynamics and Correlation of Serum Cortisol and Corticosterone under Different Physiological or Stressful Conditions in Mice. PLoS ONE 2015, 10, e0117503. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Gümüşlü, S. Immobilization Stress in Rat Tissues: Alterations in Protein Oxidation, Lipid Peroxidation and Antioxidant Defense System. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2007, 144, 342–347. [Google Scholar] [CrossRef]
- Zafir, A.; Banu, N. Induction of Oxidative Stress by Restraint Stress and Corticosterone Treatments in Rats. Indian J. Biochem. Biophys. 2009, 46, 53–58. [Google Scholar] [PubMed]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T.; Samini, F. Anti-Oxidative Effects of Curcumin on Immobilization-Induced Oxidative Stress in Rat Brain, Liver and Kidney. Biomed. Pharmacother. 2017, 87, 223–229. [Google Scholar] [CrossRef]
- Ali, F.F.; Rifaai, R.A. Preventive Effect of Omega-3 Fatty Acids in a Rat Model of Stress-Induced Liver Injury. J. Cell. Physiol. 2019, 234, 11960–11968. [Google Scholar] [CrossRef] [PubMed]
- Nirupama, M.; Devaki, M.; Nirupama, R.; Yajurvedi, H.N. Chronic Intermittent Stress-Induced Alterations in the Spermatogenesis and Antioxidant Status of the Testis Are Irreversible in Albino Rat. J. Physiol. Biochem. 2013, 69, 59–68. [Google Scholar] [CrossRef]
- García-Díaz, E.C.; Gómez-Quiroz, L.E.; Arenas-Ríos, E.; Aragón-Martínez, A.; Ibarra-Arias, J.A.; del Socorro I Retana-Márquez, M. Oxidative Status in Testis and Epididymal Sperm Parameters after Acute and Chronic Stress by Cold-Water Immersion in the Adult Rat. Syst. Biol. Reprod. Med. 2015, 61, 150–160. [Google Scholar] [CrossRef]
- Andersen, M.L.; Martins, P.J.F.; D’Almeida, V.; Bignotto, M.; Tufik, S. Endocrinological and Catecholaminergic Alterations during Sleep Deprivation and Recovery in Male Rats. J. Sleep Res. 2005, 14, 83–90. [Google Scholar] [CrossRef]
- Meerlo, P.; Koehl, M.; van der Borght, K.; Turek, F.W. Sleep Restriction Alters the Hypothalamic-Pituitary-Adrenal Response to Stress. J. Neuroendocrinol. 2002, 14, 397–402. [Google Scholar] [CrossRef]
- Suchecki, D.; Tiba, P.A.; Tufik, S. Paradoxical Sleep Deprivation Facilitates Subsequent Corticosterone Response to a Mild Stressor in Rats. Neurosci. Lett. 2002, 320, 45–48. [Google Scholar] [CrossRef]
- Suchecki, D.; Lobo, L.L.; Hipólide, D.C.; Tufik, S. Increased ACTH and Corticosterone Secretion Induced by Different Methods of Paradoxical Sleep Deprivation. J. Sleep Res. 1998, 7, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Coenen, A.M.; van Luijtelaar, E.L. Stress Induced by Three Procedures of Deprivation of Paradoxical Sleep. Physiol. Behav. 1985, 35, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Nollet, M.; Wisden, W.; Franks, N.P. Sleep Deprivation and Stress: A Reciprocal Relationship. Interface Focus 2020, 10, 20190092. [Google Scholar] [CrossRef]
- Machado, R.B.; Hipólide, D.C.; Benedito-Silva, A.A.; Tufik, S. Sleep Deprivation Induced by the Modified Multiple Platform Technique: Quantification of Sleep Loss and Recovery. Brain Res. 2004, 1004, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Suchecki, D.; Machado, R.B.; Tiba, P.A. Stress-Induced Sleep Rebound: Adaptive Behavior and Possible Mechanisms. Sleep Sci. 2009, 2, 151–160. [Google Scholar]
- Machado, R.B.; Tufik, S.; Suchecki, D. Role of Corticosterone on Sleep Homeostasis Induced by REM Sleep Deprivation in Rats. PLoS ONE 2013, 8, e63520. [Google Scholar] [CrossRef] [PubMed]
- Melgarejo-Gutiérrez, M.; Acosta-Peña, E.; Venebra-Muñoz, A.; Escobar, C.; Santiago-García, J.; Garcia-Garcia, F. Sleep Deprivation Reduces Neuroglobin Immunoreactivity in the Rat Brain. Neuroreport 2013, 24, 120–125. [Google Scholar] [CrossRef]
- Gopalakrishnan, A.; Ji, L.L.; Cirelli, C. Sleep Deprivation and Cellular Responses to Oxidative Stress. Sleep 2004, 27, 27–35. [Google Scholar] [CrossRef]
- Vaccaro, A.; Kaplan Dor, Y.; Nambara, K.; Pollina, E.A.; Lin, C.; Greenberg, M.E.; Rogulja, D. Sleep Loss Can Cause Death through Accumulation of Reactive Oxygen Species in the Gut. Cell 2020, 181, 1307–1328.e15. [Google Scholar] [CrossRef]
- Briançon-Marjollet, A.; Weiszenstein, M.; Henri, M.; Thomas, A.; Godin-Ribuot, D.; Polak, J. The Impact of Sleep Disorders on Glucose Metabolism: Endocrine and Molecular Mechanisms. Diabetol. Metab. Syndr. 2015, 7, 25. [Google Scholar] [CrossRef]
- Morselli, L.; Leproult, R.; Balbo, M.; Spiegel, K. Role of Sleep Duration in the Regulation of Glucose Metabolism and Appetite. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Boyle, P.J.; Scott, J.C.; Krentz, A.J.; Nagy, R.J.; Comstock, E.; Hoffman, C. Diminished Brain Glucose Metabolism Is a Significant Determinant for Falling Rates of Systemic Glucose Utilization during Sleep in Normal Humans. J. Clin. Investig. 1994, 93, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Holmer, B.J.; Lapierre, S.S.; Jake-Schoffman, D.E.; Christou, D.D. Effects of Sleep Deprivation on Endothelial Function in Adult Humans: A Systematic Review. Geroscience 2021, 43, 137–158. [Google Scholar] [CrossRef] [PubMed]
- Palagini, L.; Bruno, R.M.; Gemignani, A.; Baglioni, C.; Ghiadoni, L.; Riemann, D. Sleep Loss and Hypertension: A Systematic Review. Curr. Pharm. Des. 2013, 19, 2409–2419. [Google Scholar] [CrossRef]
- Sá Gomes E Farias, A.V.; de Lima Cavalcanti, M.P.; de Passos Junior, M.A.; Vechio Koike, B.D. The Association between Sleep Deprivation and Arterial Pressure Variations: A Systematic Literature Review. Sleep Med. X 2022, 4, 100042. [Google Scholar] [CrossRef] [PubMed]
- Xi, B.; He, D.; Zhang, M.; Xue, J.; Zhou, D. Short Sleep Duration Predicts Risk of Metabolic Syndrome: A Systematic Review and Meta-Analysis. Sleep Med. Rev. 2014, 18, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Antza, C.; Kostopoulos, G.; Mostafa, S.; Nirantharakumar, K.; Tahrani, A. The Links between Sleep Duration, Obesity and Type 2 Diabetes Mellitus. J. Endocrinol. 2021, 252, 125–141. [Google Scholar] [CrossRef]
- Knutson, K.L.; Spiegel, K.; Penev, P.; Van Cauter, E. The Metabolic Consequences of Sleep Deprivation. Sleep Med. Rev. 2007, 11, 163–178. [Google Scholar] [CrossRef]
- Bishir, M.; Bhat, A.; Essa, M.M.; Ekpo, O.; Ihunwo, A.O.; Veeraraghavan, V.P.; Mohan, S.K.; Mahalakshmi, A.M.; Ray, B.; Tuladhar, S.; et al. Sleep Deprivation and Neurological Disorders. Biomed Res. Int. 2020, 2020, 5764017. [Google Scholar] [CrossRef]
- Xue, R.; Wan, Y.; Sun, X.; Zhang, X.; Gao, W.; Wu, W. Nicotinic Mitigation of Neuroinflammation and Oxidative Stress After Chronic Sleep Deprivation. Front. Immunol. 2019, 10, 2546. [Google Scholar] [CrossRef]
- Chen, P.; Ban, W.; Wang, W.; You, Y.; Yang, Z. The Devastating Effects of Sleep Deprivation on Memory: Lessons from Rodent Models. Clocks Sleep 2023, 5, 276–294. [Google Scholar] [CrossRef]
- Tufik, S.; Andersen, M.L.; Bittencourt, L.R.A.; de Mello, M.T. Paradoxical Sleep Deprivation: Neurochemical, Hormonal and Behavioral Alterations. Evidence from 30 Years of Research. An. Acad. Bras. Cienc. 2009, 81, 521–538. [Google Scholar] [CrossRef]
- Maloney, K.J.; Mainville, L.; Jones, B.E. Differential C-Fos Expression in Cholinergic, Monoaminergic, and GABAergic Cell Groups of the Pontomesencephalic Tegmentum after Paradoxical Sleep Deprivation and Recovery. J. Neurosci. 1999, 19, 3057–3072. [Google Scholar] [CrossRef]
- Suchecki, D.; Tufik, S. Social Stability Attenuates the Stress in the Modified Multiple Platform Method for Paradoxical Sleep Deprivation in the Rat. Physiol. Behav. 2000, 68, 309–316. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neculicioiu, V.S.; Colosi, I.A.; Costache, C.; Toc, D.A.; Sevastre-Berghian, A.; Colosi, H.A.; Clichici, S. Sleep Deprivation-Induced Oxidative Stress in Rat Models: A Scoping Systematic Review. Antioxidants 2023, 12, 1600. https://doi.org/10.3390/antiox12081600
Neculicioiu VS, Colosi IA, Costache C, Toc DA, Sevastre-Berghian A, Colosi HA, Clichici S. Sleep Deprivation-Induced Oxidative Stress in Rat Models: A Scoping Systematic Review. Antioxidants. 2023; 12(8):1600. https://doi.org/10.3390/antiox12081600
Chicago/Turabian StyleNeculicioiu, Vlad Sever, Ioana Alina Colosi, Carmen Costache, Dan Alexandru Toc, Alexandra Sevastre-Berghian, Horațiu Alexandru Colosi, and Simona Clichici. 2023. "Sleep Deprivation-Induced Oxidative Stress in Rat Models: A Scoping Systematic Review" Antioxidants 12, no. 8: 1600. https://doi.org/10.3390/antiox12081600
APA StyleNeculicioiu, V. S., Colosi, I. A., Costache, C., Toc, D. A., Sevastre-Berghian, A., Colosi, H. A., & Clichici, S. (2023). Sleep Deprivation-Induced Oxidative Stress in Rat Models: A Scoping Systematic Review. Antioxidants, 12(8), 1600. https://doi.org/10.3390/antiox12081600