
Solanum dulcamara L. Berries: A Convenient Model System to Study Redox Processes in Relation to Fruit Ripening

 , , ,
, , ,  , , , ,
, , , ,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Metabolomics of MG and RR Bittersweet Fruits
2.1.1. Chemicals and Reagents
2.1.2. Sampling and Storage of Bittersweet Berries
2.1.3. Preparation of Methanol Extracts of Bittersweet Berries
2.1.4. UHPLC-LTQ OrbiTrap MS Qualitative Analysis of Phenolics in Bittersweet Fruits
2.1.5. UHPLC/DAD/(±)HESI−MS2 Quantification of Major Phenolics
2.2. EPR Measurements
2.2.1. Determination of the Scavenging Activity of Whole and Selected Tissue Bittersweet Extracts towards DPPH Radicals
2.2.2. Determination of the Scavenging Activity of Bittersweet Fruits Water Extracts towards Hydroxyl Radicals
2.2.3. Determination of the Capacity of Bittersweet Fruit Tissues to Reduce Pyrrolidine Spin Probes (X-Band 1D Gradient Imaging)
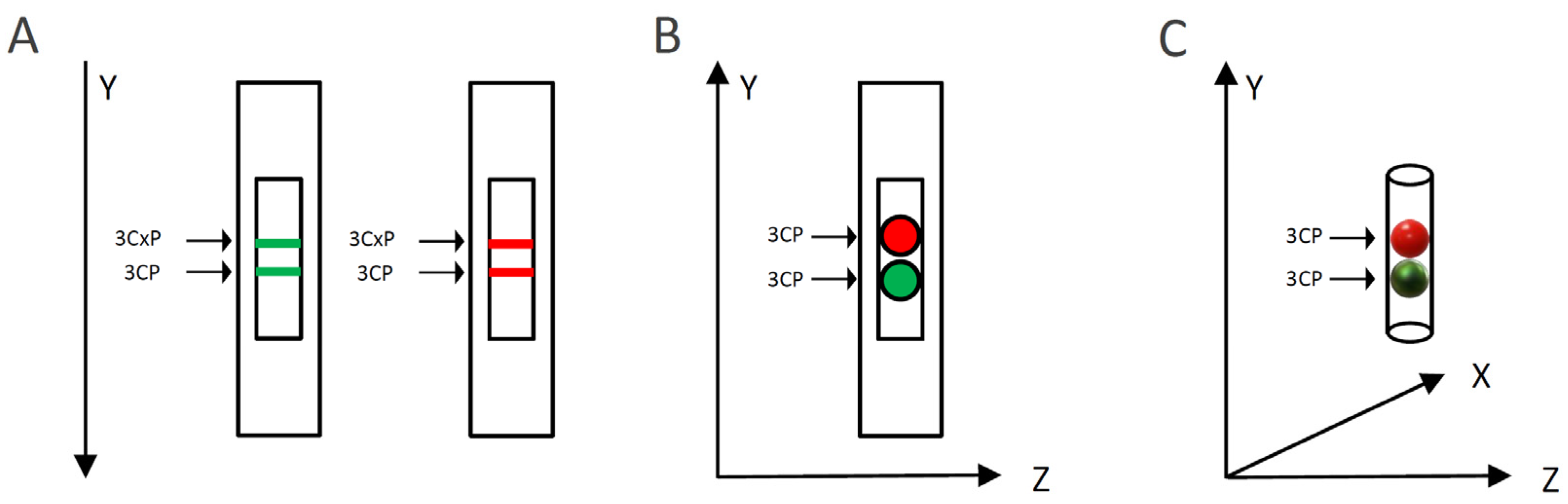
2.2.4. Spatiotemporal Visualization of the Capacity of the Bittersweet Fruits to Reduce Pyrrolidine Spin Probe (X-Band 2D Imaging)
2.2.5. Spatiotemporal Visualization of the Capacity of the Intact Bittersweet Fruits to Reduce Pyrrolidine Spin Probe (L-Band 2D Imaging)
2.3. High Performance Thin Layer Chromatography (HPTLC) and HPTLC-DPPH Bioautography Assay
2.4. Activities of Antioxidant Enzymes
2.5. Statistical Analysis
3. Results
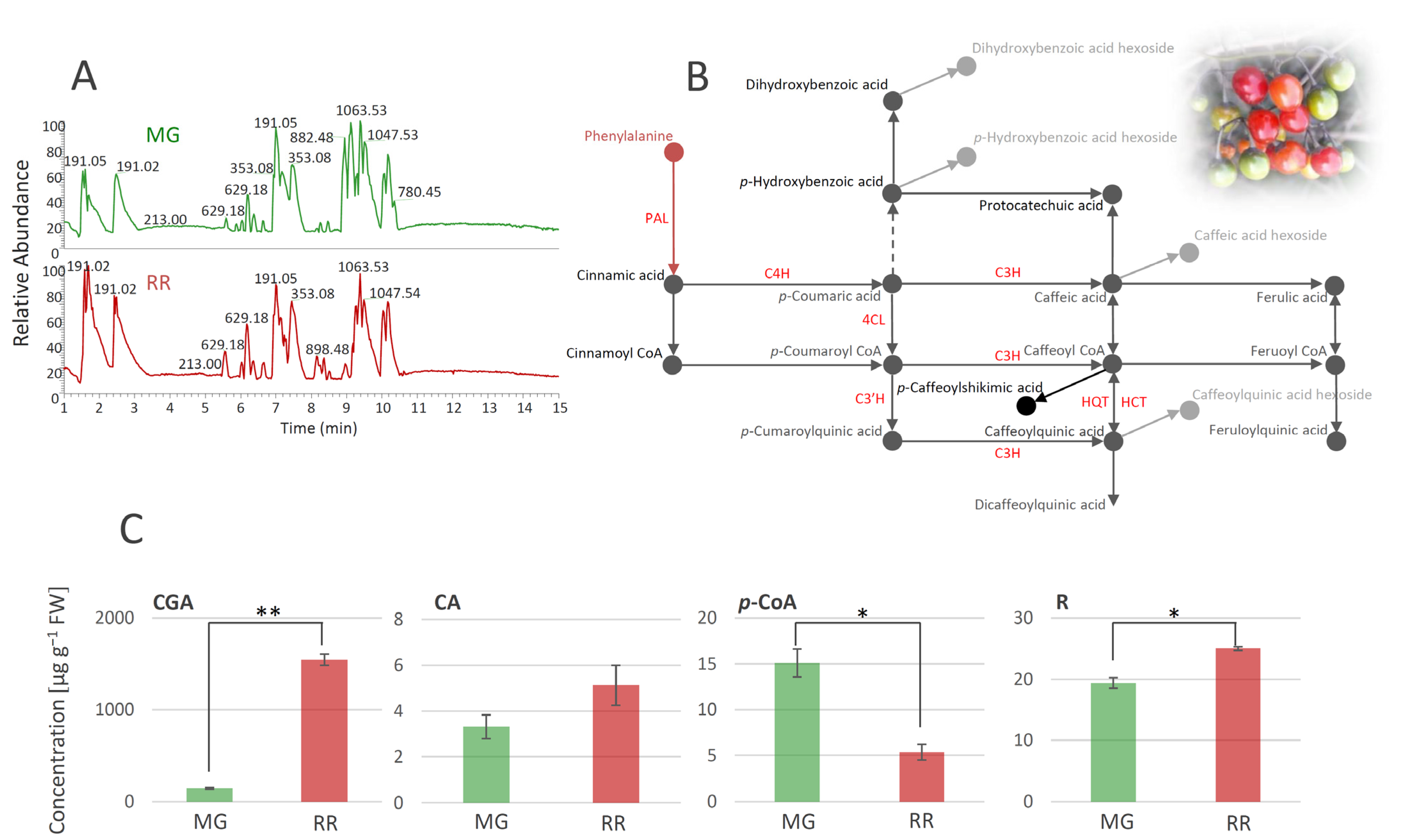
3.1. Metabolic Profiling of Solanum Dulcamara Fruits
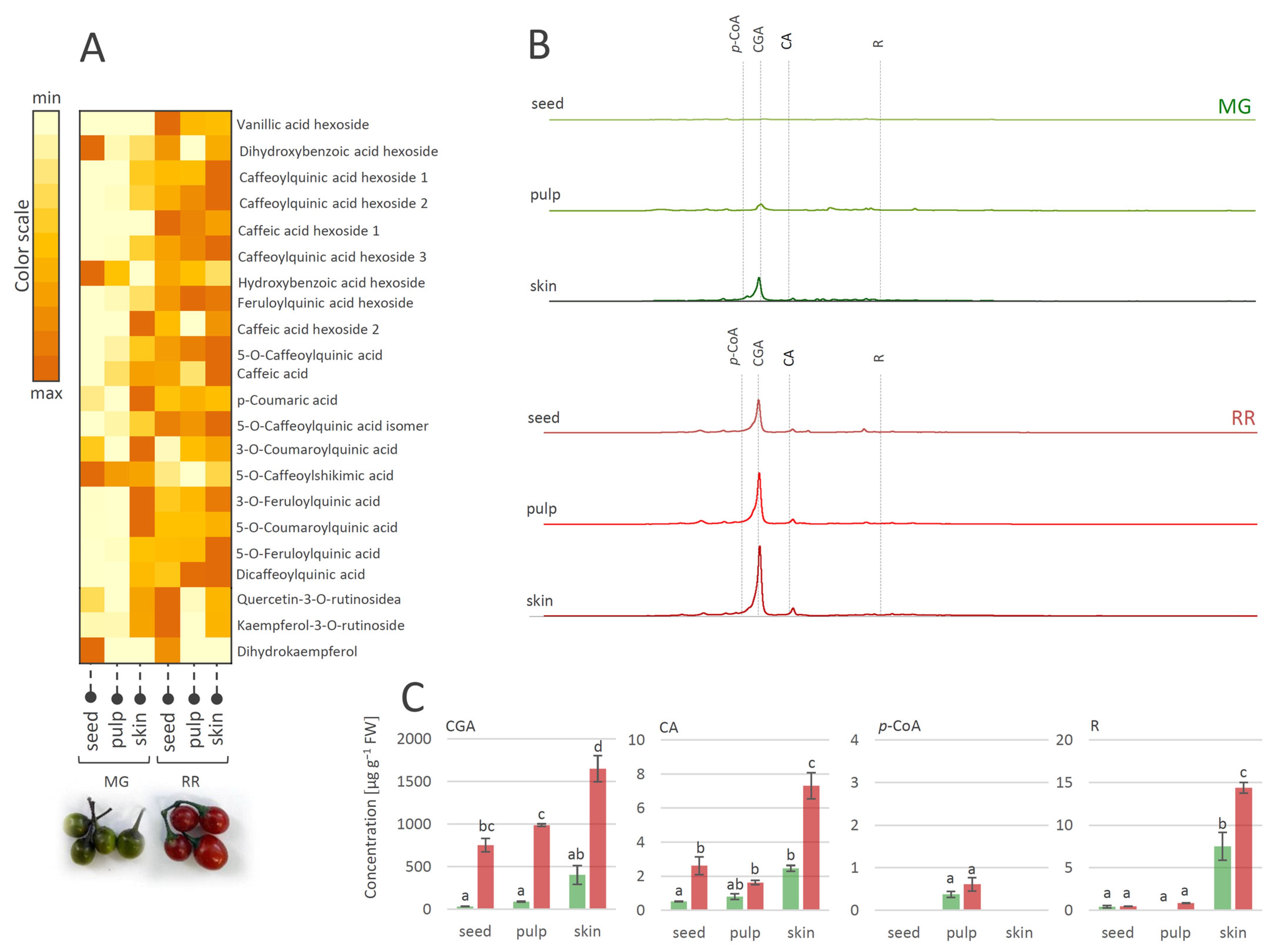
3.2. Tissue-Specific Distribution of Polyphenolics in MG and RR Berries of S. dulcamara
3.3. Tissue-Specific Redox State of Bittersweet Fruits
3.3.1. Scavenging Activity of MG and RR Bittersweet Fruits towards DPPH and Hydroxyl Radicals
3.3.2. Tissue-Specific Scavenging Activity of MG and RR Bittersweet Fruits Extracts towards DPPH Radicals
3.3.3. Determination of the Capacity of Bittersweet Fruits to Reduce Pyrrolidine Spin Probes (X-Band 1D Gradient Imaging)
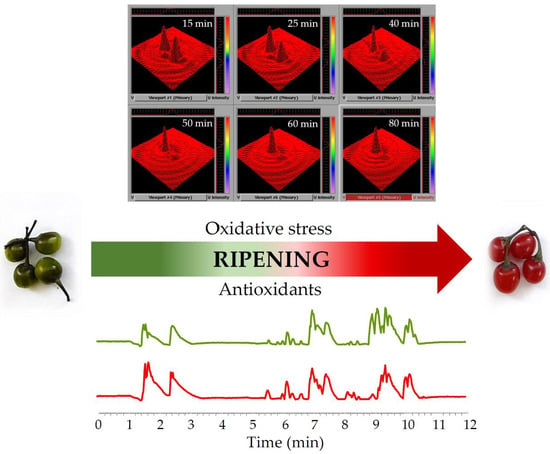
3.3.4. Spatiotemporal Visualization of the Capacity of the Bittersweet Fruits to Reduce Pyrrolidine Spin Probe (X-Band 2D Imaging)
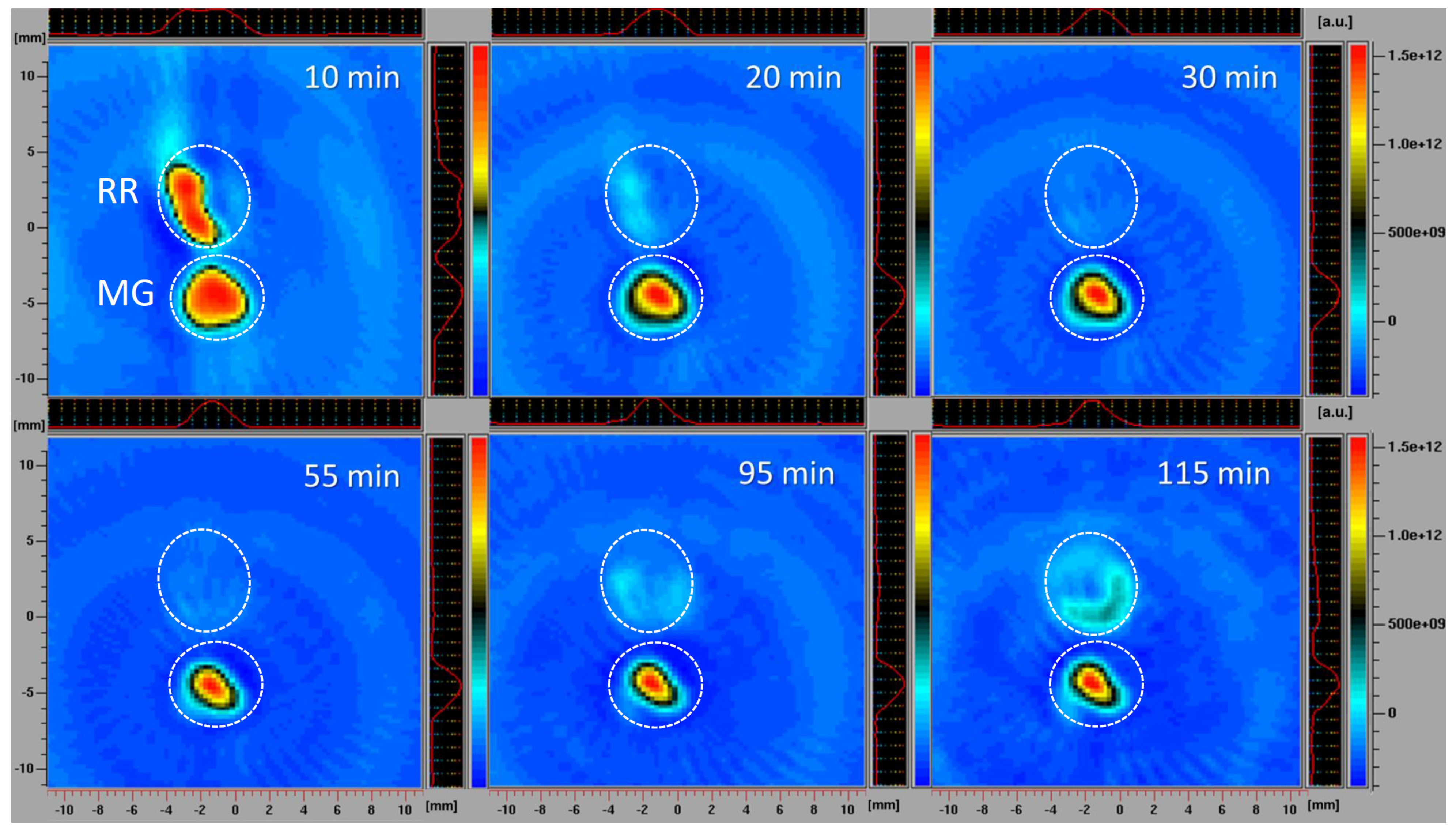
3.3.5. Spatiotemporal Visualization of the Capacity of the Intact Bittersweet Fruits to Reduce Pyrrolidine Spin Probe (L-Band 2D Imaging)
3.4. Tissue-Specific Activity of Antioxidant Enzymes in MG and RR Bittersweet Berries
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Decros, G.; Baldet, P.; Beauvoit, B.; Stevens, R.; Flandin, A.; Colombié, S.; Gibon, Y.; Pétriacq, P. Get the Balance Right: ROS Homeostasis and Redox Signalling in Fruit. Front. Plant Sci. 2019, 10, 1091. [Google Scholar] [CrossRef] [PubMed]
- King County Noxious Weed Control Program. Garlic Mustard Best Management Practices. 2010. Available online: http://your.kingcounty.gov/dnrp/library/water-and-land/weeds/BMPs/GarlicMustard-Control.pdf (accessed on 24 November 2022).
- Ui, I.; Okajo, A.; Endo, K.; Utsumi, H.; Matsumoto, K.-I. Effect of Hydrogen Peroxide in Redox Status Estimation Using Nitroxyl Spin Probe. Free. Radic. Biol. Med. 2004, 37, 2012–2017. [Google Scholar] [CrossRef]
- Dragišić Maksimović, J.; Mojović, M.; Vučinić, Ž.; Maksimović, V. Spatial Distribution of Apoplastic Antioxidative Constituents in Maize Root. Physiol. Plant. 2021, 173, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Kostić, A.Ž.; Gašić, U.M.; Pešić, M.B.; Stanojević, S.P.; Barać, M.B.; Mačukanović-Jocić, M.P.; Avramov, S.N.; Tešić, Ž.L. Phytochemical Analysis and Total Antioxidant Capacity of Rhizome, Above-Ground Vegetative Parts and Flower of Three Iris Species. Chem. Biodivers. 2019, 16, e1800565. [Google Scholar] [CrossRef]
- Gómez-Romero, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Metabolite Profiling and Quantification of Phenolic Compounds in Methanol Extracts of Tomato Fruit. Phytochemistry 2010, 71, 1848–1864. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-B.; Meyer, R.S.; Whitaker, B.D.; Litt, A.; Kennelly, E.J. A New Liquid Chromatography–Mass Spectrometry-Based Strategy to Integrate Chemistry, Morphology, and Evolution of Eggplant (Solanum) Species. J. Chromatogr. A 2013, 1314, 154–172. [Google Scholar] [CrossRef] [PubMed]
- Daji, G.; Steenkamp, P.; Madala, N.; Dlamini, B. Phytochemical Composition of Solanum retroflexum Analysed with the Aid of Ultra-Performance Liquid Chromatography Hyphenated to Quadrupole-Time-of-Flight Mass Spectrometry (UPLC-qTOF-MS). J. Food Qual. 2018, 2018, 3678795. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Gómez-Caravaca, A.M.; Guerra-Hernández, E.; Cerretani, L.; García-Villanova, B.; Verardo, V. Comprehensive Metabolite Profiling of Solanum tuberosum L. (Potato) Leaves by HPLC-ESI-QTOF-MS. Food Res. Int. 2018, 112, 390–399. [Google Scholar] [CrossRef]
- Bednarz, H.; Roloff, N.; Niehaus, K. Mass Spectrometry Imaging of the Spatial and Temporal Localization of Alkaloids in Nightshades. J. Agric. Food Chem. 2019, 67, 13470–13477. [Google Scholar] [CrossRef]
- Liu, C.; Tian, J.; An, T.; Lyu, F.; Jia, P.; Zhou, M.; Liu, Z.; Feng, Y. Secondary Metabolites from Solanum rostratum and Their Antifeedant Defense Mechanisms against Helicoverpa armigera. J. Agric. Food Chem. 2020, 68, 88–96. [Google Scholar] [CrossRef]
- Banjanac, T.; Dragicevic, M.; Siler, B.; Gasic, U.; Bohanec, B.; Nestorovic Zivkovic, J.; Trifunovic, S.; Misic, D. Chemodiversity of Two Closely Related Tetraploid Centaurium Species and Their Hexaploid Hybrid: Metabolomic Search for High-Resolution Taxonomic Classifiers. Phytochemistry 2017, 140, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Sanna, D.; Delogu, G.; Mulas, M.; Schirra, M.; Fadda, A. Determination of Free Radical Scavenging Activity of Plant Extracts Through DPPH Assay: An EPR and UV–Vis Study. Food Anal. Methods 2012, 5, 759–766. [Google Scholar] [CrossRef]
- Gođevac, D.; Vujisić, L.; Mojović, M.; Ignjatović, A.; Spasojević, I.; Vajs, V. Evaluation of Antioxidant Capacity of Allium Ursinum L. Volatile Oil and Its Effect on Membrane Fluidity. Food Chem. 2008, 107, 1692–1700. [Google Scholar] [CrossRef]
- Nakarada, Đ.J.; Marković, S.Z.; Popović, M.D.; Dimitrijević, M.S.; Rakić, A.A.; Mojović, M.D. Redox Properties of Grape Wine Skin Extracts from the Šumadija Region: An Electron Paramagnetic Resonance Study. Hosp. Pharmacol.-Int. Multidiscip. J. 2021, 8, 1004–1013. [Google Scholar] [CrossRef]
- Savić, A.G.; Mojović, M. Free Radicals Identification from the Complex EPR Signals by Applying Higher Order Statistics. Anal. Chem. 2012, 84, 3398–3402. [Google Scholar] [CrossRef]
- Nakarada, Đ.; Pejin, B.; Tommonaro, G.; Mojović, M. Liposomal Integration Method for Assessing Antioxidative Activity of Water Insoluble Compounds Towards Biologically Relevant Free Radicals: Example Of Avarol. J. Liposome Res. 2020, 30, 218–226. [Google Scholar] [CrossRef]
- Villamena, F.A.; Hadad, C.M.; Zweier, J.L. Kinetic Study and Theoretical Analysis of Hydroxyl Radical Trapping and Spin Adduct Decay of Alkoxycarbonyl and Dialkoxyphosphoryl Nitrones in Aqueous Media. J. Phys. Chem. A 2003, 107, 4407–4414. [Google Scholar] [CrossRef]
- Puač, N.; Škoro, N.; Spasić, K.; Živković, S.; Milutinović, M.; Malović, G.; Petrović, Z.L. Activity of Catalase Enzyme in Paulownia tomentosa Seeds During the Process of Germination after Treatments with Low Pressure Plasma and Plasma Activated Water. Plasma Process. Polym. 2018, 15, 1700082. [Google Scholar] [CrossRef]
- Dmitrović, S.; Simonović, A.; Mitić, N.; Savić, J.; Cingel, A.; Filipović, B.; Ninković, S. Hairy Root Exudates of Allelopathic Weed Chenopodium murale L. Induce Oxidative Stress and Down-Regulate Core Cell Cycle Genes in Arabidopsis and Wheat Seedlings. Plant Growth Regul. 2015, 75, 365–382. [Google Scholar] [CrossRef]
- Pouresmaeil, M.; Nojadeh, M.S.; Movafeghi, A.; Maggi, F. Exploring the Bio-Control Efficacy of Artemisia fragrans Essential Oil on the Perennial Weed Convolvulus arvensis: Inhibitory Effects on the Photosynthetic Machinery and Induction of Oxidative Stress. Ind. Crops Prod. 2020, 155, 112785. [Google Scholar] [CrossRef]
- Živković, S.; Popović, M.; Dragišić-Maksimović, J.; Momcilović, I.; Grubisic, D.V. Dehydration-Related Changes of Peroxidase and Polyphenol Oxidase Activity in Fronds of the Resurrection Fern Asplenium ceterach L. Arch. Biol. Sci. 2010, 62, 1071–1081. [Google Scholar] [CrossRef]
- Takács, Z.; Poór, P.; Tari, I. Comparison of Polyamine Metabolism in Tomato Plants Exposed to Different Concentrations of Salicylic Acid under Light or Dark Conditions. Plant Physiol. Biochem. 2016, 108, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Vallverdú-Queralt, A.; de Alvarenga, J.F.R.; Estruch, R.; Lamuela-Raventos, R.M. Bioactive Compounds Present in the Mediterranean Sofrito. Food Chem. 2013, 141, 3365–3372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-H.; Hu, X.; Shi, S.-Y.; Huang, L.-Q.; Chen, W.; Chen, L.; Cai, P. Typical Ultraviolet Spectra in Combination with Diagnostic Mass Fragmentation Analysis for the Rapid and Comprehensive Profiling of Chlorogenic Acids in the Buds of Lonicera macranthoides. Anal. Bioanal. Chem. 2016, 408, 3659–3672. [Google Scholar] [CrossRef] [PubMed]
- Moco, S.; Bino, R.J.; Vorst, O.; Verhoeven, H.A.; de Groot, J.; van Beek, T.A.; Vervoort, J.; De Vos, C.R. A Liquid Chromatography-Mass Spectrometry-Based Metabolome Database for Tomato. Plant Physiol. 2006, 141, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Chideh, S.; Pilard, S.; Attoumbré, J.; Saguez, R.; Hassan-Abdallah, A.; Cailleu, D.; Wadouachi, A.; Baltora-Rosset, S. 5-O-Caffeoylshikimic acid from Solanum somalense Leaves: Advantage of Centrifugal Partition Chromatography over Conventional Column Chromatography. J. Sep. Sci. 2014, 37, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Puzanskiy, R.; Yemelyanov, V.; Shavarda, A.; Gavrilenko, T.; Shishova, M. Age-And Organ-Specific Differences of Potato (Solanum phureja) Plants Metabolome. Russ. J. Plant Physiol. 2018, 65, 813–823. [Google Scholar] [CrossRef]
- Sun, J.; Huo, H.-X.; Zhang, J.; Huang, Z.; Zheng, J.; Zhang, Q.; Zhao, Y.-F.; Li, J.; Tu, P.-F. Phenylpropanoid Amides from the Roots of Solanum melongena L. (Solanaceae). Biochem. Syst. Ecol. 2015, 58, 265–269. [Google Scholar] [CrossRef]
- Zeiss, D.R.; Mhlongo, M.I.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Comparative Metabolic Phenotyping of Tomato (Solanum lycopersicum) for the Identification of Metabolic Signatures in Cultivars Differing in Resistance to Ralstonia solanacearum. Int. J. Mol. Sci. 2018, 19, 2558. [Google Scholar] [CrossRef]
- Dastmalchi, K.; Rodriguez, M.P.; Lin, J.; Yoo, B.; Stark, R.E. Temporal Resistance of Potato Tubers: Antibacterial Assays and Metabolite Profiling of Wound-Healing Tissue Extracts from Contrasting Cultivars. Phytochemistry 2019, 159, 75–89. [Google Scholar] [CrossRef]
- Oertel, A.; Matros, A.; Hartmann, A.; Arapitsas, P.; Dehmer, K.J.; Martens, S.; Mock, H.-P. Metabolite Profiling of Red and Blue Potatoes Revealed Cultivar and Tissue Specific Patterns for Anthocyanins and other Polyphenols. Planta 2017, 246, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, H.; Sashida, Y.; Mimaki, Y. Steroidal Saponins, Pardarinoside A–G from the Bulbs of Lilium pardarinum. Phytochemistry 1989, 28, 3163–3170. [Google Scholar] [CrossRef]
- Russo, M.V.; Avino, P.; Centola, A.; Notardonato, I.; Cinelli, G. Rapid and Simple Determination of Acrylamide in Conventional Cereal-Based Foods and Potato Chips through Conversion to 3-[Bis (Trifluoroethanoyl) Amino]-3-Oxopropyl Trifluoroacetate by Gas Chromatography Coupled with Electron Capture and Ion Trap Mass Spectrometry Detectors. Food Chem. 2014, 146, 204–211. [Google Scholar]
- Wang, Y.; Xiang, L.; Yi, X.; He, X. Potential Anti-Inflammatory Steroidal Saponins from the Berries of Solanum nigrum L. (European Black Nightshade). J. Agric. Food Chem. 2017, 65, 4262–4272. [Google Scholar] [CrossRef] [PubMed]
- De Nicolò, A.; Avataneo, V.; Rabbia, F.; Sciandra, M.; Tosello, F.; Cusato, J.; Perlo, E.; Mulatero, P.; Veglio, F.; Di Perri, G. UHPLC–MS/MS Method with Sample Dilution to Test Therapeutic Adherence through Quantification of Ten Antihypertensive Drugs in Urine Samples. J. Pharm. Biomed. Anal. 2017, 142, 279–285. [Google Scholar] [CrossRef]
- Sabiu, S.; Ajani, E.; Aladodo, R.; Garuba, T.; Agunbiade, M.; Alimi, A.; Lekena, N.; Abdulrazaq, M. Membrane Stabilization and Probable Mechanisms of Hypoglycemic Activity of Fruit Extract of Solanum incanum L. (Solanaceae). Comp. Clin. Pathol. 2018, 27, 1611–1619. [Google Scholar] [CrossRef]
- Zhu, X.-H.; Ikeda, T.; Nohara, T. Studies on the constituents of solanaceous plants.(46). Steroidal Glycosides from the Fruits of Solanum anguivi. Chem. Pharm. Bull. 2000, 48, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Maurya, A.; Manika, N.; Verma, R.K.; Singh, S.C.; Srivastava, S.K. Simple and Reliable Methods for the Determination of Three Steroidal Glycosides in the Eight Species of Solanum by Reversed-Phase HPLC Coupled with Diode Array Detection. Phytochem. Anal. 2013, 24, 87–92. [Google Scholar] [CrossRef]
- Lee, Y.-Y.; HASHIMOTO, F.; YAHARA, S.; NOHARA, T.; YOSHIDA, N. Steroidal Glycosides from Solanum dulcamara. Chem. Pharm. Bull. 1994, 42, 707–709. [Google Scholar] [CrossRef]
- Yamashita, T.; MATSUMOTO, T.; Yahara, S.; Yoshida, N.; Nohara, T. Structures of Two New Steroidal Glycosides, Soladulcosides A and B from Solanum dulcamara. Chem. Pharm. Bull. 1991, 39, 1626–1628. [Google Scholar] [CrossRef]
- Ono, M.; Shimode, M.; Tsutsumi, S.; Yasuda, S.; Okawa, M.; Kinjo, J.; Miyashita, H.; Ikeda, T.; Yoshimitsu, H.; Nohara, T. A New Steroidal Glycoside from the Fruits of Solanum myriacanthum. Nat. Prod. Res. 2021, 36, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Uenosono, Y.; Umaoka, H.; Shiono, Y.; Ikeda, T.; Okawa, M.; Kinjo, J.; Yoshimitsu, H.; Nohara, T. Five New Steroidal Glycosides from the Stems of Solanum sodomaeum. Chem. Pharm. Bull. 2009, 57, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Ta, C.A.K.; Guerrero-Analco, J.A.; Roberts, E.; Liu, R.; Mogg, C.D.; Saleem, A.; Otárola-Rojas, M.; Poveda, L.; Sanchez-Vindas, P.; Cal, V. Antifungal Saponins from the Maya Medicinal Plant Cestrum schlechtendahlii G. Don (Solanaceae). Phytother. Res. 2016, 30, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Villano, C.; D’Amelia, V.; Esposito, S.; Adelfi, M.G.; Contaldi, F.; Ferracane, R.; Vitaglione, P.; Aversano, R.; Carputo, D. Genome-Wide HMG Family Investigation and its Role in Glycoalkaloid Accumulation in Wild Tuber-bearing Solanum commersonii. Life 2020, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Wang, C.; Wang, L.; Song, X.; Yang, H.; Fu, Q.; Zhao, W. Preparative Isolation and Purification of Steroidal Glycoalkaloid from the Ripe Berries of Solanum nigrum L. by Preparative HPLC–MS and UHPLC–TOF-MS/MS and its Anti-non-small Cell Lung Tumors Effects in Vitro and in Vivo. J. Sep. Sci. 2019, 42, 2471–2481. [Google Scholar] [CrossRef]
- Yoshimitsu, H.; Nishida, M.; Yoshida, M.; Nohara, T. Four New 26-Aminocholestane-Type Glycosides from Solanum abutiloides. Chem. Pharm. Bull. 2002, 50, 284–286. [Google Scholar] [CrossRef]
- Pinto, F.d.C.L.; Torres, M.d.C.M.; Silveira, E.R.; Pessoa, O.D.L.; Braz-Filho, R.; Guedes, M.L.d.S. Constituintes Químicos de Solanum buddleifolium Sendtn. Química Nova 2013, 36, 1111–1115. [Google Scholar] [CrossRef]
- Xiang, L.; Wang, Y.; Yi, X.; He, X. Steroidal Alkaloid Glycosides and Phenolics from the Immature Fruits of Solanum nigrum. Fitoterapia 2019, 137, 104268. [Google Scholar] [CrossRef]
- Wang, L.; Wang, N.; Yao, X. Non-saponins from Solanum nigrum L. Zhong Yao Cai=Zhongyaocai=J. Chin. Med. Mater. 2007, 30, 792–794. [Google Scholar]
- Elbermawi, A.; Halim, A.F.; Mansour, E.-S.S.; Ahmad, K.F.; Elsbaey, M.; Ashour, A.; Amen, Y.; El-Gamil, M.M.; Tomofumi, M.; Shimizu, K. Lycium schweinfurthii: New Secondary Metabolites and their Cytotoxic Activities. Nat. Prod. Res. 2022, 36, 5134–5141. [Google Scholar] [CrossRef]
- Yang, B.-Y.; Yin, X.; Liu, Y.; Sun, Y.; Guan, W.; Zhou, Y.-Y.; Kuang, H.-X. Terpenes and Lignans from the Roots of Solanum melongena L. Nat. Prod. Res. 2020, 34, 359–368. [Google Scholar] [CrossRef]
- Miralles, P.; López, A.; Dualde, P.; Coscollà, C.; Yusà, V. Liquid Chromatography-Orbitrap Tribrid High-Resolution Mass Spectrometry using Data Dependent-Tandem Mass Spectrometry with Triple Stage Fragmentation as a Screening Tool to Perform Identification and Risk Assessment of Unknown Substances in Food Contact Epoxy Resin. J. Sep. Sci. 2021, 44, 3020–3030. [Google Scholar] [PubMed]
- Sade, D.; Shriki, O.; Cuadros-Inostroza, A.; Tohge, T.; Semel, Y.; Haviv, Y.; Willmitzer, L.; Fernie, A.R.; Czosnek, H.; Brotman, Y. Comparative Metabolomics and Transcriptomics of Plant Response to Tomato Yellow Leaf Curl Virus Infection in Resistant and Susceptible Tomato Cultivars. Metabolomics 2015, 11, 81–97. [Google Scholar] [CrossRef]
- Mosad, R.R.; Ali, M.H.; Ibrahim, M.T.; Shaaban, H.M.; Emara, M.; Wahba, A.E. New Cytotoxic Steroidal Saponins from Cestrum parqui. Phytochem. Lett. 2017, 22, 167–173. [Google Scholar] [CrossRef]
- Sugimoto, K.; Matsui, K.; Iijima, Y.; Akakabe, Y.; Muramoto, S.; Ozawa, R.; Uefune, M.; Sasaki, R.; Alamgir, K.M.; Akitake, S. Intake and Transformation to a Glycoside of (Z)-3-Hexenol from Infested Neighbors Reveals a Mode of Plant Odor Reception and Defense. Proc. Natl. Acad. Sci. USA 2014, 111, 7144–7149. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Andújar, C.; Martínez-Pérez, A.; Albacete, A.; Martínez-Melgarejo, P.A.; Dodd, I.C.; Thompson, A.J.; Mohareb, F.; Estelles-Lopez, L.; Kevei, Z.; Ferrández-Ayela, A. Overproduction of ABA in Rootstocks Alleviates Salinity Stress in Tomato Shoots. Plant Cell Environ. 2021, 44, 2966–2986. [Google Scholar] [CrossRef] [PubMed]
- Ekanayaka, E.P.; Li, C.; Jones, A.D. Sesquiterpenoid Glycosides from Glandular Trichomes of the Wild Tomato Relative Solanum habrochaites. Phytochemistry 2014, 98, 223–231. [Google Scholar] [CrossRef]
- Voynikov, Y.; Zheleva-Dimitrova, D.; Gevrenova, R.; Lozanov, V.; Zaharieva, M.M.; Tsvetkova, I.; Najdenski, H.; Yagi, S.; Almoulah, N.F.; Momekov, G. Hydroxycinnamic Acid Amide Profile of Solanum schimperianum Hochst by UPLC-HRMS. Int. J. Mass Spectrom. 2016, 408, 42–50. [Google Scholar] [CrossRef]
- Song, Y.; Mei, T.; Liu, Y.; Kong, S.; Zhang, J.; Xie, M.; Ou, S.; Liang, M.; Wang, Q. Metabolites Identification of Chemical Constituents from the Eggplant (Solanum melongena L.) Calyx in Rats by UPLC/ESI/qTOF-MS Analysis and Their Cytotoxic Activities. Front. Pharmacol. 2021, 926, 655008. [Google Scholar] [CrossRef]
- Van der Rest, B.; Danoun, S.; Boudet, A.-M.; Rochange, S.F. Down-regulation of Cinnamoyl-Coa Reductase in Tomato (Solanum lycopersicum L.) Induces Dramatic Changes in Soluble Phenolic Pools. J. Exp. Bot. 2006, 57, 1399–1411. [Google Scholar] [CrossRef]
- Helmja, K.; Vaher, M.; Gorbatšova, J.; Kaljurand, M. Characterization of Bioactive Compounds Contained in Vegetables of the Solanaceae Family by Capillary Electrophoresis. Proc. Est. Acad. Sci. Chem. 2007, 56, 4. [Google Scholar]
- Scalzo, R.L.; Florio, F.E.; Fibiani, M.; Speranza, G.; Rabuffetti, M.; Gattolin, S.; Toppino, L.; Rotino, G.L. Scrapped but Not Neglected: Insights into the Composition, Molecular Modulation and Antioxidant Capacity of Phenols in Peel of Eggplant (Solanum melongena L.) Fruits at Different Developmental Stages. Plant Physiol. Biochem. 2021, 167, 678–690. [Google Scholar] [CrossRef]
- Cocaliadis, M.F.; Fernández-Muñoz, R.; Pons, C.; Orzaez, D.; Granell, A. Increasing Tomato Fruit Quality by Enhancing Fruit Chloroplast Function. A Double-Edged Sword? J. Exp. Bot. 2014, 65, 4589–4598. [Google Scholar] [CrossRef]
- Lado, J.; Zacarías, L.; Gurrea, A.; Page, A.; Stead, A.; Rodrigo, M.J. Exploring the Diversity in Citrus Fruit Colouration to Decipher the Relationship Between Plastid Ultrastructure and Carotenoid Composition. Planta 2015, 242, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Martí, M.; Camejo, D.; Olmos, E.; Sandalio, L.; Fernández-García, N.; Jiménez, A.; Sevilla, F. Characterisation and Changes in the Antioxidant System of Chloroplasts and Chromoplasts Isolated from Green and Mature Pepper Fruits. Plant Biol. 2009, 11, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Munné-Bosch, S. Photo-Oxidative Stress During Leaf, Flower and Fruit Development. Plant Physiol. 2018, 176, 1004–1014. [Google Scholar] [CrossRef]
- Maulucci, G.; Bačić, G.; Bridal, L.; Schmidt, H.H.; Tavitian, B.; Viel, T.; Utsumi, H.; Yalçın, A.S.; De Spirito, M. Imaging Reactive Oxygen Species-Induced Modifications in Living Systems. Antioxid. Redox Signal. 2016, 24, 939–958. [Google Scholar] [CrossRef]
- Steffen-Heins, A.; Steffens, B. EPR Spectroscopy and its Use in Planta—A Promising Technique to Disentangle the Origin of Specific ROS. Front. Environ. Sci. 2015, 3, 15. [Google Scholar] [CrossRef]
- Krishna, M.C.; Grahame, D.A.; Samuni, A.; Mitchell, J.B.; Russo, A. Oxoammonium Cation Intermediate in the Nitroxide-Catalyzed Dismutation of Superoxide. Proc. Natl. Acad. Sci. USA 1992, 89, 5537–5541. [Google Scholar] [CrossRef]
- Dikalov, S.; Grigor’ev, I.; Voinov, M.; Bassenge, E. Detection of Superoxide Radicals and Peroxynitrite by 1-Hydroxy-4-Phosphonooxy-2, 2, 6, 6-Tetramethylpiperidine: Quantification of Extracellular Superoxide Radicals Formation. Biochem. Biophys. Res. Commun. 1998, 248, 211–215. [Google Scholar] [CrossRef]
- Nakagawa, K.; Promjareet, A.; Priprem, A.; Netweera, V.; Hara, H. Investigation of Scavenging Activities and Distribution of Paramagnetic Species in Zanthoxylum limonella Seeds. Free. Radic. Res. 2016, 50, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Maeda, H. EPR Imaging and HPLC Characterization of the Pigment-Based Organic Free Radical in Black Soybean Seeds. Free. Radic. Res. 2017, 51, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Epel, B. Investigating the Distribution of Stable Paramagnetic Species in an Apple Seed Using X-Band EPR and EPR Imaging. J. Oleo Sci. 2017, 66, 315–319. [Google Scholar] [CrossRef]
- Ilangovan, G.; Li, H.; Zweier, J.L.; Krishna, M.C.; Mitchell, J.B.; Kuppusamy, P. In Vivo Measurement of Regional Oxygenation and Imaging of Redox Status in RIF-1 Murine Tumor: Effect of Carbogen-Breathing. Magn. Reson. Med. Off. J. Int. Soc. Magn. Reson. Med. 2002, 48, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, P.; Afeworki, M.; Shankar, R.A.; Coffin, D.; Krishna, M.C.; Hahn, S.M.; Mitchell, J.B.; Zweier, J.L. In Vivo Electron Paramagnetic Resonance Imaging of Tumor Heterogeneity and Oxygenation in a Murine Model. Cancer Res. 1998, 58, 1562–1568. [Google Scholar]
- Kuppusamy, P.; Li, H.; Ilangovan, G.; Cardounel, A.J.; Zweier, J.L.; Yamada, K.; Krishna, M.C.; Mitchell, J.B. Noninvasive Imaging of Tumor Redox Status and its Modification by Tissue Glutathione Levels. Cancer Res. 2002, 62, 307–312. [Google Scholar]
- Yamada, K.I.; Kuppusamy, P.; English, S.; Yoo, J.; Irie, A.; Subramanian, S.; Mitchell, J.B.; Krishna, M.C. Feasibility and Assessment of Non-Invasive In Vivo Redox Status Using Electron Paramagnetic Resonance Imaging. Acta Radiol. 2002, 43, 433–440. [Google Scholar] [CrossRef]
- Utsumi, H.; Yamada, K.-I. In Vivo Electron Spin Resonance-Computed Tomography/Nitroxyl Probe Technique for Non-Invasive Analysis of Oxidative Injuries. Arch. Biochem. Biophys. 2003, 416, 1–8. [Google Scholar] [CrossRef]
- Samuni, A.; Goldstein, S.; Russo, A.; Mitchell, J.B.; Krishna, M.C.; Neta, P. Kinetics and Mechanism of Hydroxyl Radical and OH-Adduct Radical Reactions with Nitroxides and with their Hydroxylamines. J. Am. Chem. Soc. 2002, 124, 8719–8724. [Google Scholar] [CrossRef]
- Krishna, M.C.; Russo, A.; Mitchell, J.B.; Goldstein, S.; Dafni, H.; Samuni, A. Do Nitroxide Antioxidants Act as Scavengers of O2− or as SOD Mimics? J. Biol. Chem. 1996, 271, 26026–26031. [Google Scholar] [CrossRef]
- Takeshita, K.; Saito, K.; Ueda, J.-i.; Anzai, K.; Ozawa, T. Kinetic Study on ESR Signal Decay of Nitroxyl Radicals, Potent Redox Probes for In Vivo ESR Spectroscopy, Caused by Reactive Oxygen Species. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2002, 1573, 156–164. [Google Scholar] [CrossRef]
- Guo, D.L.; Wang, Z.G.; Pei, M.S.; Guo, L.L.; Yu, Y.H. Transcriptome Analysis Reveals Mechanism of Early Ripening in Kyoho Grape with Hydrogen Peroxide Treatment. BMC Genom. 2020, 21, 784. [Google Scholar] [CrossRef]
- Ren, Y.; He, J.; Liu, H.; Liu, G.; Ren, X. Nitric Oxide Alleviates Deterioration and Preserves Antioxidant Properties in ‘Tainong’ Mango Fruit During Ripening. Hortic.Environ. Biotechnol. 2017, 58, 27–37. [Google Scholar] [CrossRef]
- Qin, G.; Meng, X.; Wang, Q.; Tian, S. Oxidative Damage of Mitochondrial Proteins Contributes to Fruit Senescence: A Redox Proteomics Analysis. J. Proteome Res. 2009, 8, 2449–2462. [Google Scholar] [CrossRef] [PubMed]
- Lacan, D.; Baccou, J.-C. High Levels of Antioxidant Enzymes Correlate with Delayed Senescence in Nonnetted Muskmelon Fruits. Planta 1998, 204, 377–382. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen Peroxide- and Glutathione-Associated Mechanisms of Acclimatory Stress Tolerance and Signalling. Physiol. Plant. 1997, 100, 241–254. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of Superoxide Dismutases (SODs) in Controlling Oxidative Stress in Plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- López-Huertas, E.; Palma, J.M. Changes in Glutathione, Ascorbate, and Antioxidant Enzymes during Olive Fruit Ripening. J. Agric. Food Chem. 2020, 68, 12221–12228. [Google Scholar] [CrossRef]
- Mondal, K.; Sharma, N.S.; Malhotra, S.P.; Dhawan, K.; Singh, R. Antioxidant Systems in Ripening Tomato Fruits. Biol. Plant. 2004, 48, 49–53. [Google Scholar] [CrossRef]
- Jiménez, A.; Romojaro, F.; Gómez, J.M.; Llanos, M.R.; Sevilla, F. Antioxidant Systems and their Relationship with the Response of Pepper Fruits to Storage at 20 Degrees C. J. Agric. Food Chem. 2003, 51, 6293–6299. [Google Scholar] [CrossRef]
- Batista, K.A.; Batista, G.L.A.; Alves, G.L.; Fernandes, K.F. Extraction, Partial Purification and Characterization of Polyphenol Oxidase from Solanum lycocarpum Fruits. J. Mol. Catal. B-Enzym. 2014, 102, 211–217. [Google Scholar] [CrossRef]
- Queiroz, C.; Mendes Lopes, M.L.; Fialho, E.; Valente-Mesquita, V.L. Polyphenol Oxidase: Characteristics and Mechanisms of Browning Control. Food Rev. Int. 2008, 24, 361–375. [Google Scholar] [CrossRef]
- Mishra, B.B.; Gautam, S.; Sharma, A. Free Phenolics and Polyphenol Oxidase (PPO): The Factors Affecting Post-Cut Browning in Eggplant (Solanum melongena). Food Chem. 2013, 139, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Maioli, A.; Gianoglio, S.; Moglia, A.; Acquadro, A.; Valentino, D.; Milani, A.M.; Prohens, J.; Orzaez, D.; Granell, A.; Lanteri, S.; et al. Simultaneous CRISPR/Cas9 Editing of Three PPO Genes Reduces Fruit Flesh Browning in Solanum melongena L. Front. Plant Sci. 2020, 11, 607161. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compound Name | tR, min | Molecular Formula, [M–H]– | Calculated Mass, [M–H]– | Exact Mass, [M–H]– | Δ mDa | MS2 Fragments, (% Base Peak) | MS3 Fragments, (% Base Peak) | Ref |
|---|---|---|---|---|---|---|---|---|---|
| Phenolic acids | |||||||||
| 1 | Vanillic acid hexoside | 3.46 | C14H17O9– | 329.08781 | 329.08420 | 3.61 | 123(48), 161(100), 169(22), 282(24), 283(30), 284(24), 285(37) | 59(9), 71(15), 85(12), 97(11), 101(100), 113(38), 143(19) | [24] |
| 2 | Dihydroxybenzoic acid hexoside | 5.74 | C13H15O9– | 315.07216 | 315.06883 | 3.33 | 108(11), 109(12), 152(43), 153(100), 154(8), 163(9), 165(12) | 81(10), 108(9), 109(100) | [9] |
| 3 | Caffeoylquinic acid hexoside 1 | 5.96 | C22H27O14– | 515.14063 | 515.13512 | 5.51 | 179(4), 191(100), 192(7), 323(6), 341(7), 353(87), 354(11) | 85(100), 93(47), 109(23), 111(40), 127(91), 171(20), 173(55) | [7] |
| 4 | Caffeoylquinic acid hexoside 2 | 6.47 | C22H27O14– | 515.14063 | 515.13454 | 6.09 | 179(8), 191(31), 323(100), 324(15), 341(26), 353(21), 455(6) | 133(5), 161(100) | [7] |
| 5 | Caffeic acid hexoside 1 | 6.50 | C15H17O9– | 341.08781 | 341.08383 | 3.98 | 135(6), 161(27), 179(100), 180(4), 203(5), 281(3) | 107(20), 135(100) | [6] |
| 6 | Caffeoylquinic acid hexoside 3 | 6.64 | C22H27O14– | 515.14063 | 515.13471 | 5.92 | 179(4), 191(100), 192(6), 341(7), 353(49), 354(7), 395(14) | 85(100), 93(43), 109(20), 111(33), 127(84), 171(28), 173(61) | [7] |
| 7 | Hydroxybenzoic acid hexoside | 6.78 | C13H15O8– | 299.07724 | 299.07399 | 3.25 | 137(100) | 93(100) | [9] |
| 8 | Feruloylquinic acid hexoside b | 6.87 | C23H29O14– | 529.15628 | 529.15155 | 4.73 | 191(100), 192(7), 193(8), 365(6), 367(79), 368(11), 409(12) | 85(100), 93(52), 109(17), 111(32), 127(76), 171(18), 173(63) | [25] |
| 9 | Caffeic acid hexoside 2 | 6.87 | C15H17O9– | 341.08781 | 341.08375 | 4.06 | 135(8), 179(100), 180(8) | 107(22), 135(100) | [6] |
| 10 | 5-O-Caffeoylquinic acid a | 6.92 | C16H17O9– | 353.08781 | 353.08312 | 4.68 | 179(3), 191(100), 192(3) | 85(99), 93(64), 109(23), 111(41), 127(100), 171(31), 173(68) | [7] |
| 11 | Caffeic acid a | 7.04 | C9H7O4– | 179.03498 | 179.03301 | 1.98 | 135(100) | 91(68), 107(100), 117(59), 135(72) | [26] |
| 12 | p-Coumaric acid a | 7.08 | C9H7O3– | 163.04007 | 163.03862 | 1.45 | 103(24), 118(14), 119(100), 120(14), 121(19), 135(29), 136(26) | 91(100) | [27] |
| 13 | 5-O-Caffeoylquinic acid isomer | 7.45 | C16H17O9– | 353.08781 | 353.08321 | 4.59 | 179(3), 191(100), 192(4) | 85(100), 87(19), 93(55), 111(31), 127(88), 171(28), 173(61) | [7] |
| 14 | 3-O-Coumaroylquinic acid | 7.69 | C16H17O8– | 337.09289 | 337.08872 | 4.17 | 135(3), 163(5), 173(32), 179(13), 191(100), 192(3) | 85(89), 87(21), 93(59), 111(33), 127(100), 171(28), 173(64) | [8] |
| 15 | 5-O-Caffeoylshikimic acid | 7.72 | C16H15O8– | 335.07724 | 335.07352 | 3.73 | 135(28), 161(3), 179(100), 180(7), 191(3) | 107(18), 135(100) | [27] |
| 16 | 3-O-Feruloylquinic acid | 7.97 | C17H19O9– | 367.10346 | 367.09929 | 4.17 | 173(22), 191(100), 192(7), 193(7), 203(6) | 85(87), 93(65), 109(23), 111(37), 127(100), 171(27), 173(62) | [9] |
| 17 | 5-O-Coumaroylquinic acid | 8.11 | C16H17O8– | 337.09289 | 337.08928 | 3.61 | 135(3), 163(5), 173(4), 179(10), 191(100), 192(3) | 85(100), 87(19), 93(64), 111(31), 127(90), 171(21), 173(75) | [8] |
| 18 | 5-O-Feruloylquinic acid | 8.27 | C17H19O9– | 367.10346 | 367.09963 | 3.82 | 191(100), 193(3) | 85(97), 93(58), 109(22), 111(36), 127(100), 171(23), 173(68) | [9] |
| 19 | Dicaffeoylquinic acid | 9.04 | C25H23O12– | 515.11950 | 515.11530 | 4.20 | 173(7), 179(4), 203(11), 255(5), 299(9), 353(100), 354(10) | 93(20), 135(10), 173(100), 179(62), 191(30) | [7] |
| Phenolic amides | |||||||||
| 20 | N-Caffeoylputrescine 1 | 4.87 | C13H17N2O3– | 249.12447 | 249.12179 | 2.68 | 135(100), 136(8), 175(5), 176(5), 207(24), 208(3), 249(4) | 91(26), 93(36), 106(5), 107(100), 117(22), 135(3) | [9] |
| 21 | Pantothenic acid | 5.73 | C9H16NO5– | 218.10340 | 218.10075 | 2.65 | 88(100), 89(5), 129(6), 143(8), 144(3), 146(12), 173(10) | 59(100) | [28] |
| 22 | N-Caffeoylputrescine 2 | 5.86 | C13H17N2O3– | 249.12447 | 249.12186 | 2.61 | 135(100), 136(5), 207(17), 249(3) | 79(45), 91(5), 93(93), 107(100), 117(11), 135(25) | [9] |
| 23 | N1,N14-bis-Dihydrocaffeoylspermine (kukoamine A) | 6.03 | C28H41N4O6– | 529.30316 | 529.29671 | 6.45 | 365(100), 366(21), 367(3), 408(53), 408(11), 419(3), 511(3) | 115(3), 121(35), 122(3), 137(7), 243(100), 244(12), 323(6) | [7] |
| 24 | N1-Caffeoy-N14-dihydrocaffeoylspermine | 6.10 | C28H39N4O6– | 527.28751 | 527.28172 | 5.79 | 366(100), 391(12), 407(51), 420(4) | 109(3), 115(4), 121(33), 137(10), 243(100), 323(5) | [7] |
| 25 | N-Feruloyltyramine 1 | 8.19 | C18H18NO4– | 312.12413 | 312.12114 | 2.99 | 135(66), 148(19), 176(15), 177(12), 178(100), 270(11), 297(68) | 135(100), 136(19) | [11] |
| 26 | N-Caffeoyloctopamine | 8.65 | C17H16NO5– | 314.10340 | 314.10003 | 3.37 | 135(57), 150(16), 152(20), 161(100), 162(9), 178(24), 192(19) | 133(100) | [29] |
| 27 | Acetyl tryptophan | 8.84 | C13H13N2O3– | 245.09317 | 245.09135 | 1.82 | 74(3), 116(5), 201(3), 203(100), 204(10) | 116(40), 129(10), 142(17), 159(100), 186(7) | [30] |
| 28 | N-Feruloyltyramine 2 | 10.27 | C18H18NO4– | 312.12413 | 312.12173 | 2.40 | 135(63), 148(15), 176(15), 177(15), 178(100), 297(56), 298(11) | 93(20), 135(100), 136(13), 160(8) | [11] |
| 29 | Grossamide | 11.89 | C36H35N2O8– | 623.23989 | 623.23378 | 6.11 | 297(29), 432(9), 460(100), 461(28), 486(16), 591(26), 592(10) | 282(9), 283(18), 297(100), 298(5), 323(5), 445(16), 446(3) | [31] |
| Flavonoids | |||||||||
| 30 | Quercetin-3-O-rutinoside a | 7.99 | C27H29O16– | 609.14611 | 609.14096 | 5.15 | 255(4), 271(7), 300(12), 301(100), 302(10), 343(5) | 151(83), 179(100), 255(58), 257(12), 271(95), 272(25), 273(19) | [8] |
| 31 | Kaempferol-3-O-rutinoside | 8.47 | C27H29O15– | 593.15119 | 593.14561 | 5.58 | 229(3), 255(3), 257(4), 285(15), 285(100), 286(14), 327(3) | 197(18), 199(19), 213(24), 229(53), 241(31), 257(100), 267(43) | [8] |
| 32 | Dihydrokaempferol | 9.85 | C15H11O6– | 287.05611 | 287.05401 | 2.10 | 201(3), 243(9), 259(100), 260(9), 269(4) | 125(60), 151(17), 165(11), 172(16), 173(33), 215(100), 241(19) | [32] |
| Saponins | |||||||||
| 33 | Saponin derivative 1 b | 8.52 | C66H99O30– | 1371.62267 | 1371.62202 | 0.64 | 1048(21), 1064(40), 1065(14), 1210(100), 1211(37), 1226(52) | 755(8), 884(7), 901(55), 918(7), 1046(6), 1048(100), 1064(78) | [33] |
| 34 | Saponin derivative 2 b | 8.71 | C54H81O23– | 1097.51741 | 1097.51286 | 4.55 | 773(13), 934(15), 936(100), 937(26), 1050(9), 1051(5), 1078(8) | 594(5), 611(38), 755(7), 773(100), 774(9) | [34] |
| 35 | Saponin derivative 3 b | 8.83 | C66H99O30– | 1371.62267 | 1371.61563 | 7.04 | 1064(28), 1080(13), 1210(47), 1211(21), 1226(100), 1227(51) | 738(3), 756(10), 900(3), 902(28), 917(30), 1063(100), 1080(94) | [33] |
| 36 | Solanigroside Y6 | 8.97 | C57H93O27– | 1209.59097 | 1209.58649 | 4.48 | 902(45), 1048(100), 1049(47), 1050(29), 1064(65), 1065(37) | 738(3), 755(20), 884(10), 886(4), 902(100), 903(6) | [35] |
| 37 | Saponin derivative 4 b | 9.21 | C45H69O20– | 929.43877 | 929.43736 | 1.41 | 750(4), 767(4), 883(100), 884(44), 911(7), 912(4), 914(4) | 574(20), 720(7), 721(100), 737(6) | [36] |
| 38 | Agamenoside A | 9.21 | C56H91O28– | 1211.57024 | 1211.56963 | 0.61 | 756(6), 918(22), 919(9), 1050(38), 1051(15), 1080(100), 1081(21) | 756(17), 917(100), 919(4) | [37] |
| 39 | Scopoloside I | 9.30 | C45H71O20– | 931.45442 | 931.45039 | 4.03 | 733(23), 752(62), 753(21), 770(48), 771(24), 913(100), 915(38) | 500(17), 575(8), 708(11), 733(100), 751(21), 869(30), 895(28) | [38] |
| 40 | Indioside D | 9.38 | C51H83O23– | 1063.53306 | 1063.52670 | 6.37 | 756(17), 900(10), 902(100), 903(16), 916(8), 918(46), 919(9) | 593(20), 740(7), 755(100), 757(4) | [39] |
| 41 | Protodioscin | 9.51 | C51H83O22– | 1047.53815 | 1047.53540 | 2.75 | 756(16), 757(14), 884(9), 885(8), 902(100), 903(77), 904(46) | 576(7), 593(4), 738(18), 739(3), 740(3), 756(100), 757(12) | [35] |
| 42 | Melongoside N | 9.57 | C45H75O19– | 919.49080 | 919.48480 | 6.00 | 596(4), 756(8), 758(100), 758(18) | 434(6), 595(100), 596(7) | [7] |
| 43 | Saponin derivative 5 | 9.59 | C45H73O18– | 901.48024 | 901.47764 | 2.60 | 738(3), 740(4), 741(3), 756(100), 757(37), 758(16) | 413(25), 432(21), 575(100), 576(10), 593(80), 595(15), 738(7) | [40] |
| 44 | Soladulcoside A | 9.90 | C39H61O15– | 769.40160 | 769.40011 | 1.48 | 431(3), 710(12), 711(4), 734(9), 752(100), 753(34), 754(8) | 413(81), 431(26), 546(61), 575(27), 589(38), 707(61), 734(100) | [41] |
| 45 | Solaviaside A | 9.97 | C51H85O21– | 1033.55888 | 1033.55842 | 0.46 | 870(19), 885(14), 886(60), 887(24), 888(100), 889(43), 890(15) | 561(20), 724(9), 725(5), 742(100), 742(7) | [42] |
| 46 | Solasodoside A | 10.64 | C51H81O21– | 1029.52758 | 1029.52354 | 4.04 | 737(8), 866(22), 867(9), 884(100), 885(27), 886(33), 887(21) | 557(15), 558(3), 719(3), 722(15), 737(33), 738(100), 866(4) | [43] |
| 47 | Saponin derivative 6 | 10.88 | C39H63O14– | 755.42233 | 755.41563 | 6.70 | 696(16), 697(6), 720(11), 721(4), 737(100), 739(40), 740(8) | 512(11), 561(12), 694(10), 708(9), 720(100), 721(21), 1472(17) | [44] |
| 48 | Saponin derivative 7 | 10.88 | C45H69O18– | 897.44894 | 897.44657 | 2.37 | 605(14), 607(3), 734(14), 735(5), 751(100), 752(21) | 425(16), 443(5), 587(23), 606(100) | [40] |
| Glycoalkaloids | |||||||||
| 49 | Solanidenediol triose derivative 1 | 7.26 | C45H72NO18– | 914.47549 | 914.46822 | 7.27 | 753(100), 754(57), 755(27), 756(5), 768(3), 769(3) | 247(35), 307(6), 500(7), 540(72), 552(14), 606(10), 606(100) | [45] |
| 50 | Solanidenediol triose derivative 2 | 7.60 | C45H72NO17– | 898.48057 | 898.47411 | 6.46 | 540(5), 734(3), 750(3), 753(100), 754(43) | 247(100), 442(7), 557(14), 606(41) | [46] |
| 51 | Solanandaine | 7.90 | C45H72NO16– | 882.48566 | 882.48345 | 2.21 | 737(100) | 247(53), 306(7), 428(63), 525(68), 554(11), 590(100) | [7] |
| 52 | Solanidenediol triose derivative 3 | 8.03 | C45H72NO18– | 914.47549 | 914.46579 | 9.70 | 751(16), 752(56), 753(100), 754(10), 768(6), 768(5), 867(12) | 607(4), 722(100), 734(13), 735(3) | [45] |
| 53 | Solanidenediol triose derivative 4 | 8.16 | C45H72NO17– | 898.48057 | 898.47398 | 6.59 | 576(3), 737(37), 738(100), 739(14) | 574(34), 576(100), 591(3), 592(3) | [46] |
| 54 | β-Tomatine | 8.25 | C45H74NO17– | 900.49622 | 900.48694 | 9.28 | 577(12), 578(15), 736(5), 737(41), 738(19), 739(100), 740(89) | 576(100), 578(32) | [10] |
| 55 | Solanandaine isomer | 8.36 | C45H72NO16– | 882.48566 | 882.48044 | 5.22 | 736(100), 737(54), 738(38), 740(14) | 247(100), 307(26), 572(30), 589(24), 590(18), 1422(31) | [7] |
| 56 | β2-Solasonine | 8.36 | C39H62NO12– | 736.42775 | 736.42544 | 2.31 | 574(100) | 410(16), 410(14), 412(15), 454(18), 468(49), 483(100), 509(78) | [10] |
| 57 | α-Solasonine | 8.96 | C45H72NO16– | 882.48566 | 882.47982 | 5.84 | 720(9), 721(100), 722(51), 723(28), 724(7), 736(4) | 307(4), 497(6), 508(27), 515(14), 520(6), 556(4), 574(100) | [7] |
| 58 | γ-Solamarine | 9.13 | C39H62NO11– | 720.43284 | 720.42971 | 3.12 | 246(4), 247(17), 307(6), 358(6), 508(13), 574(100), 575(12) | 179(100) | [10] |
| 59 | α-Solanine | 9.13 | C45H72NO15– | 866.49020 | 866.48811 | 2.09 | 721(100), 722(87), 722(50), 723(10) | 247(24), 265(6), 568(6), 574(100), 1373(5), 1393(4) | [9] |
| 60 | Abutiloside H | 9.15 | C46H74NO17– | 912.49622 | 912.48906 | 7.16 | 867(100), 868(8) | 720(100) | [47] |
| 61 | Leptine II | 11.60 | C47H74NO17– | 924.49622 | 924.48626 | 9.96 | 763(33), 764(100), 764(5), 779(4), 780(3), 878(3) | 246(12), 437(56), 551(43), 616(11), 617(100), 746(26), 746(94) | [9] |
| 62 | Leptine I | 11.77 | C47H74NO16– | 908.50131 | 908.49170 | 9.61 | 550(4), 762(58), 763(100), 861(5), 863(8), 881(3), 892(4) | 247(100), 599(85), 617(65) | [9] |
| Lignans | |||||||||
| 63 | Alangilignoside C | 7.93 | C28H37O13– | 581.22396 | 581.21837 | 5.59 | 233(13), 373(13), 419(100), 420(19), 533(25), 535(26), 566(12) | 223(4), 373(7), 389(3), 404(100), 405(4) | [48] |
| 64 | Alangilignoside C isomer | 8.44 | C28H37O13– | 581.22396 | 581.21959 | 4.37 | 179(7), 401(100), 402(17), 419(7), 533(43), 534(13), 535(6) | 205(16), 220(49), 235(61), 356(37), 368(13), 371(100), 386(91) | [48] |
| 65 | Isolariciresinol 3-O-hexoside | 8.61 | C26H33O11– | 521.20284 | 521.19756 | 5.28 | 341(12), 359(9), 473(10), 474(21), 475(100), 476(31), 477(8) | 191(31), 253(56), 331(31), 343(96), 407(38), 415(100), 433(99) | [49] |
| 66 | Pinoresinol 3-O-hexoside | 8.74 | C26H31O11– | 519.18719 | 519.18198 | 5.21 | 357(100), 358(17) | 136(33), 151(100), 175(3), 311(14), 342(8) | [50] |
| 67 | Pinoresinol | 8.75 | C20H21O6– | 357.13436 | 357.13017 | 4.19 | 136(33), 151(100), 175(3), 311(14), 342(8) | 136(100) | [51] |
| 68 | Syringaresinol | 8.78 | C22H25O8– | 417.15549 | 417.15102 | 4.47 | 151(16), 166(30), 179(13), 181(100), 353(10), 371(15), 402(44) | 166(100) | [52] |
| Other compounds | |||||||||
| 69 | Quinic acid a | 1.61 | C7H11O6– | 191.05611 | 191.04617 | 9.94 | 85(100), 87(15), 93(41), 111(60), 127(61), 171(27), 173(46) | 57(100) | [31] |
| 70 | Aconitic acid a | 1.67 | C6H5O6– | 173.00861 | 173.00753 | 1.08 | 111(100), 112(9), 129(5), 143(7), 155(17) | 67(100) | [6] |
| 71 | Butanediol pentosyl-hexoside b | 5.25 | C15H27O11– | 383.15589 | 383.15155 | 4.34 | 161(11), 191(10), 251(100), 336(6), 337(11), 338(11), 346(11) | 85(4), 97(3), 101(18), 113(15), 161(100) | [53] |
| 72 | Phenethyl pentosyl-hexoside | 6.46 | C19H27O10– | 415.16097 | 415.15567 | 5.30 | 173(9), 191(100), 192(11), 235(12), 253(14), 367(6), 397(6) | 123(24), 133(41), 136(72), 149(100), 173(96), 176(35) | [54] |
| 73 | Benzyl pentosyl-hexoside | 7.25 | C18H25O10– | 401.14532 | 401.14150 | 3.82 | 175(11), 191(100), 269(13), 353(13), 354(14), 355(24), 379(16) | 113(3), 127(3),148(20), 176(100) | [55] |
| 74 | Tuberonic acid hexoside | 7.37 | C18H27O9– | 387.16606 | 387.16206 | 4.00 | 163(46), 191(13), 205(50), 207(74), 223(100), 247(49) | 164(100), 179(22), 208(36) | [9] |
| 75 | Roseoside A | 7.58 | C19H29O8– | 385.18679 | 385.18185 | 4.94 | 113(5), 143(4), 153(100), 161(26), 179(4), 205(76), 223(59) | 95(7), 97(14), 109(7), 111(5), 137(18), 138(100) | [9] |
| 76 | Methylbutyl pentosyl-hexoside | 7.66 | C16H27O10– | 381.17662 | 381.17180 | 4.82 | 161(6), 217(4), 249(100), 250(4), 336(3) | 83(7), 85(11), 99(12), 101(69), 113(20), 159(23), 161(100) | [6] |
| 77 | Hexenyl pentosyl-hexoside | 7.92 | C17H29O10– | 393.17662 | 393.17246 | 4.16 | 131(25), 149(100), 173(16), 191(73), 347(12), 353(57), 355(21) | 88(3), 89(89), 113(14), 131(100), 134(6) | [56] |
| 78 | Deacetylasperuloside | 8.01 | C16H19O10– | 371.09837 | 371.09353 | 4.84 | 121(4), 191(18), 192(12), 193(22), 231(6), 249(100), 250(4) | 85(27), 95(15), 99(11), 111(10), 113(100), 175(14), 231(82) | [9] |
| 79 | Abscisic acid a | 8.65 | C15H19O4– | 263.12888 | 263.12637 | 2.51 | 147(10), 153(100), 216(16), 217(23), 218(32), 219(46), 246(15) | 105(4), 111(8), 123(4), 124(14), 136(15), 138(100) | [57] |
| 80 | Campherenane diol dihexoside malonate ester | 10.97 | C24H39O10– | 487.25487 | 487.24932 | 5.55 | 161(10), 221(16), 440(15), 441(100), 442(4), 443(11) | 143(11), 147(8), 161(47), 221(10), 237(3), 321(4), 381(100) | [58] |
| 81 | Trihydroxyoctadecenoic acid | 11.72 | C18H33O5– | 329.23335 | 329.22954 | 3.81 | 171(100), 172(9), 201(6), 211(15), 229(22), 293(14), 311(19) | 125(62), 127(100), 153(99), 171(3) | [9] |
| 82 | 9,10,11-Trihydroxy-12, 15-octadecadienoic acid | 11.89 | C18H31O5– | 327.21770 | 327.21369 | 4.01 | 171(100), 172(7), 183(10), 201(36), 213(10), 291(12), 309(34) | 97(5), 123(3), 125(55), 127(100), 153(95) | [9] |
| 83 | 9,12,13-Trihydroxy-10-octadecenoic acid | 12.54 | C18H33O5– | 329.23335 | 329.22951 | 3.84 | 171(100), 172(8), 201(81), 202(8), 275(18), 293(19), 311(28) | 123(3), 125(71), 127(100), 153(78) | [9] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milutinović, M.; Nakarada, Đ.; Božunović, J.; Todorović, M.; Gašić, U.; Živković, S.; Skorić, M.; Ivković, Đ.; Savić, J.; Devrnja, N.; et al. Solanum dulcamara L. Berries: A Convenient Model System to Study Redox Processes in Relation to Fruit Ripening. Antioxidants 2023, 12, 346. https://doi.org/10.3390/antiox12020346
Milutinović M, Nakarada Đ, Božunović J, Todorović M, Gašić U, Živković S, Skorić M, Ivković Đ, Savić J, Devrnja N, et al. Solanum dulcamara L. Berries: A Convenient Model System to Study Redox Processes in Relation to Fruit Ripening. Antioxidants. 2023; 12(2):346. https://doi.org/10.3390/antiox12020346
Chicago/Turabian StyleMilutinović, Milica, Đura Nakarada, Jelena Božunović, Miloš Todorović, Uroš Gašić, Suzana Živković, Marijana Skorić, Đurđa Ivković, Jelena Savić, Nina Devrnja, and et al. 2023. "Solanum dulcamara L. Berries: A Convenient Model System to Study Redox Processes in Relation to Fruit Ripening" Antioxidants 12, no. 2: 346. https://doi.org/10.3390/antiox12020346
APA StyleMilutinović, M., Nakarada, Đ., Božunović, J., Todorović, M., Gašić, U., Živković, S., Skorić, M., Ivković, Đ., Savić, J., Devrnja, N., Aničić, N., Banjanac, T., Mojović, M., & Mišić, D. (2023). Solanum dulcamara L. Berries: A Convenient Model System to Study Redox Processes in Relation to Fruit Ripening. Antioxidants, 12(2), 346. https://doi.org/10.3390/antiox12020346