
Blueberry Supplementation and Skin Health



Abstract

1. Introduction
2. Cutaneous Anatomy and Physiology
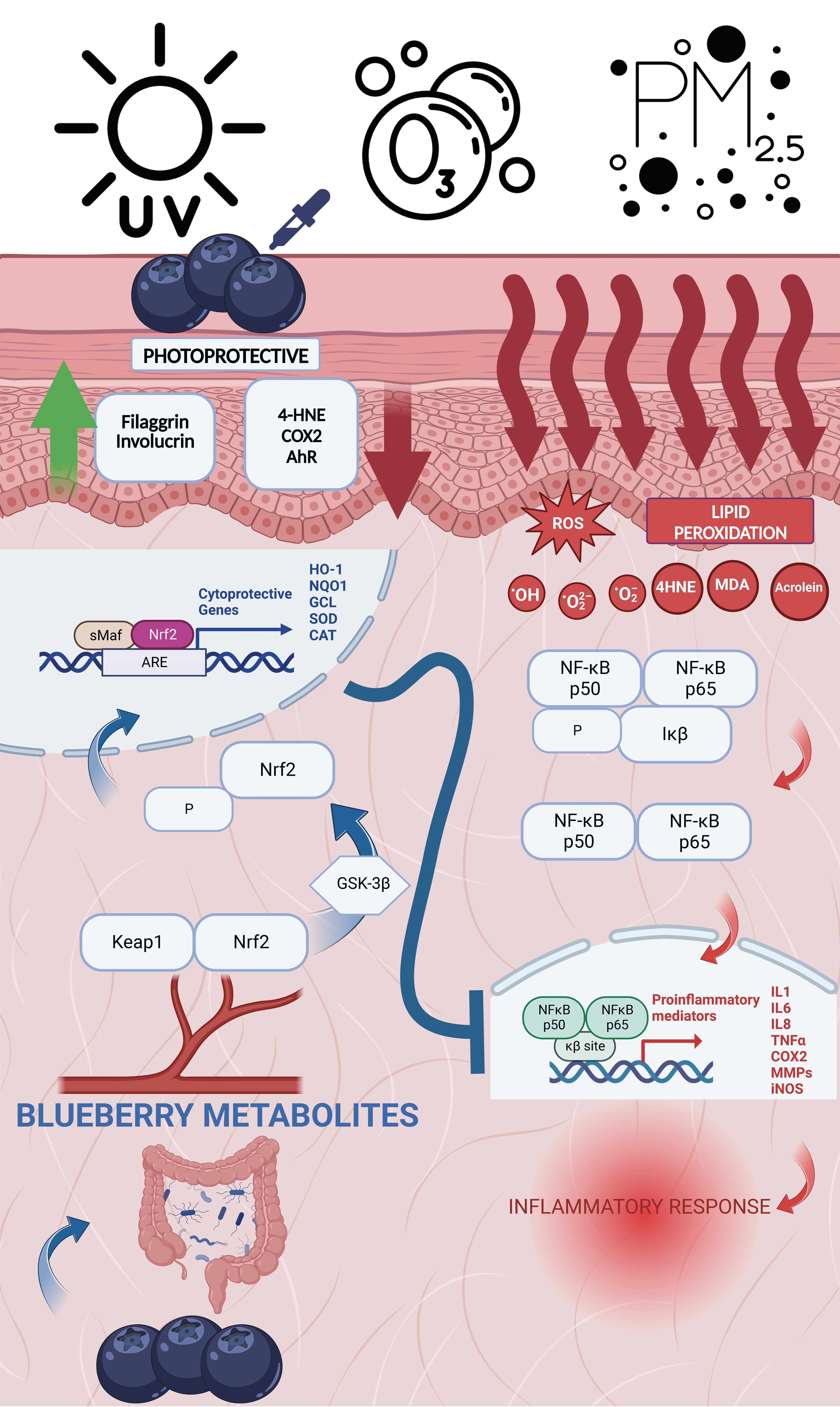
3. Principal Environmental Stressors to Cutaneous Systems
4. Blueberry Phytoactives
5. Topical Application
6. Dietary Supplementation
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maya-Cano, D.; Arango-Varela, S.; Santa-Gonzalez, G. Phenolic compounds of blueberries (Vaccinium spp.) as a protective strategy against skin cell damage induced by ROS: A review of antioxidant potential and antiproliferative capacity. Heliyon 2021, 7, e06297. [Google Scholar] [CrossRef]
- Chilcott, R.P.; Price, S. Principles and Practice of Skin Toxicology; John Wiley & Sons: Hoboken, NJ, USA, 2008; p. 370. [Google Scholar]
- Stenqvist, B.O.; Sparr, E. Tortuosity in the Brick and Mortar Model Based on Chemical Conduction. Chem. Eng. Sci. 2020, 223, 115729. [Google Scholar] [CrossRef]
- Menon, G.K.; Cleary, G.W.; Lane, M.E. The structure and function of the stratum corneum. Int. J. Pharm. 2012, 435, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Woodby, B.; Penta, K.; Pecorelli, A.; Lila, M.A.; Valacchi, G. Skin Health from the Inside Out. Annu. Rev. Food Sci. Technol. 2020, 11, 235–254. [Google Scholar] [CrossRef]
- Yousef, H.; Alhajj, M.; Sharma, S. Anatomy, Skin (Integument), Epidermis; StatPearls Publishing LLC: Treasure Island, FL, USA, 2022. [Google Scholar]
- Watt, F.M.; Fujiwara, H. Cell-extracellular matrix interactions in normal and diseased skin. Cold Spring Harb. Perspect. Biol. 2011, 3, a005124. [Google Scholar] [CrossRef]
- McKnight, G.; Shah, J.; Hargest, R. Physiology of the skin. Surgery 2022, 40, 8–12. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Aguilar, T.A.F.; Navarro, B.C.H.; Pérez, J.A.M. Endogenous Antioxidants: A Review of their Role in Oxidative Stress. In The Transcription Factor Nrf2; Jose, A.M.-G., Angel, M.-G., Eduardo, O.M.-S., Eds.; IntechOpen: Rijeka, Croatia, 2016. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Godic, A.; Poljšak, B.; Adamic, M.; Dahmane, R. The role of antioxidants in skin cancer prevention and treatment. Oxid. Med. Cell Longev. 2014, 2014, 860479. [Google Scholar] [CrossRef] [PubMed]
- Keen, M.A.; Hassan, I. Vitamin E in dermatology. Indian Derm. Online J. 2016, 7, 311–315. [Google Scholar] [CrossRef]
- Shim, J.H. Prostaglandin E2 induces skin aging via e-prostanoid 1 in normal human dermal fibroblasts. Int. J. Mol. Sci. 2019, 20, 5555. [Google Scholar] [CrossRef]
- Serbecic, N.; Beutelspacher, S.C. Vitamins inhibit oxidant-induced apoptosis of corneal endothelial cells. Jpn. J. Ophthalmol. 2005, 49, 355–362. [Google Scholar] [CrossRef]
- Pullar, J.M.; Carr, A.C.; Vissers, M.C.M. The roles of vitamin C in skin health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef] [PubMed]
- Pambianchi, E. Topical Intervention to Prevent Pollution Induced Extrinsic Aging and Aberrant OxInflammatory Responses in Orthodox and Innovative Skin Models; North Carolina State University: Ann Arbor, MI, USA, 2021. [Google Scholar]
- Matsumura, Y.; Ananthaswamy, H.N. Toxic effects of ultraviolet radiation on the skin. Toxicol. Appl. Pharmacol. 2004, 195, 298–308. [Google Scholar] [CrossRef]
- Valacchi, G.; Sticozzi, C.; Pecorelli, A.; Cervellati, F.; Cervellati, C.; Maioli, E. Cutaneous responses to environmental stressors. Ann. N. Y. Acad. Sci. 2012, 1271, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.B.; Gniadecki, R.; Vicanova, J.; Thorn, T.; Wulf, H.C. Hydrogen peroxide is responsible for UVA-induced DNA damage measured by alkaline comet assay in HaCaT keratinocytes. J. Photochem. Photobiol. B Biol. 2000, 59, 123–131. [Google Scholar] [CrossRef]
- Zegarska, B.; Pietkun, K.; Zegarski, W.; Bolibok, P.; Wiśniewski, M.; Roszek, K.; Czarnecka, J.; Nowacki, M. Air pollution, UV irradiation and skin carcinogenesis: What we know, where we stand and what is likely to happen in the future? Adv. Dermatol. Allergol. 2017, 34, 6–14. [Google Scholar] [CrossRef]
- McDaniel, D.; Farris, P.; Valacchi, G. Atmospheric skin aging-Contributors and inhibitors. J. Cosmet. Dermatol. 2018, 17, 124–137. [Google Scholar] [CrossRef]
- Lelieveld, J.; Dentener, F.J. What controls tropospheric ozone? J. Geophys. Res. Atmos. 2000, 105, 3531–3551. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, Y.; Fang, Z. Ozone pollution: A major health hazard worldwide. Front. Immunol. 2019, 10, 2518. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Regional Office, for Europe Review of Evidence on Health Aspects of Air Pollution: REVIHAAP Project: Technical Report. 2021. Available online: https://apps.who.int/iris/handle/10665/341712 (accessed on 7 March 2023).
- Packer, L.; Valacchi, G. Antioxidants and the response of skin to oxidative stress: Vitamin E as a key indicator. Skin Pharmacol. Appl. Skin Physiol. 2002, 15, 282–290. [Google Scholar] [CrossRef]
- Valacchi, G.; van der Vliet, A.; Schock, B.C.; Okamoto, T.; Obermuller-Jevic, U.; Cross, C.E.; Packer, L. Ozone exposure activates oxidative stress responses in murine skin. Toxxicology 2002, 179, 163–170. [Google Scholar] [CrossRef]
- Fuks, K.B.; Woodby, B.; Valacchi, G. Skin damage by tropospheric ozone. Hautarzt 2019, 70, 163–168. [Google Scholar] [CrossRef]
- Farris, P.K.; Valacchi, G. Ultraviolet light protection: Is it really enough? Antioxidants 2022, 11, 1484. [Google Scholar] [CrossRef]
- Pecorelli, A.; Woodby, B.; Prieux, R.; Valacchi, G. Involvement of 4-hydroxy-2-nonenal in pollution-induced skin damage. Biofactors 2019, 45, 536–547. [Google Scholar] [CrossRef]
- Valacchi, G.; Pagnin, E.; Okamoto, T.; Corbacho, A.M.; Olano, E.; Davis, P.A.; Vliet, A.v.d.; Packer, L.; Cross, C.E. Induction of stress proteins and MMP-9 by 0.8ppm of ozone in murine skin. Biochem. Biophys. Res. Commun. 2003, 305, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Burnett, R.; Chen, H.; Szyszkowicz, M.; Fann, N.; Hubbell, B.; Pope, C.A., III; Apte, J.S.; Brauer, M.; Cohen, A.; Weichenthal, S.; et al. Global estimates of mortality associated with long-term exposure to outdoor fine particulate matter. Proc. Natl. Acad. Sci. USA 2018, 115, 9592–9597. [Google Scholar] [CrossRef]
- Kim, B.E.; Kim, J.; Goleva, E.; Berdyshev, E.; Lee, J.; Vang, K.A.; Lee, U.H.; Han, S.; Leung, S.; Hall, C.F.; et al. Particulate matter causes skin barrier dysfunction. JCI Insight 2021, 6, e145185. [Google Scholar] [CrossRef]
- Li, N.; Sioutas, C.; Cho, A.; Schmitz, D.; Misra, C.; Sempf, J.; Wang, M.; Oberley, T.; Froines, J.; Nel, A. Ultrafine particulate pollutants induce oxidative stress and mitochondrial damage. Environ. Health Perspect. 2003, 111, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Magnani, N.D.; Muresan, X.M.; Belmonte, G.; Cervellati, F.; Sticozzi, C.; Pecorelli, A.; Miracco, C.; Marchini, T.; Evelson, P.; Valacchi, G. Skin Damage Mechanisms Related to Airborne Particulate Matter Exposure. Toxicol. Sci. 2016, 149, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent research on the health benefits of blueberries and their anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Guo, Y.; Liu, M.; Chen, X.; Xiao, X.; Wang, S.; Gong, P.; Ma, Y.; Chen, F. Structure and function of blueberry anthocyanins: A review of recent advances. J. Funct. Foods 2022, 88, 104864. [Google Scholar] [CrossRef]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Mazza, G.; Kay, C.D.; Cottrell, T.; Holub, B.J. Absorption of anthocyanins from blueberries and serum antioxidant status in human subjects. J. Agric. Food Chem. 2002, 50, 7731–7737. [Google Scholar] [CrossRef]
- Chandra, P.; Rathore, A.S.; Kay, K.L.; Everhart, J.L.; Curtis, P.; Burton-Freeman, B.; Cassidy, A.; Kay, C.D. Contribution of Berry Polyphenols to the Human Metabolome. Molecules 2019, 24, 4220. [Google Scholar] [CrossRef]
- Carvalho Templeton, M. Evaluating Plant-Derived (Poly)phenol Bioavailability via Broad-Spectrum Metabolomics Using Highbush Blueberries as a Model Fruit. Ph.D. Dissertation, University of North Carolina State, Raleigh, NC, USA, 2022. Available online: https://www.lib.ncsu.edu/resolver/1840.20/39930 (accessed on 7 March 2023).
- Rathinasabapathy, T.; Lomax, J.; Srikanth, K.; Esposito, D.; Kay, C.D.; Komarnytsky, S. Effect of wild blueberry metabolites on biomarkers of gastrointestinal and immune health in vitro. Immuno 2022, 2, 293–306. [Google Scholar] [CrossRef]
- Duan, Y.; Tarafdar, A.; Chaurasia, D.; Singh, A.; Bhargava, P.C.; Yang, J.; Li, Z.; Ni, X.; Tian, Y.; Li, H.; et al. Blueberry fruit valorization and valuable constituents: A review. Int. J. Food Microbiol. 2022, 381, 109890. [Google Scholar] [CrossRef]
- Liao, M.; Ma, L.; Miao, S.; Hu, X.; Liao, X.; Chen, F.; Ji, J. The in-vitro digestion behaviors of milk proteins acting as wall materials in spray-dried microparticles: Effects on the release of loaded blueberry anthocyanins. Food Hydrocoll. 2021, 115, 106620. [Google Scholar] [CrossRef]
- Mukherjee, S.; Date, A.; Patravale, V.; Korting, H.C.; Roeder, A.; Weindl, G. Retinoids in the treatment of skin aging: An overview of clinical efficacy and safety. Clin. Interv. Aging 2006, 1, 327–348. [Google Scholar] [CrossRef]
- VanBuren, C.A.; Everts, H.B. Vitamin A in skin and hair: An update. Nutrients 2022, 14, 2952. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mateos, A.; Feliciano, R.P.; Cifuentes-Gomez, T.; Spencer, J.P.E. Bioavailability of wild blueberry (poly)phenols at different levels of intake. J. Berry Res. 2016, 6, 137–148. [Google Scholar] [CrossRef]
- Eberlein-König, B.; Ring, J. Relevance of vitamins C and E in cutaneous photoprotection. J. Cosmet. Dermatol. 2005, 4, 4–9. [Google Scholar] [CrossRef]
- Aguilera, J.; de Gálvez, M.V.; Sánchez, C.; Herrera-Ceballos, E. Changes in photoinduced cutaneous erythema with topical application of a combination of vitamins C and E before and after UV exposure. J. Dermatol. Sci. 2012, 66, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Valacchi, G.; Muresan, X.M.; Sticozzi, C.; Belmonte, G.; Pecorelli, A.; Cervellati, F.; Demaude, J.; Krol, Y.; Oresajo, C. Ozone-induced damage in 3D-Skin Model is prevented by topical vitamin C and vitamin E compound mixtures application. J. Dermatol. Sci. 2016, 82, 209–212. [Google Scholar] [CrossRef]
- Lin, J.; Selim, M.A.; Shea, C.R.; Grichnik, J.M.; Omar, M.M.; Monteiro-Riviere, N.A.; Pinnell, S.R. UV photoprotection by combination topical antioxidants vitamin C and vitamin E. J. Am. Acad. Dermatol. 2003, 48, 866–874. [Google Scholar] [CrossRef]
- Cao, C.; Lu, S.; Kivlin, R.; Wallin, B.; Card, E.; Bagdasarian, A.; Tamakloe, T.; Wang, W.; Song, X.; Chu, W.; et al. SIRT1 confers protection against UVB- and H2O2-induced cell death via modulation of p53 and JNK in cultured skin keratinocytes. J. Cell Mol. Med. 2009, 13, 3632–3643. [Google Scholar] [CrossRef]
- Chung, S.; Yao, H.; Caito, S.; Hwang, J.; Arunachalam, G.; Rahman, I. Regulation of SIRT1 in cellular functions: Role of polyphenols. Arch. Biochem. Biophys. 2010, 501, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Pambianchi, E.; Ferrara, F.; Pecorelli, A.; Woodby, B.; Grace, M.; Therrien, J.; Lila, M.A.; Valacchi, G. Blueberry extracts as a novel approach to prevent ozone-induced cutaneous inflammasome activation. Oxid. Med. Cell Longev. 2020, 2020, 9571490. [Google Scholar] [CrossRef]
- Roy, S.; Khanna, S.; Alessio, H.M.; Vider, J.; Bagchi, D.; Bagchi, M.; Sen, C.K. Anti-angiogenic property of edible berries. Free Radic. Res. 2002, 36, 1023–1031. [Google Scholar] [CrossRef]
- Pambianchi, E.; Hagenberg, Z.; Pecorelli, A.; Grace, M.; Therrien, J.; Lila, M.A.; Valacchi, G. Alaskan bog blueberry (vaccinium uliginosum) extract as an innovative topical approach to prevent UV-induced skin damage. Cosmetics 2021, 8, 112. [Google Scholar] [CrossRef]
- Yadav, U.C.S.; Ramana, K.V. Regulation of NF-κB-induced inflammatory signaling by lipid peroxidation-derived aldehydes. Oxid. Med. Cell Longev. 2013, 2013, 690545. [Google Scholar] [CrossRef]
- Zhong, J.L.; Edwards, G.P.; Raval, C.; Li, H.; Tyrrell, R.M. The role of Nrf2 in ultraviolet A mediated heme oxygenase 1 induction in human skin fibroblasts. Photochem. Photobiol. Sci. 2010, 9, 18–24. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y.M.; Boguniewicz, M.; Howell, M.D. Loricrin and involucrin expression is down-regulated by Th2 cytokines through STAT-6. Clin. Immunol. 2008, 126, 332–337. [Google Scholar] [CrossRef]
- Sandilands, A.; O’Regan, G.M.; Liao, H.; Zhao, Y.; Terron-Kwiatkowski, A.; Watson, R.M.; Cassidy, A.J.; Goudie, D.R.; Smith, F.J.D.; McLean, W.H.I.; et al. Prevalent and rare mutations in the gene encoding filaggrin cause ichthyosis vulgaris and predispose individuals to atopic dermatitis. J. Investig. Dermatol. 2006, 126, 1770–1775. [Google Scholar] [CrossRef]
- Draelos, Z.D.; Yatskayer, M.; Raab, S.; Oresajo, C. An evaluation of the effect of a topical product containing C-xyloside and blueberry extract on the appearance of type II diabetic skin. J. Cosmet. Dermatol. 2009, 8, 147–151. [Google Scholar] [CrossRef]
- Park, C.H.; Kwak, Y.S.; Seo, H.K.; Kim, H.Y. Assessing the values of blueberries intake on exercise performance, TAS, and inflammatory factors. Iran J. Public Health 2018, 47, 27–32. [Google Scholar]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Nieman, D.C.; Gillitt, N.D.; Chen, G.; Zhang, Q.; Sha, W.; Kay, C.D.; Chandra, P.; Kay, K.L.; Lila, M.A. Blueberry and/or banana consumption mitigate arachidonic, cytochrome P450 oxylipin generation during recovery from 75-Km cycling: A randomized trial. Front. Nutr. 2020, 7, 121. [Google Scholar] [CrossRef]
- Nayeem, M.A. Role of oxylipins in cardiovascular diseases. Acta Pharmacol. Sin. 2018, 39, 1142–1154. [Google Scholar] [CrossRef]
- Martini, D.; Marino, M.; Venturi, S.; Tucci, M.; Klimis-Zacas, D.; Riso, P.; Porrini, M.; Del Bo’, C. Blueberries and their bioactives in the modulation of oxidative stress, inflammation and cardio/vascular function markers: A systematic review of human intervention studies. J. Nutr. Biochem. 2023, 111, 109154. [Google Scholar] [CrossRef]
- Palma, X.; Thomas-Valdés, S.; Cruz, G. Acute consumption of blueberries and short-term blueberry supplementation improve glucose management and insulin levels in sedentary subjects. Nutrients 2021, 13, 1458. [Google Scholar] [CrossRef]
- McLeay, Y.; Barnes, M.J.; Mundel, T.; Hurst, S.M.; Hurst, R.D.; Stannard, S.R. Effect of New Zealand blueberry consumption on recovery from eccentric exercise-induced muscle damage. J. Int. Soc. Sport. Nutr. 2012, 9, 19. [Google Scholar] [CrossRef]
- Hojman, L.; Karsulovic, C. Cardiovascular Disease-Associated Skin Conditions. Vasc. Health Risk Manag. 2022, 18, 43–53. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, J.G.; Jang, Y.A.; Bayazid, A.B.; Ou Lim, B. Fermented black rice and blueberry with Lactobacillus plantarum MG4221 improve UVB-induced skin injury. Food Agric. Immunol. 2021, 32, 499–515. [Google Scholar] [CrossRef]
- Segger, D.; Schönlau, F. Supplementation with Evelle® improves skin smoothness and elasticity in a double-blind, placebo-controlled study with 62 women. J. Dermatol. Treat. 2004, 15, 222–226. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef]
- Jeon, H.Y.; Kim, J.K.; Kim, W.G.; Lee, S.J. Effects of oral epigallocatechin gallate supplementation on the minimal erythema dose and UV-induced skin damage. Skin Pharmacol. Physiol. 2009, 22, 137–141. [Google Scholar] [CrossRef]
- De Pessemier, B.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut-Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef]
- Song, H.; Yoo, Y.; Hwang, J.; Na, Y.; Kim, H.S. Faecalibacterium prausnitzii subspecies–level dysbiosis in the human gut microbiome underlying atopic dermatitis. J. Allergy Clin. Immunol. 2016, 137, 852–860. [Google Scholar] [CrossRef]
- Lee, E.; Lee, S.; Kang, M.; Kim, K.; Won, S.; Kim, B.; Choi, K.Y.; Kim, B.; Cho, H.; Kim, Y.; et al. Clostridia in the gut and onset of atopic dermatitis via eosinophilic inflammation. Ann. Allergy Asthma Immunol. 2016, 117, 91–92.e1. [Google Scholar] [CrossRef]
- Nam, J.; Yun, Y.; Kim, H.; Kim, H.; Jung, H.J.; Chang, Y.; Ryu, S.; Shin, H.; Kim, H.; Kim, W. Rosacea and its association with enteral microbiota in Korean females. Exp. Dermatol. 2018, 27, 37–42. [Google Scholar] [CrossRef]
- Sikora, M.; Stec, A.; Chrabaszcz, M.; Knot, A.; Waskiel-Burnat, A.; Rakowska, A.; Olszewska, M.; Rudnicka, L. Gut Microbiome in Psoriasis: An Updated Review. Pathogens 2020, 9, 463. [Google Scholar] [CrossRef]
- Bosman, E.S.; Albert, A.Y.; Lui, H.; Dutz, J.P.; Vallance, B.A. Skin exposure to narrow band ultraviolet (UVB) light modulates the human intestinal microbiome. Front. Microbiol. 2019, 10, 2410. [Google Scholar] [CrossRef]
- Rodríguez-Daza, M.; Daoust, L.; Boutkrabt, L.; Pilon, G.; Varin, T.; Dudonné, S.; Levy, É.; Marette, A.; Roy, D.; Desjardins, Y. Wild blueberry proanthocyanidins shape distinct gut microbiota profile and influence glucose homeostasis and intestinal phenotypes in high-fat high-sucrose fed mice. Sci. Rep. 2020, 10, 2217. [Google Scholar] [CrossRef]
- Tan, L.; Zhao, S.; Zhu, W.; Wu, L.; Li, J.; Shen, M.; Lei, L.; Chen, X.; Peng, C. The Akkermansia muciniphila is a gut microbiota signature in psoriasis. Exp. Dermatol. 2018, 27, 144–149. [Google Scholar] [CrossRef]
- Thye, A.Y.; Bah, Y.; Law, J.W.; Tan, L.T.; He, Y.; Wong, S.; Thurairajasingam, S.; Chan, K.; Lee, L.; Letchumanan, V. Gut-skin axis: Unravelling the connection between the gut microbiome and psoriasis. Biomedicines 2022, 10, 1037. [Google Scholar] [CrossRef]
- Vendrame, S.; Guglielmetti, S.; Riso, P.; Arioli, S.; Klimis-Zacas, D.; Porrini, M. Six-week consumption of a wild blueberry powder drink increases bifidobacteria in the human gut. J. Agric. Food Chem. 2011, 59, 12815–12820. [Google Scholar] [CrossRef]
- Moludi, J.; Khedmatgozar, H.; Saiedi, S.; Razmi, H.; Alizadeh, M.; Ebrahimi, B. Probiotic supplementation improves clinical outcomes and quality of life indicators in patients with plaque psoriasis: A randomized double-blind clinical trial. Clin. Nutr. ESPEN 2021, 46, 33–39. [Google Scholar] [CrossRef]
- Ntemiri, A.; Ghosh, T.S.; Gheller, M.E.; Tran, T.T.T.; Blum, J.E.; Pellanda, P.; Vlckova, K.; Neto, M.C.; Howell, A.; Thalacker-Mercer, A.; et al. Whole blueberry and isolated polyphenol-rich fractions modulate specific gut microbes in an in vitro colon model and in a pilot study in human consumers. Nutrients 2020, 12, 2800. [Google Scholar] [CrossRef]
- Volkmann, E.R. Intestinal microbiome in scleroderma: Recent progress. Curr. Opin. Rheumatol. 2017, 29, 553–560. [Google Scholar] [CrossRef]
- Catinean, A.; Neag, M.A.; Mitre, A.O.; Bocsan, C.I.; Buzoianu, A.D. Microbiota and immune-mediated skin diseases-an overview. Microorganisms 2019, 7, 279. [Google Scholar] [CrossRef]
- Scher, J.U.; Ubeda, C.; Artacho, A.; Attur, M.; Isaac, S.; Reddy, S.M.; Marmon, S.; Neimann, A.; Brusca, S.; Patel, T.; et al. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis Rheumatol. 2015, 67, 128–139. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.; Pang, D.; Li, T.; Liu, R. Mechanisms underlying the protective effects of blueberry extract against ultraviolet radiation in a skin cell co-culture system. J. Funct. Foods 2019, 52, 603–610. [Google Scholar] [CrossRef]
- Hong, S.; Kang, M.C.; Jin, M.; Lee, T.H.; Lim, B.; Kim, S. Fermented blueberry and black rice containing Lactobacillus plantarum MG4221: A novel functional food for particulate matter (PM 2.5)/dinitrochlorobenzene (DNCB)-induced atopic dermatitis. Food Funct. 2021, 12, 3611–3623. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Krutmann, J.; Wilkens, K.; D’Amato, K. Effect of a blueberry-derived antioxidant matrix on infrared-a induced gene expression in human dermal fibroblasts. J. Drugs Dermatol. 2017, 16, s125–s128. [Google Scholar]
- Bae, J.; Lim, S.S.; Kim, S.J.; Choi, J.; Park, J.; Ju, S.M.; Han, S.J.; Kang, I.; Kang, Y. Bog blueberry anthocyanins alleviate photoaging in ultraviolet-B irradiation-induced human dermal fibroblasts. Mol. Nutr. Food Res. 2009, 53, 726–738. [Google Scholar] [CrossRef]
- Yamasaki, S.; Mizoguchi, K.; Kodama, N.; Iseki, J. Lowbush blueberry extract protects normal human (Homo sapiens L.) epidermal keratinocytes from damage by UV-B irradiation. Eur. Chem. Bull. 2018, 7, 162. [Google Scholar] [CrossRef]
- Schiavon, D.; Martini, D.N.; Brocco, G.; Santos, J.S.; Anzolin, A.P.; Rossato-Grando, L.G.; Omidian, H.; Bertol, C.D. Multifunctional cosmetic containing blueberry and tinosorb M®-loaded microparticles improves sunscreen performance. Adv. Pharm. Bull. 2019, 9, 241–248. [Google Scholar] [CrossRef]

{kind=link}
| Metabolite | Concentration in Plasma (nM) |
|---|---|
| Hippuric acid | 5456 ± 1274 |
| α-Hydroxyhippuric acid | 4322 ± 1607 |
| Benzoic acid | 385 ± 16 |
| Syringic acid | 241 ± 201 |
| 3-and 4-Hydroxyphenylacetic acid | 241 ± 50 327 ± 22 |
| Hydroxycinnamic acids (Ferulic, Dihydroferulic, Isoferulic) | 194 ± 2 256 ± 52 217 ± 44 |
| Authors | Model | Findings |
|---|---|---|
| Roy, S. et al., 2002 [56] | In vitro | Blueberry extract-treated human HaCaT keratinocytes displayed significant reductions in hydrogen peroxide and TNF⍺-induced vascular endothelial growth factor (VEGF) expression. |
| Wang, H. et al., 2019 [90] | In vitro | On human HaCaT keratinocytes and human foreskin fibroblast cells exposed to UV-C radiation, blueberry extract exhibited protective effects: decrease DNA fragmentation, inhibited MMP-1 and other inflammatory factors expression. |
| Hong, S. et al., 2021 [91] | In vitro/In vivo | Fermented blueberry and black rice (FBBBR) extract mitigated particulate matter-induced inflammation in HaCaT cells through proinflammatory cytokine reduction (IL-1β, IL-6, IL-8). FBBBR improved transepidermal water loss, decreased skin erythema, AD-like symptoms, cytokines (IFN-γ, IL-2, IL-4, and IL-10), and scratching tendency in mice exposed to particulate matter. |
| Grether-beck, S, et al., 2017 [92] | In vitro | Blueberry-derived antioxidant matrix (Berrimatrix™) successfully prevented infrared A-induced MMP-1 expression in primary human skin fibroblasts. |
| Bae, J. et al., 2009 [93] | In vitro | Bog blueberry extract attenuated UV-B triggered collagen damage and release of IL-6 and IL-8 in human dermal fibroblasts. |
| Yamasaki, S. et al., 2018 [94] | In vitro | Normal human epidermal keratinocytes pretreated with lowbush blueberry extract exhibited significant attenuation in UV-B-induced damage. |
| Park, S. et al., 2021 [71] | In vivo | In UVB-irradiated hairless mice, FBBBR oral supplementation improved stratum corneum hydration, epidermal thickness, and increased filaggrin, involucrin, TGM-1, and COL1A1 mRNA expression in skin. |
| Pambianchi, E. et al., 2020 [55] | In vitro/Ex vivo | Pretreatment with blueberry extract displayed protective effects in HaCaT, reconstructed human epidermis, and human skin explants exposed to ozone. Blueberry extract enhanced keratinocyte wound closure and prevented O3-induced ROS production and inflammasome activation (reduction of CASP1, IL-18). Blueberry extract also prevented O3-induced increase in 4-HNE, ASC, and NLRP1 in human skin explants. |
| Pambianchi, E. et al., 2021 [57] | Ex vivo | Topical application of Alaskan bog blueberry extract prevented UVA/UVB-induced 4-HNE, HO-1, COX2 expression and decreased the loss of cutaneous structural proteins (filaggrin and involucrin) in human skin explants. |
| Draelos, Z.D. et al., 2009 [62] | Clinical | /Topical application of blueberry extract-containing product for 12 weeks increased cutaneous moisturization, smoothness, radiance, and firmness in type II diabetic skin. |
| Segger, D. et al., 2004 [72] | Clinical | Dietary supplementation of Evelle (a blueberry extract-containing product) for 12 weeks successfully improved skin elasticity and reduced skin roughness. |
| Schiavon, D. et al., 2019 [95] | Clinical | Blueberry extract- and microparticles-formulated sunscreen exhibited increased photoprotective capacity in the stratum corneum of humans exposed to UV light. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivarsson, J., Jr.; Pecorelli, A.; Lila, M.A.; Valacchi, G. Blueberry Supplementation and Skin Health. Antioxidants 2023, 12, 1261. https://doi.org/10.3390/antiox12061261
Ivarsson J Jr., Pecorelli A, Lila MA, Valacchi G. Blueberry Supplementation and Skin Health. Antioxidants. 2023; 12(6):1261. https://doi.org/10.3390/antiox12061261
Chicago/Turabian StyleIvarsson, John, Jr., Alessandra Pecorelli, Mary Ann Lila, and Giuseppe Valacchi. 2023. "Blueberry Supplementation and Skin Health" Antioxidants 12, no. 6: 1261. https://doi.org/10.3390/antiox12061261
APA StyleIvarsson, J., Jr., Pecorelli, A., Lila, M. A., & Valacchi, G. (2023). Blueberry Supplementation and Skin Health. Antioxidants, 12(6), 1261. https://doi.org/10.3390/antiox12061261