
Addition of Spirulina to Craft Beer: Evaluation of the Effects on Volatile Flavor Profile and Cytoprotective Properties


 ,
, 
 ,
, 
 and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Production Process
- (1)
- 45 °C for 10 min (protease enzymes react to hydrolyze low-weight protein as nourishment for yeast);
- (2)
- 66 °C for 40 min (β-amylase activity, pH 5.0–5.5, enzymatic synergy points between amylases);
- (3)
- 72 °C for 20 min (α-amylase activity, pH 5.6–5.8, maximum activity);
- (4)
- 78 °C for 5 min (enzymatical inactivation phase).
2.2. Materials
2.3. Chemicals
2.4. SPME Sampling
2.5. GC-MS Analisys of Beers
2.6. GC-MS Analysis of Spirulina after Derivatization
2.7. PTR-ToF-MS Analysis of Beers
2.8. Spectrophotometric Analysis of Total Polyphenols and Tannins
2.9. Radical Scavenging Activity
2.10. Confocal Microscopy of Beer Yeast Cells
2.11. Cytoprotective Activity towards Oxidative Stress
2.11.1. Cell Culture
2.11.2. Cytotoxicity
2.11.3. Cytoprotection towards the Oxidative Damage induced by Tert-Butyl Hydroperoxide (t-BOOH)
2.11.4. Immunofluorescence Analysis of Nrf2
2.12. Statistical Analysis
3. Results
3.1. Vapor-Phase Beer Chemical Composition
3.2. Chemical Composition of Spirulina after Derivatization
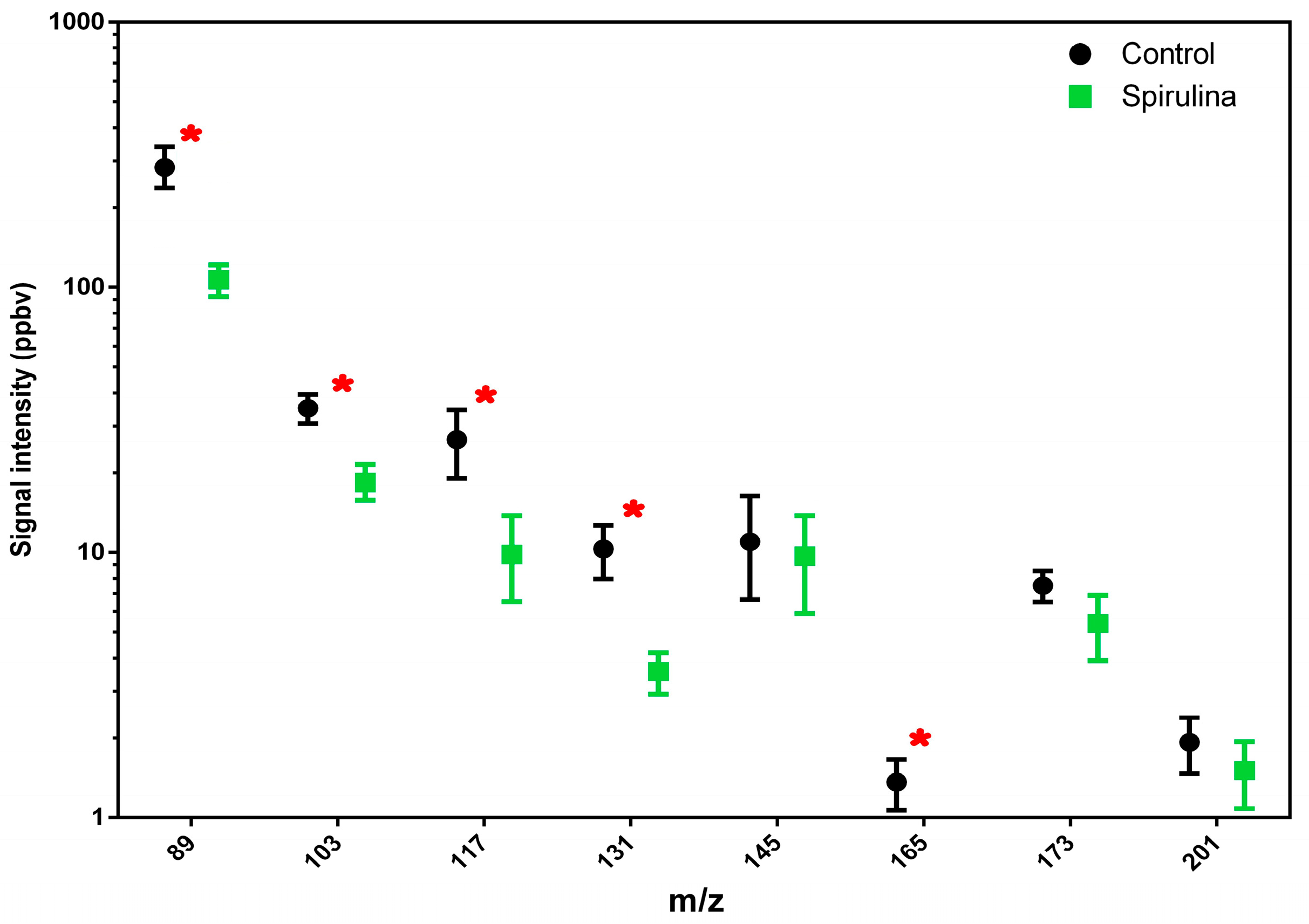
3.3. PTR-ToF-MS: Determination of Volatile Compounds from Beers
3.4. Spectrophotometric Analysis of Total Polyphenols and Tannins
3.5. Radical Scavenging Activity
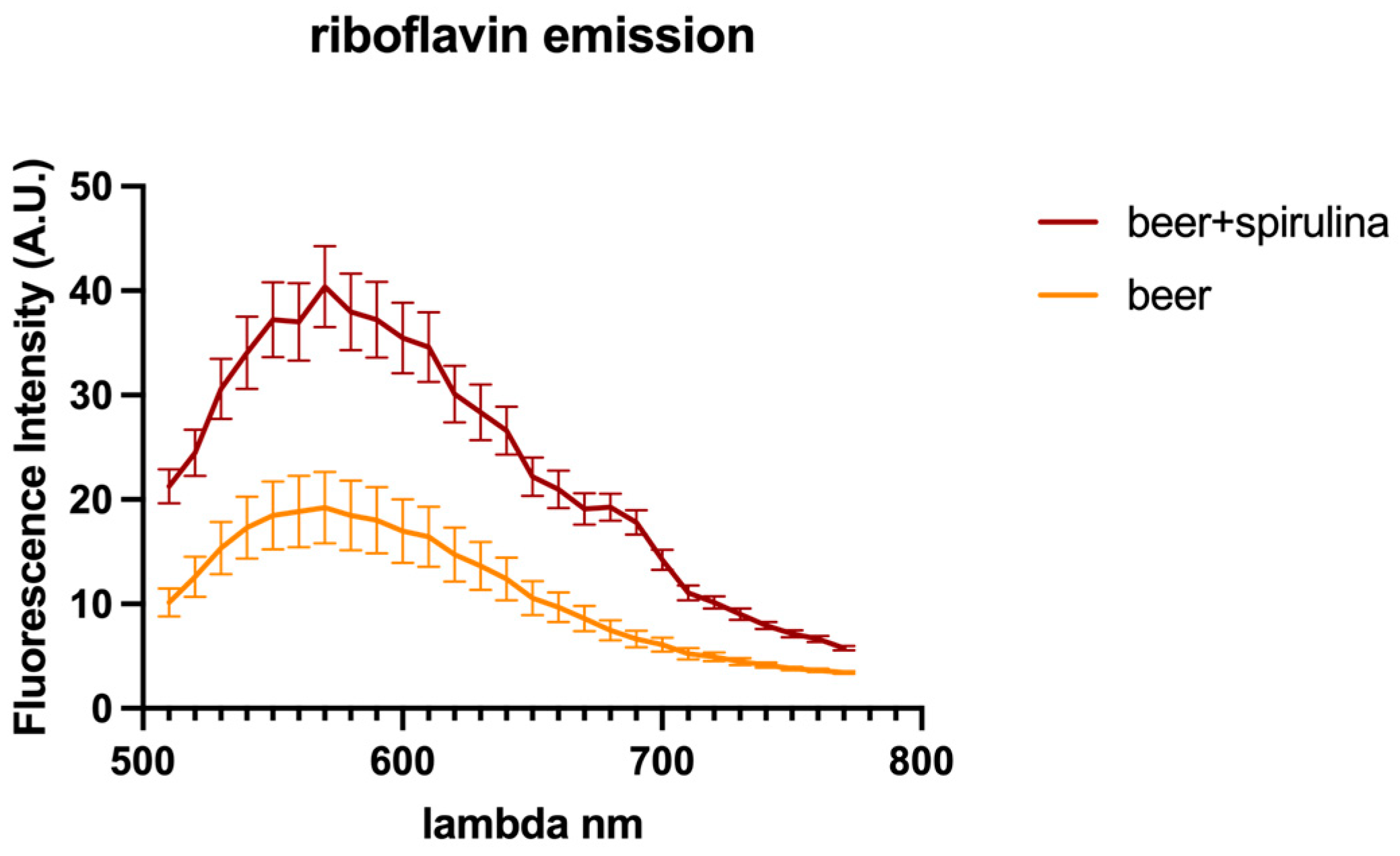
3.6. Confocal Microscopy Analysis
3.7. Antioxidant Cytoprotective Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palamand, S.R.; Aldenhoff, J.M. Bitter tasting compounds of beer: Chemistry and taste properties of some hop resin compounds. J. Agric. Food Chem. 1973, 21, 535–543. [Google Scholar] [CrossRef]
- Ravindra, P.A. Value-added food: Single cell protein. Biotechnol. Adv. 2000, 18, 459–479. [Google Scholar]
- Verdelho, V.V.; Cadoret, J.P.; Acien, F.G.; Benemann, J. Clarification of most relevant concepts related to the microalgae production sector. Processes 2022, 10, 175. [Google Scholar] [CrossRef]
- Sibiya, T.; Ghazi, T.; Mohan, J.; Nagiah, S.; Chuturgoon, A.A. Spirulina platensis ameliorates oxidative stress associated with antiretroviral drugs in hepg2 cells. Plants 2022, 11, 3143. [Google Scholar] [CrossRef] [PubMed]
- Gogna, S.; Kaur, J.; Sharma, K.; Prasad, R.; Singh, J.; Bhadariya, V.; Kumar, P.; Jarial, S. Spirulina-an edible cyanobacterium with potential therapeutic health benefits and toxicological consequences. J. Am. Nutr. Assoc. 2022, 2, 1–14. [Google Scholar] [CrossRef]
- Metekia, W.A.; Ulusoy, B.H.; Habte-Tsion, H.M. Spirulina phenolic compounds: Natural food additives with antimicrobial properties. Int. Food Res. J. 2021, 28, 1109–1118. [Google Scholar] [CrossRef]
- Belay, A.; Ota, Y.; Miyakawa, K.; Shimamatsu, H. Current knowledge on potential health benefits of Spirulina. J. Appl. Phycol. 1993, 5, 235–241. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; De-Paula, J.C.; da S. Pereira, L.O.; Marquez, U.M.L. Amino acid composition, protein content and calculation of nitrogen-to-protein conversion factors for 19 tropical seaweeds. Phycol. Res. 2002, 50, 233–241. [Google Scholar] [CrossRef]
- Santos, T.D.; de Freitas, B.C.B.; Moreira, J.B.; Zanfonato, K.; Costa, J.A.V. Development of powdered food with the addition of Spirulina for food supplementation of the elderly population. Innov. Food Sci. Emerg. Technol. 2016, 37, 216–220. [Google Scholar] [CrossRef]
- Fradique, M.; Batista, A.P.; Nunes, M.C.; Gouveia, L.; Bandarra, N.M.; Raymundo, A. Incorporation of Chlorella vulgaris and Spirulina maxima biomass in pasta products. Part 1: Preparation and evaluation. J. Sci. Food Agric. 2010, 90, 1656–1664. [Google Scholar] [CrossRef]
- Madhubalaji, C.K.; Rashmi, V.; Chauhan, V.S.; Shylaja, M.D.; Sarada, R. Improvement of vitamin B12 status with Spirulina supplementation in Wistar rats validated through functional and circulatory markers. J. Food Biochem. 2019, 43, e13038. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ramamoorthy, D.; Verma, D.K.; Kumar, A.; Kumar, N.; Kanak, K.R.; Marwein, B.M.; Mohan, K. Antioxidant and phytonutrient activities of Spirulina platensis. Energy Nexus 2022, 6, 16. [Google Scholar] [CrossRef]
- Romero-Ogawa, M.A.; Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Parra-Saldivar, R.; Ornelas-Soto, N. Extraction and purification of high-value metabolites from microalgae: Essential lipids, astaxanthin and phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar]
- Li, Y. The Bioactivities of Phycocyanobilin from Spirulina. J. Immunol. Res. 2022, 2022, 4008991. [Google Scholar] [CrossRef]
- Pereira, A.M.; Lisboa, C.R.; Santos, T.D.; Costa, J.A.V. Bioactive stability of microalgal protein hydrolysates under food processing and storage conditions. J. Food Sci. Technol. 2019, 56, 4543–4551. [Google Scholar] [CrossRef]
- Ragusa, I.; Nardone, G.N.; Zanatta, S.; Bertin, W.; Amadio, E. Spirulina for skin care: A bright blue future. Cosmetics 2021, 8, 7. [Google Scholar] [CrossRef]
- Souza, C.; Campos, P.M.B.G.M. Development and photoprotective effect of a sunscreen containing the antioxidants Spirulina and dimethylmethoxy chromanol on sun-induced skin damage. Eur. J. Pharm. Sci. 2017, 15, 52–64. [Google Scholar] [CrossRef]
- Habschied, K.; Živković, A.; Krstanović, V.; Mastanjević, K. Functional Beer—A Review on Possibilities. Beverages 2020, 6, 51. [Google Scholar] [CrossRef]
- Iannone, M.; Ovidi, E.; Vitalini, S.; Masci, V.L.; Marianelli, A.; Iriti, M.; Tiezzi, A.; Garzoli, S. From hops to craft beers: Production process, VOCs profile characterization, total polyphenol and flavonoid content determination and antioxidant activity evaluation. Processes 2022, 10, 517. [Google Scholar] [CrossRef]
- Nezi, P.; Cicaloni, V.; Tinti, L.; Salvini, L.; Iannone, M.; Vitalini, S.; Garzoli, S. Metabolomic and Proteomic Profile of Dried Hop Inflorescences (Humulus lupulus L. cv. Chinook and cv. Cascade) by SPME-GC-MS and UPLC-MS-MS. Separations 2022, 9, 204. [Google Scholar] [CrossRef]
- Aprea, E.; Biasioli, F.; Märk, T.D.; Gasperi, F. PTR-MS study of esters in water and water/ethanol solutions: Fragmentation patterns and partition coefficients. Int. J. Mass Spectrom. 2007, 262, 114–121. [Google Scholar] [CrossRef]
- Capozzi, V.; Makhoul, S.; Aprea, E.; Romano, A.; Cappellin, L.; Jimena, A.S.; Biasioli, F. PTR-MS characterization of VOCs associated with commercial aromatic bakery yeasts of wine and beer origin. Molecules 2016, 21, 483. [Google Scholar] [CrossRef] [PubMed]
- Boscaini, E.; Mikoviny, T.; Wisthaler, A.; von Hartungen, E.; Märk, T.D. Characterization of wine with PTR-MS. Int. J. Mass Spectrom. 2004, 239, 215–219. [Google Scholar] [CrossRef]
- Vitalone, A.; Di Sotto, A.; Mammola, C.L.; Heyn, R.; Miglietta, S.; Mariani, P.; Sciubba, F.; Passarelli, F.; Nativio, P.; Mazzanti, G. Phytochemical analysis and effects on ingestive behaviour of a Caralluma fimbriata extract. Food Chem. Toxicol. 2017, 108, 63–73. [Google Scholar] [CrossRef]
- Di Sotto, A.; Locatelli, M.; Macone, A.; Toniolo, C.; Cesa, S.; Carradori, S.; Eufemi, M.; Mazzanti, G.; Di Giacomo, S. Hypoglycemic, antiglycation, and cytoprotective properties of a phenol-rich extract from waste peel of Punica granatum L. var. Dente di Cavallo DC2. Molecules 2019, 27, 3103. [Google Scholar] [CrossRef]
- Di Sotto, A.; Durazzi, F.; Sarpietro, M.G.; Mazzanti, G. Antimutagenic and antioxidant activities of some bioflavours from wine. Food Chem. Toxicol. 2013, 60, 141–146. [Google Scholar] [CrossRef]
- Maslanka, R.; Kwolek-Mirek, M.; Zadrag-Tecza, R. Autofluorescence of yeast Saccharomyces cerevisiae cells caused by glucose metabolism products and its methodological implications. J. Microbiol. 2018, 146, 55–60. [Google Scholar] [CrossRef]
- Grubman, S.A.; Perrone, R.D.; Lee, D.W.; Murray, S.L.; Rogers, L.C.; Wolkoff, L.I.; Mulberg, A.E.; Cherington, V.; Jefferson, D.M. Regulation of intracellular pH by immortalized human intrahepatic biliary epithelial cell lines. Am. J. Physiol. 1994, 266, 1060–1070. [Google Scholar] [CrossRef]
- Di Sotto, A.; Di Giacomo, S.; Rubini, E.; Macone, A.; Gulli, M.; Mammola, C.L.; Eufemi, M.; Mancinelli, R.; Mazzanti, G. Modulation of stat3 signaling, cell redox defenses and cell cycle checkpoints by β-caryophyllene in cholangiocarcinoma cells: Possible mechanisms accounting for doxorubicin chemosensitization and chemoprevention. Cells 2020, 9, 858. [Google Scholar] [CrossRef]
- ISO 10993-5:2009; Biological Evaluation of Medical Devices Part 5: Tests for Invitro Cytotoxicity, 2nd ed. International Organization for Standardization/ANSI: Geneva, Switzerland, 2009.
- Nguyen, T.; Nioi, P.; Pickett, C.B. the nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef]
- Lermusieau, G.; Bulens, M.; Collin, S. Use of GC–olfactometry to identify the hop aromatic compounds in beer. J. Agric. Food Chem. 2001, 49, 3867–3874. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, S.; Hong, H.; Wang, K.; Yang, Q.; Gahruie, H.H.; Zhuang, S.; Li, Y.; Liang, Y.; Tan, Y.; Luo, Y. Ultrasonic-assisted food-grade solvent extraction of high-value added compounds from microalgae Spirulina platensis and evaluation of their antioxidant and antibacterial properties. Algal Res. 2021, 60, 102493. [Google Scholar] [CrossRef]
- Sandmann, M.; Beisler, N. Integration of Arthrospira platensis (spirulina) into the brewing process to develop new beers with unique sensory properties. Front. Sustain. Food Syst. 2022, 6, 918772. [Google Scholar] [CrossRef]
- Saerens, S.; Thevelein, J.; Delvaux, F. Ethyl ester production during brewery fermentation, a review. Cerevisia 2008, 33, 82–90. [Google Scholar]
- Serban, M.-C.; Sahebkar, A.; Dragan, S.; Stoichescu-Hogea, G.; Ursoniu, S.; Andrica, F.; Banach, M. A systematic review and meta-analysis of the impact of Spirulina supplementation on plasma lipid concentrations. Clin. Nutr. 2015, 35, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Finamore, A.; Palmery, M.; Bensehaila, S.; Peluso, I. Antioxidant, immunomodulating, and microbial-modulating activities of the sustainable and ecofriendly spirulina. Oxidative Med. Cell. Longev. 2017, 2017, 3247528. [Google Scholar] [CrossRef]
- Asghari, A.; Fazilati, M.; Latifi, A.M.; Salavati, H.; Choopani, A. A review on antioxidant properties of Spirulina. J. Appl. Biotechnol. Rep. 2016, 3, 345–351. [Google Scholar]
- Oriquat, G.A.; Ali, M.A.; Mahmoud, S.A.; Eid, R.M.; Hassan, R.; Kamel, M.A. Improving hepatic mitochondrial biogenesis as a postulated mechanism for the antidiabetic effect of Spirulina platensis in comparison with metformin. Appl. Physiol. Nutr. Metab. 2019, 44, 357–364. [Google Scholar] [CrossRef]
- El-Baky, H.; El Baz, F.K.; El-Baroty, G.S. Production of phenolic compounds from Spirulina maxima microalgae and its protective effects in vitro toward hepatotoxicity model. Afr. J. Pharm. Pharmacol. 2009, 3, 133–139. [Google Scholar]
- Papalia, T.; Sidari, R.; Panuccio, M.R. Impact of different storage methods on bioactive compounds in Arthrospira platensis biomass. Molecules 2019, 24, 2810. [Google Scholar] [CrossRef]
- Seghiri, R.; Kharbach, M.; Essamri, A. Functional composition, nutritional properties, and biological activities of Moroccan Spirulina microalga. J. Food Qual. 2019, 2019, 3707219. [Google Scholar] [CrossRef]
- Wannenmacher, J.; Gastl, M.; Becker, T. Phenolic substances in beer: Structural diversity, reactive potential and relevance for brewing process and beer quality. Compr. Rev. Food Sci. Food Saf. 2018, 17, 953–988. [Google Scholar] [CrossRef] [PubMed]
- Habschied, K.; Lončarić, A.; Mastanjević, K. Screening of polyphenols and antioxidative activity in industrial beers. Foods 2020, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.C., Jr.; Nicola, P.D.M.; Viegas, O.; Santos, M.C.; Faria, M.A.; Ferreira, I.M.P.L.V.O. Bioactive properties and phenolic composition of wood-aged beers: Influence of oak origin and the use of pale and dark malts. Foods 2023, 12, 1237. [Google Scholar] [CrossRef] [PubMed]
- Tefera, M.; Ayele, D. Investigation of total polyphenol, antioxidant activity, and levels of metals in Ethiopian commercial beers. Cogent Chem. 2020, 6, 1824336. [Google Scholar] [CrossRef]
- Tassoult, M.; Kati, D.E.; Fernández-Prior, M.Á.; Bermúdez-Oria, A.; Fernandez-Bolanos, J.; Rodríguez-Gutiérrez, G. Antioxidant capacity and phenolic and sugar profiles of date fruits extracts from six different algerian cultivars as influenced by ripening stages and extraction systems. Foods 2021, 10, 503. [Google Scholar] [CrossRef]
- Dhibi, M.; Mechri, B.; Brahmi, F.; Skhiri, F.; Alsaif, M.A.; Hammami, M.M. Fatty acid profiles, antioxidant compounds and antiradical properties of Pinus halepensis Mill. cones and seeds. J. Sci. Food Agric. 2012, 92, 1702–1708. [Google Scholar] [CrossRef]
- Sheraz, M.A.; Kazi, S.H.; Ahmed, S.; Anwar, Z.; Ahmad, I. Photo, thermal and chemical degradation of riboflavin. J. Org. Chem. 2014, 10, 1999. [Google Scholar] [CrossRef]
- Alonso-Andrés, P.; Martín, M.; Albasanz, J.É.L. Modulation of adenosine receptors and antioxidative effect of beer extracts in in vitro models. Nutrients 2019, 11, 1258. [Google Scholar] [CrossRef]
- Piovan, A.; Battaglia, J.; Filippini, R.; Costa, V.D.; Facci, L.; Argentini, C.; Pagetta, A.; Giusti, P.; Zusso, M. Pre- and Early Post-treatment with Arthrospira platensis (Spirulina) extract impedes lipopolysaccharide-triggered neuroinflammation in microglia. Front. Pharmacol. 2021, 9, 724993. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | COMPONENT 1 | LRI 2 | LRI 3 | Bc (%) | Bs (%) |
|---|---|---|---|---|---|
| 1 | ethyl acetate | 601 | 599 | 5.0 ± 0.02 | 0.5 ± 0.02 |
| 2 | 1-butanol, 3-methyl | 744 | 740 | 11.2 ± 0.03 | 8.9 ± 0.03 |
| 3 | 1-butanol, 2-methyl | 748 | 744 | 8.4 ± 0.02 | 8.0 ± 0.02 |
| 4 | propanal, 2-methyl,-oxime | 795 | 797 | - | 10.5 ± 0.04 |
| 5 | isoamyl acetate | 861 | 859 | 2.8 ± 0.02 | 0.5 ± 0.01 |
| 6 | 1H-1,2,4-triazole | 877 | 869 | - | 2.4 ± 0.02 |
| 7 | 2-methyl butyl acetate | 882 | 879 | 0.8 ± 0.01 | - |
| 8 | ethyl caproate | 998 | 996 | 2.4 ± 0.02 | 2.2 ± 0.02 |
| 9 | ethyl caprylate | 1183 | 1181 | 33.5 ± 0.06 | 32.9 ± 0.06 |
| 10 | phenyl ethyl acetate | 1233 | 1229 | 2.3 ± 0.02 | 1.6 ± 0.02 |
| 11 | ethyl caprate | 1385 | 1382 | 28.5 ± 0.05 | 26.9 ± 0.05 |
| 12 | phenyl ethyl butyrate | 1445 | 1440 | 0.8 ± 0.01 | 0.5 ± 0.01 |
| 13 | ethyl laurate | 1590 | 1581 | 4.3 ± 0.02 | 3.4 ± 0.02 |
| SUM | 100.0 | 98.3 |
| N° | COMPONENT | Spirulina (%) |
|---|---|---|
| CARBOXYLIC ACIDS | ||
| 1 | lactic acid | 64.7 |
| 2 | malonic acid | 0.3 |
| 3 | succinic acid | 4.2 |
| 4 | L-idopyranuronic acid | 0.2 |
| FATTY ACIDS | ||
| 5 | palmitic acid | 2.1 |
| 6 | stearic acid | 0.5 |
| SUGARS | ||
| 7 | L-rahmnose | 2.2 |
| 8 | D-xylose | 2.3 |
| 9 | arabinofuranose | 2.3 |
| 10 | D-lyxofuranose | 1.5 |
| 11 | 3-α-mannobiose | 1.8 |
| 12 | D-galactofuranoside | 0.2 |
| SUGAR ACIDS | ||
| 13 | glyceric acid | 0.6 |
| SUGAR ALCOHOLS | ||
| 14 | glycerol | 2.9 |
| TERPENES | ||
| 15 | phytol | 0.4 |
| OTHERS | ||
| 16 | 2,3-butanediol | 0.2 |
| 17 | hexadecane | 8.0 |
| 18 | 2-pentanol | 0.3 |
| 19 | cyclohexanol, trans- | 0.8 |
| 20 | pulchelloside I | 0.9 |
| M/Z | Chemical Formula | Beer | Beer with Spirulina | ||
|---|---|---|---|---|---|
| Average | SD | Average | SD | ||
| 27 | C2H3+ | 494.70 | 93.99 | 227.13 | 53.45 |
| 31 | CH3O+ | 5.97 | 1.63 | 3.60 | 0.54 |
| 41 | C3H5+ | 96.18 | 14.99 | 78.06 | 13.04 |
| 43 | C2H3O+ | 1074.06 | 134.61 | 693.08 | 120.14 |
| 43 | C3H7+ | 129.27 | 27.51 | 74.84 | 19.09 |
| 45 | C2H5O+ | 44.68 | 4.04 | 20.88 | 6.12 |
| 57 | C4H9+ | 120.94 | 18.81 | 151.64 | 20.31 |
| 59 | C3H7O+ | 25.28 | 7.09 | 27.67 | 7.44 |
| 61 | C2H5O2+ | 31.97 | 7.52 | 46.82 | 11.84 |
| 63 | C2H7O2+ | 12.57 | 1.01 | 18.96 | 3.21 |
| 65 | C5H5+ | 17.29 | 2.63 | 5.56 | 2.88 |
| 67 | C5H7+ | 5.80 | 1.51 | 1.87 | 0.29 |
| 69 | C5H9+ | 6.04 | 1.32 | 3.80 | 3.15 |
| 71 | C5H11+ | 61.32 | 3.91 | 70.79 | 13.10 |
| 73 | C4H9O+ | 13.05 | 1.88 | 10.57 | 3.77 |
| 75 | C4H11O+ | 105.89 | 14.09 | 54.74 | 6.61 |
| 77 | C6H5+ | 4.12 | 2.39 | 1.69 | 0.34 |
| 79 | C6H7+ | 2.33 | 0.79 | 1.95 | 0.32 |
| 85 | C6H9O+ | 5.91 | 0.85 | 2.27 | 0.93 |
| 87 | C5H11O+ | 14.61 | 6.60 | 9.50 | 1.47 |
| 89 | C4H9O2+ | 316.65 | 18.36 | 127.06 | 10.21 |
| 95 | C7H11+ | 22.79 | 3.20 | 7.58 | 1.76 |
| 101 | C6H13O+ | 28.14 | 6.68 | 15.93 | 3.21 |
| 103 | C5H11O2+ | 38.78 | 6.36 | 18.06 | 5.68 |
| 105 | C8H9+ | 12.17 | 1.19 | 8.60 | 3.07 |
| 107 | C8H11+ | 16.50 | 3.79 | 6.37 | 1.13 |
| 109 | C8H13+ | 3.68 | 0.93 | 1.83 | 0.38 |
| 113 | C6H9O2+ | 4.63 | 1.18 | 0.00 | 0.00 |
| 115 | C6H11O2+ | 8.14 | 2.62 | 1.90 | 0.71 |
| 117 | C6H13O2+ | 27.57 | 8.22 | 9.47 | 3.00 |
| 131 | C7H15O2+ | 10.15 | 3.32 | 3.74 | 1.02 |
| 135 | C10H15+ | 10.11 | 0.83 | 5.44 | 0.44 |
| 145 | C8H17O2+ | 12.38 | 4.19 | 10.17 | 4.11 |
| 147 | C9H7O2+ | 3.89 | 0.85 | 8.13 | 0.73 |
| 161 | C10H9O2+ | 5.82 | 1.12 | 1.92 | 0.25 |
| 173 | C10H21O2+ | 5.51 | 1.00 | 3.51 | 1.52 |
| 177 | C9H21O3+ | 8.34 | 2.44 | 0.00 | 0.00 |
| 189 | C8H13O5+ | 14.13 | 2.31 | 2.77 | 0.35 |
| 201 | C12H25O2+ | 1.60 | 0.38 | 1.39 | 0.20 |
| 203 | C12H11O3+ | 3.44 | 1.14 | 0.00 | 0.00 |
| Total VOC emission | 2636.81 | 1928.87 | |||
| Total number of signals | 40 | 37 | |||
| Sample | DPPH Scavenging Activity | ABTS Scavenging Activity | |
|---|---|---|---|
| Beer | IC50 (CL) µL/mL | 57.7 (43.1–77.3) | 5.4 (1.8–9.3) |
| Beer + spirulina 0.25% w/v | 49.9 (25.2–95.0) | 1.9 (0.7–4.9) # | |
| Spirulina | IC50 (CL) µg/mL | ne | ne |
| Trolox | 7.0 (5.0–9.7) | 5.8 (4.2–8.4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taiti, C.; Stefano, G.; Percaccio, E.; Di Giacomo, S.; Iannone, M.; Marianelli, A.; Di Sotto, A.; Garzoli, S. Addition of Spirulina to Craft Beer: Evaluation of the Effects on Volatile Flavor Profile and Cytoprotective Properties. Antioxidants 2023, 12, 1021. https://doi.org/10.3390/antiox12051021
Taiti C, Stefano G, Percaccio E, Di Giacomo S, Iannone M, Marianelli A, Di Sotto A, Garzoli S. Addition of Spirulina to Craft Beer: Evaluation of the Effects on Volatile Flavor Profile and Cytoprotective Properties. Antioxidants. 2023; 12(5):1021. https://doi.org/10.3390/antiox12051021
Chicago/Turabian StyleTaiti, Cosimo, Giovanni Stefano, Ester Percaccio, Silvia Di Giacomo, Matteo Iannone, Andrea Marianelli, Antonella Di Sotto, and Stefania Garzoli. 2023. "Addition of Spirulina to Craft Beer: Evaluation of the Effects on Volatile Flavor Profile and Cytoprotective Properties" Antioxidants 12, no. 5: 1021. https://doi.org/10.3390/antiox12051021
APA StyleTaiti, C., Stefano, G., Percaccio, E., Di Giacomo, S., Iannone, M., Marianelli, A., Di Sotto, A., & Garzoli, S. (2023). Addition of Spirulina to Craft Beer: Evaluation of the Effects on Volatile Flavor Profile and Cytoprotective Properties. Antioxidants, 12(5), 1021. https://doi.org/10.3390/antiox12051021