
MOMAST® Reduces the Plasmatic Lipid Profile and Oxidative Stress and Regulates Cholesterol Metabolism in a Hypercholesterolemic Mouse Model: The Proof of Concept of a Sustainable and Innovative Antioxidant and Hypocholesterolemic Ingredient

 ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
MOMAST® Description
2.2. In Vitro MOMAST® Antioxidant Activity
2.2.1. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Assay
2.2.2. Ferric Reducing Antioxidant Power (FRAP) Assay
2.3. Study Design
2.4. Biochemical Parameters
2.5. In Vivo Antioxidant Activity of MOMAST®
2.5.1. DPPH and FRAP Assays
2.5.2. Determination of Hepatic Malondialdehyde (MDA) Levels
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
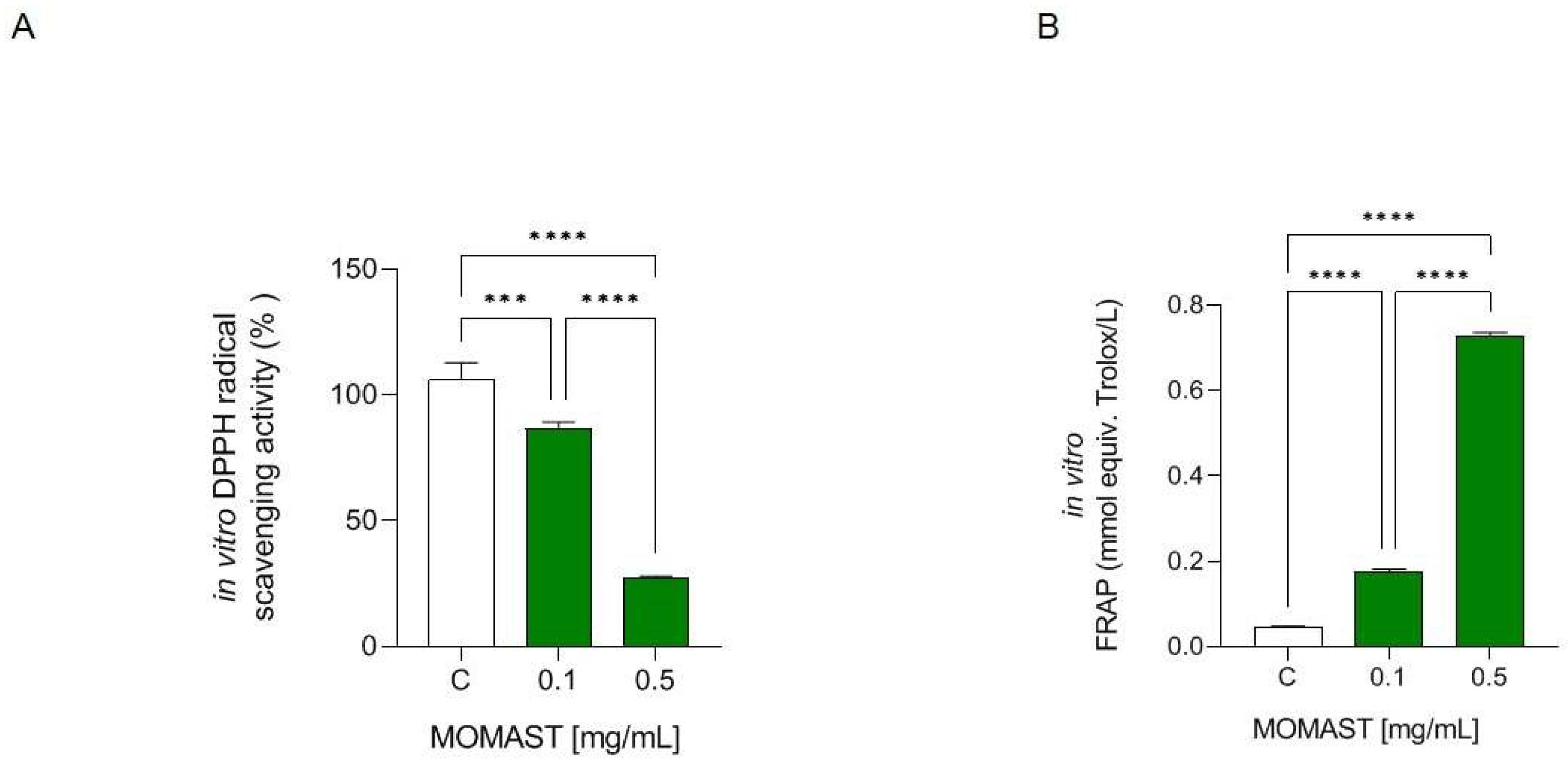
3.1. MOMAST® Has In Vitro DPPH Radical Scavenging and Ferric Reducing Capacities
3.2. Effect of MOMAST® on Body Weight
3.3. MOMAST® Exerts Antioxidant Effects at Serum and Hepatic Levels
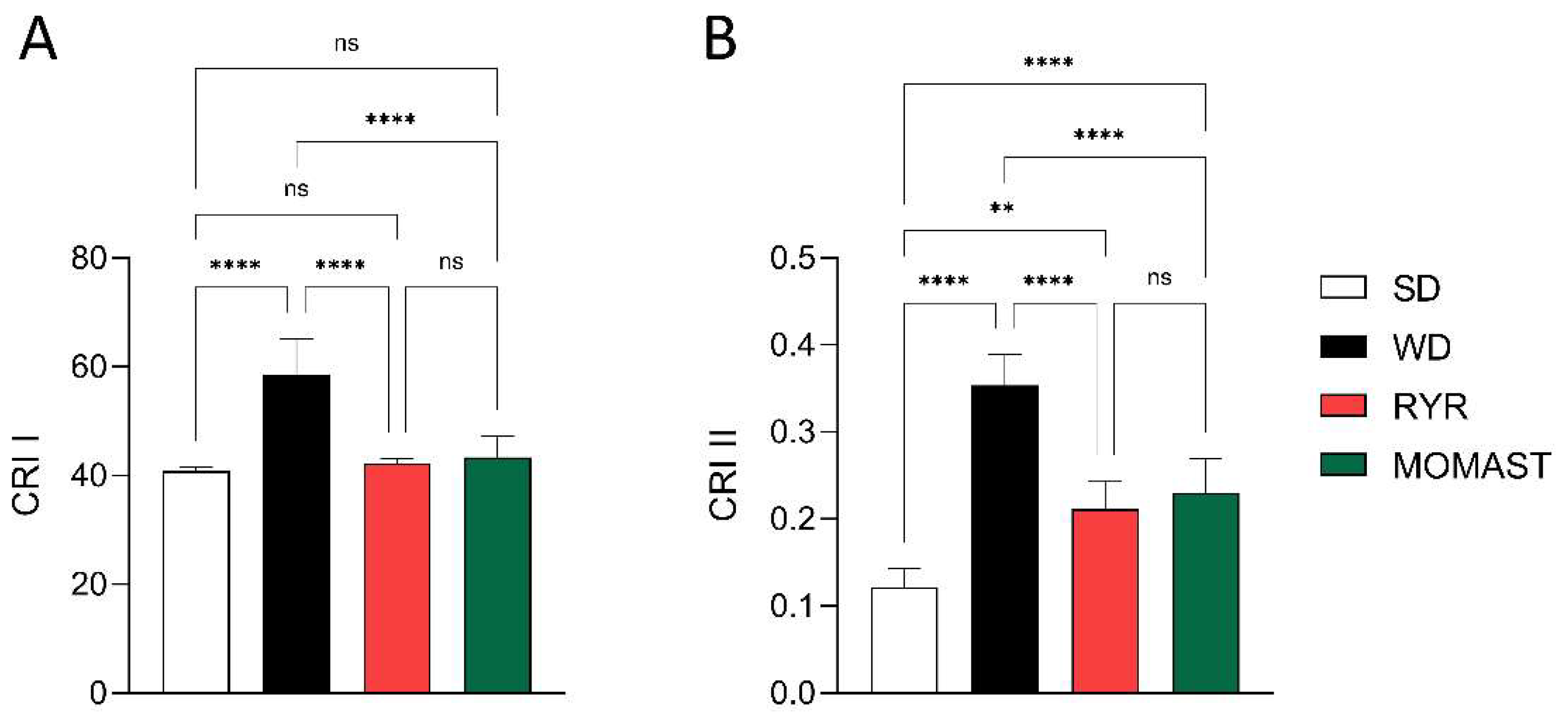
3.4. MOMAST® Improves the Plasmatic Lipidic Profile
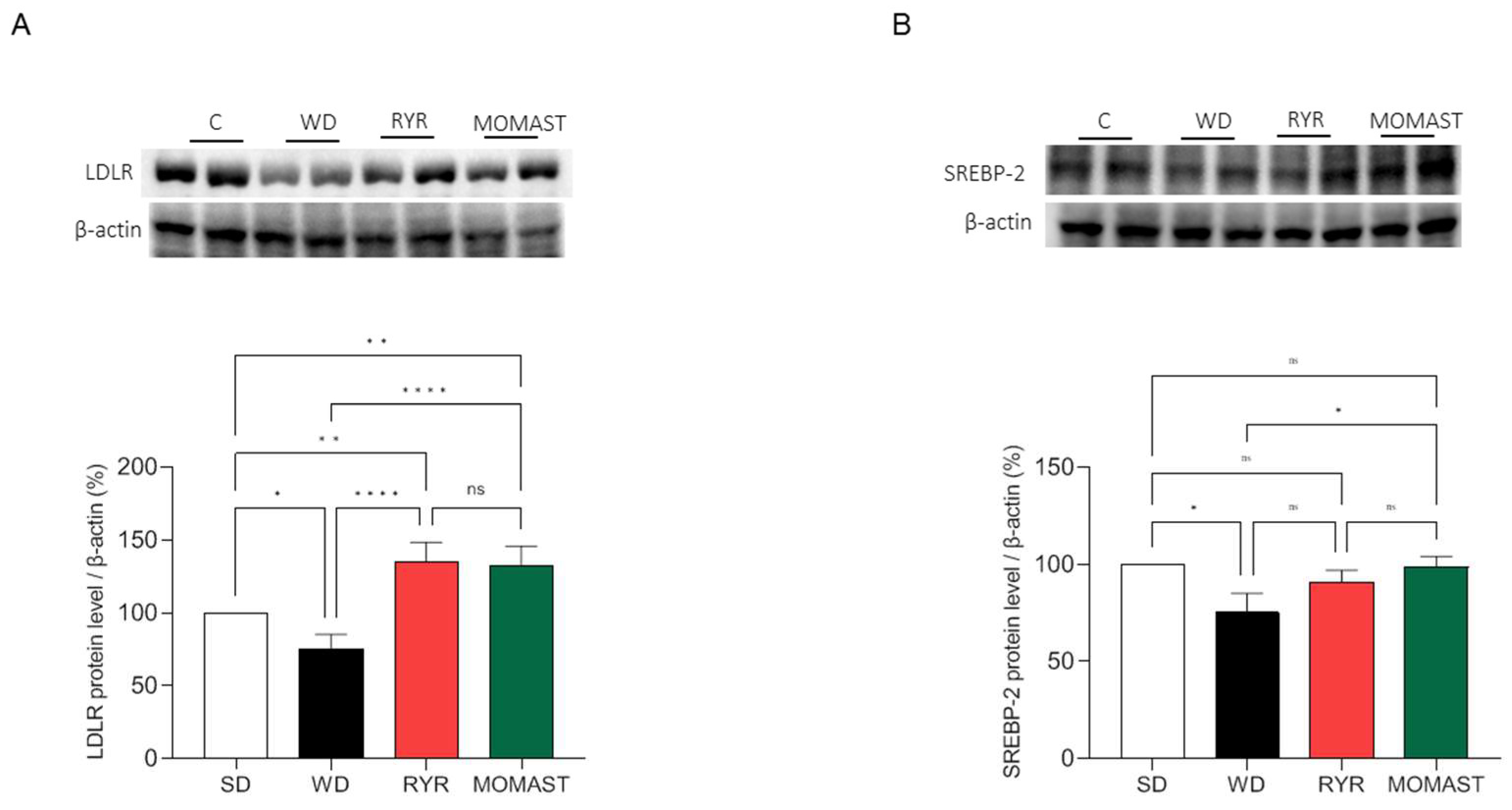
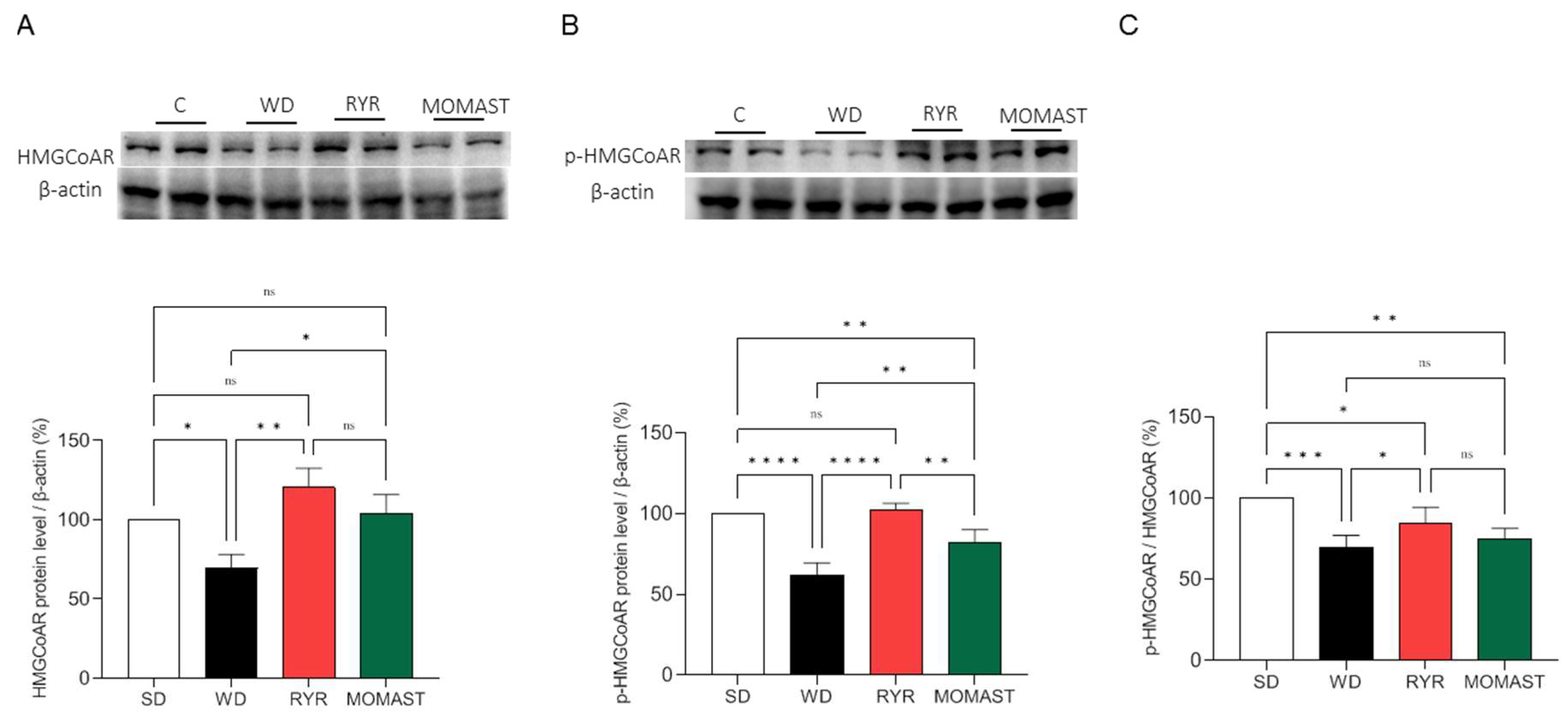
3.5. MOMAST® Activates the SREBP-2/LDLR Pathway and Modulates the Active HMGCoAR Enzyme
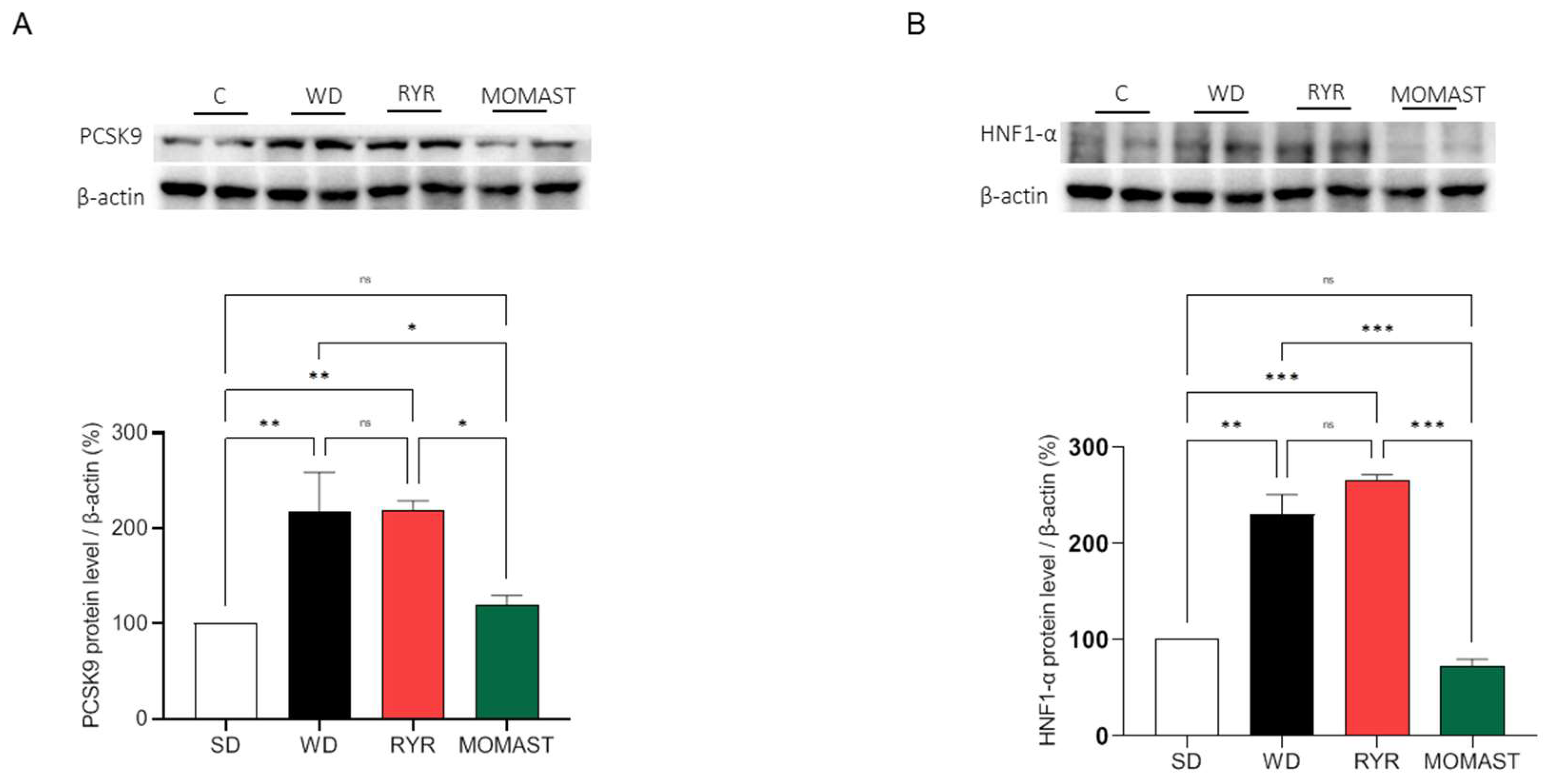
3.6. MOMAST® Treatment Reduces the PCSK9 Protein Levels Increased by WD Ingestion
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obied, H.K.; Karuso, P.; Prenzler, P.D.; Robards, K. Novel Secoiridoids with Antioxidant Activity from Australian Olive Mill Waste. J. Agric. Food Chem. 2007, 55, 2848–2853. [Google Scholar] [CrossRef]
- Mulinacci, N.; Romani, A.; Galardi, C.; Pinelli, P.; Giaccherini, C.; Vincieri, F.F. Polyphenolic Content in Olive Oil Waste Waters and Related Olive Samples. J. Agric. Food Chem. 2001, 49, 3509–3514. [Google Scholar] [CrossRef]
- De Luca, P.; Sicilia, V.; Candamano, S.; Macario, A. Olive Vegetation Waters (OVWs): Characteristics, Treatments and Environmental Problems. IOP Conf. Ser. Mater. Sci. Eng. 2022, 1251, 012011. [Google Scholar] [CrossRef]
- Bartolomei, M.; Bollati, C.; Li, J.; Arnoldi, A.; Lammi, C. Assessment of the Cholesterol-Lowering Effect of MOMAST®: Biochemical and Cellular Studies. Nutrients 2022, 14, 493. [Google Scholar] [CrossRef]
- McBean, G.A. Integrating Science to Address Food and Health within Global Agenda 2030. npj Sci. Food 2021, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, J.; Haumont, M.; Prost-Camus, E.; Durand, P.; Prost, M.; Lizard, G.H.; Latruffe, N. Protection of C2C12 Skeletal Muscle Cells towards Oxidation by a Polyphenol Rich Plant Extract. Redox Exp. Med. 2023, 2023, e230002. [Google Scholar] [CrossRef]
- Aires, V.; Labbé, J.; Deckert, V.; Pais de Barros, J.P.; Boidot, R.; Haumont, M.; Maquart, G.; Le Guern, N.; Masson, D.; Prost-Camus, E.; et al. Healthy Adiposity and Extended Lifespan in Obese Mice Fed a Diet Supplemented with a Polyphenol-Rich Plant Extract. Sci. Rep. 2019, 9, 9134. [Google Scholar] [CrossRef] [PubMed]
- Recinella, L.; Chiavaroli, A.; Orlando, G.; Menghini, L.; Ferrante, C.; Di Cesare Mannelli, L.; Ghelardini, C.; Brunetti, L.; Leone, S. Protective Effects Induced by Two Polyphenolic Liquid Complexes from Olive (Olea Europaea, Mainly Cultivar Coratina) Pressing Juice in Rat Isolated Tissues Challenged with LPS. Molecules 2019, 24, 3002. [Google Scholar] [CrossRef]
- Lammi, C.; Aiello, G.; Bollati, C.; Li, J.; Bartolomei, M.; Ranaldi, G.; Ferruzza, S.; Fassi, E.M.A.; Grazioso, G.; Sambuy, Y.; et al. Trans-epithelial Transport, Metabolism and Biological Activity Assessment of the Multi-target Lupin Peptide Lilpkhsdad (P5) and Its Metabolite Lpkhsdad (P5-met). Nutrients 2021, 13, 863. [Google Scholar] [CrossRef]
- Lammi, C.; Bellumori, M.; Cecchi, L.; Bartolomei, M.; Bollati, C.; Clodoveo, M.L.; Corbo, F.; Arnoldi, A.; Mulinacci, N. Extra Virgin Olive Oil Phenol Extracts Exert Hypocholesterolemic Effects through the Modulation of the LDLR Pathway: In Vitro and Cellular Mechanism of Action Elucidation. Nutrients 2020, 12, 1723. [Google Scholar] [CrossRef]
- Pugliese, G.; Liccardi, A.; Graziadio, C.; Barrea, L.; Muscogiuri, G.; Colao, A. Obesity and Infectious Diseases: Pathophysiology and Epidemiology of a Double Pandemic Condition. Int. J. Obes. 2022, 46, 449–465. [Google Scholar] [CrossRef]
- De Lorenzo, A.; Gratteri, S.; Gualtieri, P.; Cammarano, A.; Bertucci, P.; Di Renzo, L. Why Primary Obesity Is a Disease? J. Transl. Med. 2019, 17, 169. [Google Scholar] [CrossRef] [PubMed]
- Al-Jawaldeh, A.; Abbass, M.M.S. Unhealthy Dietary Habits and Obesity: The Major Risk Factors Beyond Non-Communicable Diseases in the Eastern Mediterranean Region. Front. Nutr. 2022, 9, 817808. [Google Scholar] [CrossRef] [PubMed]
- Pi-Sunyer, X. The Medical Risks of Obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef]
- Nagao, M.; Nakajima, H.; Toh, R.; Hirata, K.I.; Ishida, T. Cardioprotective Effects of High-Density Lipoprotein beyond Its Anti-Atherogenic Action. J. Atheroscler. Thromb. 2018, 25, 985–993. [Google Scholar] [CrossRef]
- Cai, G.; Shi, G.; Xue, S.; Lu, W. The Atherogenic Index of Plasma Is a Strong and Independent Predictor for Coronary Artery Disease in the Chinese Han Population. Medicine 2017, 96, e8058. [Google Scholar] [CrossRef]
- Olamoyegun, M.; Oluyombo, R.; Asaolu, S. Evaluation of Dyslipidemia, Lipid Ratios, and Atherogenic Index as Cardiovascular Risk Factors among Semi-Urban Dwellers in Nigeria. Ann. Afr. Med. 2016, 15, 194. [Google Scholar] [CrossRef]
- Millán, J.; Pintó, X.; Muñoz, A.; Zúñiga, M.; Rubiés-Prat, J.; Pallardo, L.F.; Masana, L.; Mangas, A.; Hernández-Mijares, A.; González-Santos, P.; et al. Lipoprotein Ratios: Physiological Significance and Clinical Usefulness in Cardiovascular Prevention. Vasc. Health Risk Manag. 2009, 5, 757–765. [Google Scholar] [PubMed]
- Zammit, V.A.; Easom, R.A. Regulation of Hepatic HMG-CoA Reductase in Vivo by Reversible Phosphorylation. BBA Mol. Cell Res. 1987, 927, 223–228. [Google Scholar] [CrossRef]
- Parker, R.A.; Miller, S.J.; Gibson, D.M. Phosphorylation State of HMG CoA Reductase Affects Its Catalytic Activity and Degradation. Adv. Enzym. Regul. 1986, 25, 329–343. [Google Scholar] [CrossRef]
- Zou, J.; Yan, C.; Wan, J.B. Red Yeast Rice Ameliorates Non-Alcoholic Fatty Liver Disease through Inhibiting Lipid Synthesis and NF-ΚB/NLRP3 Inflammasome-Mediated Hepatic Inflammation in Mice. Chin. Med. 2022, 17, 17. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Fogacci, F.; Banach, M. Red Yeast Rice for Hypercholesterolemia. Methodist Debakey Cardiovasc. J. 2019, 15, 192. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.N.; Fisher, E.A.; Breslow, J.L. Overexpression of PCSK9 Accelerates the Degradation of the LDLR in a Post-Endoplasmic Reticulum Compartment. Proc. Natl. Acad. Sci. USA 2005, 102, 2069–2074. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Chamorro, I.; Santos-Sánchez, G.; Ponce-España, E.; Bollati, C.; d’Adduzio, L.; Bartolomei, M.; Li, J.; Carrillo-Vico, A.; Lammi, C. MOMAST® Reduces the Plasmatic Lipid Profile and Oxidative Stress and Regulates Cholesterol Metabolism in a Hypercholesterolemic Mouse Model: The Proof of Concept of a Sustainable and Innovative Antioxidant and Hypocholesterolemic Ingredient. Antioxidants 2023, 12, 1335. https://doi.org/10.3390/antiox12071335
Cruz-Chamorro I, Santos-Sánchez G, Ponce-España E, Bollati C, d’Adduzio L, Bartolomei M, Li J, Carrillo-Vico A, Lammi C. MOMAST® Reduces the Plasmatic Lipid Profile and Oxidative Stress and Regulates Cholesterol Metabolism in a Hypercholesterolemic Mouse Model: The Proof of Concept of a Sustainable and Innovative Antioxidant and Hypocholesterolemic Ingredient. Antioxidants. 2023; 12(7):1335. https://doi.org/10.3390/antiox12071335
Chicago/Turabian StyleCruz-Chamorro, Ivan, Guillermo Santos-Sánchez, Eduardo Ponce-España, Carlotta Bollati, Lorenza d’Adduzio, Martina Bartolomei, Jianqiang Li, Antonio Carrillo-Vico, and Carmen Lammi. 2023. "MOMAST® Reduces the Plasmatic Lipid Profile and Oxidative Stress and Regulates Cholesterol Metabolism in a Hypercholesterolemic Mouse Model: The Proof of Concept of a Sustainable and Innovative Antioxidant and Hypocholesterolemic Ingredient" Antioxidants 12, no. 7: 1335. https://doi.org/10.3390/antiox12071335
APA StyleCruz-Chamorro, I., Santos-Sánchez, G., Ponce-España, E., Bollati, C., d’Adduzio, L., Bartolomei, M., Li, J., Carrillo-Vico, A., & Lammi, C. (2023). MOMAST® Reduces the Plasmatic Lipid Profile and Oxidative Stress and Regulates Cholesterol Metabolism in a Hypercholesterolemic Mouse Model: The Proof of Concept of a Sustainable and Innovative Antioxidant and Hypocholesterolemic Ingredient. Antioxidants, 12(7), 1335. https://doi.org/10.3390/antiox12071335