
Antioxidant and Immune-Related Implications of Minerals in COVID-19: A Possibility for Disease Prevention and Management


 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. The Link between Immunity, Oxidative Stress, and COVID-19
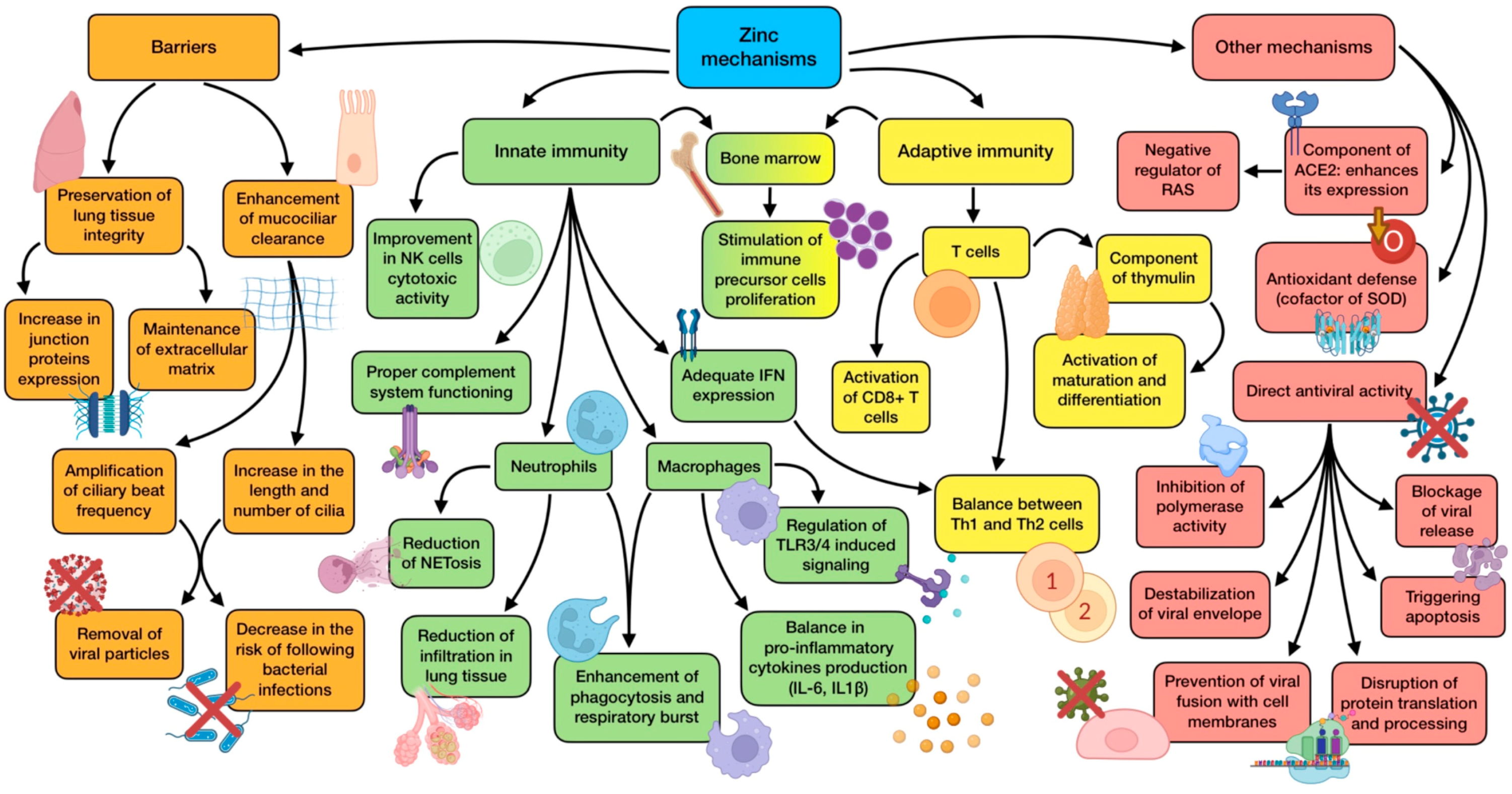
3.2. The Role of Zinc (Zn)
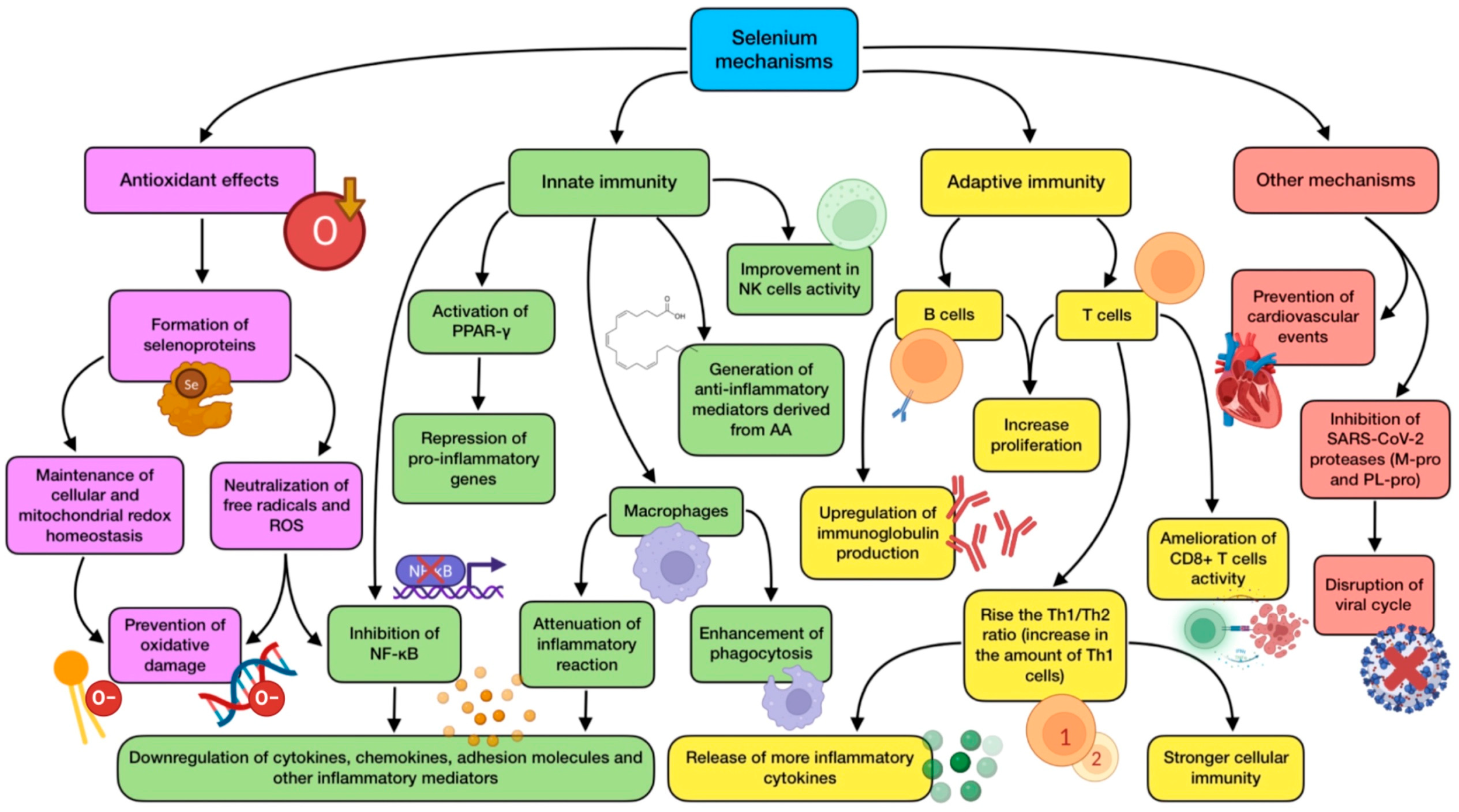
3.3. The Role of Selenium (Se)
- Antioxidant activity: the majority of selenoproteins have a role in the mitigation of oxidative stress, being able to neutralize free radicals triggered by infections, immune response, and inflammation [99].
- Maintenance of cellular redox homeostasis: TXNRDs and GPXs are required for preserving mitochondrial integrity and redox tone in immune cells, reversing oxidative damage inflicted on them (in part due to its own oxidative burst) [100].
- Anti-inflammatory effects: selenoproteins take part in the generation of anti-inflammatory mediators derived from arachidonic acid (AA), which protect cells against the expression of pro-inflammatory genes. They can also inhibit NF-κB, thus down-regulating inflammatory genes. In addition, these proteins activate the peroxisome proliferator-activated nuclear receptor-γ (PPAR-γ), which leads to a repression of pro-inflammatory genes [101,102].
- Immunomodulatory effects: these types of proteins can modify immunity through an up-regulation of IL-2 receptor’s expression. Therefore, T and B cells can respond easily to this cytokine, increasing their proliferation and activation. Moreover, selenoproteins P, W, and K modify circulating levels of some cytokines, such as IL-6, TNF-α, and IL-1β [102].
- Other functions: TXNRD1 is necessary for DNA synthesis during T cell proliferation. GPX1 activity is related to a reduction in cardiovascular events in patients suffering from coronary disease, and TXNRD2 has been proven to preserve heart functioning in the elderly [103].
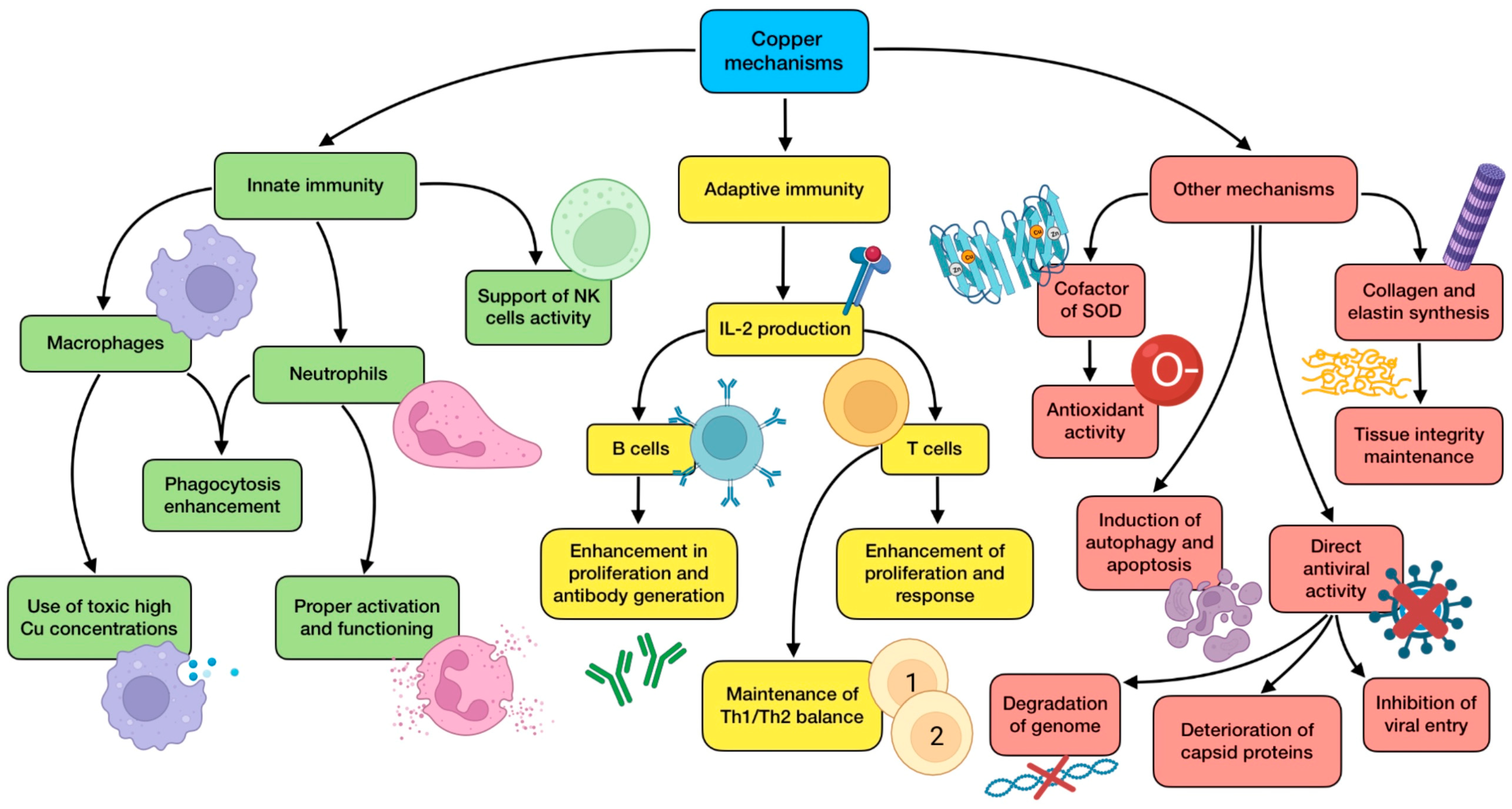
3.4. The Role of Copper (Cu)
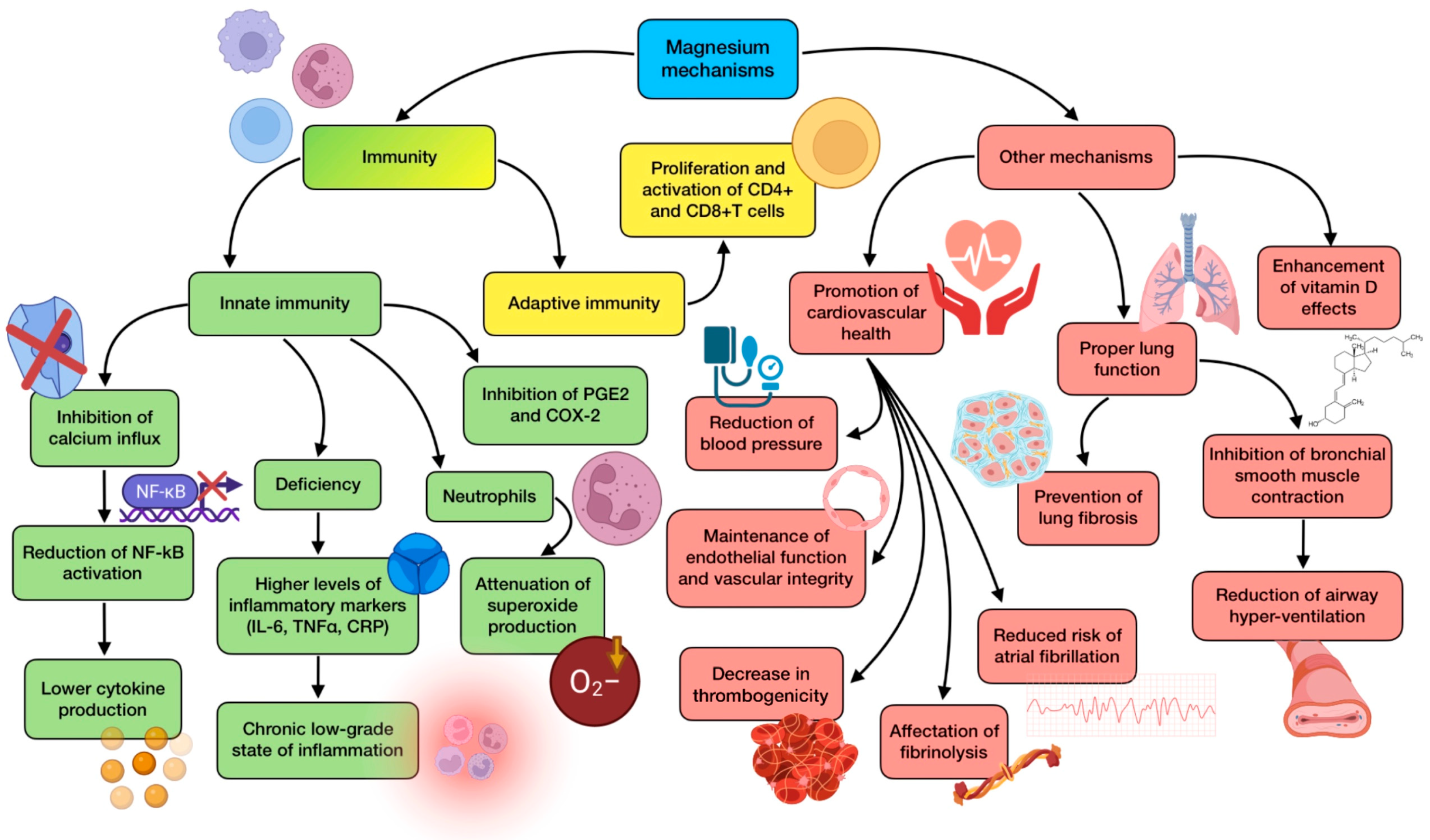
3.5. The Role of Magnesium (Mg)
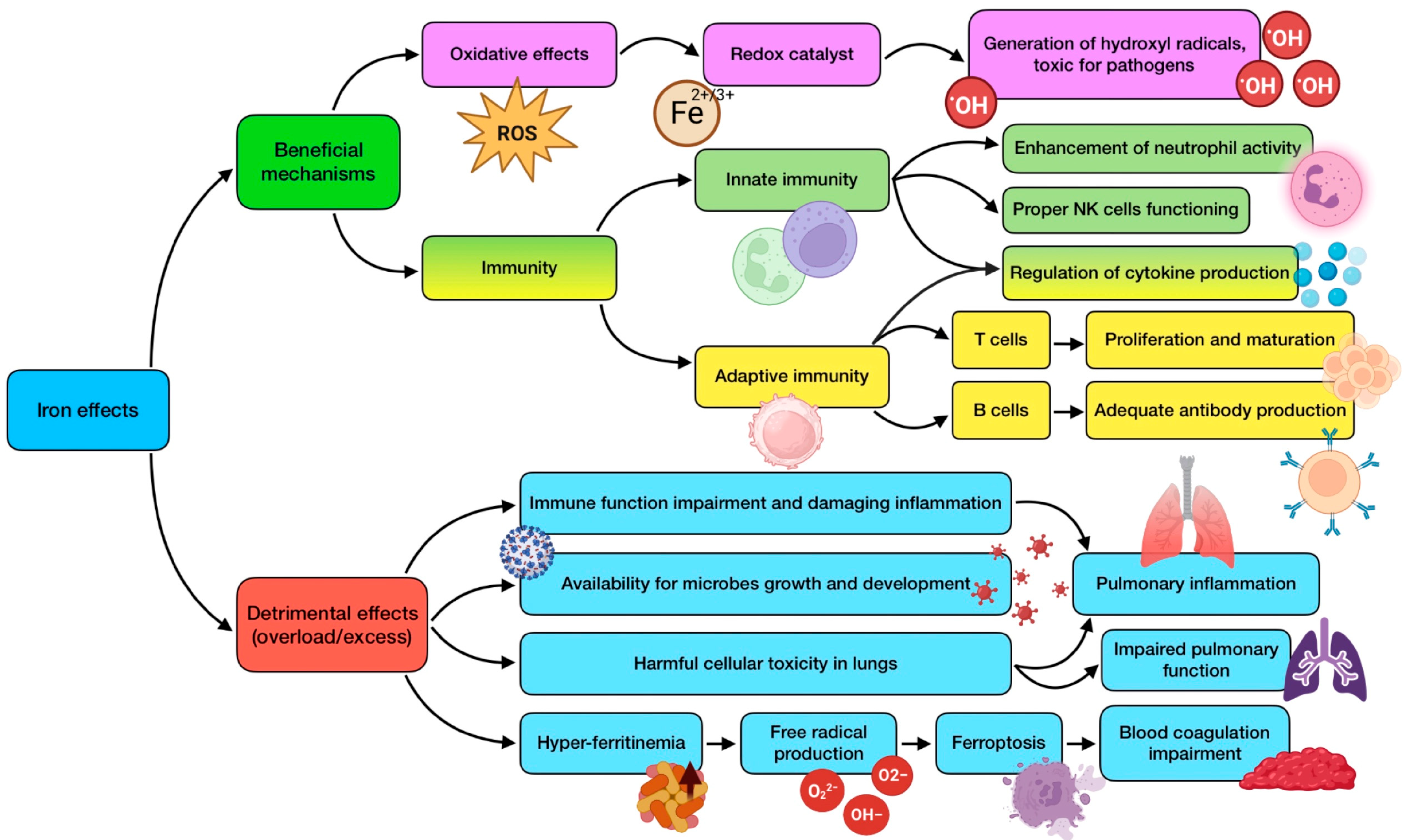
3.6. The Role of Iron (Fe)
3.7. The Role of Other Minerals
3.8. Limitations of the Study
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rothan, H.A.; Byrareddy, S.N. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Ye, D.; Liu, Q. Review of the 2019 novel coronavirus (SARS-CoV-2) based on current evidence. Int. J. Antimicrob. Agents 2020, 55, 105948. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.C.; Shih, T.P.; Ko, W.C.; Tang, H.J.; Hsueh, P.R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor Recognition by the Novel Coronavirus from Wuhan: An Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J. Virol. 2020, 94, e00127-20. [Google Scholar] [CrossRef]
- Singhal, T. A Review of Coronavirus Disease-2019 (COVID-19). Indian J. Pediatr. 2020, 87, 281–286. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Shao, C.; Huang, J.; Gan, J.; Huang, X.; Bucci, E.; Piacentini, M.; Ippolito, G.; Melino, G. COVID-19 infection: The perspectives on immune responses. Cell Death Differ. 2020, 27, 1451–1454. [Google Scholar] [CrossRef]
- Gombart, A.F.; Pierre, A.; Maggini, S. A Review of Micronutrients and the Immune System-Working in Harmony to Reduce the Risk of Infection. Nutrients 2020, 12, 236. [Google Scholar] [CrossRef]
- Žarković, N.; Orehovec, B.; Milković, L.; Baršić, B.; Tatzber, F.; Wonisch, W.; Tarle, M.; Kmet, M.; Mataić, A.; Jakovčević, A.; et al. Preliminary Findings on the Association of the Lipid Peroxidation Product 4-Hydroxynonenal with the Lethal Outcome of Aggressive COVID-19. Antioxidants 2021, 10, 1341. [Google Scholar] [CrossRef]
- Wessels, I.; Pupke, J.T.; von Trotha, K.T.; Gombert, A.; Himmelsbach, A.; Fischer, H.J.; Jacobs, M.J.; Rink, L.; Grommes, J. Zinc supplementation ameliorates lung injury by reducing neutrophil recruitment and activity. Thorax 2020, 75, 253–261. [Google Scholar] [CrossRef]
- Jothimani, D.; Kailasam, E.; Danielraj, S.; Nallathambi, B.; Ramachandran, H.; Sekar, P.; Manoharan, S.; Ramani, V.; Narasimhan, G.; Kaliamoorthy, I.; et al. COVID-19: Poor outcomes in patients with zinc deficiency. Int. J. Infect. Dis. 2020, 100, 343–349. [Google Scholar] [CrossRef]
- Te Velthuis, A.J.; van den Worm, S.H.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn(2+) inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef]
- Roscioli, E.; Jersmann, H.P.; Lester, S.; Badiei, A.; Fon, A.; Zalewski, P.; Hodge, S. Zinc deficiency as a codeterminant for airway epithelial barrier dysfunction in an ex vivo model of COPD. Int. J. Chronic Obstruct. Pulmon. Dis. 2017, 12, 3503–3510. [Google Scholar] [CrossRef]
- Hemilä, H. Zinc lozenges and the common cold: A meta-analysis comparing zinc acetate and zinc gluconate, and the role of zinc dosage. JRSM Open. 2017, 8, 2054270417694291. [Google Scholar] [CrossRef]
- Science, M.; Johnstone, J.; Roth, D.E.; Guyatt, G.; Loeb, M. Zinc for the treatment of the common cold: A systematic review and meta-analysis of randomized controlled trials. Cmaj 2012, 184, E551–E561. [Google Scholar] [CrossRef]
- Mao, S.; Zhang, A.; Huang, S. Meta-analysis of Zn, Cu and Fe in the hair of Chinese children with recurrent respiratory tract infection. Scand. J. Clin. Lab. Investig. 2014, 74, 561–567. [Google Scholar] [CrossRef]
- Anuk, A.T.; Polat, N.; Akdas, S.; Erol, S.A.; Tanacan, A.; Biriken, D.; Keskin, H.L.; Tekin, O.M.; Yazihan, N.; Sahin, D. The Relation Between Trace Element Status (Zinc, Copper, Magnesium) and Clinical Outcomes in COVID-19 Infection During Pregnancy. Biol. Trace Elem. Res. 2021, 199, 3608–3617. [Google Scholar] [CrossRef]
- Heller, R.A.; Sun, Q.; Hackler, J.; Seelig, J.; Seibert, L.; Cherkezov, A.; Minich, W.B.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Prediction of survival odds in COVID-19 by zinc, age and selenoprotein P as composite biomarker. Redox Biol. 2021, 38, 101764. [Google Scholar] [CrossRef]
- Ben Abdallah, S.; Mhalla, Y.; Trabelsi, I.; Sekma, A.; Youssef, R.; Ali, K.B.H.; Soltane, H.B.; Yacoubi, H.; Msolli, M.A.; Stambouli, N.; et al. Twice-Daily Oral Zinc in the Treatment of Patients With Coronavirus Disease 2019: A Randomized Double-Blind Controlled Trial. Clin. Infect. Dis. 2023, 76, 185–191. [Google Scholar] [CrossRef]
- Reino-Gelardo, S.; Palop-Cervera, M.; Aparisi-Valero, N.; Miguel, I.E.-S.; Lozano-Rodríguez, N.; Llop-Furquet, G.; Sanchis-Artero, L.; Cortés-Castell, E.; Rizo-Baeza, M.; Cortés-Rizo, X. Effect of an Immune-Boosting, Antioxidant and Anti-Inflammatory Food Supplement in Hospitalized COVID-19 Patients: A Prospective Randomized Pilot Study. Nutrients 2023, 15, 1736. [Google Scholar] [CrossRef]
- Natarajan, S.; Anbarasi, C.; Sathiyarajeswaran, P.; Manickam, P.; Geetha, S.; Kathiravan, R.; Prathiba, P.; Pitchiahkumar, M.; Parthiban, P.; Kanakavalli, K.; et al. Kabasura Kudineer (KSK), a poly-herbal Siddha medicine, reduced SARS-CoV-2 viral load in asymptomatic COVID-19 individuals as compared to vitamin C and zinc supplementation: Findings from a prospective, exploratory, open-labeled, comparative, randomized controlled trial, Tamil Nadu, India. Trials 2021, 22, 623. [Google Scholar] [PubMed]
- Finzi, E. Treatment of SARS-CoV-2 with high dose oral zinc salts: A report on four patients. Int. J. Infect. Dis. 2020, 99, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Patel, O.; Chinni, V.; El-Khoury, J.; Perera, M.; Neto, A.S.; McDonald, C.; See, E.; Jones, D.; Bolton, D.; Bellomo, R.; et al. A pilot double-blind safety and feasibility randomized controlled trial of high-dose intravenous zinc in hospitalized COVID-19 patients. J. Med. Virol. 2021, 93, 3261–3267. [Google Scholar] [CrossRef] [PubMed]
- Abdelmaksoud, A.A.; Ghweil, A.A.; Hassan, M.H.; Rashad, A.; Khodeary, A.; Aref, Z.F.; Sayed, M.A.A.; Elsamman, M.K.; Bazeed, S.E.S. Olfactory Disturbances as Presenting Manifestation Among Egyptian Patients with COVID-19: Possible Role of Zinc. Biol. Trace Element Res. 2021, 199, 4101–4108. [Google Scholar] [CrossRef]
- Stambouli, N.; Driss, A.; Gargouri, F.; Bahrini, K.; Arfaoui, B.; Abid, R.; Taamallah, K.; Hannachi, S.; Boughariou, S.; Rebai, A.; et al. COVID-19 prophylaxis with doxycycline and zinc in health care workers: A prospective, randomized, double-blind clinical trial. Int. J. Infect. Dis. 2022, 122, 553–558. [Google Scholar] [CrossRef]
- Rodriguez, J.A.M.; Bifano, M.; Goma, E.R.; Plasencia, C.M.; Torralba, A.O.; Font, M.S.; Millán, P.R. Effect and Tolerability of a Nutritional Supplement Based on a Synergistic Combination of β-Glucans and Selenium- and Zinc-Enriched Saccharomyces cerevisiae (ABB C1(®)) in Volunteers Receiving the Influenza or the COVID-19 Vaccine: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2021, 13, 4347. [Google Scholar]
- Thomas, S.; Patel, D.; Bittel, B.; Wolski, K.; Wang, Q.; Kumar, A.; Il, Z.J.; Mehra, R.; McWilliams, C.; Nissen, S.E.; et al. Effect of High-Dose Zinc and Ascorbic Acid Supplementation vs Usual Care on Symptom Length and Reduction Among Ambulatory Patients With SARS-CoV-2 Infection: The COVID A to Z Randomized Clinical Trial. JAMA Netw. Open 2021, 4, e210369. [Google Scholar] [CrossRef]
- Abd-Elsalam, S.; Soliman, S.; Esmail, E.S.; Khalaf, M.; Mostafa, E.F.; Medhat, M.A.; Ahmed, O.A.; El Ghafar, M.S.A.; Alboraie, M.; Hassany, S.M. Do Zinc Supplements Enhance the Clinical Efficacy of Hydroxychloroquine?: A Randomized, Multicenter Trial. Biol. Trace Elem. Res. 2021, 199, 3642–3646. [Google Scholar] [CrossRef]
- Maywald, M.; Rink, L. Zinc supplementation induces CD4(+)CD25(+)Foxp3(+) antigen-specific regulatory T cells and suppresses IFN-γ production by upregulation of Foxp3 and KLF-10 and downregulation of IRF-1. Eur. J. Nutr. 2017, 56, 1859–1869. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.; Sun, Y.; Stubbs, D.; He, J.; Li, W.; Wang, F.; Liu, Z.; Ruzicka, J.A.; Taylor, E.W.; et al. SARS-CoV-2 suppresses mRNA expression of selenoproteins associated with ferroptosis, endoplasmic reticulum stress and DNA synthesis. Food Chem. Toxicol. 2021, 153, 112286. [Google Scholar] [CrossRef]
- Lee, Y.H.; Lee, S.J.; Lee, M.K.; Lee, W.Y.; Yong, S.J.; Kim, S.H. Serum selenium levels in patients with respiratory diseases: A prospective observational study. J. Thorac. Dis. 2016, 8, 2068–2078. [Google Scholar] [CrossRef]
- Tseng, C.K.; Ho, C.T.; Hsu, H.S.; Lin, C.H.; Li, C.I.; Li, T.C.; Liu, C.S.; Lin, C.C.; Lin, W.Y. Selenium is inversely associated with interleukin-6 in the elderly. J. Nutr. Health Aging 2013, 17, 280–284. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of M(pro) from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef]
- Mahmoodpoor, A.; Hamishehkar, H.; Shadvar, K.; Ostadi, Z.; Sanaie, S.; Saghaleini, S.H.; Nader, N.D. The Effect of Intravenous Selenium on Oxidative Stress in Critically Ill Patients with Acute Respiratory Distress Syndrome. Immunol. Investig. 2019, 48, 147–159. [Google Scholar] [CrossRef]
- Zhang, J.; Taylor, E.W.; Bennett, K.; Saad, R.; Rayman, M.P. Association between regional selenium status and reported outcome of COVID-19 cases in China. Am. J. Clin. Nutr. 2020, 111, 1297–1299. [Google Scholar] [CrossRef]
- Moghaddam, A.; Heller, R.A.; Sun, Q.; Seelig, J.; Cherkezov, A.; Seibert, L.; Hackler, J.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Selenium Deficiency Is Associated with Mortality Risk from COVID-19. Nutrients 2020, 12, 2098. [Google Scholar] [CrossRef]
- Warnes, S.L.; Little, Z.R.; Keevil, C.W. Human Coronavirus 229E Remains Infectious on Common Touch Surface Materials. mBio 2015, 6, e01697-15. [Google Scholar] [CrossRef]
- Wazir, S.M.; Ghobrial, I. Copper deficiency, a new triad: Anemia, leucopenia, and myeloneuropathy. J. Community Hosp. Intern. Med. Perspect. 2017, 74, 265–268. [Google Scholar] [CrossRef]
- Lee, Y.H.; Bang, E.S.; Lee, J.H.; Lee, J.D.; Kang, D.R.; Hong, J.; Lee, J.M. Serum Concentrations of Trace Elements Zinc, Copper, Selenium, and Manganese in Critically Ill Patients. Biol. Trace Elem. Res. 2019, 188, 316–325. [Google Scholar] [CrossRef]
- Hackler, J.; Heller, R.A.; Sun, Q.; Schwarzer, M.; Diegmann, J.; Bachmann, M.; Moghaddam, A.; Schomburg, L. Relation of Serum Copper Status to Survival in COVID-19. Nutrients. 2021, 13, 1898. [Google Scholar] [CrossRef] [PubMed]
- Kanellopoulou, C.; AGeorge, B.; Masutani, E.; Cannons, J.L.; Ravell, J.C.; Yamamoto, T.N.; Smelkinson, M.G.; Jiang, P.D.; Matsuda-Lennikov, M.; Reilley, J.; et al. Mg(2+) regulation of kinase signaling and immune function. J. Exp. Med. 2019, 216, 1828–1842. [Google Scholar] [CrossRef] [PubMed]
- Afshar Ebrahimi, F.; Foroozanfard, F.; Aghadavod, E.; Bahmani, F.; Asemi, Z. The Effects of Magnesium and Zinc Co-Supplementation on Biomarkers of Inflammation and Oxidative Stress, and Gene Expression Related to Inflammation in Polycystic Ovary Syndrome: A Randomized Controlled Clinical Trial. Biol. Trace Elem. Res. 2018, 184, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhang, P.; Liu, T.; Zhang, X.; Pan, X.; Cen, Y.; Liu, Y.; Zhang, H.; Chen, X. Magnesium isoglycyrrhizinate ameliorates radiation-induced pulmonary fibrosis by inhibiting fibroblast differentiation via the p38MAPK/Akt/Nox4 pathway. Biomed. Pharmacother. 2019, 115, 108955. [Google Scholar] [CrossRef] [PubMed]
- Nasser, R.; Naffaa, M.E.; Mashiach, T.; Azzam, Z.S.; Braun, E. The association between serum magnesium levels and community-acquired pneumonia 30-day mortality. BMC Infect. Dis. 2018, 18, 698. [Google Scholar] [CrossRef]
- Quilliot, D.; Bonsack, O.; Jaussaud, R.; Mazur, A. Dysmagnesemia in Covid-19 cohort patients: Prevalence and associated factors. Magnes. Res. 2020, 33, 114–122. [Google Scholar] [CrossRef]
- Tan, C.W.; Ho, L.P.; Kalimuddin, S.; Cherng, B.P.Z.; Teh, Y.E.; Thien, S.Y.; Wong, H.M.; Tern, P.J.W.; Chandran, M.; Chay, J.W.M.; et al. Cohort study to evaluate the effect of vitamin D, magnesium, and vitamin B(12) in combination on progression to severe outcomes in older patients with coronavirus (COVID-19). Nutrition 2020, 79, 111017. [Google Scholar] [CrossRef]
- Fan, L.; Zhu, X.; Zheng, Y.; Zhang, W.; Seidner, D.L.; Ness, R.; Murff, H.J.; Yu, C.; Huang, X.; Shrubsole, M.J.; et al. Magnesium treatment on methylation changes of transmembrane serine protease 2 (TMPRSS2). Nutrition 2021, 89, 111340. [Google Scholar] [CrossRef]
- Agoro, R.; Taleb, M.; Quesniaux, V.F.J.; Mura, C. Cell iron status influences macrophage polarization. PLoS ONE 2018, 13, e0196921. [Google Scholar] [CrossRef]
- Jayaweera, J.; Reyes, M.; Joseph, A. Childhood iron deficiency anemia leads to recurrent respiratory tract infections and gastroenteritis. Sci. Rep. 2019, 9, 12637. [Google Scholar] [CrossRef]
- Ponikowski, P.; BKirwan, A.; Anker, S.D.; McDonagh, T.; Dorobantu, M.; Drozdz, J.; Fabien, V.; Filippatos, G.; Göhring, U.M.; Keren, A.; et al. Ferric carboxymaltose for iron deficiency at discharge after acute heart failure: A multicentre, double-blind, randomised, controlled trial. Lancet 2020, 396, 1895–1904. [Google Scholar] [CrossRef]
- Vinke, J.S.J.; Altulea, D.H.A.; Eisenga, M.F.; Jagersma, R.L.; Niekolaas, T.M.; van Baarle, D.; Heiden, M.V.; Steenhuis, M.; Rispens, T.; Abdulahad, W.H.; et al. Ferric carboxymaltose and SARS-CoV-2 vaccination-induced immunogenicity in kidney transplant recipients with iron deficiency: The COVAC-EFFECT randomized controlled trial. Front. Immunol. 2022, 13, 1017178. [Google Scholar] [CrossRef]
- Bolondi, G.; Russo, E.; Gamberini, E.; Circelli, A.; Meca, M.C.C.; Brogi, E.; Viola, L.; Bissoni, L.; Poletti, V.; Agnoletti, V. Iron metabolism and lymphocyte characterisation during Covid-19 infection in ICU patients: An observational cohort study. World J. Emerg. Surg. 2020, 15, 41. [Google Scholar] [CrossRef]
- Zhao, K.; Huang, J.; Dai, D.; Feng, Y.; Liu, L.; Nie, S. Serum Iron Level as a Potential Predictor of Coronavirus Disease 2019 Severity and Mortality: A Retrospective Study. Open Forum Infect. Dis. 2020, 7, ofaa250. [Google Scholar] [CrossRef]
- Dinc, M.E.; Dalgic, A.; Ulusoy, S.; Dizdar, D.; Develioglu, O.; Topak, M. Does iron deficiency anemia affect olfactory function? Acta Otolaryngol. 2016, 136, 754–757. [Google Scholar] [CrossRef]
- Ali, M.K.; Kim, R.Y.; Brown, A.C.; Donovan, C.; Vanka, K.S.; Mayall, J.R.; Liu, G.; Pillar, A.L.; Jones-Freeman, B.; Xenaki, D.; et al. Critical role for iron accumulation in the pathogenesis of fibrotic lung disease. J. Pathol. 2020, 251, 49–62. [Google Scholar] [CrossRef]
- Lippi, G.; South, A.M.; Henry, B.M. Electrolyte imbalances in patients with severe coronavirus disease 2019 (COVID-19). Ann. Clin. Biochem. 2020, 57, 62–265. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; Cardona-Ospina, J.A.; Gutiérrez-Ocampo, E.; Villamizar-Peña, R.; Holguin-Rivera, Y.; Escalera-Antezana, J.P.; Alvarado-Arnez, L.E.; Bonilla-Aldana, D.K.; Franco-Paredes, C.; Henao-Martinez, A.F.; et al. Clinical, laboratory and imaging features of COVID-19: A systematic review and meta-analysis. Travel. Med. Infect. Dis. 2020, 34, 101623. [Google Scholar] [CrossRef]
- Sun, Y.; Yin, Y.; Gong, L.; Liang, Z.; Zhu, C.; Ren, C.; Zheng, N.; Zhang, Q.; Liu, H.; Liu, W.; et al. Manganese nanodepot augments host immune response against coronavirus. Nano Res. 2021, 14, 1260–1272. [Google Scholar] [CrossRef]
- Lauridsen, C. From oxidative stress to inflammation: Redox balance and immune system. Poult. Sci. 2019, 98, 4240–4246. [Google Scholar] [CrossRef]
- Okba, N.M.A.; Müller, M.A.; Li, W.; Wang, C.; GeurtsvanKessel, C.H.; Corman, V.M.; Lamers, M.M.; Sikkema, R.S.; de Bruin, E.; Chandler, F.D.; et al. Severe Acute Respiratory Syndrome Coronavirus 2-Specific Antibody Responses in Coronavirus Disease Patients. Emerg. Infect. Dis. 2020, 26, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Paces, J.; Strizova, Z.; Smrz, D.; Cerny, J. COVID-19 and the immune system. Physiol. Res. 2020, 69, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Siu, K.L.; Kok, K.H.; Ng, M.J.; Poon, V.K.M.; Yuen, K.Y.; Zheng, B.J.; Jin, D.Y. Severe acute respiratory syndrome coronavirus M protein inhibits type I interferon production by impeding the formation of TRAF3.TANK.TBK1/IKKepsilon complex. J. Biol. Chem. 2009, 284, 16202–16209. [Google Scholar] [CrossRef] [PubMed]
- Iddir, M.; Brito, A.; Dingeo, G.; Del Campo, S.S.F.; Samouda, H.; La Frano, M.R.; Bohn, T. Strengthening the Immune System and Reducing Inflammation and Oxidative Stress through Diet and Nutrition: Considerations during the COVID-19 Crisis. Nutrients 2020, 12, 1562. [Google Scholar] [CrossRef]
- Libby, P. Inflammatory mechanisms: The molecular basis of inflammation and disease. Nutr. Rev. 2007, 65 Pt 2, S140–S146. [Google Scholar] [CrossRef]
- Fu, Y.; Cheng, Y.; Wu, Y. Understanding SARS-CoV-2-Mediated Inflammatory Responses: From Mechanisms to Potential Therapeutic Tools. Virol. Sin. 2020, 35, 266–271. [Google Scholar] [CrossRef]
- McGonagle, D.; Sharif, K.; O’Regan, A.; Bridgewood, C. The Role of Cytokines including Interleukin-6 in COVID-19 induced Pneumonia and Macrophage Activation Syndrome-Like Disease. Autoimmun. Rev. 2020, 19, 102537. [Google Scholar] [CrossRef]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Xu, Y.; Tian, Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell Mol. Immunol. 2020, 17, 533–535. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, Y.; Zhang, F.; Wang, Q.; Li, T.; Liu, Z.; Wang, J.; Qin, Y.; Zhang, X.; Yan, X.; et al. The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The Perspectives of clinical immunologists from China. Clin. Immunol. 2020, 214, 108393. [Google Scholar] [CrossRef]
- Lopresti, A.L.; Maker, G.L.; Hood, S.D.; Drummond, P.D. A review of peripheral biomarkers in major depression: The potential of inflammatory and oxidative stress biomarkers. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2014, 48, 102–111. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Pohanka, M. Role of oxidative stress in infectious diseases. A review. Folia Microbiol. 2013, 58, 503–513. [Google Scholar] [CrossRef]
- Laforge, M.; Elbim, C.; Frère, C.; Hémadi, M.; Massaad, C.; Nuss, P.; Benoliel, J.J.; Becker, C. Tissue damage from neutrophil-induced oxidative stress in COVID-19. Nat. Rev. Immunol. 2020, 20, 515–516. [Google Scholar] [CrossRef]
- Chams, N.; Chams, S.; Badran, R.; Shams, A.; Araji, A.; Raad, M.; Mukhopadhyay, S.; Stroberg, E.; Duval, E.J.; Barton, L.M.; et al. COVID-19: A Multidisciplinary Review. Front. Public Health 2020, 8, 383. [Google Scholar] [CrossRef]
- Langley-Evans, S.C.; Carrington, L.J. Diet and the developing immune system. Lupus 2006, 15, 746–752. [Google Scholar] [CrossRef]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting Enzyme 2: SARS-CoV-2 Receptor and Regulator of the Renin-Angiotensin System: Celebrating the 20th Anniversary of the Discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef]
- Skalny, A.V.; Rink, L.; Ajsuvakova, O.P.; Aschner, M.; Gritsenko, V.A.; Alekseenko, S.I.; Svistunov, A.A.; Petrakis, D.; Spandidos, D.A.; Aaseth, J.; et al. Zinc and respiratory tract infections: Perspectives for COVID-19 (Review). Int. J. Mol. Med. 2020, 46, 17–26. [Google Scholar] [CrossRef]
- Pal, A.; Squitti, R.; Picozza, M.; Pawar, A.; Rongioletti, M.; Dutta, A.K.; Sahoo, S.; Goswami, K.; Sharma, P.; Prasad, R. Zinc and COVID-19: Basis of Current Clinical Trials. Biol. Trace Elem. Res. 2021, 199, 2882–2892. [Google Scholar] [CrossRef]
- Gammoh, N.Z.; Rink, L. Zinc in Infection and Inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef]
- Wintergerst, E.S.; Maggini, S.; Hornig, D.H. Immune-enhancing role of vitamin C and zinc and effect on clinical conditions. Ann. Nutr. Metab. 2006, 50, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Nutrition, immunity and COVID-19. BMJ Nutr. Prev. Health 2020, 3, 74–92. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.I.; Ledford, J.R.; Zhou, P.; Page, K. Zinc supplementation alters airway inflammation and airway hyperresponsiveness to a common allergen. J. Inflamm. 2011, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.; Rink, L.; Haase, H. Zinc signals in neutrophil granulocytes are required for the formation of neutrophil extracellular traps. Innate Immun. 2013, 19, 253–264. [Google Scholar] [CrossRef]
- Jovic, T.H.; Ali, S.R.; Ibrahim, N.; Jessop, Z.M.; Tarassoli, S.P.; Dobbs, T.D.; Holford, P.; Thornton, C.A.; Whitaker, I.S. Could Vitamins Help in the Fight Against COVID-19? Nutrients 2020, 12, 2550. [Google Scholar] [CrossRef]
- Choudhury, A.; Mukherjee, S. In silico studies on the comparative characterization of the interactions of SARS-CoV-2 spike glycoprotein with ACE-2 receptor homologs and human TLRs. J. Med. Virol. 2020, 92, 2105–2113. [Google Scholar] [CrossRef]
- Wong, C.P.; Rinaldi, N.A.; Ho, E. Zinc deficiency enhanced inflammatory response by increasing immune cell activation and inducing IL6 promoter demethylation. Mol. Nutr. Food Res. 2015, 59, 991–999. [Google Scholar] [CrossRef]
- Berg, K.; Bolt, G.; Andersen, H.; Owen, T.C. Zinc potentiates the antiviral action of human IFN-alpha tenfold. J. Interferon Cytokine Res. 2001, 21, 471–474. [Google Scholar] [CrossRef]
- Wessels, I.; Rolles, B.; Rink, L. The Potential Impact of Zinc Supplementation on COVID-19 Pathogenesis. Front. Immunol. 2020, 11, 1712. [Google Scholar] [CrossRef]
- De Almeida Brasiel, P.G. The key role of zinc in elderly immunity: A possible approach in the COVID-19 crisis. Clin. Nutr. ESPEN 2020, 38, 65–66. [Google Scholar] [CrossRef]
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The Role of Zinc in Antiviral Immunity. Adv. Nutr. 2019, 10, 696–710. [Google Scholar] [CrossRef]
- Subramanian Vignesh, K.; Deepe, G.S., Jr. Metallothioneins: Emerging Modulators in Immunity and Infection. Int. J. Mol. Sci. 2017, 18, 2197. [Google Scholar] [CrossRef]
- Woodworth, B.A.; Zhang, S.; Tamashiro, E.; Bhargave, G.; Palmer, J.N.; Cohen, N.A. Zinc increases ciliary beat frequency in a calcium-dependent manner. Am. J. Rhinol. Allergy 2010, 24, 6–10. [Google Scholar] [CrossRef]
- The World Health report 2002. Midwifery 2003, 19, 72–73.
- Mossad, S.B.; Macknin, M.L.; Medendorp, S.V.; Mason, P. Zinc gluconate lozenges for treating the common cold. A randomized, double-blind, placebo-controlled study. Ann. Intern. Med. 1996, 125, 81–88. [Google Scholar] [CrossRef]
- Barnett, J.B.; Hamer, D.H.; Meydani, S.N. Low zinc status: A new risk factor for pneumonia in the elderly? Nutr. Rev. 2010, 68, 30–37. [Google Scholar] [CrossRef]
- Razzaque, M.S. COVID-19 Pandemic: Can Maintaining Optimal Zinc Balance Enhance Host Resistance? Tohoku J. Exp. Med. 2020, 251, 175–181. [Google Scholar] [CrossRef]
- Galmés, S.; Serra, F.; Palou, A. Current State of Evidence: Influence of Nutritional and Nutrigenetic Factors on Immunity in the COVID-19 Pandemic Framework. Nutrients 2020, 12, 2738. [Google Scholar] [CrossRef]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-Dependent Antioxidant Enzymes: Actions and Properties of Selenoproteins. Antioxidants 2018, 7, 667. [Google Scholar] [CrossRef]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef]
- Qian, F.; Misra, S.; Prabhu, K.S. Selenium and selenoproteins in prostanoid metabolism and immunity. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 484–516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Saad, R.; Taylor, E.W.; Rayman, M.P. Selenium and selenoproteins in viral infection with potential relevance to COVID-19. Redox Biol. 2020, 37, 101715. [Google Scholar] [CrossRef] [PubMed]
- Blankenberg, S.; Rupprecht, H.J.; Bickel, C.; Torzewski, M.; Hafner, G.; Tiret, L.; Smieja, M.; Cambien, F.; Meyer, J.; Lackner, K.J. Glutathione peroxidase 1 activity and cardiovascular events in patients with coronary artery disease. N. Engl. J. Med. 2003, 349, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.M.; Lei, X.; Prabhu, K.S. Selenium levels affect the IL-4-induced expression of alternative activation markers in murine macrophages. J. Nutr. 2011, 141, 1754–1761. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, Z.; Chen, J.; Wang, W.; Song, D.; Li, S.; Yang, H.; Xue, S.; Chen, C. Increased levels of IL-6, IL-1β, and TNF-α in Kashin-Beck disease and rats induced by T-2 toxin and selenium deficiency. Rheumatol. Int. 2014, 34, 995–1004. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Al-Quraishy, S.; Dkhil, M.A.; Wunderlich, F.; Sies, H. Dietary selenium in adjuvant therapy of viral and bacterial infections. Adv. Nutr. 2015, 6, 73–82. [Google Scholar] [CrossRef]
- Nelson, H.K.; Shi, Q.; Van Dael, P.; Schiffrin, E.J.; Blum, S.; Barclay, D.; Levander, O.A.; Beck, M.A. Host nutritional selenium status as a driving force for influenza virus mutations. Faseb J. 2001, 15, 1727–1738. [Google Scholar] [CrossRef]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, Selenoproteins and Viral Infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef]
- Taylor, E.W.; Radding, W. Understanding Selenium and Glutathione as Antiviral Factors in COVID-19: Does the Viral M(pro) Protease Target Host Selenoproteins and Glutathione Synthesis? Front. Nutr. 2020, 7, 143. [Google Scholar] [CrossRef]
- Gopalakrishna, R.; Gundimeda, U.; Zhou, S.; Zung, K.; Forell, K.; Holmgren, A. Imbalance in Protein Thiol Redox Regulation and Cancer-Preventive Efficacy of Selenium. React. Oxyg. Species 2016, 2, 272–289. [Google Scholar] [CrossRef]
- Broome, C.S.; McArdle, F.; Kyle, J.A.; Andrews, F.; Lowe, N.M.; Hart, C.A.; Arthur, J.R.; Jackson, M.J. An increase in selenium intake improves immune function and poliovirus handling in adults with marginal selenium status. Am. J. Clin. Nutr. 2004, 80, 154–162. [Google Scholar] [CrossRef]
- Stoffaneller, R.; Morse, N.L. A review of dietary selenium intake and selenium status in Europe and the Middle East. Nutrients 2015, 7, 1494–1537. [Google Scholar] [CrossRef]
- Peng, Y.; Meng, K.; He, M.; Zhu, R.; Guan, H.; Ke, Z.; Leng, L.; Wang, X.; Liu, B.; Hu, C.; et al. Clinical Characteristics and Prognosis of 244 Cardiovascular Patients Suffering From Coronavirus Disease in Wuhan, China. J. Am. Heart Assoc. 2020, 9, e016796. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium intake, status, and health: A complex relationship. Hormones 2020, 19, 9–14. [Google Scholar] [CrossRef]
- Zabetakis, I.; Lordan, R.; Norton, C.; Tsoupras, A. COVID-19: The Inflammation Link and the Role of Nutrition in Potential Mitigation. Nutrients 2020, 12, 1466. [Google Scholar] [CrossRef]
- Raha, S.; Mallick, R.; Basak, S.; Duttaroy, A.K. Is copper beneficial for COVID-19 patients? Med. Hypotheses. 2020, 142, 109814. [Google Scholar] [CrossRef]
- Fooladi, S.; Matin, S.; Mahmoodpoor, A. Copper as a potential adjunct therapy for critically ill COVID-19 patients. Clin. Nutr. ESPEN 2020, 40, 90–91. [Google Scholar] [CrossRef]
- Besold, A.N.; Culbertson, E.M.; Culotta, V.C. The Yin and Yang of copper during infection. J. Biol. Inorg. Chem. 2016, 21, 137–144. [Google Scholar] [CrossRef]
- Andreou, A.; Trantza, S.; Filippou, D.; Sipsas, N.; Tsiodras, S. COVID-19: The Potential Role of Copper and N-acetylcysteine (NAC) in a Combination of Candidate Antiviral Treatments Against SARS-CoV-2. In Vivo 2020, 34, 1567–1588. [Google Scholar] [CrossRef]
- Wintergerst, E.S.; Maggini, S.; Hornig, D.H. Contribution of selected vitamins and trace elements to immune function. Ann. Nutr. Metab. 2007, 51, 301–323. [Google Scholar] [CrossRef]
- Bonham, M.; O’Connor, J.M.; Hannigan, B.M.; Strain, J.J. The immune system as a physiological indicator of marginal copper status? Br. J. Nutr. 2002, 87, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.S. Copper and immunity. Am. J. Clin. Nutr. 1998, 67, 1064s–1068s. [Google Scholar] [CrossRef] [PubMed]
- Pearson, P.; Britton, J.; McKeever, T.; Lewis, S.A.; Weiss, S.; Pavord, I.; Fogarty, A. Lung function and blood levels of copper, selenium, vitamin C and vitamin E in the general population. Eur. J. Clin. Nutr. 2005, 59, 1043–1048. [Google Scholar] [CrossRef]
- Xu, D.; Liu, D.; Wang, B.; Chen, C.; Chen, Z.; Li, D.; Yang, Y.; Chen, H.; Kong, M.G. In Situ OH Generation from O2- and H2O2 Plays a Critical Role in Plasma-Induced Cell Death. PLoS ONE 2015, 10, e0128205. [Google Scholar] [CrossRef] [PubMed]
- Báez-Santos, Y.M.; John, S.E.S.; Mesecar, A.D. The SARS-coronavirus papain-like protease: Structure, function and inhibition by designed antiviral compounds. Antiviral. Res. 2015, 115, 21–38. [Google Scholar] [CrossRef]
- Turnlund, J.R.; Jacob, R.A.; Keen, C.L.; Strain, J.J.; Kelley, D.S.; Domek, J.M.; Keyes, W.R.; Ensunsa, J.L.; Lykkesfeldt, J.; Coulter, J. Long-term high copper intake: Effects on indexes of copper status, antioxidant status, and immune function in young men. Am. J. Clin. Nutr. 2004, 79, 1037–1044. [Google Scholar] [CrossRef]
- Wallace, T.C. Combating COVID-19 and Building Immune Resilience: A Potential Role for Magnesium Nutrition? J. Am. Coll. Nutr. 2020, 39, 685–693. [Google Scholar] [CrossRef]
- Costello, R.B.; Elin, R.J.; Rosanoff, A.; Wallace, T.C.; Guerrero-Romero, F.; Hruby, A.; Lutsey, P.L.; Nielsen, F.H.; Rodriguez-Moran, M.; Song, Y.; et al. Perspective: The Case for an Evidence-Based Reference Interval for Serum Magnesium: The Time Has Come. Adv. Nutr. 2016, 7, 977–993. [Google Scholar] [CrossRef]
- Tang, C.F.; Ding, H.; Jiao, R.Q.; Wu, X.X.; Kong, L.D. Possibility of magnesium supplementation for supportive treatment in patients with COVID-19. Eur. J. Pharmacol. 2020, 886, 173546. [Google Scholar] [CrossRef]
- Sugimoto, J.; Romani, A.M.; Valentin-Torres, A.M.; Luciano, A.A.; Kitchen, C.M.R.; Funderburg, N.; Mesiano, S.; Bernstein, H.B. Magnesium decreases inflammatory cytokine production: A novel innate immunomodulatory mechanism. J. Immunol. 2012, 188, 6338–6346. [Google Scholar] [CrossRef]
- Iotti, S.; Wolf, F.; Mazur, A.; Maier, J.A. The COVID-19 pandemic: Is there a role for magnesium? Hypotheses and perspectives. Magnes. Res. 2020, 33, 21–27. [Google Scholar] [CrossRef]
- Toledano, J.M.; Moreno-Fernandez, J.; Puche-Juarez, M.; Ochoa, J.J.; Diaz-Castro, J. Implications of Vitamins in COVID-19 Prevention and Treatment through Immunomodulatory and Anti-Oxidative Mechanisms. Antioxidants 2021, 11, 5. [Google Scholar] [CrossRef]
- Reddy, P.; Edwards, L.R. Magnesium Supplementation in Vitamin D Deficiency. Am. J. Ther. 2019, 26, e124–e132. [Google Scholar] [CrossRef]
- Costello, R.B.; Nielsen, F. Interpreting magnesium status to enhance clinical care: Key indicators. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 504–511. [Google Scholar] [CrossRef]
- Banjanin, N.; Belojevic, G. Changes of Blood Pressure and Hemodynamic Parameters after Oral Magnesium Supplementation in Patients with Essential Hypertension-An Intervention Study. Nutrients 2018, 10, 581. [Google Scholar] [CrossRef]
- Torres, S.; Sticco, N.; Bosch, J.J.; Iolster, T.; Siaba, A.; Rivarola, M.R.; Schnitzler, E. Effectiveness of magnesium sulfate as initial treatment of acute severe asthma in children, conducted in a tertiary-level university hospital: A randomized, controlled trial. Arch. Argent. Pediatr. 2012, 110, 291–296. [Google Scholar]
- Edeas, M.; Saleh, J.; Peyssonnaux, C. Iron: Innocent bystander or vicious culprit in COVID-19 pathogenesis? Int. J. Infect. Dis. 2020, 97, 303–305. [Google Scholar] [CrossRef]
- Maggini, S.; Pierre, A.; Calder, P.C. Immune Function and Micronutrient Requirements Change over the Life Course. Nutrients 2018, 10, 1531. [Google Scholar] [CrossRef]
- Ali, M.K.; Kim, R.Y.; Karim, R.; Mayall, J.R.; Martin, K.L.; Shahandeh, A.; Abbasian, F.; Starkey, M.R.; Loustaud-Ratti, V.; Johnstone, D.; et al. Role of iron in the pathogenesis of respiratory disease. Int. J. Biochem. Cell Biol. 2017, 88, 181–195. [Google Scholar] [CrossRef]
- Cherayil, B.J. Iron and immunity: Immunological consequences of iron deficiency and overload. Arch. Immunol. Ther. Exp. 2010, 58, 407–415. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Iron homeostasis in host defence and inflammation. Nat. Rev. Immunol. 2015, 15, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Malan, L.; Baumgartner, J.; Calder, P.C.; Zimmermann, M.B.; Smuts, C.M. n-3 Long-chain PUFAs reduce respiratory morbidity caused by iron supplementation in iron-deficient South African schoolchildren: A randomized, double-blind, placebo-controlled intervention. Am. J. Clin. Nutr. 2015, 101, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Phua, J.; Weng, L.; Ling, L.; Egi, M.; Lim, C.M.; Divatia, J.V.; Shrestha, B.R.; Arabi, Y.M.; Ng, J.; Gomersall, C.D.; et al. Intensive care management of coronavirus disease 2019 (COVID-19): Challenges and recommendations. Lancet Respir. Med. 2020, 8, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Daher, R.; Manceau, H.; Karim, Z. Iron metabolism and the role of the iron-regulating hormone hepcidin in health and disease. Presse Med. 2017, 46, e272–e278. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, P.; Zhai, B.; Zhang, M.; Xiang, Y.; Fang, J.; Xu, S.; Gao, Y.; Chen, X.; Sui, X.; et al. The emerging role of ferroptosis in inflammation. Biomed. Pharmacother. 2020, 127, 110108. [Google Scholar] [CrossRef]
- Yilmaz, B.; Li, H. Gut Microbiota and Iron: The Crucial Actors in Health and Disease. Pharmaceuticals 2018, 11, 98. [Google Scholar] [CrossRef]
- Dalamaga, M.; Karampela, I.; Mantzoros, C.S. Commentary: Could iron chelators prove to be useful as an adjunct to COVID-19 Treatment Regimens? Metabolism 2020, 108, 154260. [Google Scholar] [CrossRef]
- Williams, A.; Meyer, D. Desferrioxamine as immunomodulatory agent during microorganism infection. Curr. Pharm. Des. 2009, 15, 1261–1268. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, M.; Bedi, O.; Gupta, M.; Kumar, S.; Jaiswal, G.; Rahi, V.; Yedke, N.G.; Bijalwan, A.; Sharma, S.; et al. Role of vitamins and minerals as immunity boosters in COVID-19. Inflammopharmacology 2021, 29, 1001–1016. [Google Scholar] [CrossRef]
- Haase, H. Innate Immune Cells Speak Manganese. Immunity 2018, 48, 616–618. [Google Scholar] [CrossRef]
- Chang, E.L.; Simmers, C.; Knight, D.A. Cobalt Complexes as Antiviral and Antibacterial Agents. Pharmaceuticals 2010, 3, 1711–1728. [Google Scholar] [CrossRef]
- Evgen’ev, M.B.; Frenkel, A. Possible application of H(2)S-producing compounds in therapy of coronavirus (COVID-19) infection and pneumonia. Cell Stress Chaperones 2020, 25, 713–715. [Google Scholar] [CrossRef]
- Dharmalingam, K.; Birdi, A.; Tomo, S.; Sreenivasulu, K.; Charan, J.; Yadav, D.; Purohit, P.; Sharma, P. Trace Elements as Immunoregulators in SARS-CoV-2 and Other Viral Infections. Indian J. Clin. Biochem. 2021, 36, 416–426. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Mineral | Study Design | Major Findings |
|---|---|---|---|
| Wessels I et al., 2020 [10] | Zinc | Animal model | Zinc supplementation improves lung injury through a reduction in neutrophil recruitment and activity. |
| Jothimani D et al., 2020 [11] | Zinc | Observational | A significant number of COVID-19 patients are zinc deficient, they develop more complications, and the deficiency is associated with a prolonged hospital stay and higher mortality rate. |
| te Velthuis AJ et al., 2010 [12] | Zinc | In vitro | Zn(2+) inhibits coronavirus RNA polymerase activity and its ionophores block their replication in cell culture. |
| Roscioli E et al., 2017 [13] | Zinc | In vitro | Zinc deficiency as a codeterminant for airway epithelial barrier dysfunction through an increase in its permeability. |
| Hemilä H, 2017 [14] | Zinc | Meta-analysis | Zinc lozenges are useful for common cold treatment, significantly reducing its duration. |
| Science M et al., 2012 [15] | Zinc | Meta-analysis | Oral zinc formulations shorten the duration of symptoms of the common cold. |
| Mao S et al., 2014 [16] | Zinc, Copper, Iron | Meta-analysis | Zn, Cu, and Fe deficiency may be contributing factors for the susceptibility of recurrent respiratory tract infections in children. |
| Anuk AT et al., 2021 [17] | Zinc, Copper, Magnesium | Observational | Pregnant women with COVID-19 show lower levels of serum zinc and Zn/Cu ratio, as well as higher levels of serum copper and magnesium. Serum zinc and Zn/Cu ratio concentrations had a negative relationship with acute phase markers such as IL-6 and CRP. |
| Heller RA et al., 2021 [18] | Zinc, Selenium | Observational | Zn and SELENOP status within the reference ranges is related to a higher survival odd in SARS-CoV-2 infection, and correcting a proven deficit in these trace elements by a personalized supplementation supports recovery. |
| Ben Abdallah S et al., 2023 [19] | Zinc | Clinical trial | Oral zinc can reduce 30-day death, ICU admission rate, and hospital stay, also shortening symptom duration in COVID-19 patients. |
| Reino-Gelardo S et al., 2023 [20] | Zinc, Selenium | Clinical trial | The administration of a food supplement in severe COVID-19 patients has protective effect, allowing early recovery from digestive symptoms and shorter hospital stay. |
| Natarajan S et al., 2021 [21] | Zinc | Clinical trial | In patients without clinical symptoms and comorbidities, viral load was significantly reduced on the seventh day after zinc and vitamin C supplementation, without progression to a symptomatic state. |
| Finzi E et al., 2020 [22] | Zinc | Case reports | All COVID-19 patients experienced relevant improvement after one day of high-dose zinc therapy. |
| Patel O et al., 2021 [23] | Zinc | Clinical trial | The use of high-dose intravenous Zn in hospitalized COVID-19 patients appears safe and feasible. No outcomes nor conclusions about recovery or ventilation need were drawn due to a lack of enrolment. |
| Abdelmaksoud AA et al., 2021 [24] | Zinc | Clinical trial | Zinc therapy might have a significant role in reducing the duration of smell and taste recovery in COVID-19 patients, without affecting the total recovery duration from the disease. |
| Stambouli N et al., 2022 [25] | Zinc | Clinical trial | A significant decrease in SARS-CoV-2 infection among healthcare professionals is reported when doxycycline is administered with zinc instead of just the drug. |
| Rodriguez JAM et al., 2021 [26] | Zinc, Selenium | Clinical trial | Mean levels of CD4+T, CD3+T, and CD8+T cells increase after the second dose of the COVID-19 vaccine due to the administration of a combination of β-glucans with Se- and Zn-enriched Saccharomyces cerevisiae. An augmentation in both IgG and IgM is also reported. |
| Thomas S et al., 2021 [27] | Zinc | Clinical trial | Ambulatory COVID-19 patients treated with high-dose zinc gluconate, ascorbic acid, or a combination of both, do not show a significant decrease in the duration of symptoms. |
| Abd-Elsalam S et al., 2021 [28] | Zinc | Clinical trial | Zinc supplementation in COVID-19 hospitalized patients did not improve the clinical efficacy of hydroxychloroquine. |
| Maywald M et al., 2017 [29] | Zinc | In vitro | Zinc can ameliorate the allogeneic immune reaction by enhancing antigen-specific iTreg, so it can be considered as a possible tool for inducing tolerance in adverse immune reactions. |
| Wang Y et al., 2021 [30] | Selenium | In vitro | SARS-CoV-2 suppresses mRNA expression of several selenoproteins involved in ferroptosis, endoplasmic reticulum stress, and DNA synthesis. |
| Lee YH et al., 2016 [31] | Selenium | Observational | Low selenium levels in patients suffering from respiratory diseases have a major correlation with poor nutritional status and prognosis on admission. |
| Tseng CK et al., 2013 [32] | Selenium | Observational | Serum selenium is inversely associated with inflammatory cytokine IL-6 in the elderly. |
| Gordon DE et al., 2020 [33] | Selenium | In vitro | SARS-CoV-2 viral proteases such as M-pro are able to induce the degradation of host selenoproteins. |
| Jin Z et al., 2020 [34] | Selenium | In silico | An organoselenium compound called “ebselen” shows a strong inhibitory activity against SARS-CoV-2 protease M-pro. It also performs as an important suppressor of PL-pro. |
| Mahmoodpoor A et al., 2019 [35] | Selenium | Clinical trial | Selenium administration restores lung antioxidant capacity, moderates inflammatory responses, and improves respiratory mechanics in critically ill patients with ARDS, not showing a relevant effect on overall survival, duration of mechanical ventilation, and ICU stay. |
| Zhang J et al., 2020 [36] | Selenium | Observational | There is a positive correlation between selenium concentrations and COVID-19 recovery rate. |
| Moghaddam A et al., 2020 [37] | Selenium | Observational | Selenium status was significantly higher in surviving COVID-19 patients compared with non-survivors, showing a relationship between selenium levels and mortality risk in this disease. |
| Warnes SL et al., 2015 [38] | Copper, Zinc | In vitro | Human coronavirus 229E is rapidly inactivated on a range of copper alloys, and Cu/Zn surfaces are very effective at lower copper concentrations. |
| Wazir SM et al., 2017 [39] | Copper | Case reports | Copper deficiency is associated with anemia, leucopenia, and myeloneuropathy. |
| Lee YH et al., 2019 [40] | Copper, Zinc | Observational | At the time of ICU admission, serum concentrations of copper and zinc are lower than the normal values in critically ill patients. Serum levels measured on day 14 of ICU stay are higher than those measured at the time of ICU admission for zinc and copper, being associated with a significantly lower mortality. |
| Hackler J et al., 2021 [41] | Copper, Selenium | Clinical trial | Modifications in serum biomarkers of Cu and Se status during SARS-CoV-2 infection are not compatible with a simple acute phase response, copper and SELENOP serum concentrations contribute to a good prediction of disease survival. |
| Kanellopoulou C et al., 2019 [42] | Magnesium | Animal model and in vitro | Mg2+ regulates the active site of certain kinases during T cell responses, so keeping high serum Mg2+ concentration is important in order to boost antiviral immunity. |
| Afshar Ebrahimi F et al., 2018 [43] | Magnesium, Zinc | Clinical trial | Magnesium and zinc co-supplementation has beneficial effects on serum hs-CRP, oxidative stress markers, and gene expression of IL-1 and TNF-α. |
| Yang Q et al., 2019 [44] | Magnesium | Animal model and in vitro | Magnesium isoglycyrrhizinate attenuates pulmonary fibrosis partly by inhibiting fibroblast differentiation. |
| Nasser R et al., 2018 [45] | Magnesium | Observational | Hypomagnesemia and hypermagnesemia on admission were related to an increased rate of 30-day mortality among patients hospitalized with community-acquired pneumonia. |
| Quilliot D et al., 2020 [46] | Magnesium | Observational | The risk of Mg deficiency is negatively related to SARS-CoV-2 infection severity, oxygen therapy, and ICU stay in critical care unit. There is a high prevalence of hypomagnesemia in COVID-19 hospitalized patients. |
| Tan CW et al., 2020 [47] | Magnesium | Clinical trial | A magnesium, vitamin D, and vitamin B12 combination in older COVID-19 patients is associated with a relevant reduction in clinical deterioration requiring oxygen support, ICU support, or both. |
| Fan L et al., 2021 [48] | Magnesium | Clinical trial | Twelve-week personalized Mg treatment in the elderly led to methylation changes in TMPRSS2 gene, confirming another potential role of magnesium intervention in COVID-19 prevention. |
| Agoro R et al., 2018 [49] | Iron | Animal model | Iron modulates the inflammatory response outcome through an influence in macrophage polarization. |
| Jayaweera JAAS et al., 2019 [50] | Iron | Observational | Iron deficiency makes children more prone to develop acute respiratory tract infections. Once iron deficiency is corrected, the rate of recurrent infections is reduced. |
| Ponikowski P et al., 2020 [51] | Iron | Clinical trial | In iron-deficient patients who were stabilized after acute heart failure, treatment with ferric carboxymaltose decreases the hazard of heart failure hospitalizations, without an apparent effect on cardiovascular death risk. |
| Vinke JSJ et al., 2023 [52] | Iron | Clinical trial | Intravenous iron supplementation efficiently restores its status, but humoral and cellular immune response against SARS-CoV-2 does not improve after three vaccinations. |
| Bolondi G et al., 2020 [53] | Iron | Observational | Lymphopenia is severe and constant in COVID-19 critically ill patients. Transferrin saturation is extremely reduced at ICU admission, with the same trend observed for serum iron. Hyperferritinemia is constant during ICU stay. |
| Zhao K et al., 2020 [54] | Iron | Observational | Serum iron deficiency is detected in the patients with SARS-CoV-2 infection. The severity and mortality of COVID-19 is tightly correlated with serum iron concentrations. Low iron level is an independent risk factor for death in these patients. |
| Dinc ME et al., 2016 [55] | Iron | Observational | Patients with iron deficiency have an altered olfactory function with a significantly lower threshold, discrimination, and identification capacity. |
| Ali MK et al., 2020 [56] | Iron | Animal model | Augmented accumulation of pulmonary iron has a key role in the pathogenesis of pulmonary fibrosis and lung function affectation. |
| Lippi G et al., 2020 [57] | Sodium, Potassium, Calcium | Observational | COVID-19 severity is related to lower serum levels of electrolytes such as sodium, potassium, and calcium. |
| Rodriguez-Morales AJ et al., 2020 [58] | Calcium | Meta-analysis | Calcium concentration has reported to be lower in critical COVID-19 patients, with these levels being inversely proportional to the severity of the disease. |
| Sun Y et al., 2021 [59] | Manganese | Animal model | Manganese nanodepot boosts host immune response against coronavirus, offering a simple, safe, and robust strategy against the disease. |
| Reference | Mineral | Dose | Population | Major Findings |
|---|---|---|---|---|
| Ben Abdallah S et al., 2023 [19] | Zinc | 50 mg/day of elemental Zn for 15 days | COVID-19 ambulatory and hospitalized patients | Oral zinc can reduce 30-day death, ICU admission rate, and hospital stay, also shortening symptom duration. |
| Reino-Gelardo S et al., 2023 [20] | Zinc, Selenium | 1.5 mg/day of Zn and 8.25 µg/day of Se combined with vitamin D, probiotics, and prebiotics | Severe COVID-19 and hospitalized patients | The administration of a food supplement has protective effect, allowing early recovery from digestive symptoms and shorter hospital stay. |
| Natarajan S et al., 2021 [21] | Zinc | 100 mg/day of Zn combined with ascorbic acid for 7 days | Positive COVID-19 patients without clinical symptoms | Viral load declines significantly after zinc and vitamin C supplementation, without progression to a symptomatic state. |
| Patel O et al., 2021 [23] | Zinc | 0.24 mg/kg/day of elemental Zn 7 days maximum | COVID-19 hospitalized patients | Treatment with high-dose intravenous zinc appears safe and feasible, only being associated with minimal peripheral infusion site irritation. |
| Abdelmaksoud AA et al., 2021 [24] | Zinc | 440 mg/day of Zn sulfate (50 mg elemental Zn) | COVID-19 ambulatory and hospitalized patients | Zinc therapy might have a significant role in reducing the duration of smell and taste recovery, without affecting the total recovery duration from the disease. |
| Stambouli N et al., 2022 [25] | Zinc | 15 mg/day of zinc for 6 weeks | Healthcare workers | A significant decrease in SARS-CoV-2 infection is reported when doxycycline is administered with zinc instead of just the drug. |
| Rodriguez JAM et al., 2021 [26] | Zinc, Selenium | 15 mg/day of Zn and 100 µg/day of Se combined with β-1,3/1,6-glucan for 35 days | General population | Mean levels of CD4+T, CD3+T, and CD8+T cells increase after the second dose of the COVID-19 vaccine due to the intervention. An augmentation in both IgG and IgM is also reported. |
| Thomas S et al., 2021 [27] | Zinc | 50 mg/day of zinc gluconate alone and combined with ascorbic acid for 10 days | COVID-19 ambulatory patients | Treatment with high-dose zinc gluconate alone or combined with vitamin C does not significantly decrease symptoms’ duration compared with standard care. |
| Abd-Elsalam S et al., 2021 [28] | Zinc | 220 mg of Zn sulfate (50 mg of elemental Zn) twice daily | COVID-19 hospitalized patients | Zinc supplementation did not improve the clinical efficacy of hydroxychloroquine. |
| Tan CW et al., 2020 [47] | Magnesium | 150 mg/day of Mg for 14 days maximum, combined with vitamin D and B12 | Elderly COVID-19 hospitalized patients | The administration of this oral combination in older COVID-19 patients is associated with a relevant reduction in clinical deterioration requiring oxygen support, ICU support, or both. |
| Fan L et al., 2021 [48] | Magnesium | Personalized to reach a calcium-to-magnesium intake ratio ≥ 2.6 | Population over 65 years old | Twelve-week personalized Mg treatment led to methylation changes in TMPRSS2 gene, confirming another potential role of magnesium intervention in COVID-19 prevention. |
| Ponikowski P et al., 2021 [51] | Iron | Not specified | Iron-deficient patients stabilized after acute heart failure | Treatment with ferric carboxymaltose decreases the hazard of heart failure hospitalizations, without an apparent effect on cardiovascular death risk |
| Vinke JSJ et al., 2023 [52] | Iron | 1–4 doses of 500 mg of ferric carboxymaltose with six-week intervals | Iron-deficient kidney transplanted patients | Intravenous supplementation efficiently restores iron status, but humoral and cellular immune response against the virus does not improve after three vaccinations. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toledano, J.M.; Puche-Juarez, M.; Moreno-Fernandez, J.; Ochoa, J.J.; Diaz-Castro, J. Antioxidant and Immune-Related Implications of Minerals in COVID-19: A Possibility for Disease Prevention and Management. Antioxidants 2023, 12, 1104. https://doi.org/10.3390/antiox12051104
Toledano JM, Puche-Juarez M, Moreno-Fernandez J, Ochoa JJ, Diaz-Castro J. Antioxidant and Immune-Related Implications of Minerals in COVID-19: A Possibility for Disease Prevention and Management. Antioxidants. 2023; 12(5):1104. https://doi.org/10.3390/antiox12051104
Chicago/Turabian StyleToledano, Juan M., María Puche-Juarez, Jorge Moreno-Fernandez, Julio J. Ochoa, and Javier Diaz-Castro. 2023. "Antioxidant and Immune-Related Implications of Minerals in COVID-19: A Possibility for Disease Prevention and Management" Antioxidants 12, no. 5: 1104. https://doi.org/10.3390/antiox12051104
APA StyleToledano, J. M., Puche-Juarez, M., Moreno-Fernandez, J., Ochoa, J. J., & Diaz-Castro, J. (2023). Antioxidant and Immune-Related Implications of Minerals in COVID-19: A Possibility for Disease Prevention and Management. Antioxidants, 12(5), 1104. https://doi.org/10.3390/antiox12051104