
Stability and Bioaccessibility of Phenolic Compounds in Rosehip Extracts during In Vitro Digestion

 ,
, 
 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Extracts
2.2. In Vitro Digestions
2.2.1. Gastrointestinal Digestion
2.2.2. Colonic Digestion
2.3. Determination of Phenolic Compounds
2.4. Determination of Antioxidant Activity
2.5. Statistical Treatment
3. Results and Discussion
3.1. Characterization of Rosehip Extracts
3.1.1. Antioxidant Capacity
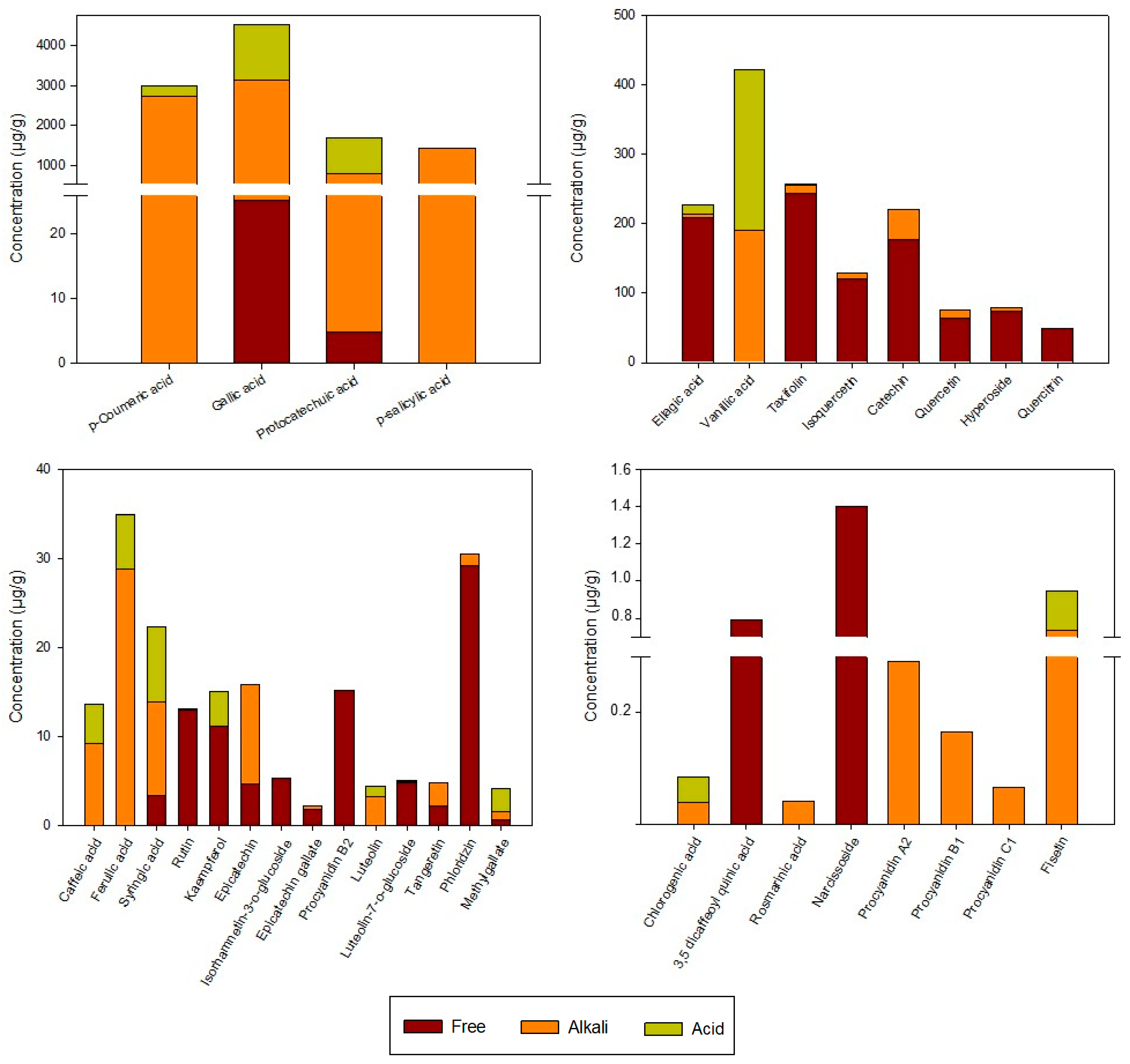
3.1.2. Individual Phenols
3.2. Effect of Gastrointestinal and Colonic Digestion on Total and Individual Phenolic Content and Antioxidant Properties of Rosehip Extracts
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- García-Lomillo, J.; González-SanJosé, M.L. Applications of wine pomace in the food industry: Approaches and functions. Compr. Rev. Food Sci. Food Saf. 2017, 16, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Özkan, G.; Sagdiç, O.; Baydar, N.G.; Baydar, H. Note: Antioxidant and antibacterial activities of Rosa damascena flower extracts. Food Sci. Technol. Int. 2004, 10, 277–281. [Google Scholar] [CrossRef]
- Jiménez, S.; Gascón, S.; Luquin, A.; Laguna, M.; Ancin-Azpilicueta, C.; Rodríguez-Yoldi, M.J. Rosa canina extracts have antiproliferative and antioxidant effects on Caco-2 human colon cancer. PLoS ONE 2016, 11, e0159136. [Google Scholar] [CrossRef] [PubMed]
- Warholm, O.; Skaar, S.; Hedman, E.; Mølmen, H.M.; Eik, L. The effects of a standardized herbal remedy made from a subtype of Rosa canina in patients with osteoarthritis: A double-blind, randomized, placebo-controlled clinical trial. Curr. Ther. Res. 2003, 64, 21–31. [Google Scholar] [CrossRef]
- Cohen, M. Rosehip—An evidence based herbal medicine for inflammation and arthritis. Aust. Fam. Physician 2012, 41, 495–498. [Google Scholar]
- Paunović, D.; Kalušević, A.; Petrović, T.; Urošević, T.; Djinović, D.; Nedović, V.; Popović-Djordjević, J. Assessment of chemical and antioxidant properties of fresh and dried rosehip (Rosa canina L.). Not. Bot. Horti. Agrobot. 2019, 47, 108–113. [Google Scholar] [CrossRef]
- Popović-Djordjević, J.; Paunović, D.; Milić, A.; Krstić, Đ.; Siavash Moghaddam, S.; Roje, V. Multi-elemental analysis, pattern recognition techniques of wild and cultivated rosehips from Serbia, and nutritional aspect. Biol. Trace Elem. Res. 2021, 199, 1110–1122. [Google Scholar] [CrossRef]
- Kulaitienė, J.; Medveckienė, B.; Levickienė, D.; Vaitkevičienė, N.; Makarevičienė, V.; Jarienė, E. Changes in fatty acids content in organic rosehip (Rosa spp.) seeds during ripening. Plants 2020, 9, 1793. [Google Scholar] [CrossRef]
- Winther, K.; Vinther Hansen, A.S.; Campbell-Tofte, J. Bioactive ingredients of rose hips (Rosa canina L) with special reference to antioxidative and anti-inflammatory properties: In vitro studies. Bot. Targets Ther. 2016, 6, 11–23. [Google Scholar] [CrossRef]
- Shahidi, F. Functional foods: Their role in health promotion and disease prevention. J. Food Sci. 2006, 69, R146–R149. [Google Scholar] [CrossRef]
- Vaz, A.Á.; Odriozola-Serrano, I.; Oms-Oliu, G.; Martín-Belloso, O. Physicochemical properties and bioaccessibility of phenolic compounds of dietary fibre concentrates from vegetable by-products. Foods 2022, 11, 2578. [Google Scholar] [CrossRef]
- Reboul, E.; Richelle, M.; Perrot, E.; Desmoulins-Malezet, C.; Pirisi, V.; Borel, P. Bioaccessibility of carotenoids and vitamin e from their main dietary sources. J. Agric. Food Chem. 2006, 54, 8749–8755. [Google Scholar] [CrossRef]
- Al-Yafeai, A.; Böhm, V. In vitro bioaccessibility of carotenoids and vitamin E in rosehip products and tomato paste as affected by pectin contents and food processing. J. Agric. Food Chem. 2018, 66, 3801–3809. [Google Scholar] [CrossRef]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Gonçalves, S.; Hernanz, D.; Heredia, F.J.; Romano, A. Effects of in vitro gastrointestinal digestion on phenolic compounds and antioxidant activity of different white winemaking byproducts extracts. Food Res. Int. 2018, 109, 433–439. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, T.; Zhang, Y.; Chen, Y.; Ge, X.; Sui, W.; Zhu, Q.; Geng, J.; Zhang, M. Release of bound polyphenols from wheat bran soluble dietary fiber during simulated gastrointestinal digestion and colonic fermentation in vitro. Food Chem. 2023, 402, 134111. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Durand, M.; Dumay, C.; Beaumatin, P.; Morel, M.T. Use of the rumen simulation technique (rusitec) to compare microbial digestion of various by-products. Anim. Feed Sci. Technol. 1988, 21, 197–204. [Google Scholar] [CrossRef]
- Mattila, P.; Kumpulainen, J. Determination of free and total phenolic acids in plant-derived foods by HPLC with diode-array detection. J. Agric. Food Chem. 2002, 50, 3660–3667. [Google Scholar] [CrossRef]
- Delpino-Rius, A.; Eras, J.; Vilaró, F.; Cubero, M.A.; Balcells, M.; Canela-Garayoa, R. Characterisation of phenolic compounds in processed fibres from the juice industry. Food Chem. 2015, 172, 575–584. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by Ferric Reducing Antioxidant Power (FRAP), ABTS bleaching assay (TEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Su, L.; Yin, J.-J.; Charles, D.; Zhou, K.; Moore, J.; Yu, L.L. Total phenolic contents, chelating capacities, and radical-scavenging properties of black peppercorn, nutmeg, rosehip, cinnamon and oregano leaf. Food Chem. 2007, 100, 990–997. [Google Scholar] [CrossRef]
- Goztepe, B.; Selma Kayacan, S.; Bozkurt, F.; Tomas, M.; Osman Sagdic, O.; Karasu, S. Drying kinetics, total bioactive compounds, antioxidant activity, phenolic profile, lycopene and β-carotene content and color quality of rosehip dehydrated by different methods. LWT-Food Sci. Technol. 2022, 153, 112476. [Google Scholar] [CrossRef]
- Demir, N.; Yildiz, O.; Alpaslan, M.; Hayaloglu, A.A. Evaluation of volatiles, phenolic compounds and antioxidant activities of rose hip (Rosa L.) fruits in Turkey. LWT-Food Sci. Technol. 2014, 57, 126–133. [Google Scholar] [CrossRef]
- Elmastaς, M.; Demir, A.; Genç, N.; Dölek, U.; Günes, M. Changes in flavonoid and phenolic acid contents in some Rosa species during ripening. Food Chem. 2017, 235, 154–159. [Google Scholar] [CrossRef]
- Zhu, L.; Li, W.; Deng, Z.; Li, H.; Zhang, B. The composition and antioxidant activity of bound phenolics in three legumes, and their metabolism and bioaccessibility of gastrointestinal tract. Foods 2020, 9, 1816. [Google Scholar] [CrossRef]
- Olech, M.; Pietrzak, W.; Nowak, R. Characterization of free and bound phenolic acids and flavonoid aglycones in Rosa rugosa Thunb. leaves and achenes using LC–ESI–MS/MS–MRM methods. Molecules 2020, 25, 1804. [Google Scholar] [CrossRef]
- Jiménez, S.; Jiménez-Moreno, N.; Luquin, A.; Laguna, M.; Rodríguez-Yoldi, M.J.; Ancín-Azpilicueta, C. Chemical composition of rosehips from different Rosa species: An alternative source of antioxidants for the food industry. Food Addit. Contam. Part A 2017, 34, 1121–1130. [Google Scholar] [CrossRef]
- Huang, D.; Li, C.; Chen, Q.; Xie, X.; Fu, X.; Chen, C.; Huang, Q.; Huang, Z.; Dong, H. Identification of polyphenols from Rosa roxburghii Tratt pomace and evaluation of in vitro and in vivo antioxidant activity. Food Chem. 2022, 377, 131922. [Google Scholar] [CrossRef]
- Ghendov-Mosanu, A.; Cristea, E.; Patras, A.; Sturza, R.; Niculaua, M. Rose hips, a valuable source of antioxidants to improve gingerbread characteristics. Molecules 2020, 25, 5659. [Google Scholar] [CrossRef]
- Terao, J. Potential role of quercetin glycosides as anti-atherosclerotic food-derived factors for human health. Antioxidants 2023, 12, 258. [Google Scholar] [CrossRef]
- Guimarães, R.; Barros, L.; Dueñas, M.; Carvalho, A.M.; Queiroz, M.J.R.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Characterisation of phenolic compounds in wild fruits from Northeastern Portugal. Food Chem. 2013, 141, 3721–3730. [Google Scholar] [CrossRef]
- Tumbas, V.T.; Canadanović-Brunet, J.M.; Dragana, D.; Četojević-Simin, D.D.; Cetković, G.S.; Dilas, S.M.; Gillec, L. Effect of rosehip (Rosa canina L.) phytochemicals on stable free radicals and human cancer cells. J. Sci. Food Agric. 2012, 92, 1273–1281. [Google Scholar] [CrossRef]
- Fecka, I. Qualitative and quantitative determination of hydrolysable tannins and other polyphenols in herbal products from meadowsweet and dog rose. Phytochem. Anal. 2009, 20, 177–190. [Google Scholar] [CrossRef]
- Bhave, A.; Schulzova, V.; Chmelarova, H.; Mrnka, L.; Hajslova, J. Assessment of rosehips based on the content of their biologically active compounds. J. Food Drug. Anal. 2017, 25, 681–690. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accesibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Xie, J.; Schaich, K.-M. Re-evaluation of the 2,2-Diphenyl-1-picrylhydrazyl free radical (DPPH) assay for antioxidant activity. J. Agric. Food Chem. 2014, 62, 4251–4260. [Google Scholar] [CrossRef]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin-Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Fawole, O.A.; Opara, U.L. Stability of total phenolic concentration and antioxidant capacity of extracts from pomegranate co-products subjected to in vitro digestion. BCM Complement. Altern. Med. 2016, 16, 358. [Google Scholar] [CrossRef] [PubMed]
- Campos-Vega, R.; Vázquez-Sánchez, K.; López-Barrera, D.; Loarca-Piña, G.; Mendoza-Díaz, S.; Oomah, B.D. Simulated gastrointestinal digestion and in vitro colonic fermentation of spent coffee (Coffea arabica L.): Bioaccessibility and intestinal permeability. Food Res. Int. 2015, 77, 156–161. [Google Scholar] [CrossRef]
- Correa, V.G.; Gonçalves, G.A.; de Sá-Nakanishi, A.B.; Ferreira, I.C.F.R.; Barros, L.; Dias, M.I.; Koehnlein, E.A.; de Souza, C.G.M.; Bracht, A.; Peralta, R.M. Effects of in vitro digestion and in vitro colonic fermentation on stability and functional properties of yerba mate (Ilex paraguariensis A. St. Hil.) beverages. Food Chem. 2017, 237, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Sirisena, S.; Ajlouni, S.; Ng, K. Simulated gastrointestinal digestion and in vitro colonic fermentation of date (Phoenix dactylifera L.) seed polyphenols. Int. J. Food Sci. Technol. 2018, 53, 412–422. [Google Scholar] [CrossRef]
- Chait, Y.A.; Gunenc, A.; Bendali, F.; Hosseinian, F. Simulated gastrointestinal digestion and in vitro colonic fermentation of carob polyphenols: Bioaccessibility and bioactivity. Food Sci. Technol. 2020, 117, 108623. [Google Scholar] [CrossRef]
- Medveckiene, B.; Kulaitiene, J.; Jariene, E.; Vaitkeviciene, N.; Hallman, E. Carotenoids, polyphenols, and ascorbic acid in organic rosehips (Rosa spp.) cultivated in Lithuania. Appl. Sci. 2020, 10, 5337. [Google Scholar] [CrossRef]
- Bas-Bellver, C.; Andrés, C.; Seguí, L.; Barrera, C.; Jiménez-Hernández, N.; Artacho, A.; Betoret, N.; Gosalbes, M.J. Valorization of persimmon and blueberry byproducts to obtain functional powders: In vitro digestion and fermentation by gut microbiota. J. Agric. Food Chem. 2020, 68, 8080–8090. [Google Scholar] [CrossRef]
- Padayachee, A.; Netzel, G.; Netzel, M.; Day, L.; Mikkelsena, D.; Gidley, M.J. Lack of release of bound anthocyanins and phenolic acids from carrot plant cell walls and model composites during simulated gastric and small intestinal digestion. Food Funct. 2013, 4, 906. [Google Scholar] [CrossRef]
- Pérez-Jimenez, J.; Díaz-Rubio, M.E.; Saura-Calixto, F. Non-extractable polyphenols, a major dietary antioxidant: Occurrence, metabolic fate and health effects. Nutr. Res. Rev. 2013, 26, 118–129. [Google Scholar] [CrossRef]
- Ruíz-García, Y.; Beres, C.; Chávez, D.W.H.; Pereira, D.C.S.; Santiago, M.C.P.A.; Godoy, R.O.; dos Santos Gomes, F.; Antoniassi, R.; Tonon, R.V.; Cabral, L.M.C. In vitro digestion and colonic fermentation of an Alicante Bouschet (Vitis vinifera L.) skin extract. LWT-Food Sci. Technol. 2022, 157, 113083. [Google Scholar] [CrossRef]
- Ortega, N.; Macia, A.; Romero, M.P.; Reguant, J.; Motilva, M.J. Matrix composition effect on the digestibility of carob flour phenols by an in vitro digestion model. Food Chem. 2011, 124, 65–71. [Google Scholar] [CrossRef]
- Xue, M.; Shi, H.; Zhang, J.; Liu, Q.Q.; Guan, J.; Zhang, J.Y.; Ma, Q. Stability and degradation of caffeoylquinic acids under different storage conditions studied by High-Performance Liquid Chromatography with photo diode array detection and High-Performance Liquid Chromatography with Electrospray Ionization Collision-Induced Dissociation Tandem Mass Spectrometry. Molecules 2016, 21, 948. [Google Scholar] [CrossRef]
- Targino Dutraa, R.L.; Macedo Dantasa, A.; de Araújo Marquesa, D.; Ferreira Batistaa, J.D.; Lins de Albuquerque Meirelesb, B.R.; Tribuzy de Magalhães Cordeirob, A.M.; Magnanic, M.; da Silva Campelo Borge, G. Bioaccessibility and antioxidant activity of phenolic compounds in frozen pulps of Brazilian exotic fruits exposed to simulated gastrointestinal conditions. Food Res. Int. 2017, 100, 650–657. [Google Scholar] [CrossRef]
- Bermúdez-Soto, M.J.; Tomás-Barberán, F.A.; García-Conesa, M.T. Stability of polyphenols in chokeberry (Aronia melanocarpa) subjected to in vitro gastric and pancreatic digestion. Food Chem. 2007, 102, 865–874. [Google Scholar] [CrossRef]
- Rocha-Amador, O.G.; Gallegos-Infante, J.A.; Huang, Q.; González-Laredo, R.F. Effect of glycosylation degree of quercetin on its in vitro bioaccessibility in food grade organogels. Int. J. Food Eng. 2017, 13, 20170166. [Google Scholar] [CrossRef]
- Lucas-González, R.; Viuda-Martos, M.; Pérez Álvarez, J.A.; Fernández-López, J. Changes in bioaccessibility, polyphenol profile and antioxidant potential of flours obtained from persimmon fruit (Diospyros kaki) co-products during in vitro gastrointestinal digestion. Food Chem. 2018, 256, 252–258. [Google Scholar] [CrossRef]
- Niwano, Y.; Kohzaki, H.; Shirato, M.; Shishido, S.; Nakamura, K. Metabolic fate of orally ingested proanthocyanidins through the digestive tract. Antioxidants 2023, 12, 17. [Google Scholar] [CrossRef]
- Neilson, A.P.; George, J.C.; Janle, E.M.; Mattes, R.D.; Rudolph, R.; Matusheski, N.V.; Ferruzzi, M.G. Influence of chocolate matrix composition on cocoa flavan-3-ol bioaccessibility in vitro and bioavailability in humans. J. Agric. Food Chem. 2009, 57, 9418–9426. [Google Scholar] [CrossRef]
- Green, R.J.; Murphy, A.S.; Schulz, B.; Watkins, B.A.; Ferruzzi, M.G. Common tea formulations modulate in vitro digestive recovery of green tea catechins. Mol. Nutr. Food Res. 2007, 51, 1152–1162. [Google Scholar] [CrossRef]
- Yan, F.; Zhao, L.; Chena, W.; Lu, Q.; Tang, C.; Wang, C.; Liu, R. Comparison of the inhibitory effects of procyanidins with different structures and their digestion products against acrylamide-induced cytotoxicity in IPEC-J2 cells. J. Funct. Foods. 2020, 72, 104073. [Google Scholar] [CrossRef]
- Li, N.; Taylor, L.S.; Ferruzzi, M.G.; Mauer, L.J. Kinetic study of catechins stability: Effects of pH, concentration, and temperature. J. Agric. Food Chem. 2012, 60, 12531–12539. [Google Scholar] [CrossRef] [PubMed]
- Ortega, N.; Reguant, J.; Romero, M.P.; Macià, A.; Motilva, M.J. Effect of fat content on the digestibility and bioaccessibility of cocoa polyphenol by an in vitro digestion model. J. Agric. Food Chem. 2009, 57, 5743–5749. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez-Díaz, J.L.; Moreno-Ortega, A.M.; Roldán-Guerra, F.J.; Ortíz-Somovilla, V.; Moreno-Rojas, J.M.; Pereira-Caro, G. In vitro gastrointestinal digestion and colonic catabolism of mango (Mangifera indica L.) pulp polyphenols. Foods 2020, 9, 1836. [Google Scholar] [CrossRef] [PubMed]
- Urpí-Sarda, M.; Monagas, M.; Khan, N.; Llorach, R.; Lamuela-Raventós, R.M.; Jáuregui, O.; Estruch, R.; Izquierdo-Pulido, M.; Andrés-Lacueva, C. Targeted metabolic profiling of phenolics in urine and plasma after regular consumption of cocoa by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2009, 1216, 7258–7267. [Google Scholar] [CrossRef]
- Ou, K.; Sarnoski, P.; Schneider, K.R.; Song, K.; Khoo, C.; Gu, L. Microbial catabolism of procyanidins by human gut microbiota. Mol. Nutr. Food Res. 2014, 58, 2196–2205. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Tran, M.H.; Lee, E.Y. Co-upgrading of ethanol-assisted depolymerized lignin: A new biological lignin valorization approach for the production of protocatechuic acid and polyhydroxyalkanoic acid. Bioresour. Technol. 2021, 338, 125563. [Google Scholar] [CrossRef]
- Cassidy, A.; Minihane, A.M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef]
- Alegría, A.; García-Llatas, G.; Cilla, A. Static digestion models: General Introduction. In The Impact of Food Bioactives on Health In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; SpringerOpen: London, UK, 2015; Chapter 1; pp. 3–12. [Google Scholar] [CrossRef]
- Lucas-González, R.; Viuda-Martos, M.; Pérez-Álvarez, J.A. In vitro digestion models suitable for foods: Opportunities for new fields of application and challenges. Food Res. Int. 2018, 107, 423–436. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Initial (Free Fraction) (µg/g) | SGD 1 (µg/g) | SPSID 2 (µg/g) | BI 3 (%) | IPSID 4 (µg/g) | SCD 5 (µg/g) | |
|---|---|---|---|---|---|---|
| ACIDS | ||||||
| Caffeic acid | Nd | 0.83 b | 1.8 a | 13.4 | 0.016 d | 0.060 c |
| Chlorogenic acid | Nd | 2.2 a | 2.7 a | 148 | 0.013 b | 0.072 b |
| p-Coumaric acid | Nd | 2.5 b | 65.3 a | 2.2 | 0.118 c | 0.075 c |
| 3,5-Dicaffeoylquinic acid | 0.79 | nd | nd | - | nd | nd |
| Ellagic acid | 208.0 a | 100.7 b | 212.3 a | 93.4 | 2.115 c | 0.936 d |
| Ferulic acid | Nd | nd | nd | - | nd | nd |
| Gallic acid | 25.2 a | 13.9 b | 30.9 a | 0.68 | 0.247 c | 0.085 d |
| Protocatechuic acid | 4.9 b | 32.1 a | 43.3 a | 2.5 | 0.163 d | 0.397 c |
| Rosmarinic acid | Nd | nd | nd | - | nd | nd |
| p-Salicylic acid | Nd | 1.4 a | nd | - | 0.111 b | 0.042 c |
| o-Salicylic acid | Nd | nd | nd | - | nd | 0.235 |
| Syringic acid | 3.4 a | 1.4 c | 1.7 b | 7.6 | 0.048 e | 0.122 d |
| Vanillic acid | Nd | nd | nd | - | 0.103 | nd |
| Total phenolic acids | 242 | 155 | 358 | 3 | 2.934 | 2.024 |
| FLAVONOLS | ||||||
| Quercetin | 64.6 a | 43.0 b | 33.0 c | 43.2 | 1.285 d | 0.114 e |
| Taxifolin | 243.7 a | 189.9 b | 155.6 b | 61.1 | 2.101 c | 0.089 d |
| Isoquercetin | 120.2 a | 109.0 a | 111.7 a | 86.8 | 2.129 b | 0.059 c |
| Hyperoside | 73.6 b | 83.0 a | 74.8 b | 95.2 | 2.446 c | nd |
| Quercitrin | 53.7 a | 56.3 a | 57.4 a | 107 | 0.687 b | 0.037 c |
| Kaempferol | 11.1 | nd | nd | - | nd | nd |
| Rutin | 15.6 b | 19.3 a | 21.8 a | 138 | 0.179 c | nd |
| Narcissoside | 1.4 b | 1.8 a | 1.7 ab | 121 | 0.020 d | 0.032 c |
| Isorhamnetin-3-O-glucoside | 5.4 a | 4.7 a | 4.5 a | 83.3 | 0.163 b | nd |
| Total flavonols | 589 | 507 | 460 | 73 | 9.010 | 0.331 |
| FLAVAN-3-OLS | ||||||
| Catechin | 177 a | 109.7 b | 91.8 c | 41.9 | 2.857 d | 0.140 e |
| Epicatechin | 4.7 a | 1.8 c | 2.4 b | 15.2 | 0.070 d | 0.051 d |
| Epicatechin gallate | 1.5 b | 1.9 a | 1.8 a | 92.3 | 0.081 c | nd |
| Procyanidin A2 | 1.1 b | 1.5 a | 1.5 a | 104 | 0.173 c | 0.165 c |
| Procyanidin B1 | 56.5 b | 42.1 b | 120.7 a | 213 | 1.420 c | 0.063 d |
| Procyanidin B2 | 15.2 a | 9.3 b | 6.1 b | 40.1 | 0.155 c | nd |
| Procyanidin C1 | 2.6 a | 3.7 a | nd | - | 0.066 b | nd |
| Total flavanols | 259 | 170 | 224 | 72 | 4.822 | 0.419 |
| FLAVONES | ||||||
| Luteolin | Nd | nd | nd | - | nd | Nd |
| Luteolin-7-O-glucoside | 7.8 a | 7.4 a | 7.5 a | 92.6 | 0.121 b | Nd |
| Tangeretin | 2.3 a | nd | 1.9 b | 39.4 | nd | 0.114 c |
| Fisetin | Nd | nd | nd | - | nd | Nd |
| Total flavones | 10 | 7 | 9 | 50 | 0.121 | 0.114 |
| OTHERS | ||||||
| Phlorizin | 29.2 b | 52.6 a | 46.7 a | 153.1 | 0.531 c | Nd |
| Methyl gallate | 0.70 c | 1.1 b | 4.1 a | 97.6 | 0.153 d | 0.074 e |
| TOTAL FREE PHENOLIC COMPOUNDS | 1130 | 893 | 1103 | 9 | 17.571 | 2.962 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Odriozola-Serrano, I.; Nogueira, D.P.; Esparza, I.; Vaz, A.A.; Jiménez-Moreno, N.; Martín-Belloso, O.; Ancín-Azpilicueta, C. Stability and Bioaccessibility of Phenolic Compounds in Rosehip Extracts during In Vitro Digestion. Antioxidants 2023, 12, 1035. https://doi.org/10.3390/antiox12051035
Odriozola-Serrano I, Nogueira DP, Esparza I, Vaz AA, Jiménez-Moreno N, Martín-Belloso O, Ancín-Azpilicueta C. Stability and Bioaccessibility of Phenolic Compounds in Rosehip Extracts during In Vitro Digestion. Antioxidants. 2023; 12(5):1035. https://doi.org/10.3390/antiox12051035
Chicago/Turabian StyleOdriozola-Serrano, Isabel, Danielle P. Nogueira, Irene Esparza, Ana A. Vaz, Nerea Jiménez-Moreno, Olga Martín-Belloso, and Carmen Ancín-Azpilicueta. 2023. "Stability and Bioaccessibility of Phenolic Compounds in Rosehip Extracts during In Vitro Digestion" Antioxidants 12, no. 5: 1035. https://doi.org/10.3390/antiox12051035
APA StyleOdriozola-Serrano, I., Nogueira, D. P., Esparza, I., Vaz, A. A., Jiménez-Moreno, N., Martín-Belloso, O., & Ancín-Azpilicueta, C. (2023). Stability and Bioaccessibility of Phenolic Compounds in Rosehip Extracts during In Vitro Digestion. Antioxidants, 12(5), 1035. https://doi.org/10.3390/antiox12051035