
Effects of UV Stress in Promoting Antioxidant Activities in Fungal Species Тrametes versicolor (L.) Lloyd and Flammulina velutipes (Curtis) Singer

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Material
2.2. Submerged Cultivation and Preparation of Extracts
2.3. LC-MS/MS Quantification of Selected Phenolic Compounds
2.4. Determination of Total Phenolic (TP) and Total Protein (TPR) Contents
2.5. DPPH Radical Scavenging Assay
2.6. Hydroxyl Radical (OH•) Scavenging Assay
2.7. Superoxide Anion Radical Scavenging Assay (SOA)
2.8. Ferrous Ion Chelating Assay (FRAP)
2.9. Statistical Analysis
3. Results аnd Discussion
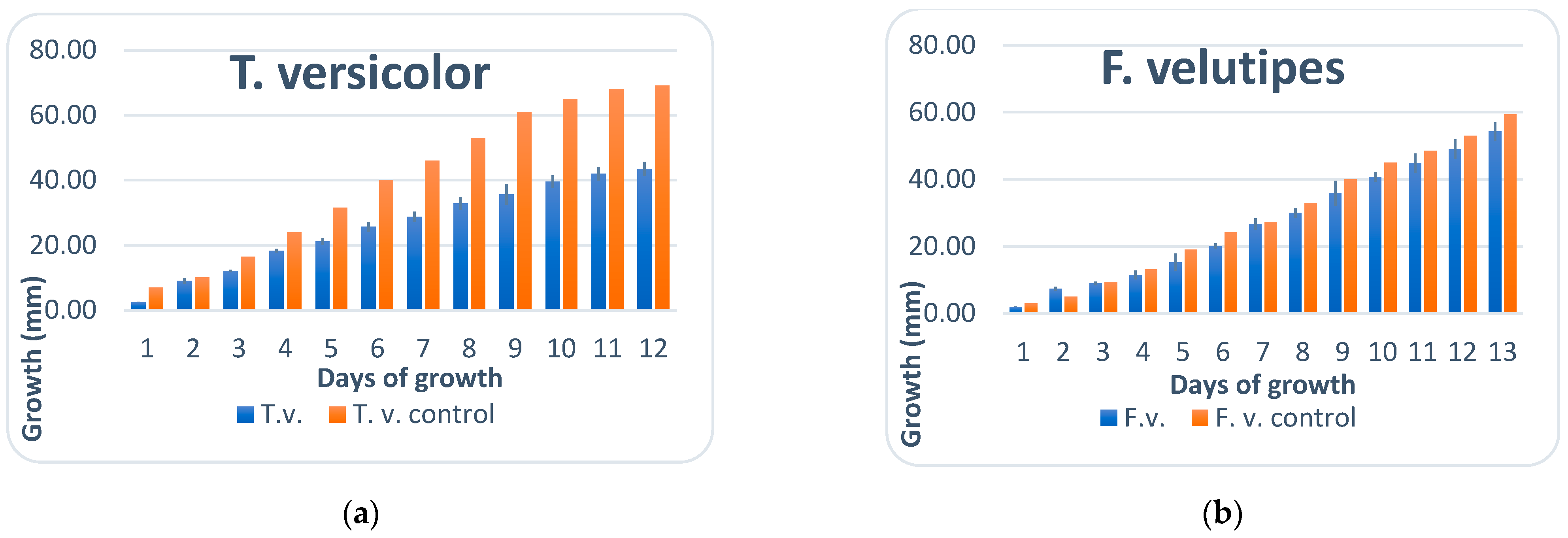
3.1. Mycelial Growth
3.2. Mycochemical Characterization
3.2.1. LC–MS/MS Analysis
3.2.2. Total Phenolic Content (TP)
3.2.3. Total Protein Content (TPR)
3.3. In Vitro AO Activity
3.3.1. DPPH Assay
3.3.2. SOA Assay
3.3.3. OH Assay
3.3.4. FRAP Assay
3.4. Correlation Analysis
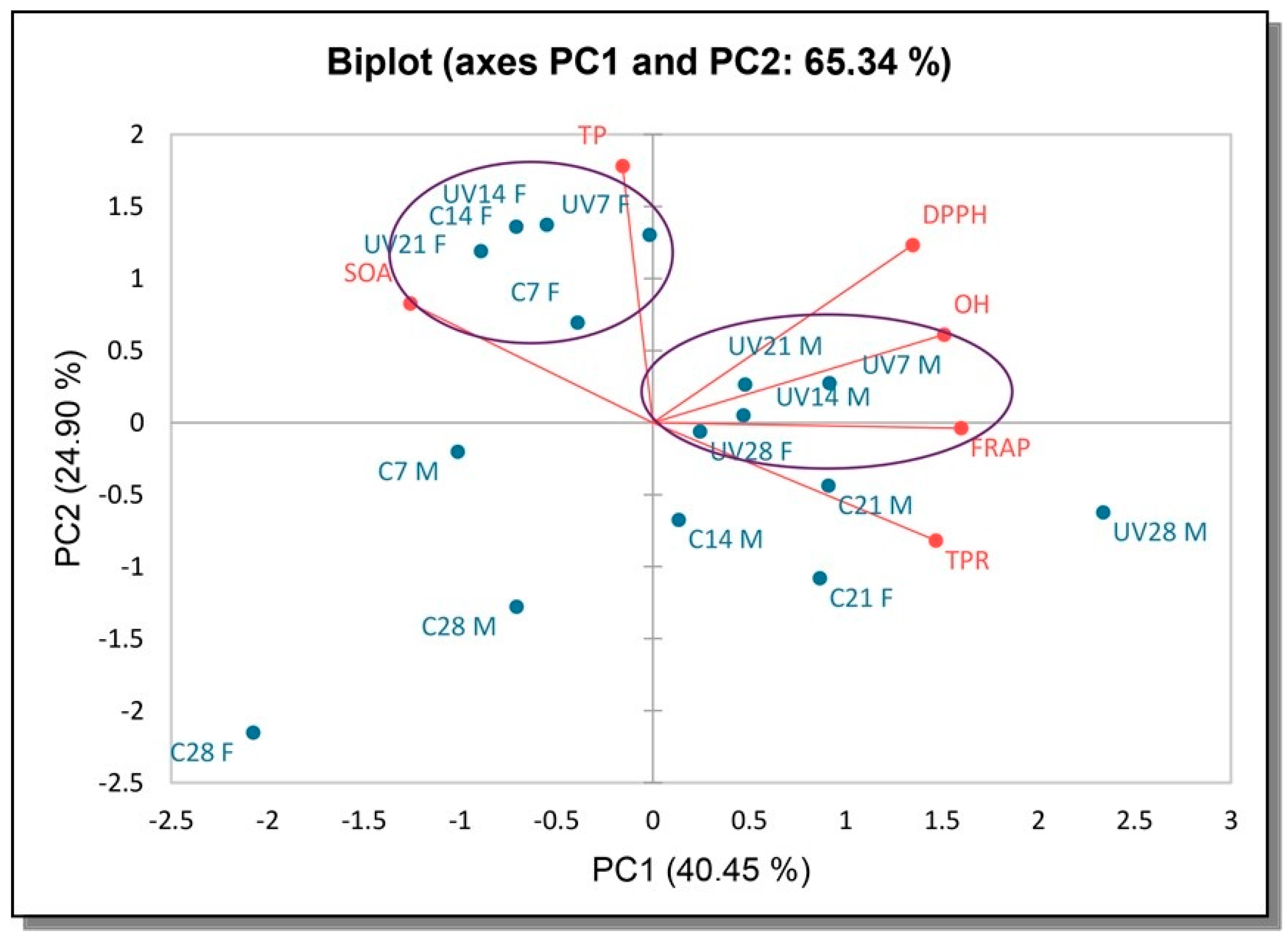
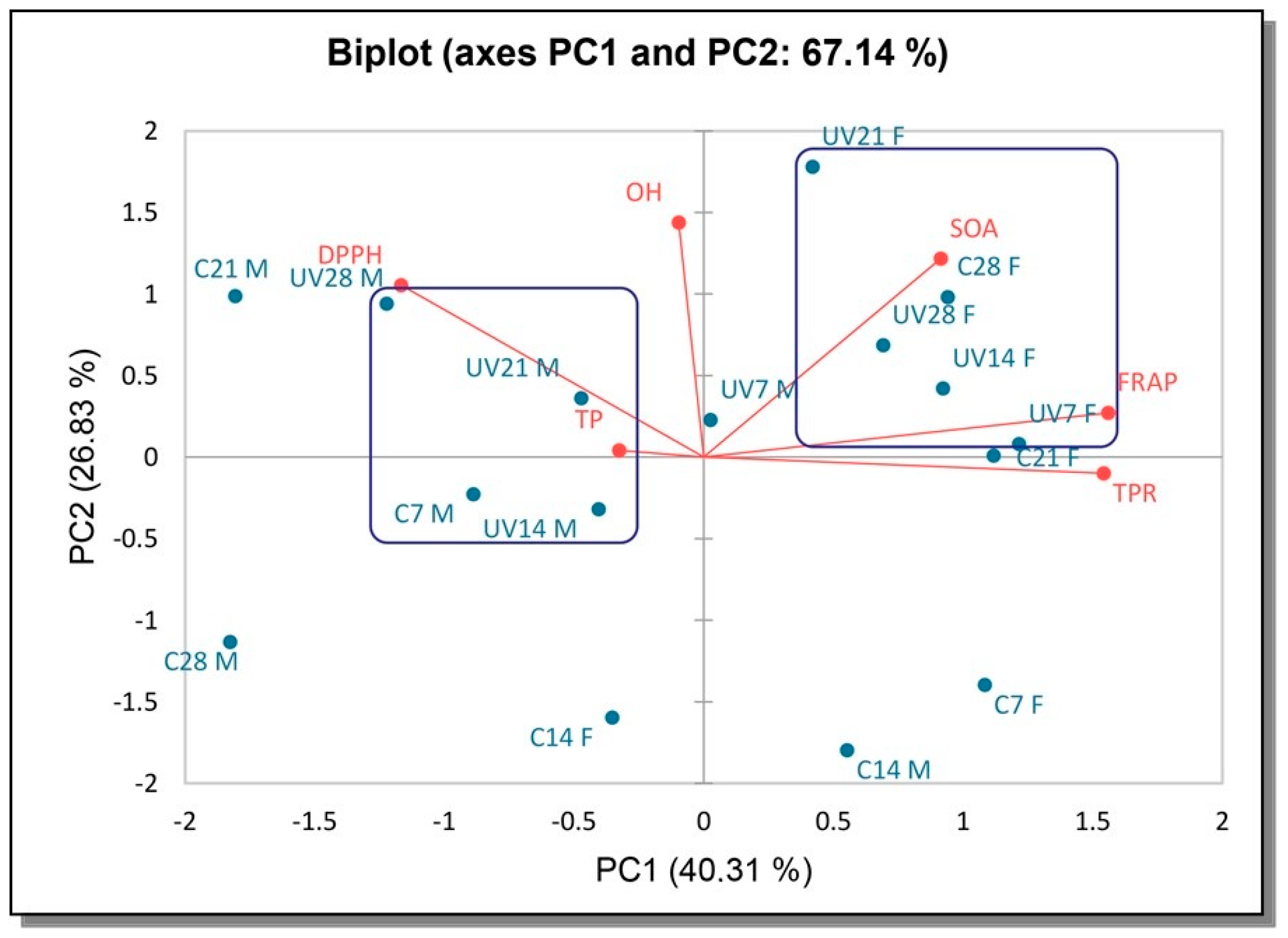
3.5. Principal Component Analysis (PCA)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singaravelan, N.; Grishkan, I.; Beharav, A.; Wakamatsu, K.; Ito, S.; Nevo, E. Adaptive melanin response of the soil fungus Aspergillus niger to UV radiation stress at “Evolution Canyon”, Mount Carmel, Israel. PLoS ONE 2008, 3, e2993. [Google Scholar] [CrossRef] [PubMed]
- Hessling, M.; Haag, R.; Sieber, N.; Vatter, P. The impact of far-UVC radiation (200–230 nm) on pathogens, cells, skin, and eyes—A collection and analysis of a hundred years of data. GMS Hyg. Infect. Control. 2021, 16, Doc07. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S.; Knowland, J. Ultraviolet radiation screening compounds. Biol. Rev. 1999, 74, 311–345. [Google Scholar] [CrossRef] [PubMed]
- Majerus, M.E.N. Melanism: Evolution in Action; Oxford University Press: Oxford, UK, 1998; p. 338. [Google Scholar]
- Bell, A.A.; Wheeler, M.H. Biosynthesis and functions of fungal melanins. Annu. Rev. Phytopathol. 1986, 24, 411–451. [Google Scholar] [CrossRef]
- Butler, M.J.; Day, A.W. Fungal melanins: A review. Can. J. Microbiol. 1998, 44, 1115–1136. [Google Scholar] [CrossRef]
- Fuller, K.K.; Loros, J.J.; Dunlap, J.C. Fungal photobiology: Visible light as a signal for stress, space and time. Curr. Genet. 2015, 61, 275–288. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated damage to cellular DNA by UVB and UVA radiation. Photochem. Photobiol. 2015, 91, 140–155. [Google Scholar] [CrossRef]
- Dulay, R. Antioxidant activity and total phenolic content of Volvariella volvacea and Schizophyllum commune mycelia cultured in indigenous liquid media. Mycosphere 2016, 7, 131–138. [Google Scholar] [CrossRef]
- Karaman, M.; Čapelja, E.; Rašeta, M.; Rakić, M. Diversity, Chemistry, and Environmental Contamination of Wild Growing Medicinal Mushroom Species as Sources of Biologically Active Substances (Antioxidants, Anti-Diabetics, and AChE Inhibitors). In Biology, Cultivation and Applications of Mushrooms; Arya, A., Rusevska, K., Eds.; Springer: Singapore, 2022; Volume 8, pp. 203–257. [Google Scholar]
- Rašeta, M.; Karaman, M.; Jakšić, M.; Šibul, F.; Kebert, M.; Novaković, A.; Popović, M. Mineral composition, antioxidant and cytotoxic biopotentials of wild-growing Ganoderma species (Serbia): G. lucidum (Curtis) P. Karst vs. G. applanatum (Pers.) Pat. Int. J. Food Sci. Technol. 2016, 51, 2583–2590. [Google Scholar] [CrossRef]
- Papagianni, М. Fungal morphology and metabolite production in submerged mycelial processes (review). Biotechnol. Adv. 2004, 22, 189–259. [Google Scholar] [CrossRef]
- Janjušević, L. Biological Activity and Chemical Characteristics of Selected Extracts of Autochthonous Macrofungi. Ph.D. Thesis, Faculty of Sciences, University of Novi Sad, Novi Sad, Serbia, 2017. [Google Scholar]
- Karaman, M.; Jovin, E.; Malbaša, R.; Matavuly, M.; Popović, M. Medicinal and edible lignicolous fungi as natural sources of antioxidative and antibacterial agents. Phytother. Res. 2010, 24, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Ikekawa, T. Beneficial effects of edible and medicinal mushrooms on health care. Int. J. Med. Mushrooms 2001, 3, 291–298. [Google Scholar]
- Rašeta, M.; Popović, M.; Knežević, P.; Šibul, F.; Kaišarević, S.; Karaman, M. Bioactive phenolic compounds of two medicinal mushroom species Trametes versicolor and Stereum subtomentosum as antioxidant and antiproliferative agents. Chem. Biodivers. 2020, 17, e2000683. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Ra, K.S.; Noh, D.O.; Suh, H.J. Optimization of submerged culture conditions for the production of angiotensin converting enzyme inhibitor from Flammulina velutipes. J. Ind. Microbiol. Biotechnol. 2002, 29, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.; Ismail, S.M.; Aminudin, N.; Shuib, A.S.; Lau, B.F. Evaluation of selected culinary-medicinal mushrooms for antioxidant and ace inhibitory activities. Evid. Based Complement. Altern. Med. 2012, 2012, 464238. [Google Scholar] [CrossRef]
- Orhan, I.; Üstün, O. Determination of total phenol content, antioxidant activity and acetylcholinesterase inhibition in selected mushrooms from Turkey. J. Food Compos. Anal. 2011, 24, 386–390. [Google Scholar] [CrossRef]
- Janjušević, L.; Pejin, B.; Kaišarević, S.; Gorjanović, S.; Pastor, F.; Tešanović, K.; Karaman, M. Trametes versicolor ethanol extract, a promising candidate for health–promoting food supplement. Nat. Prod. Res. 2018, 32, 963967. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Nikšić, M.; Vrvić, M.M.; Todorović, N.; Jakovljević, D.; Van Griensven, L.J. Antioxidative activities and chemical characterization of polysaccharides extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinus edodes and Trametes versicolor. J. Food Compos. Anal. 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Yeh, M.Y.; Ko, W.C.; Lin, L.Y. Hypolipidemic and antioxidant activity of Enoki mushrooms (Flammulina velutipes). BioMed Res. Int. 2014, 2014, 352–385. [Google Scholar] [CrossRef]
- Barros, A.B.; Bell, V.; Ferrão, J.; Calabrese, V.; Fernandes, T.H. Mushroom biomass: Some clinical implications of β-glucans and enzymes. Curr. Res. Nutr. Food Sci. 2016, 4, 37–47. [Google Scholar] [CrossRef]
- Orčić, D.; Francišković, M.; Bekvalac, K.; Svirčev, E.; Beara, I.; Lesjak, M.; Mimica-Dukić, N. Quantitative determination of plant phenolics in Urtica dioica extracts by high-performance liquid chromatography coupled with tandem mass-spectrometric detection. Food Chem. 2014, 143, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Espin, C.J.; Soler-Rivas, G.; Wichers, J.H. Characterization of the total free radical scavenger capacity of vegetable oils and oil fractions using 2,2-diphenyl-1-picrylhydrazyl radical. J. Agric. Food Chem. 2002, 48, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.I. The deoxyribose method: A simple test tube assay for determination of rate constants for reaction of hydroxyl radical. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–853. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid and concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar]
- Roleira, F.M.; Tavares-da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant derived and dietary phenolic antioxidants: Anticancer properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef]
- Mišković, J.; Karaman, M.; Rašeta, M.; Krsmanović, N.; Berežni, S.; Jakovljević, D.; Piattoni, F.; Zambonelli, A.; Gargano, M.L.; Venturella, G. Comparison of two Schizophyllum commune strains in production of acetylcholinesterase inhibitors and antioxidants from submerged cultivation. J. Fungi 2021, 7, 115. [Google Scholar] [CrossRef]
- Terpinc, P.; Čeh, B.; Poklar Ulrih, N.; Abramovič, H. Studies of the correlation between antioxidant properties and the total phenolic content of different oil cake extracts. Ind. Crops Prod. 2012, 39, 210–217. [Google Scholar] [CrossRef]
- Tešanović, K.; Pejin, B.; Šibul, F.; Matavulj, M.; Rašeta, M.; Janjušević, L.; Karaman, M. A comparative overview of antioxidative properties and phenolic profiles of different fungal origins: Fruiting bodies and submerged cultures of Coprinus comatus and Coprinellus truncorum. J. Food Sci. Technol. 2017, 54, 430–438. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Class of Analyzed Compounds | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Hydroxybenzoic Acid | Hydroxycinnamic Acid | Cyclohexanecarboxylic Acid | Isoflavonoid | Biflavonoid | Flavone | |||||
| Analyzed Sample | Incubation Time (Days) | p-Hydroxybenzoic Acid | Protocatechuic Acid | Vanillic Acid | p-Coumaric Acid | Quinic Acid | Daidzein | Genistein | Amentoflavone | Apiin |
| Flammulina velutipes | ||||||||||
| F | 7 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| 14 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 21 | 26,996.5 | 5405.0 | 18.0 | 8.0 | 63.5 | 642.0 | 440.5 | 0 | 0 | |
| 28 | 14,234.5 | 1759.5 | 70.0 | 6853.0 | 68.5 | 854.5 | 345.5 | 0 | 0 | |
| Fcont. | 7 | 15,233.5 | 2261.0 | 17.0 | 75.5 | 63.5 | 1316.0 | 943.5 | 306 | 0 |
| 14 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 21 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 28 | 19,580.5 | 4501.5 | 107.5 | 327.5 | 30 | 641 | 390 | 0 | 0 | |
| M | 7 | 11,102.0 | 970 | 0 | 107 | 158.5 | 1702.5 | 3210.5 | 1115.5 | 114 |
| 14 | 12,027.0 | 1475.5 | 21.5 | 106.5 | 81.5 | 1906.5 | 3152 | 0 | 266 | |
| 21 | 18,650,0 | 1555.0 | 56.5 | 52 | 72.5 | 1226 | 1966.5 | 0 | 85 | |
| 28 | 8900.0 | 3680.5 | 28 | 4756 | 43 | 1109.5 | 1304 | 0 | 89 | |
| M cont. | 7 | 15,364.5 | 1475.5 | 21.5 | 102.5 | 204.5 | 2539 | 4824.0 | 0 | 1589 |
| 14 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 21 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 28 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| Range | 8900–26,996.5 | 970–5405.0 | 17.0–107.5 | 8.0–6853.0 | 30–204.5 | 641–1906 | 345.5–4824.0 | 306–1115.5 | 85–1589 | |
| Trametes versicolor | ||||||||||
| F | 7 | 2081.0 | 19.5 | 0 | 36.5 | 341.5 | 189.5 | 148.5 | 0 | 0 |
| 14 | 365 | 29 | 0 | 0 | 147.5 | 9 | 12 | 0 | 0 | |
| 21 | 275.5 | 33.5 | 0 | 0 | 127 | 13.5 | 0 | 782 | 96.5 | |
| 28 | 417.5 | 13 | 0 | 0 | 85 | 0 | 0 | 1934.5 | 0 | |
| F cont. | 7 | 984.5 | 18.5 | 0 | 0 | 440.5 | 8 | 19.5 | 0 | 0 |
| 14 | 275.5 | 36.5 | 0 | 0 | 84.5 | 5 | 2.5 | 404 | 0 | |
| 21 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 28 | 45 | 0 | 0 | 0 | 53.5 | 0 | 0 | 0 | 0 | |
| M | 7 | 172.5 | 5.5 | 0 | 21 | 82 | 0 | 0 | 0 | 56 |
| 14 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 21 | 83 | 7 | 0 | 11 | 70 | 5 | 5 | 0 | 25 | |
| 28 | 1200 | 26 | 0 | 0 | 170.5 | 4 | 13.5 | 594.5 | 140.5 | |
| M cont. | 7 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| 14 | 166 | 43.5 | 0 | 0 | 103 | 0 | 12 | 0 | 134.5 | |
| 21 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 28 | 262 | 11.5 | 29 | 0 | 48 | 5 | 35 | 0 | 104 | |
| Range | 45–2081 | 5.5–43.5 | 0–29 | 11–36.5 | 48–440.5 | 4–189.5 | 5–148.5 | 404–1934.5 | 25–140.5 | |
| Extracts | TP (mg GAE/mg d.w.) | TPR (mg BSAE/g d.w.) | |||
|---|---|---|---|---|---|
| T. versicolor | UV exposed | Control | UV exposed | Control | |
| F | 7 days | 49.28 ± 0.83 a,c | 43.74 ± 0.45 a,c | 102.90 ± 2.20 a,c | 103.93 ± 3.59 a,c |
| 14 days | 48.43 ± 0.13 a,c | 47.22 ± 0.49 a,c | 79.40 ± 0.40 a,c | 64.80 ± 3.64 a,c | |
| 21 days | 50.03 ± 0.66 a,c | 19.22 ± 0.22 a,c | 77.50 ± 1.50 a,c | 115.29 ± 1.11 a,c | |
| 28 days | 18.78 ± 0.37 a,c | 8.61 ± 0.14 a,c | 103.16 ± 1.55 a,c | 117.73 ± 2.83 a,c | |
| M | 7 days | 47.36 ± 0.61 a,c | 12.67 ± 0.44 a,c | 122.98 ± 2.07 a,c | 74.33 ± 0.36 a,c |
| 14 days | 18.03 ± 0.39 a,c | 15.56 ± 0.30 a,c | 100.82 ± 0.50 a,c | 106.37 ± 3.27 a,c | |
| 21 days | 17.86 ± 0.40 a,c | 15.94 ± 0.15 a,c | 99.15 ± 2.60 a,c | 114.95 ± 3.27 a,c | |
| 28 days | 18.84 ± 0.35 a,c | 11.19 ± 0.33 a,c | 183.53 ± 2.84 a,c | 75.02 ± 0.86 a,c | |
| F. velutipes | UV exposed | Control | UV exposed | Control | |
| F | 7 days | 17.26 ± 0.26b,c | 17.81 ± 0.03 b,c | 112.90 ± 1.20 b,c | 103.93 ± 3.59 b,c |
| 14 days | 18.35 ± 0.33 b,c | 14.86 ± 0.55 b,c | 133.16 ± 1.45 b,c | 117.73 ± 2.83 b,c | |
| 21 days | 16.98 ± 0.15 b,c | 16.69 ± 0.39 b,c | 99.50 ± 1.40 b,c | 95.29 ± 1.11 b,c | |
| 28 days | 16.91 ± 0.11 b,c | 19.03 ± 0.39 b,c | 77.40 ± 0.0 b,c | 64.80 ± 3.64 b,c | |
| M | 7 days | 54.24 ± 0.35 b,d | 24.95 ± 1.28 b,d | 98.65 ± 1.58 b,c | 15.60 ± 0.88 b,c |
| 14 days | 59.60 ± 0.14 b,d | 58.34 ± 1.70 b,d | 75.58 ± 0.36 b,c | 19.67 ± 1.39 b,c | |
| 21 days | 25.19 ± 2.65 b,d | 34.06 ± 0.96 b,d | 42.07 ± 2.25 b,c | 25.34 ± 0.17 b,c | |
| 28 days | 21.94 ± 0.23 b,d | 13.71 ± 0.14 b,d | 8.56 ± 0.45 b,c | 14.01 ± 0.56 b,c | |
| Extracts | DPPH (%) | OH (%) | SOA (%) | FRAP (mg AAE/g d.w.) | |||||
|---|---|---|---|---|---|---|---|---|---|
| T. versicolor | |||||||||
| UV exposed | Control | UV exposed | Control | UV exposed | Control | UV exposed | Control | ||
| F | 7 days | 8.20 ± 0.11 a,c,e | 7.48 ± 0.22 a,c,e | 36.34 ± 2.45 a,c,e | 23.93 ± 0.60 a,d,e | 72.53 ± 0.38 a,c,e | 73.07 ± 2.23 a,c,e | 15.80 ± 0.21 a,c,e | 16.87 ± 0.20 a,c,e |
| 14 days | 8.21 ± 0.12 a,c,e | 7.95 ± 0.06 a,c,e | 24.65 ± 0.46 a,c,e | 26.79 ± 1.92 a,d,e | 73.55 ± 1.81 a,c,e | 71.90 ± 1.55 a,c,e | 15.43 ± 0.28 a,c,e | 15.55 ± 0.30 a,c,e | |
| 21 days | 7.94 ± 0.09 a,c,e | 7.99 ± 0.15 a,c,e | 15.68 ± 2.00 a,c,e | 24.69 ± 0.19 a,d,e | 73.34 ± 1.14 a,c,e | 34.35 ± 0.52 a,c,e | 15.44 ± 0.42 a,c,e | 18.63 ± 1.30 a,c,e | |
| 28 days | 7.88 ± 0.31 a,c,e | 3.96 ± 0.73 a,c,e | 36.36 ± 1.06 a,c,e | N.A. | 66.00 ± 0.48 a,c,e | 70.58 ± 0.10 a,c,e | 17.76 ± 0.01 a,c,e | 5.05 ± 0.27 a,c,e | |
| M | 7 days | 8.22 ± 0.41 a,c,e | 6.97 ± 0.14 a,c,e | 30.33 ± 1.15 a,c,e | 29.16 ± 1.91 a,c,e | 44.83 ± 1.24 a,c,e | 72.74 ± 0.10 a,c,e | 19.88 ± 0.34 a,c,e | 9.95 ± 0.21 a,c,e |
| 14 days | 8.36 ± 0.08 a,c,e | 8.70 ± 0.08 a,c,e | 30.88 ± 3.84 a,c,e | 4.98 ± 0.31 a,c,e | 69.43 ± 0.57 a,c,e | 58.45 ± 1.43 a,c,e | 24.11 ± 0.59 a,c,e | 22.85 ± 0.50 a,c,e | |
| 21 days | 8.86 ± 0.13 a,c,e | 8.05 ± 0.47 a,c,e | 32.22 ± 1.98 a,c,e | 39.19 ± 2.96 a,c,e | 70.92 ± 1.72 a,c,e | 55.82 ± 2.10 a,c,e | 22.90 ± 0.40 a,c,e | 21.25 ± 0.90 a,c,e | |
| 28 days | 8.56 ± 0.22 a,c,e | 4.29 ± 0.07 a,c,e | 51.78 ± 0.61 a,c,e | 22.82 ± 1.67 a,c,e | 57.57 ± 0.95 a,c,e | 68.15 ± 0.86 a,c,e | 26.79 ± 0.33 a,c,e | 24.89 ± 0.25 a,c,e | |
| F. velutipes | |||||||||
| UV exposed | Control | UV exposed | Control | UV exposed | Control | UV exposed | Control | ||
| F | 7 days | 3.13 ± 0.52 b,c,e | 1.17 ± 0.18 b,c,e | 23.15 ± 1.57 a,c,e | 16.61 ± 1.96 a,c,e | 68.99 ± 0.70 b,c,e | 29.44 ± 0.30 b,c,e | 74.62 ± 0.09 b,c,e | 78.25 ± 1.10 b,c,e |
| 14 days | 4.53 ± 0.29 b,c,e | 3.10 ± 0.33 b,c,e | 24.55 ± 1.70 a,c,e | 8.04 ± 1.48 a,c,e | 68.06 ± 0.01 b,c,e | 25.46 ± 0.33 b,c,e | 85.36 ± 0.50 b,c,e | 23.72 ± 0.61 b,c,e | |
| 21 days | 7.08 ± 0.58 b,c,e | 4.04 ± 0.05 b,c,e | 51.46 ± 0.02 a,c,e | 15.67 ± 2.11 a,c,e | 71.29 ± 0.54 b,c,e | 71.12 ± 0.31 b,c,e | 74.66 ± 0.23 b,c,e | 64.71 ± 1.74 b,c,e | |
| 28 days | 5.25 ± 0.05 b,c,e | 7.26 ± 0.27 b,c,e | 30.58 ± 4.84 a,c,e | 25.40 ± 1.64 a,c,e | 71.16 ± 0.93 b,c,e | 71.45 ± 0.31 b,c,e | 56.08 ± 0.53 b,c,e | 83.10 ± 2.72 b,c,e | |
| M | 7 days | 6.23 ± 0.19 b,c,e | 7.00 ± 0.12 b,c,e | 26.10 ± 2.96 a,c,e | N.D. | 48.04 ± 1.70 b,c,f | 59.74 ± 2.79 b,c,f | 55.87 ± 0.36 b,c,f | 32.12 ± 1.65 b,c,f |
| 14 days | 7.04 ± 0.12 b,c,e | 2.90 ± 0.02 b,c,e | 12.53 ± 1.52 a,c,e | N.D. | 34.69 ± 2.01 b,c,f | 23.96 ± 1.39 b,c,f | 61.39 ± 0.14 b,c,f | 60.09 ± 1.75 b,c,f | |
| 21 days | 6.60 ± 0.62 b,c,e | 8.14 ± 0.10 b,c,e | 42.68 ± 2.17 a,c,e | 39.96 ± 2.14 a,c,e | 37.75 ± 0.93 b,c,f | 40.21 ± 2.24 b,c,f | 25.94 ± 2.73 b,c,f | 35.08 ± 0.99 b,c,f | |
| 28 days | 8.00 ± 0.05 b,c,e | 7.68 ± 0.32 b,c,e | 49.49 ± 2.62 a,c,e | N.D. | 41.36 ± 0.31 b,c,f | 21.12 ± 0.71 b,c,f | N.A. | N.A. | |
| Correlation Coefficient—R2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Extracts | DPPH/TP | OH/TP | SOA/TP | FRAP/TP | |||||
| T. versicolor | |||||||||
| UV exposed | Control | UV exposed | Control | UV exposed | Control | UV exposed | Control | ||
| F | 7 days | 0.01 | 0.58 * | 0.01 | 0.03 | 0.01 | 0.53 * | 0.13 | 0.22 |
| 14 days | 0.73 * | 0.45 | 0.63 * | 0.23 | 0.61 * | 0.26 | 0.91 * | 0.08 | |
| 21 days | 0.97 * | 0.35 | 0.08 | 0.85 * | 0.33 | 0.86 * | 0.87 * | 0.03 | |
| 28 days | 0.01 | 0.48 | 0.44 | / | 0.44 | 1.00 | 0.05 | 0.21 | |
| M | 7 days | 0.99 * | 0.70 * | 0.83 * | 0.93 * | 0.73 * | 0.08 | 0.88 * | 0.09 |
| 14 days | 0.97 * | 0.42 | 0.01 | 0.64 * | 0.85 * | 0.85 * | 0.40 | 0.89 * | |
| 21 days | 0.74 * | 0.36 | 0.76 * | 0.42 | 0.02 | 0.97 | 0.16 | 1.00 * | |
| 28 days | 0.21 | 0.65* | 0.40 | 0.99 * | 0.40 | 0.16 | 0.01 | 0.82 * | |
| F. velutipes | |||||||||
| UV exposed | Control | UV exposed | Control | UV exposed | Control | UV exposed | Control | ||
| F | 7 days | 0.36 | 0.73 * | 0.36 | 0.07 | 0.36 | 0.01 | 0.01 | 0.56 * |
| 14 days | 0.04 | 0.54 * | 0.56 * | 0.91 * | 0.10 | 0.18 | 0.02 | 0.04 | |
| 21 days | 0.81 * | 0.30 | 0.98 * | 0.27 | 0.68 * | 1.00 * | 0.98 * | 0.45 | |
| 28 days | 0.01 | 0.22 | 0.01 | 0.08 | 0.01 | 0.48 | 0.96 * | 0.59 * | |
| M | 7 days | 0.55 * | 0.93 * | 0.67 * | / | 0.01 | 0.99 * | 1.00 * | 0.16 |
| 14 days | 0.97 * | 0.83 * | 0.01 | / | 0.65 * | 0.03 | 1.00 * | 0.03 | |
| 21 days | 0.12 | 0.47 | 0.40 | 0.43 | 0.12 | 0.88 * | 1.00 * | 1.00 * | |
| 28 days | 0.07 | 0.98 * | 0.84 * | / | 0.64 * | 0.64 * | / | / | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krsmanović, N.; Rašeta, M.; Mišković, J.; Bekvalac, K.; Bogavac, M.; Karaman, M.; Isikhuemhen, O.S. Effects of UV Stress in Promoting Antioxidant Activities in Fungal Species Тrametes versicolor (L.) Lloyd and Flammulina velutipes (Curtis) Singer. Antioxidants 2023, 12, 302. https://doi.org/10.3390/antiox12020302
Krsmanović N, Rašeta M, Mišković J, Bekvalac K, Bogavac M, Karaman M, Isikhuemhen OS. Effects of UV Stress in Promoting Antioxidant Activities in Fungal Species Тrametes versicolor (L.) Lloyd and Flammulina velutipes (Curtis) Singer. Antioxidants. 2023; 12(2):302. https://doi.org/10.3390/antiox12020302
Chicago/Turabian StyleKrsmanović, Nenad, Milena Rašeta, Jovana Mišković, Kristina Bekvalac, Mirjana Bogavac, Maja Karaman, and Omoanghe S. Isikhuemhen. 2023. "Effects of UV Stress in Promoting Antioxidant Activities in Fungal Species Тrametes versicolor (L.) Lloyd and Flammulina velutipes (Curtis) Singer" Antioxidants 12, no. 2: 302. https://doi.org/10.3390/antiox12020302
APA StyleKrsmanović, N., Rašeta, M., Mišković, J., Bekvalac, K., Bogavac, M., Karaman, M., & Isikhuemhen, O. S. (2023). Effects of UV Stress in Promoting Antioxidant Activities in Fungal Species Тrametes versicolor (L.) Lloyd and Flammulina velutipes (Curtis) Singer. Antioxidants, 12(2), 302. https://doi.org/10.3390/antiox12020302