
Melatonin Supplementation during the Late Gestational Stage Enhances Reproductive Performance of Sows by Regulating Fluid Shear Stress and Improving Placental Antioxidant Capacity

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Agents
2.2. Animals
2.3. Animal Study Design
2.4. Blood Sample Collection
2.5. Measurement of Piglet Weight after Birth and Placental Tissue Collection
2.6. Melatonin Detection
2.7. Detection of Hormone and Biochemical Indicators
2.8. Real-Time Quantitative PCR
2.9. Western Blotting
2.10. Immunofluorescence Detection of AANAT, MTNR1A and MTNR1B
2.11. RNA-Seq for Placenta Tissue
2.12. Statistical Analysis
3. Results
3.1. Effects of Melatonin Supplementation on Levels of Melatonin in Blood and Colostrum
3.2. Effects of Melatonin Supplementation on Reproductive Performance of Sows
3.3. Effects of Melatonin Supplementation on Blood Reproductive Hormone and Other Biochemical Parameters
3.4. Effects of Melatonin Supplementation on Melatonin Synthetic Enzyme AANAT and Melatonin Receptor Expression in Placental Tissue
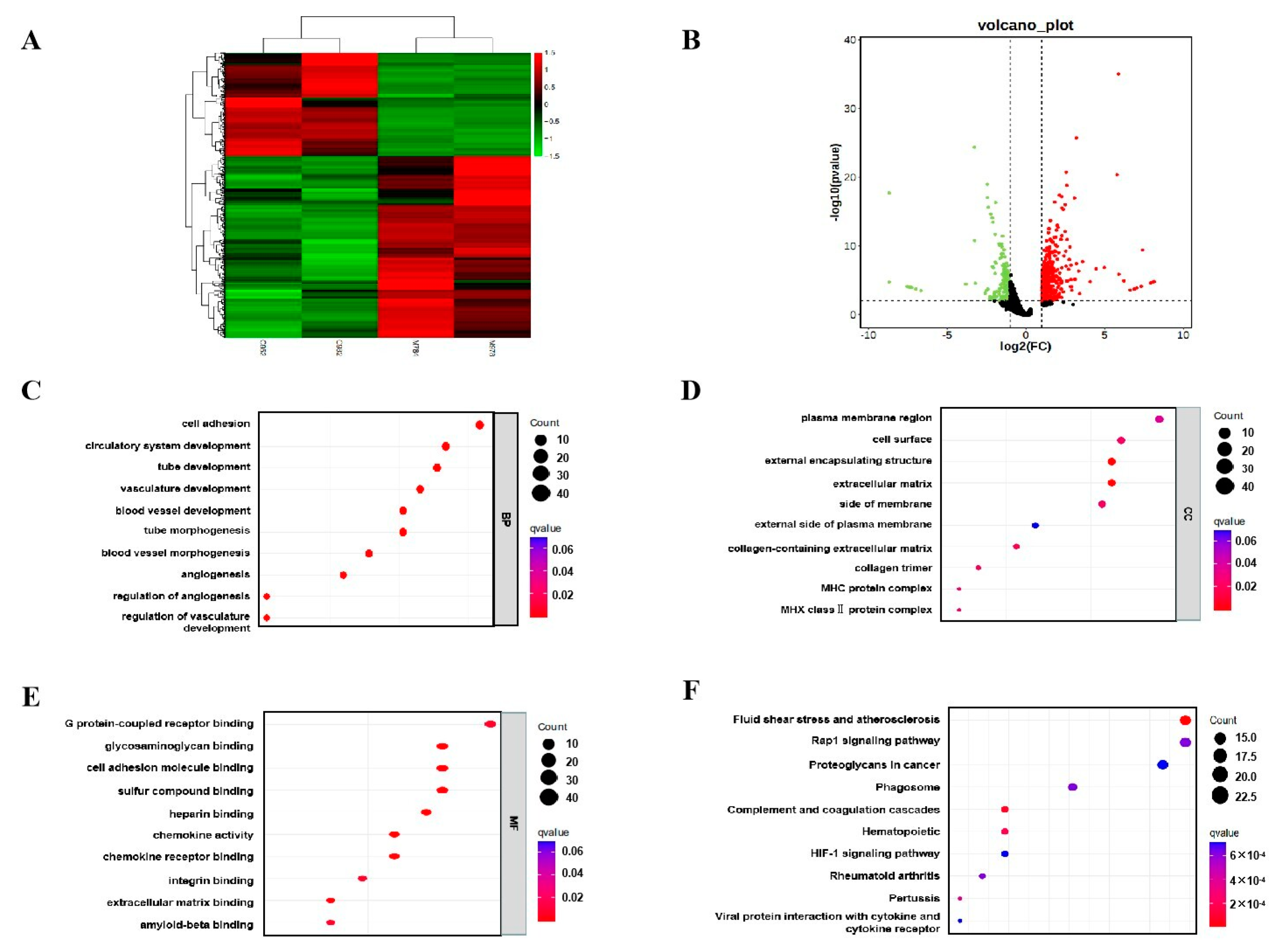
3.5. Effects of Melatonin Supplementation on the Transcriptome of Placental Tissue of Sows
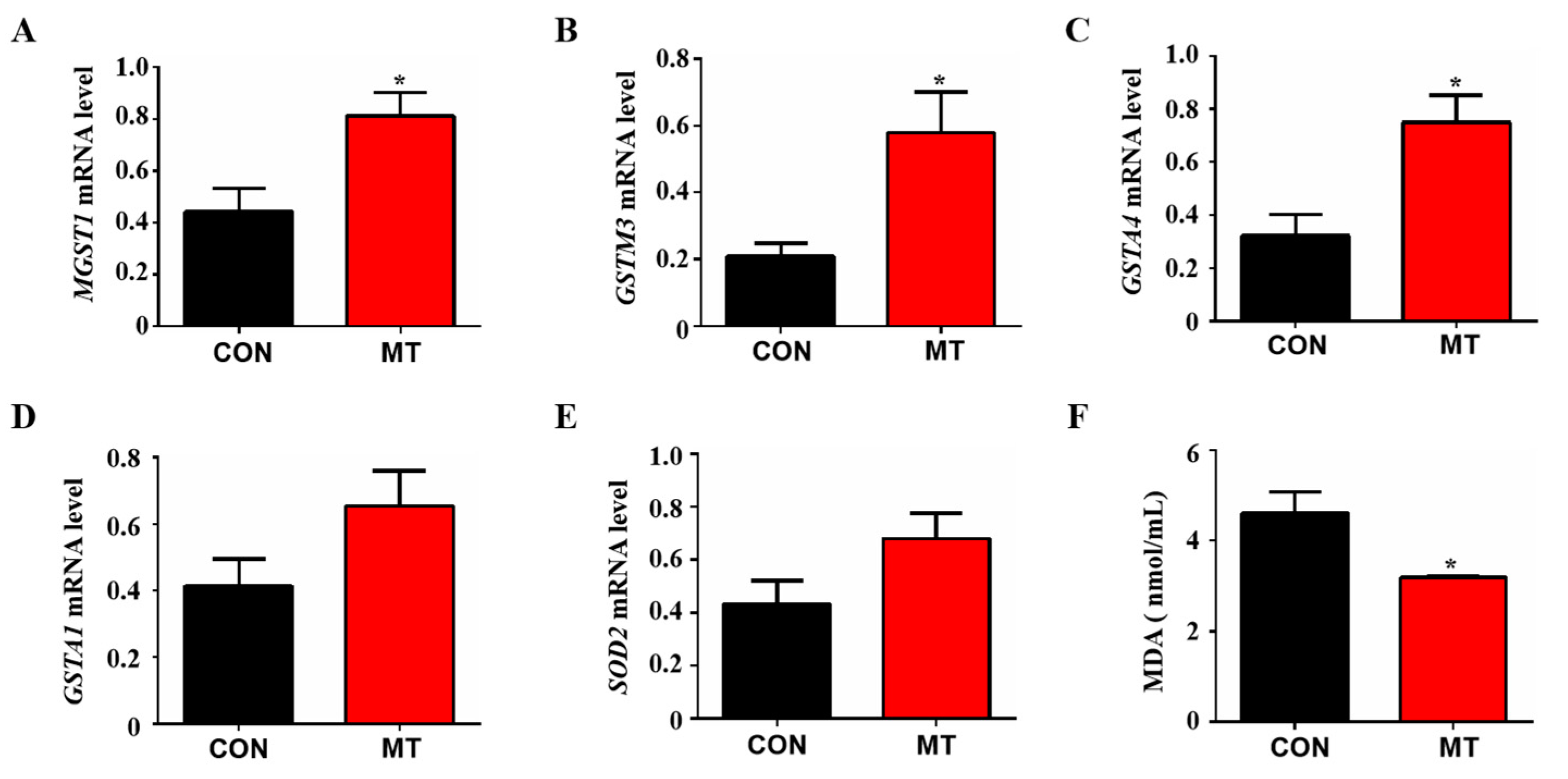
3.6. Placental Tissue Antioxidant Capacity Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, C.; Groom, K.M.; Oyston, C.; Chamley, L.W.; Clark, A.R.; James, J.L. The Placenta in Fetal Growth Restriction: What is Going Wrong? Placenta 2020, 96, 10–18. [Google Scholar] [CrossRef]
- Yajnik, C.S.; Deshmukh, U.S. Maternal Nutrition, Intrauterine Programming and Consequential Risks in the Offspring. Rev. Endocr. Metab. Disord. 2008, 9, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-González, G.L.; Vargas-Hernández, L.; Reyes-Castro, L.A.; Ibáñez, C.A.; Bautista, C.J.; Lomas-Soria, C.; Itani, N.; Estrada-Gutierrez, G.; Espejel-Nuñez, A.; Flores-Pliego, A.; et al. Resveratrol Supplementation in Obese Pregnant Rats Improves Maternal Metabolism and Prevents Increased Placental Oxidative Stress. Antioxidants 2022, 11, 1871. [Google Scholar] [CrossRef]
- Behlen, J.C.; Lau, C.H.; Pendleton, D.; Li, Y.; Hoffmann, A.R.; Golding, M.C.; Zhang, R.; Johnson, N.M. Nrf2-Dependent Placental Effects Vary by Sex and Dose Following Gestational Exposure to Ultrafine Particles. Antioxidants 2022, 11, 352. [Google Scholar] [CrossRef]
- Nagai, R.; Watanabe, K.; Wakatsuki, A.; Hamada, F.; Shinohara, K.; Hayashi, Y.; Imamura, R.; Fukaya, T. Melatonin Preserves Fetal Growth in Rats by Protecting Against Ischemia/Reperfusion-Induced Oxidative/Nitrosative Mitochondrial Damage in the Placenta. J. Pineal Res. 2008, 45, 271–276. [Google Scholar] [CrossRef]
- Qi, L.; Jiang, J.; Zhang, J.; Zhang, L.; Wang, T. Curcumin Protects Human Trophoblast Htr8/Svneo Cells From H2O2-Induced Oxidative Stress by Activating Nrf2 Signaling Pathway. Antioxidants 2020, 9, 121. [Google Scholar] [CrossRef]
- Hu, C.; Yang, Y.; Li, J.; Wang, H.; Cheng, C.; Yang, L.; Li, Q.; Deng, J.; Liang, Z.; Yin, Y.; et al. Maternal Diet-Induced Obesity Compromises Oxidative Stress Status and Angiogenesis in the Porcine Placenta by Upregulating Nox2 Expression. Oxidative Med. Cell. Longev. 2019, 2019, 2481592. [Google Scholar] [CrossRef]
- Bouroutzika, E.; Kouretas, D.; Papadopoulos, S.; Veskoukis, A.S.; Theodosiadou, E.; Makri, S.; Paliouras, C.; Michailidis, M.; Caroprese, M.; Valasi, I. Effects of Melatonin Administration to Pregnant Ewes Under Heat-Stress Conditions, in Redox Status and Reproductive Outcome. Antioxidants 2020, 9, 266. [Google Scholar] [CrossRef]
- Lian, S.; Guo, J.; Wang, L.; Li, W.; Wang, J.; Ji, H.; Kong, F.; Xu, B.; Li, S.; Yang, H. Impact of Prenatal Cold Stress on Placental Physiology, Inflammatory Response, and Apoptosis in Rats. Oncotarget 2017, 8, 115304–115314. [Google Scholar] [CrossRef]
- Zhang, J.; Fan, J.; Li, Y.; Liang, S.; Huo, S.; Wang, X.; Zuo, Y.; Cui, D.; Li, W.; Zhong, Z.; et al. Porcine Parvovirus Infection Causes Pig Placenta Tissue Damage Involving Nonstructural Protein 1 (Ns1)-Induced Intrinsic Ros/Mitochondria-Mediated Apoptosis. Viruses 2019, 11, 389. [Google Scholar] [CrossRef]
- Tan, C.; Ji, Y.; Zhao, X.; Xin, Z.; Li, J.; Huang, S.; Cui, Z.; Wen, L.; Liu, C.; Kim, S.W.; et al. Effects of Dietary Supplementation of Nucleotides From Late Gestation to Lactation on the Performance and Oxidative Stress Status of Sows and their Offspring. Anim. Nutr. 2021, 7, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Tian, X.; Peng, Y.; Sun, Z.; Wang, C.; Tang, N.; Li, B.; Jian, Y.; Wang, W.; Huo, X.; et al. Mitochondrial Cytochrome P450 (Cyp) 1B1 is Responsible for Melatonin Induced Apoptosis in Neural Cancer Cells. J. Pineal Res. 2018, 65, e12478. [Google Scholar] [CrossRef]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; Fougerou, C. Melatonin: Pharmacology, Functions and Therapeutic Benefits. Curr. Neuropharmacol. 2017, 15, 434–443. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but Over Delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin: A Versatile Protector Against Oxidative Dna Damage. Molecules 2018, 23, 530. [Google Scholar] [CrossRef]
- Gomes, P.; Motta-Teixeira, L.C.; Gallo, C.C.; Carmo, B.D.; Camargo, L.S.; Quintela, T.; Reiter, R.J.; Amaral, F.; Cipolla-Neto, J. Maternal Pineal Melatonin in Gestation and Lactation Physiology, and in Fetal Development and Programming. Gen. Comp. Endocrinol. 2021, 300, 113633. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.; Lacasse, A.; Lanoix, D.; Sagrillo-Fagundes, L.; Boulard, V.; Vaillancourt, C. Placental Melatonin System is Present Throughout Pregnancy and Regulates Villous Trophoblast Differentiation. J. Pineal Res. 2015, 59, 38–46. [Google Scholar] [CrossRef]
- Berbets, A.M.; Davydenko, I.S.; Barbe, A.M.; Konkov, D.H.; Albota, O.M.; Yuzko, O.M. Melatonin 1a and 1B Receptors’ Expression Decreases in the Placenta of Women with Fetal Growth Restriction. Reprod. Sci. 2021, 28, 197–206. [Google Scholar] [CrossRef]
- Berbets, A.M.; Barbe, A.M.; Andriiets, O.A.; Andriiets, A.V.; Yuzko, O.M. Melatonin Levels Decrease in the Umbilical Cord in Case of Intrauterine Growth Restriction. J. Med. Life 2020, 13, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Richter, H.G.; Hansell, J.A.; Raut, S.; Giussani, D.A. Melatonin Improves Placental Efficiency and Birth Weight and Increases the Placental Expression of Antioxidant Enzymes in Undernourished Pregnancy. J. Pineal Res. 2009, 46, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Renshall, L.J.; Morgan, H.L.; Moens, H.; Cansfield, D.; Finn-Sell, S.L.; Tropea, T.; Cottrell, E.C.; Greenwood, S.; Sibley, C.P.; Wareing, M.; et al. Melatonin Increases Fetal Weight in Wild-Type Mice but Not in Mouse Models of Fetal Growth Restriction. Front. Physiol. 2018, 9, 1141. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Cai, X.; Li, J.; Huang, Y.; Liu, H.; He, J.; Fang, Z.; Feng, B.; Tang, J.; Lin, Y.; et al. Effects of Melatonin Supplementation During Pregnancy On Reproductive Performance, Maternal–Placental–Fetal Redox Status, and Placental Mitochondrial Function in a Sow Model. Antioxidants 2021, 10, 1867. [Google Scholar] [CrossRef]
- Yang, Y.; Jin, H.; Qiu, Y.; Liu, Y.; Wen, L.; Fu, Y.; Qi, H.; Baker, P.N.; Tong, C. Reactive Oxygen Species are Essential for Placental Angiogenesis During Early Gestation. Oxid. Med. Cell. Longev. 2022, 2022, 4290922. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, P.; Zhu, F.; Liao, J.; Wu, Y.; Hu, M.; Fu, H.; Qiao, J.; Lin, L.; Huang, B.; et al. The Potent Antioxidant Mitoq Protects Against Preeclampsia During Late Gestation but Increases the Risk of Preeclampsia When Administered in Early Pregnancy. Antioxid. Redox Signal. 2021, 34, 118–136. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wei, H.; Sun, H.; Ao, J.; Long, G.; Jiang, S.; Peng, J. Effects of Dietary Supplementation of Oregano Essential Oil to Sows On Oxidative Stress Status, Lactation Feed Intake of Sows, and Piglet Performance. Biomed. Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Jiang, J.; Zhang, J.; Zhang, L.; Wang, T. Maternal Curcumin Supplementation Ameliorates Placental Function and Fetal Growth in Mice with Intrauterine Growth Retardation†. Biol. Reprod. 2020, 102, 1090–1101. [Google Scholar] [CrossRef]
- Miller, S.L.; Yawno, T.; Alers, N.O.; Castillo-Melendez, M.; Supramaniam, V.G.; Vanzyl, N.; Sabaretnam, T.; Loose, J.M.; Drummond, G.R.; Walker, D.W.; et al. Antenatal Antioxidant Treatment with Melatonin to Decrease Newborn Neurodevelopmental Deficits and Brain Injury Caused by Fetal Growth Restriction. J. Pineal Res. 2014, 56, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Melhuish, B.L.; Brown, G.M.; Goncalves, V.F.; Kennedy, J.L. Melatonin’s Neuroprotective Role in Mitochondria and its Potential as a Biomarker in Aging, Cognition and Psychiatric Disorders. Transl. Psychiatry 2021, 11, 339. [Google Scholar] [CrossRef]
- Ma, W.; Wu, H.; Li, G.; Yan, L.; Wang, L.; Zhao, M.; Guan, S.; Xu, S.; Guo, X.; Liu, F.; et al. Melatonin Promotes the Growth and Development of Lambs by Increasing Growth Hormone and Testosterone, Targeting On Apoptosis Signaling Pathway and Intestinal Microflora. Front. Endocrinol. 2022, 13, 966120. [Google Scholar] [CrossRef]
- Marquardt, R.M.; Kim, T.H.; Shin, J.; Jeong, J. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef]
- Lv, D.; Tan, T.; Zhu, T.; Wang, J.; Zhang, S.; Zhang, L.; Hu, X.; Liu, G.; Xing, Y. Leptin Mediates the Effects of Melatonin On Female Reproduction in Mammals. J. Pineal Res. 2019, 66, e12559. [Google Scholar] [CrossRef]
- Salak-Johnson, J.L. Social Status and Housing Factors Affect Reproductive Performance of Pregnant Sows in Groups. Mol. Reprod. Dev. 2017, 84, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Huang, J.; Wen, A.; Yan, P. Impaired Mitochondrial Function Results From Oxidative Stress in the Full-Term Placenta of Sows with Excessive Back-Fat. Animals 2020, 10, 360. [Google Scholar] [CrossRef]
- Misztal, T.; Romanowicz, K.; Barcikowski, B. Effects of Melatonin Infused Into the Iii Ventricle On Prolactin, Beta-Endorphin and Luteotropin Secretion in Ewes During the Different Stages of the Reproductive Cycle. Acta Neurobiol. Exp. 1996, 56, 769–778. [Google Scholar]
- Iwasaki, S.; Nakazawa, K.; Sakai, J.; Kometani, K.; Iwashita, M.; Yoshimura, Y.; Maruyama, T. Melatonin as a Local Regulator of Human Placental Function. J. Pineal Res. 2005, 39, 261–265. [Google Scholar] [CrossRef]
- Lanoix, D.; Beghdadi, H.; Lafond, J.; Vaillancourt, C. Human Placental Trophoblasts Synthesize Melatonin and Express its Receptors. J. Pineal Res. 2008, 45, 50–60. [Google Scholar] [CrossRef]
- Rashdan, N.A.; Zhai, B.; Lovern, P.C. Fluid Shear Stress Regulates Placental Growth Factor Expression via Heme Oxygenase 1 and Iron. Sci. Rep. 2021, 11, 14912. [Google Scholar] [CrossRef]
- Miura, S.; Sato, K.; Kato-Negishi, M.; Teshima, T.; Takeuchi, S. Fluid Shear Triggers Microvilli Formation via Mechanosensitive Activation of Trpv6. Nat. Commun. 2015, 6, 8871. [Google Scholar] [CrossRef]
- Psefteli, P.; Kitscha, P.; Vizcay, G.; Fleck, R.; Chapple, S.J.; Mann, G.E.; Fowler, M.; Siow, R.C. Glycocalyx Sialic Acids Regulate Nrf2-Mediated Signaling by Fluid Shear Stress in Human Endothelial Cells. Redox Biol. 2021, 38, 101816. [Google Scholar] [CrossRef]
- Warabi, E.; Takabe, W.; Minami, T.; Inoue, K.; Itoh, K.; Yamamoto, M.; Ishii, T.; Kodama, T.; Noguchi, N. Shear Stress Stabilizes Nf-E2-Related Factor 2 and Induces Antioxidant Genes in Endothelial Cells: Role of Reactive Oxygen/Nitrogen Species. Free Radic. Biol. Med. 2007, 42, 260–269. [Google Scholar] [CrossRef]
- Deng, C.C.; Zhang, J.P.; Huo, Y.N.; Xue, H.Y.; Wang, W.; Zhang, J.J.; Wang, X.Z. Melatonin Alleviates the Heat Stress-Induced Impairment of Sertoli Cells by Reprogramming Glucose Metabolism. J. Pineal Res. 2022, 73, e12819. [Google Scholar] [CrossRef]
- Sun, T.C.; Liu, X.C.; Yang, S.H.; Song, L.L.; Zhou, S.J.; Deng, S.L.; Tian, L.; Cheng, L.Y. Melatonin Inhibits Oxidative Stress and Apoptosis in Cryopreserved Ovarian Tissues via Nrf2/Ho-1 Signaling Pathway. Front. Mol. Biosci. 2020, 7, 163. [Google Scholar] [CrossRef]
- Bartolini, D.; Arato, I.; Mancuso, F.; Giustarini, D.; Bellucci, C.; Vacca, C.; Aglietti, M.C.; Stabile, A.M.; Rossi, R.; Cruciani, G.; et al. Melatonin Modulates Nrf2 Activity to Protect Porcine Pre-Pubertal Sertoli Cells From the Abnormal H2O2 Generation and Reductive Stress Effects of Cadmium. J. Pineal Res. 2022, 73, e12808. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Yan, L.; Han, Q.; Li, G.; Wu, H.; Ma, X.; Zhao, M.; Ma, W.; Ji, P.; Zhang, R.; et al. Melatonin Supplementation during the Late Gestational Stage Enhances Reproductive Performance of Sows by Regulating Fluid Shear Stress and Improving Placental Antioxidant Capacity. Antioxidants 2023, 12, 688. https://doi.org/10.3390/antiox12030688
Wang L, Yan L, Han Q, Li G, Wu H, Ma X, Zhao M, Ma W, Ji P, Zhang R, et al. Melatonin Supplementation during the Late Gestational Stage Enhances Reproductive Performance of Sows by Regulating Fluid Shear Stress and Improving Placental Antioxidant Capacity. Antioxidants. 2023; 12(3):688. https://doi.org/10.3390/antiox12030688
Chicago/Turabian StyleWang, Likai, Laiqing Yan, Qi Han, Guangdong Li, Hao Wu, Xiao Ma, Mengmeng Zhao, Wenkui Ma, Pengyun Ji, Ran Zhang, and et al. 2023. "Melatonin Supplementation during the Late Gestational Stage Enhances Reproductive Performance of Sows by Regulating Fluid Shear Stress and Improving Placental Antioxidant Capacity" Antioxidants 12, no. 3: 688. https://doi.org/10.3390/antiox12030688
APA StyleWang, L., Yan, L., Han, Q., Li, G., Wu, H., Ma, X., Zhao, M., Ma, W., Ji, P., Zhang, R., & Liu, G. (2023). Melatonin Supplementation during the Late Gestational Stage Enhances Reproductive Performance of Sows by Regulating Fluid Shear Stress and Improving Placental Antioxidant Capacity. Antioxidants, 12(3), 688. https://doi.org/10.3390/antiox12030688