
Studies on the Effect of Oxidation on Bioactivity of Phenolics and Wine Lees Extracts



Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Wine Lees Sample Preparation
2.2.1. Preparation of Flavonoid Model
2.2.2. Wine Lees
2.2.3. Oxidation of Flavonoid Model and Real Lees Samples
2.3. Determination of Total Phenol Content (TPC)
2.4. Determination of Total Tannin Content (TTC)
2.5. Determination of Phenolic Profile
2.6. Determination of the Mean Degree of Polymerization (mDP)
2.7. Determination of Antioxidant Activities
2.8. Determination of Antimicrobial Activity
2.9. Statistical Analysis
3. Results and Discussion
3.1. Effect of Oxidation on Total Phenolic Content (TPC) and Total Tannin Content (TTC)
3.2. Effect of Oxidation on Phenolic Compounds and mDP
3.2.1. Phenolic Compounds
3.2.2. Mean Degree of Polymerization (mDP)
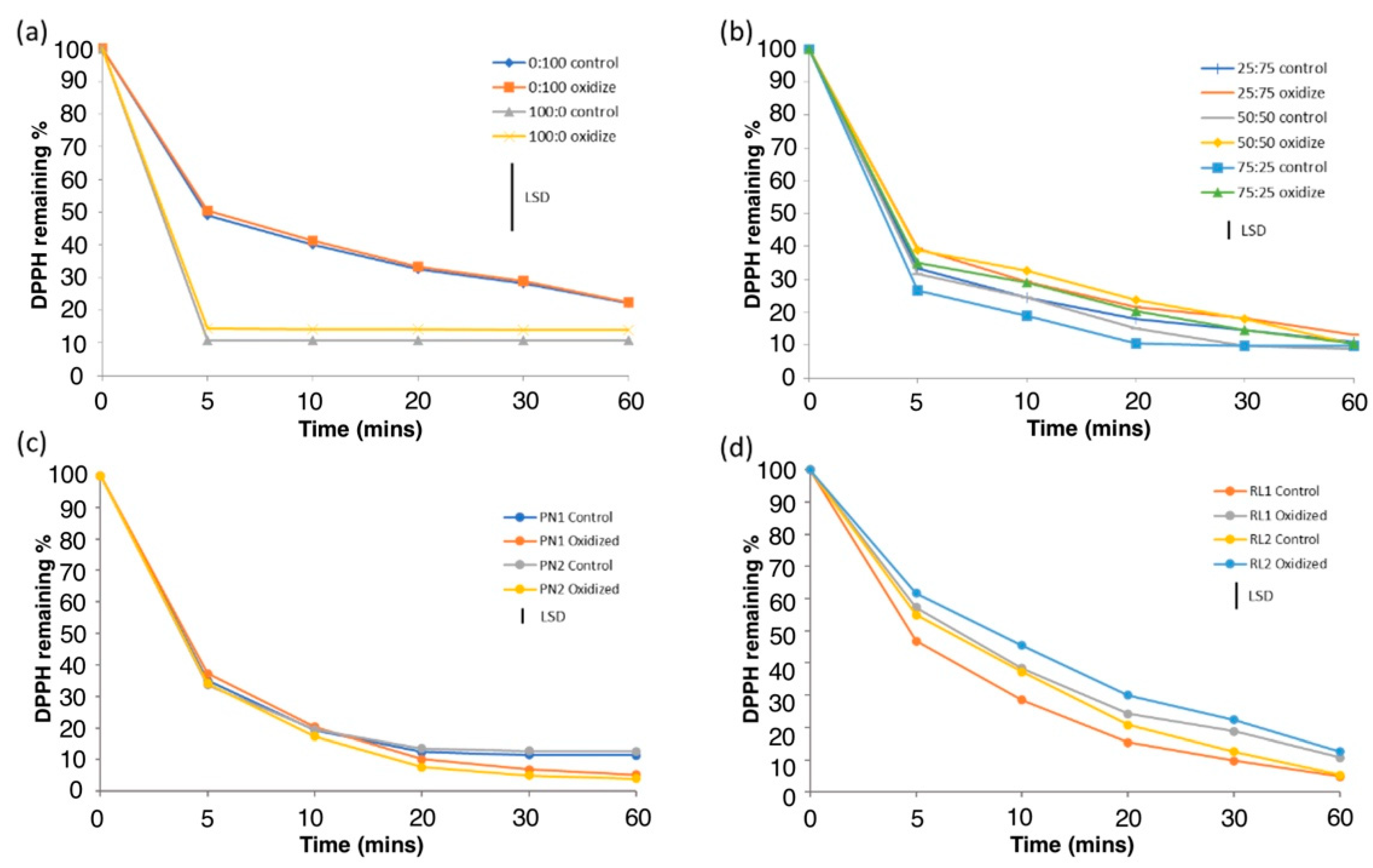
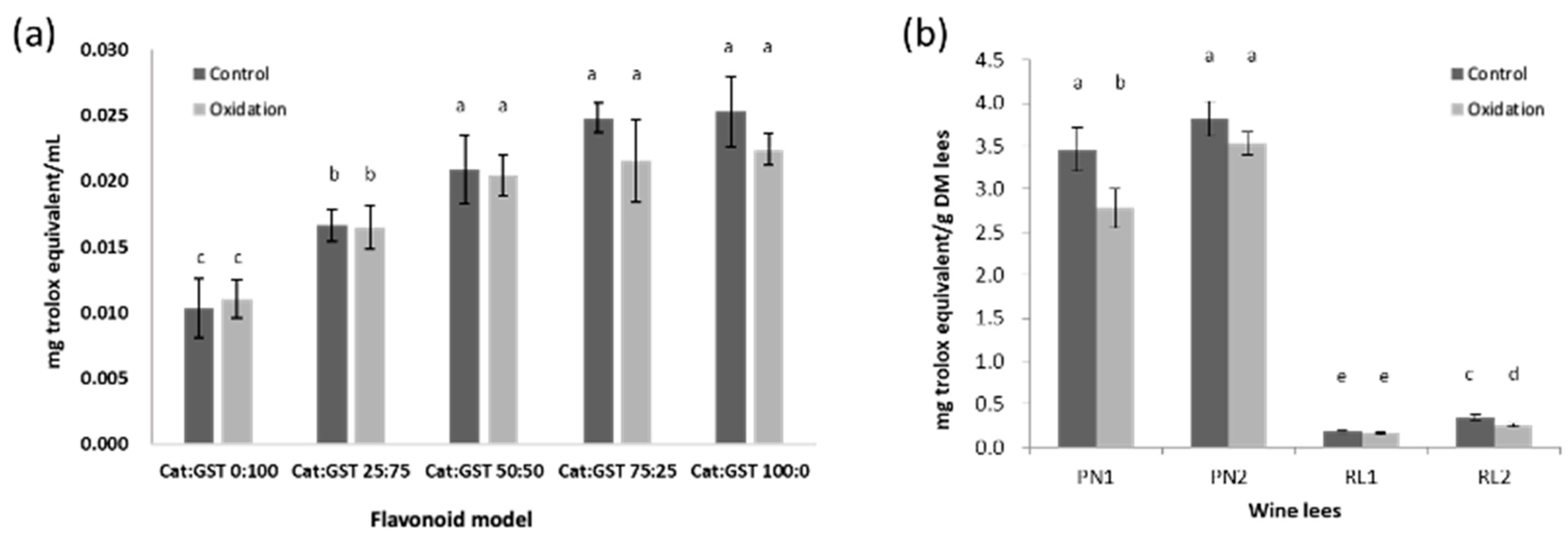
3.3. Effect of Oxidation on Antioxidant Activities
3.4. Antibacterial and Antifungal Activities of Flavonoid Model Samples
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Juice Type | Vessel | Yeast (N/C) | Fermentation Temp (°C) | Maceration | Vinification | Lees Collection |
|---|---|---|---|---|---|---|---|
| PN1 | FR | PV | N | 34 | Pre and Post | PO, P | Pre-MLF |
| PN2 | FR | PV | N | 35 | Pre and Post | PO, P | Pre-MLF |
| RL1 | - | Tank | C | 17 | - | - | Post-ALC |
| RL2 | - | Tank | C | 15 | - | - | Post-ALC |
Appendix B

References
- Bekhit, A.E.-D.A.; Cheng, V.J.; McConnell, M.; Zhao, J.H.; Sedcole, R.; Harrison, R. Antioxidant Activities, Sensory and Anti-Influenza Activity of Grape Skin Tea Infusion. Food Chem. 2011, 129, 837–845. [Google Scholar] [CrossRef]
- Cheng, V.J. Evaluation of the Bioactivity of New Zealand Wine By-Products. Ph.D. Thesis, University of Otago, Dunedin, New Zealand, 2011. [Google Scholar]
- Lachman, J.; Hejtmánková, A.; Hejtmánková, K.; Horníčková, Š.; Pivec, V.; Skala, O.; Dědina, M.; Přibyl, J. Towards Complex Utilisation of Winemaking Residues: Characterisation of Grape Seeds by Total Phenols, Tocols and Essential Elements Content as a by-Product of Winemaking. Ind. Crops Prod. 2013, 49, 445–453. [Google Scholar] [CrossRef]
- Melo, P.S.; Massarioli, A.P.; Denny, C.; dos Santos, L.F.; Franchin, M.; Pereira, G.E.; de Souza Vieira, T.M.; Rosalen, P.L.; de Alencar, S.M. Winery By-Products: Extraction Optimization, Phenolic Composition and Cytotoxic Evaluation to Act as a New Source of Scavenging of Reactive Oxygen Species. Food Chem. 2015, 181, 160–169. [Google Scholar] [CrossRef]
- Silva, A.; Silva, V.; Igrejas, G.; Aires, A.; Falco, V.; Valentão, P.; Poeta, P. Phenolic Compounds Classification and Their Distribution in Winemaking By-Products. Eur. Food Res. Technol. 2022, 249, 207–239. [Google Scholar] [CrossRef]
- Moro, K.I.B.; Bender, A.B.B.; da Silva, L.P.; Penna, N.G. Green Extraction Methods and Microencapsulation Technologies of Phenolic Compounds From Grape Pomace: A Review. Food Bioprocess Technol. 2021, 14, 1407–1431. [Google Scholar] [CrossRef]
- Katiki, L.M.; Ferreira, J.F.; Gonzalez, J.M.; Zajac, A.M.; Lindsay, D.S.; Chagas, A.C.S.; Amarante, A.F. Anthelmintic Effect of Plant Extracts Containing Condensed and Hydrolyzable Tannins on Caenorhabditis Elegans, and Their Antioxidant Capacity. Vet. Parasitol. 2013, 192, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.-C.; Lin, Y.-M.; Wei, S.-D.; Tam, N.F. Structural Diversity and Antioxidant Activity of Condensed Tannins Fractionated from Mangosteen Pericarp. Food Chem. 2011, 129, 1710–1720. [Google Scholar] [CrossRef]
- Gu, H.-F.; Li, C.-M.; Xu, Y.; Hu, W.; Chen, M.; Wan, Q. Structural Features and Antioxidant Activity of Tannin from Persimmon Pulp. Food Res. Int. 2008, 41, 208–217. [Google Scholar] [CrossRef]
- Gris, E.F.; Mattivi, F.; Ferreira, E.A.; Vrhovsek, U.; Pedrosa, R.C.; Bordignon-Luiz, M.T. Proanthocyanidin Profile and Antioxidant Capacity of Brazilian Vitis Vinifera Red Wines. Food Chem. 2011, 126, 213–220. [Google Scholar] [CrossRef]
- Jackson, R.S. Chemical Constituents of Grapes and Wine. In Wine Science Principles and Applications, 3rd ed.; Jackson, R.S., Ed.; Academic Press: Cambridge, MA, USA, 2008; pp. 270–331. [Google Scholar] [CrossRef]
- Zhijing, Y.; Shavandi, A.; Harrison, R.; Bekhit, A.E.-D.A. Characterization of Phenolic Compounds in Wine Lees. Antioxidants 2018, 7, 48. [Google Scholar] [CrossRef]
- Ye, Z.; Qin, Y.; Harrison, R.; Hider, R.; Bekhit, A.E.-D.A. Characterization of Bioactive Compounds in Lees from New Zealand Wines with Different Vinification Backgrounds. Antioxidants 2022, 11, 2335. [Google Scholar] [CrossRef] [PubMed]
- Jongberg, S.; Lund, M.N.; Østdal, H.; Skibsted, L.H. Phenolic Antioxidant Scavenging of Myosin Radicals Generated by Hypervalent Myoglobin. J. Agric. Food Chem. 2012, 60, 12020–12028. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, M.D.; Dambergs, R.G.; Herderich, M.J.; Smith, P.A. High Throughput Analysis of Red Wine and Grape Phenolics Adaptation and Validation of Methyl Cellulose Precipitable Tannin Assay and Modified Somers Color Assay to a Rapid 96 Well Plate Format. J. Agric. Food Chem. 2007, 55, 4651–4657. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Oxidants and Antioxidants Part A; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Sarneckis, C.J.; Dambergs, R.G.; Jones, P.; Mercurio, M.; Herderich, M.J.; Smith, P.A. Quantification of Condensed Tannins by Precipitation with Methyl Cellulose: Development and Validation of an Optimised Tool for Grape and Wine Analysis. Aust. J. Grape Wine Res. 2006, 12, 39–49. [Google Scholar] [CrossRef]
- Kemp, B.S.; Harrison, R.; Creasy, G.L. Effect of Mechanical Leaf Removal and Its Timing on Flavan-3-ol Composition and Concentrations in Vitis Vinifera L. Cv. Pinot Noir Wine. Aust. J. Grape Wine Res. 2011, 17, 270–279. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Jones, G.P. Analysis of Proanthocyanidin Cleavage Products Following Acid-Catalysis in the Presence of Excess Phloroglucinol. J. Agric. Food Chem. 2001, 49, 1740–1746. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A Procedure to Measure the Antiradical Efficiency of Polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Woods, G.L. Antibacterial Susceptibility Tests: Dilution and Disk Diffusion Methods. In Manual of Clinical Microbiology, 8th ed.; Murray, P.R., Baron, E.J., Jorgenson, J.H., Pfaller, M.A., Yolken, R.H., Eds.; ASM Press: Washington, DC, USA, 1995; pp. 1327–1341. [Google Scholar]
- Blainski, A.; Lopes, G.C.; de Mello, J.C.P. Application and Analysis of the Folin Ciocalteu Method for the Determination of the Total Phenolic Content from Limonium Brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef]
- Nagarajan, S.; Nagarajan, R.; Braunhut, S.J.; Bruno, F.; McIntosh, D.; Samuelson, L.; Kumar, J. Biocatalytically Oligomerized Epicatechin with Potent and Specific Anti-Proliferative Activity for Human Breast Cancer Cells. Molecules 2008, 13, 2704–2716. [Google Scholar] [CrossRef]
- Hosny, M.; Rosazza, J.P. Novel Oxidations of (+)-Catechin by Horseradish Peroxidase and Laccase. J. Agric. Food Chem. 2002, 50, 5539–5545. [Google Scholar] [CrossRef] [PubMed]
- Zanchi, D.; Guyot, S.; Konarev, P.; Baron, A.; Svergun, D. Physical Properties of Tannins Aged by Oxidation. 2007. Available online: http://hasyweb.desy.de/science/annual_reports/2007_report/part2/contrib/73/20261.pdf (accessed on 12 October 2022).
- Kusano, R.; Tanaka, T.; Matsuo, Y.; Kouno, I. Structures of Epicatechin Gallate Trimer and Tetramer Produced by Enzymatic Oxidation. Chem. Pharm. Bull. 2007, 55, 1768–1772. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Matsuo, Y.; Kouno, I. A Novel Black Tea Pigment and Two New Oxidation Products of Epigallocatechin-3-O-Gallate. J. Agric. Food Chem. 2005, 53, 7571–7578. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Tian, S.; Meng, X.; Stark, R.E.; Rosen, R.T.; Yang, C.S.; Ho, C.-T. Theadibenzotropolone A, a New Type Pigment from Enzymatic Oxidation of (-)-Epicatechin and (-)-Epigallocatechin Gallate and Characterized from Black Tea Using LC/MS/MS. Tetrahedron Lett. 2002, 43, 7129–7133. [Google Scholar] [CrossRef]
- Vernhet, A.; Dubascoux, S.; Cabane, B.; Fulcrand, H.; Dubreucq, E.; Poncet-Legrand, C. Characterization of Oxidized Tannins: Comparison of Depolymerization Methods, Asymmetric Flow Field-Flow Fractionation and Small-Angle X-Ray Scattering. Anal. Bioanal. Chem. 2011, 401, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Mouls, L.; Fulcrand, H. UPLC-ESI-MS Study of the Oxidation Markers Released from Tannin Depolymerization: Toward a Better Characterization of the Tannin Evolution over Food and Beverage Processing. J. Mass Spectrom. 2012, 47, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Herderich, M.J.; Smith, P.A. Analysis of Grape and Wine Tannins: Methods, Applications and Challenges. Aust. J. Grape Wine Res. 2005, 11, 205–214. [Google Scholar] [CrossRef]
- Singleton, V.L.; Trousdale, E.K. Anthocyanin-Tannin Interactions Explaining Differences in Polymeric Phenols between White and Red Wines. Am. J. Enol. Vitic. 1992, 43, 63–70. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-Antioxidant Activity Relationships of Flavonoids and Phenolic Acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Zhou, H.-C.; Tam, N.F.; Lin, Y.-M.; Ding, Z.-H.; Chai, W.-M.; Wei, S.-D. Relationships between Degree of Polymerization and Antioxidant Activities: A Study on Proanthocyanidins from the Leaves of a Medicinal Mangrove Plant Ceriops Tagal. PLoS ONE 2014, 9, e107606. [Google Scholar] [CrossRef]
- Gañan, M.; Martínez-Rodríguez, A.J.; Carrascosa, A.V. Antimicrobial Activity of Phenolic Compounds of Wine against Campylobacter Jejuni. Food Control 2009, 20, 739–742. [Google Scholar] [CrossRef]
- Daglia, M.; Papetti, A.; Grisoli, P.; Aceti, C.; Dacarro, C.; Gazzani, G. Antibacterial Activity of Red and White Wine against Oral Streptococci. J. Agric. Food Chem. 2007, 55, 5038–5042. [Google Scholar] [CrossRef]
- Aziz, N.H.; Farag, S.E.; Mousa, L.A.; Abo-Zaid, M.A. Comparative Antibacterial and Antifungal Effects of Some Phenolic Compounds. Microbios 1998, 93, 43–54. [Google Scholar]
- Chan, M.M.-Y. Antimicrobial Effect of Resveratrol on Dermatophytes and Bacterial Pathogens of the Skin. Biochem. Pharmacol. 2002, 63, 99–104. [Google Scholar] [CrossRef]
- Wen, A.; Delaquis, P.; Stanich, K.; Toivonen, P. Antilisterial Activity of Selected Phenolic Acids. Food Microbiol. 2003, 20, 305–311. [Google Scholar] [CrossRef]




| Catechin (mg) | GST (mg) | Cat:GST | |
|---|---|---|---|
| Control | 0 | 200 | 0:100 |
| 50 | 150 | 25:75 | |
| 100 | 100 | 50:50 | |
| 150 | 50 | 75:25 | |
| 200 | 0 | 100:0 | |
| Treatment | 0 | 200 | 0:100 |
| 50 | 150 | 25:75 | |
| 100 | 100 | 50:50 | |
| 150 | 50 | 75:25 | |
| 200 | 0 | 100:0 |
| Sample | Variety | Weight (g) | Solvent | Volume (mL) | w/v Ratio |
|---|---|---|---|---|---|
| PN1 | Pinot noir | 0.5000 | 50% acidified EtOH | 5 | 1:10 |
| PN2 | Pinot noir | 0.5000 | 50% acidified EtOH | 5 | 1:10 |
| RL1 | Riesling | 3.0000 | 50% acidified EtOH | 5 | 6:10 |
| RL2 | Riesling | 3.0000 | 50% acidified EtOH | 5 | 6:10 |
| Phenolic Compounds | Cat:GST 0:100 | Cat:GST 25:75 | Cat:GST 50:50 | Cat:GST 75:25 | Cat:GST 100:0 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Oxidized | Control | Oxidized | Control | Oxidized | Control | Oxidized | Control | Oxidized | |
| Non-flavonoids | ||||||||||
| Gallic acid | 11.92 ± 0.14 a | 8.90 ± 3.52 abc | 9.62 ± 0.36 b | 0.84 ± 0.30 e | 6.04 ± 0.04 c | 1.72 ± 0.32 d | 2.46 ± 0.92 d | - | - | - |
| Hydroxycinnamic acids | ||||||||||
| Caftaric acid | 2.44 ± 0.36 bc | 2.28 ± 0.64 bc | 2.72 ± 0.14 bc | 3.40 ± 0.94 b | 3.54 ± 0.64 bc | 3.68 ± 1.22 bc | 1.68 ± 0.36 d | 3.96 ± 0.24 a | 6.54 ± 2.82 a | 4.30 ± 0.10 b |
| p-coumaric acid | 0.04 ± 0.01 a | 0.04 ± 0.02 a | 0.06 ± 0.04 a | 0.04 ± 0.01 a | 0.12 ± 0.08 a | 0.10 ± 0.04 a | - | - | 0.04 ± 0.02 a | 0.02 ± 0.01 a |
| Hydroxybenzoric acid | - | - | - | - | - | 3.32 ± 0.01 a | - | 2.30 ± 0.14 b | - | - |
| Flavonoids | ||||||||||
| Flavan-3-ols | ||||||||||
| Catechin | 189.36 ± 7.22 g | 45.80 ± 2.06 h | 530.66 ± 16.78 e | 351.08 ± 4.42 f | 1085.14 ± 36.32 c | 852.28 ± 17.40 d | 1571.44 ± 30.86 b | 850.30 ± 17.40 d | 1879.42 ± 124.30 a | 835.40 ± 70.22 d |
| Epicatechin | 66.30 ± 0.58 a | 63.20 ± 2.40 a | 45.54 ± 0.38 b | 2.90 ± 0.18 d | 4.00 ± 0.06 c | 1.32 ± 0.28 e | 2.96 ± 1.20 d | 1.62 ± 0.14 e | - | - |
| Epicatechin gallate | - | - | - | - | - | - | - | - | - | - |
| Epigallocatechin | 0.38 ± 0.01 f | 0.36 ± 0.02 f | 4.08 ± 0.12 d | 2.70 ± 0.04 e | 8.34 ± 0.28 b | 6.54 ± 0.14 c | 12.08 ± 0.24 a | 6.54 ± 0.10 c | 14.44 ± 0.96 a | 6.42 ± 0.54 bc |
| Procyanidin A * | - | - | 7.70 ± 0.18 e | 8.14 ± 0.36 e | 21.02 ± 0.42 d | 47.76 ± 2.16 bc | 43.94 ± 0.90 c | 59.16 ± 0.82 b | 64.66 ± 0.42 a | 75.30 ± 5.58 a |
| Procyanidin B * | 9.68 ± 0.24 a | 9.54 ± 0.14 a | 5.94 ± 0.12 b | - | - | - | - | - | - | - |
| Procyanidin C * | 7.24 ± 0.06 a | 6.08 ± 0.14 b | 5.18 ± 0.30 c | - | - | - | - | - | - | - |
| Phenolic Compounds | PN1 | PN2 | RL1 | RL2 | ||||
|---|---|---|---|---|---|---|---|---|
| Control | Oxidized | Control | Oxidized | Control | Oxidized | Control | Oxidized | |
| Non-flavonoids | ||||||||
| Gallic acid | 1.64 ± 0.01 b | 0.07 ± 0.01 d | 2.03 ± 0.01 a | 0.43 ± 0.18 c | 0.13 ± 0.01 c | 0.02 ± 0.01 e | 0.10 ± 0.04 c | 0.07 ± 0.01 d |
| Hydroxycinnamic acids | ||||||||
| Caftaric acid | - | - | - | - | - | - | - | - |
| Cinnamic acid | - | - | - | - | 0.05 ± 0.01 a | 0.05 ± 0.01 a | - | - |
| p-coumaric acid | - | - | - | - | - | - | - | - |
| Hydroxybenzoric acid | - | - | - | - | - | - | - | - |
| Stillbenoids | ||||||||
| Resveratrol | 0.42 ± 0.01 b | 0.34 ± 0.01 c | 0.62 ± 0.01 a | 0.40 ± 0.01 b | 0.10 ± 0.01 e | 0.08 ± 0.01 e | 0.29 ± 0.01 d | 0.41 ± 0.01 b |
| Flavonoids | ||||||||
| Flavan-3-ols | ||||||||
| Catechin | 3.80 ± 1.50 c | 0.11 ± 0.04 g | 3.63 ± 0.09 c | 0.69 ± 0.04 e | 0.92 ± 0.06 d | 0.22 ± 0.02 f | 16.34 ± 0.49 a | 8.45 ± 1.48 b |
| Epicatechin | 1.70 ± 0.05 b | 0.28 ± 0.05 d | 2.33 ± 0.08 a | 0.48 ± 0.03 c | 2.43 ± 0.14 a | 0.40 ± 0.05 c | 0.17 ± 0.04 e | 0.19 ± 0.01 e |
| Epicatechin gallate | 0.76 ± 0.02 c | 0.29 ± 0.02 d | 0.83 ± 0.02 b | 0.29 ± 0.01 d | 0.32 ± 0.18 d | 0.13 ± 0.05 e | 1.57 ± 0.93 a | 2.59 ± 0.08 a |
| Epigallocatechin | 0.02 ± 0.01 c | - | 0.03 ± 0.01 c | - | - | - | 0.13 ± 0.01 a | 0.06 ± 0.01 b |
| Procyanidin A | 2.39 ± 0.01 d | 2.55 ± 0.14 d | 2.14 ± 0.09 e | 2.97 ± 0.15 c | 0.48 ± 0.08 b | 0.40 ± 0.02 b | 5.77 ± 1.27 a | 4.46 ± 0.14 a |
| Procyanidin B | 1.65 ± 0.02 a | 1.16 ± 0.04 a | 1.43 ± 0.03 a | 1.39 ± 0.02 a | - | - | 2.55 ± 1.92 a | 0.05 ± 0.04 b |
| Procyanidin C | 1.79 ± 0.17 b | 4.22 ± 0.10 a | 1.61 ± 0.02 b | 3.88 ± 0.23 a | - | - | - | - |
| Anthocyanins | ||||||||
| Cyanidin-3-glucoside | - | - | - | - | - | - | - | - |
| Delphinidin-3-glucoside | - | - | - | - | - | - | - | - |
| Malvidin-3-glucoside | 0.02 ± 0.01 b | - | 0.05 ± 0.01 a | - | - | - | - | - |
| Peonidin-3-glucoside | - | - | - | - | - | - | - | - |
| Petunidin-3-glucoside | - | - | - | - | - | - | - | - |
| Flavanols | ||||||||
| Quercetin | 1.80 ± 0.05 b | 0.97 ± 0.01 d | 2.38 ± 0.01 a | 1.50 ± 0.09 c | 0.77 ± 0.04 e | 0.21 ± 0.01 f | 1.10 ± 0.11 d | 1.26 ± 0.01 d |
| Quercetin methyl-glucoside | 1.16 ± 0.01 a | 1.03 ± 0.03 b | 0.79 ± 0.03 c | 0.92 ± 0.02 b | 0.42 ± 0.01 d | 0.20 ± 0.01 e | 1.01 ± 0.15 ab | 0.91 ± 0.01 b |
| Terminal Units (%) | Extension Units (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sample | mDP | Treatment | C | EC | ECG | EGC | C | EC | ECG | EGC |
| PN1 | 10.7 ± 0.5 a | Control | 64.40 | 31.59 | 4.01 | 0.00 | 6.89 | 59.54 | 3.09 | 30.48 |
| PN2 | 6.7 ± 0.4 c | Control | 58.63 | 37.52 | 3.85 | 0.00 | 7.21 | 58.10 | 3.00 | 31.69 |
| PN1 | 10.6 ± 0.9 a | Oxidized | 54.37 | 37.82 | 7.81 | 0.00 | 12.03 | 44.22 | 2.36 | 41.38 |
| PN2 | 6.8 ± 1.1 bc | Oxidized | 55.30 | 39.81 | 4.89 | 0.00 | 12.22 | 41.70 | 2.10 | 43.98 |
| Antibacterial/Antifungal Activity (MIC50) mg/mL | |||||||
|---|---|---|---|---|---|---|---|
| S. aureus 1 | E. coli 1 | C. albicans 2 | |||||
| Sample | (MIC) mg/mL | (MBC) mg/mL | (MIC) mg/mL | (MBC) mg/mL | (MIC) mg/mL | (MBC) mg/mL | |
| Control | 0:100 | 6.25 | 6.25 | 12.50 * | 12.50 * | 12.50 * | 12.50 * |
| 25:75 | 3.13 | 3.13 | 3.13 | 3.13 | 6.25 | 12.50 * | |
| 50:50 | 12.50 * | 12.50 | 6.25 | 6.25 | 6.25 | 12.50 * | |
| 75:25 | 3.13 | 3.13 | 3.13 | 3.13 | 6.25 | 12.50 * | |
| 100:0 | 6.25 | 6.25 | 1.56 | 3.13 | 12.5 * | 12.50 * | |
| Oxidized | 0:100 | 3.13 | 3.13 | 1.56 | 3.13 | 12.5 * | 12.50 * |
| 25:75 | 1.56 | 3.13 | 0.78 | 0.78 | 6.25 | 6.25 | |
| 50:50 | 3.13 | 3.13 | 0.39 | 0.78 | 6.25 | 12.50 * | |
| 75:25 | 1.56 | 3.13 | 0.39 | 0.78 | 6.25 | 6.25 | |
| 100:0 | 3.13 | 3.13 | 0.39 | 0.39 | 6.25 | 12.50 * | |
| Control | PN1 | 5.00 | 10.00 | 5.00 | 10.00 | 2.50 | 5.00 |
| PN2 | 5.00 | 10.00 | 5.00 | 5.00 | 5.00 | 10.00 | |
| RL1 | 10.00 | 20.00 | 10.00 | 20.00 | 10.00 | 20.00 | |
| RL2 | 5.00 | 10.00 | 5.00 | 10.00 | 5.00 | 10.00 | |
| Oxidized | PN1 | 5.00 | 10.00 | 5.00 | 10.00 | 2.50 | 10.00 |
| PN2 | 5.00 | 10.00 | 5.00 | 10.00 | 5.00 | 10.00 | |
| RL1 | 10.00 | 20.00 | 10.00 | 20.00 | 10.00 | 20.00 | |
| RL2 | 5.00 | 5.00 | 5.00 | 10.00 | 5.00 | 10.00 | |
| Antibacterial/antifungal activity (MIC)µg/mL | |||||||
| Antimicrobial Agent | S. aureus 1 | E. coli 1 | C. albicans 2 | ||||
| Ampicillin | 0.781 | 3.125 | |||||
| Amphotericin B | 6.25 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Z.; Shi, J.; Harrison, R.; Hider, R.; Bekhit, A.E.-D.A. Studies on the Effect of Oxidation on Bioactivity of Phenolics and Wine Lees Extracts. Antioxidants 2023, 12, 931. https://doi.org/10.3390/antiox12040931
Ye Z, Shi J, Harrison R, Hider R, Bekhit AE-DA. Studies on the Effect of Oxidation on Bioactivity of Phenolics and Wine Lees Extracts. Antioxidants. 2023; 12(4):931. https://doi.org/10.3390/antiox12040931
Chicago/Turabian StyleYe, Zhijing, Jinlin Shi, Roland Harrison, Richard Hider, and Alaa El-Din A. Bekhit. 2023. "Studies on the Effect of Oxidation on Bioactivity of Phenolics and Wine Lees Extracts" Antioxidants 12, no. 4: 931. https://doi.org/10.3390/antiox12040931
APA StyleYe, Z., Shi, J., Harrison, R., Hider, R., & Bekhit, A. E.-D. A. (2023). Studies on the Effect of Oxidation on Bioactivity of Phenolics and Wine Lees Extracts. Antioxidants, 12(4), 931. https://doi.org/10.3390/antiox12040931