
Cannabidiol (CBD) Supports the Honeybee Worker Organism by Activating the Antioxidant System



Abstract
1. Introduction
2. Materials and Methods
2.1. Preparatory Activities
2.1.1. Obtaining Queens for the Experiment
2.1.2. Preparation of the Mating Hives
2.2. Collection and Tagging of Day-Old Bees
2.3. Preparation and Administration of CBD Extract
- –
- CSy: ad libitum, on the second, fourth, and sixth days of the experiment, in a mixture with sugar syrup (1:1 water with sugar) and glycerin in the volume ratio of 0.01:0.5:0.5 (extract:distilled water:glycerin);
- –
- CSt: mixture with water and glycerin in a volume ratio of 0.8:1.5:1.5 (extract:distilled water:glycerin), textile strips (2 × 10 cm) were evenly moistened with 10 mL of the mixture and placed in the hives, wetted with the mixture on the second, fourth, and sixth days of the experiment.
2.4. Sampling of Bees
2.5. Collection of Hemolymph
2.6. Biochemical Analyses
2.6.1. Antioxidant System
- –
- TAC—OxiSelectTM Total Antioxidant Capacity Assay Kit (Cell BioLabs, Inc., Upper Heyford, UK, no. STA-360);
- –
- SOD Assay Kit (Sigma Aldrich, Schnelldorf, Germany, no. 1916-1KT-F);
- –
- CAT Assay Kit (Sigma Aldrich, Schnelldorf, Germany no. CAT100-1KT);
- –
- GPx—Glutathione peroxidase Assay Kit (Sigma Aldrich, Schnelldorf, Germany, no. MAK437);
- –
- GSH—EnzyChromTM GSH/GSSG Assay Kit (Bio Assay Systems, Hayward, CA, USA, no. EGTT-100).
2.6.2. Ions Concentration
- –
- Calcium (Alpha Diagnostics, arsenazo III method, reagent composition: TRIS buffer pH 8.5; arsenazo III, 8-hydroxy-quinoline-5-sulfonic acid, inactive stabilizers and detergents 630–670 nm);
- –
- Magnesium (Alpha Diagnostics Magnesium Xylidyl Blue, reagent composition: trioglycolic acid, DMSO, Xylidyl Blue, measurement at 550 nm);
- –
- Phosphorus (Alpha Diagnostics, direct method with phosphomolybdate, reagent composition: sulfuric acid, ammonium molybdate, measurement at 340 nm).
2.7. Statistical Analysis
3. Results
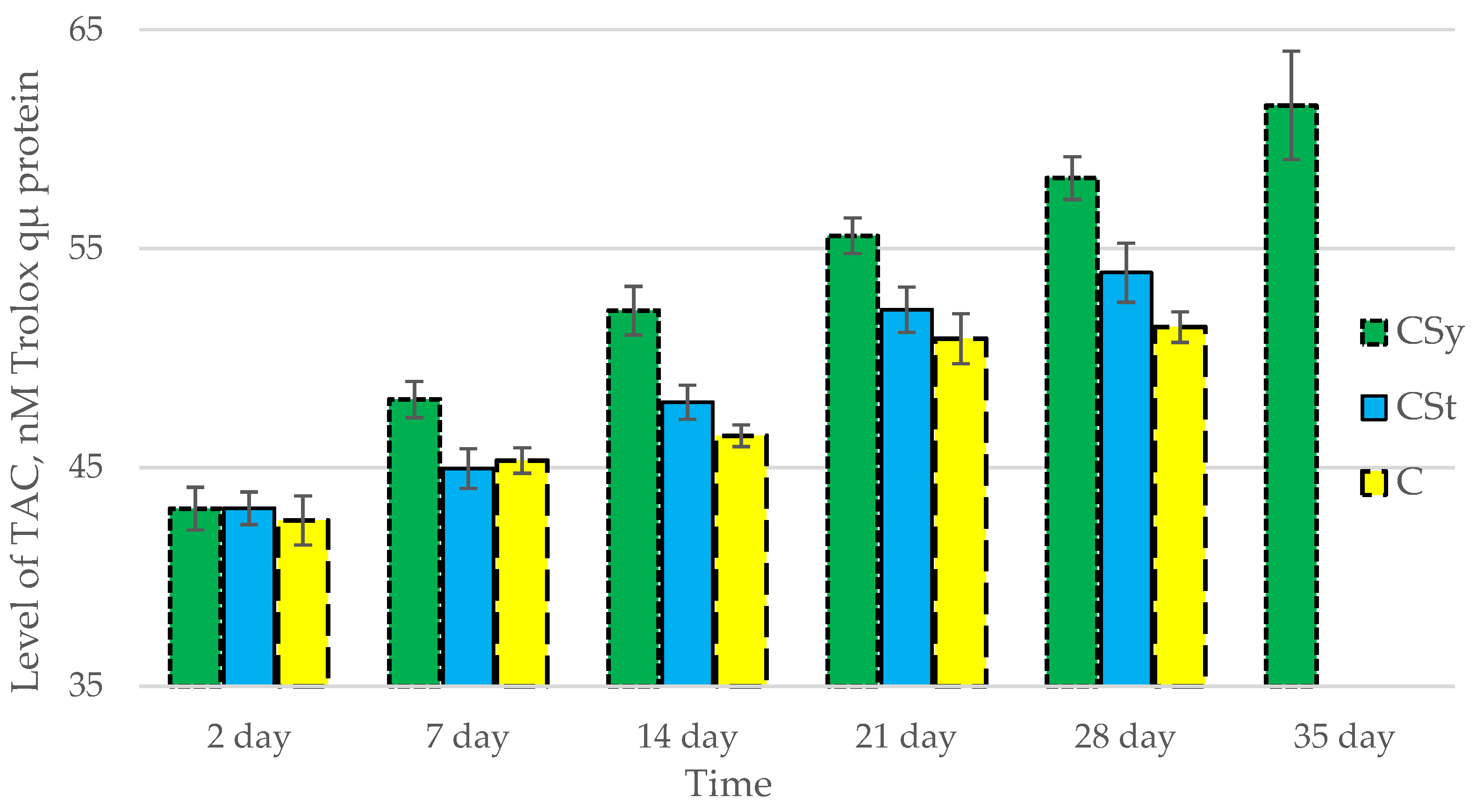
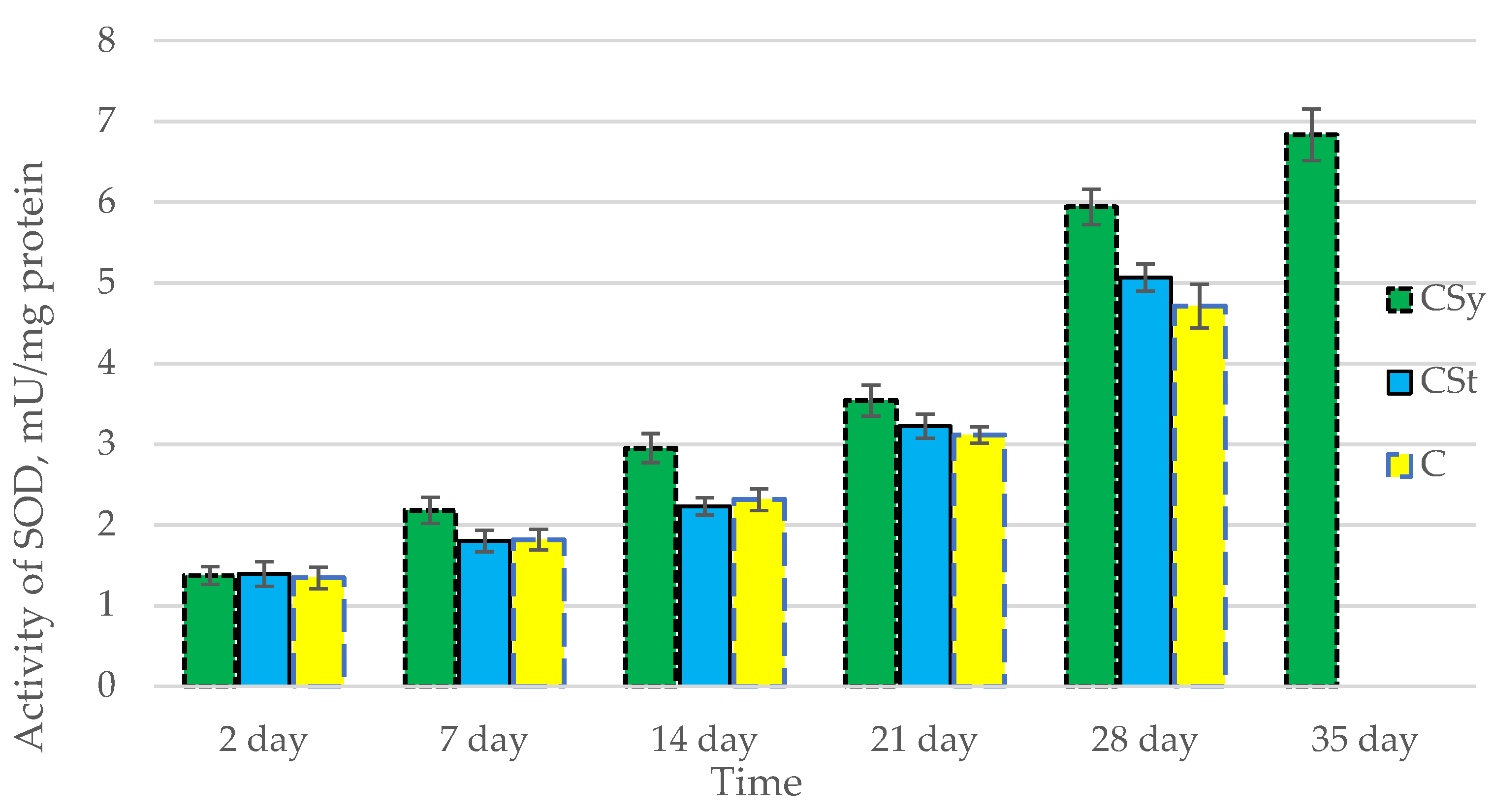
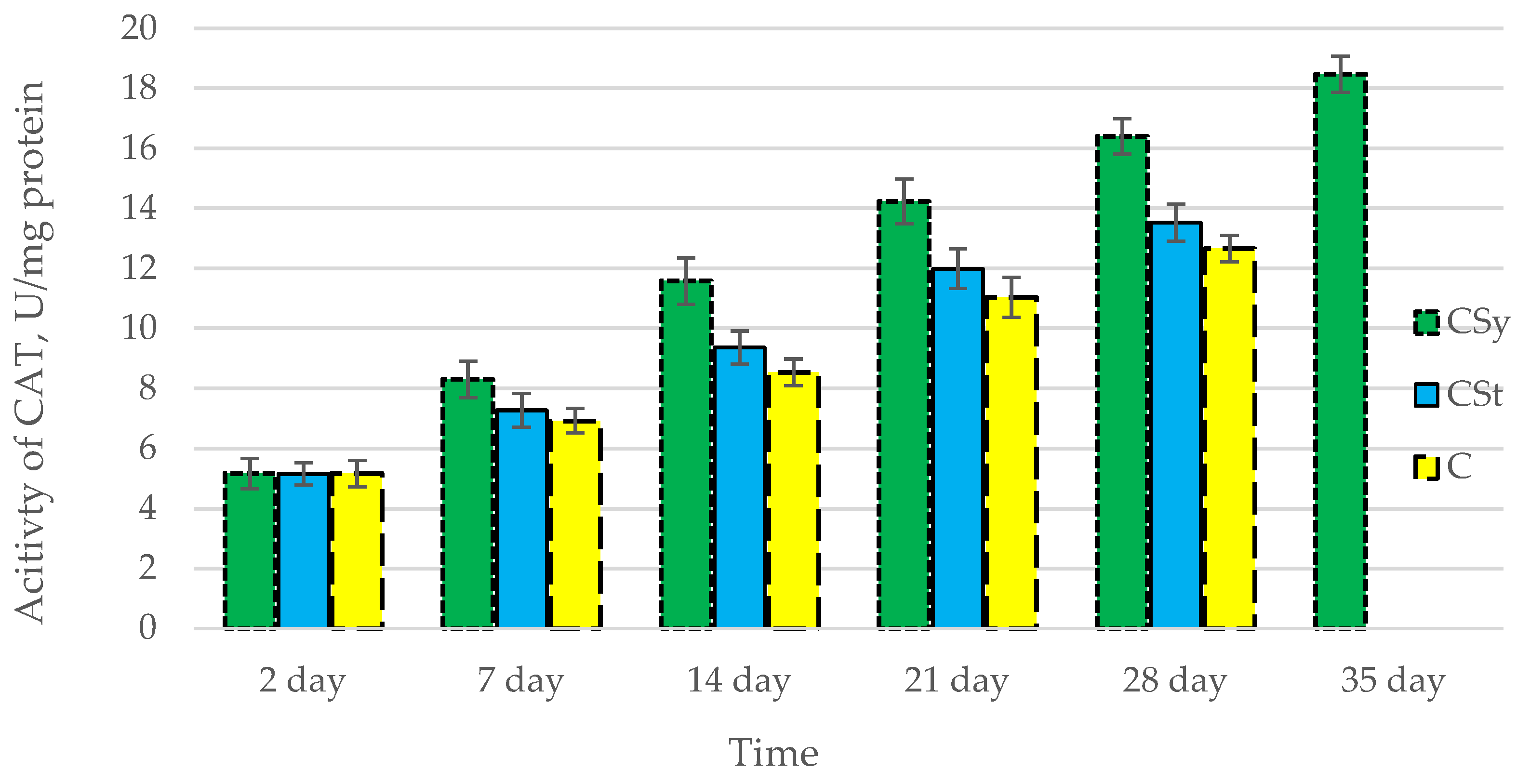
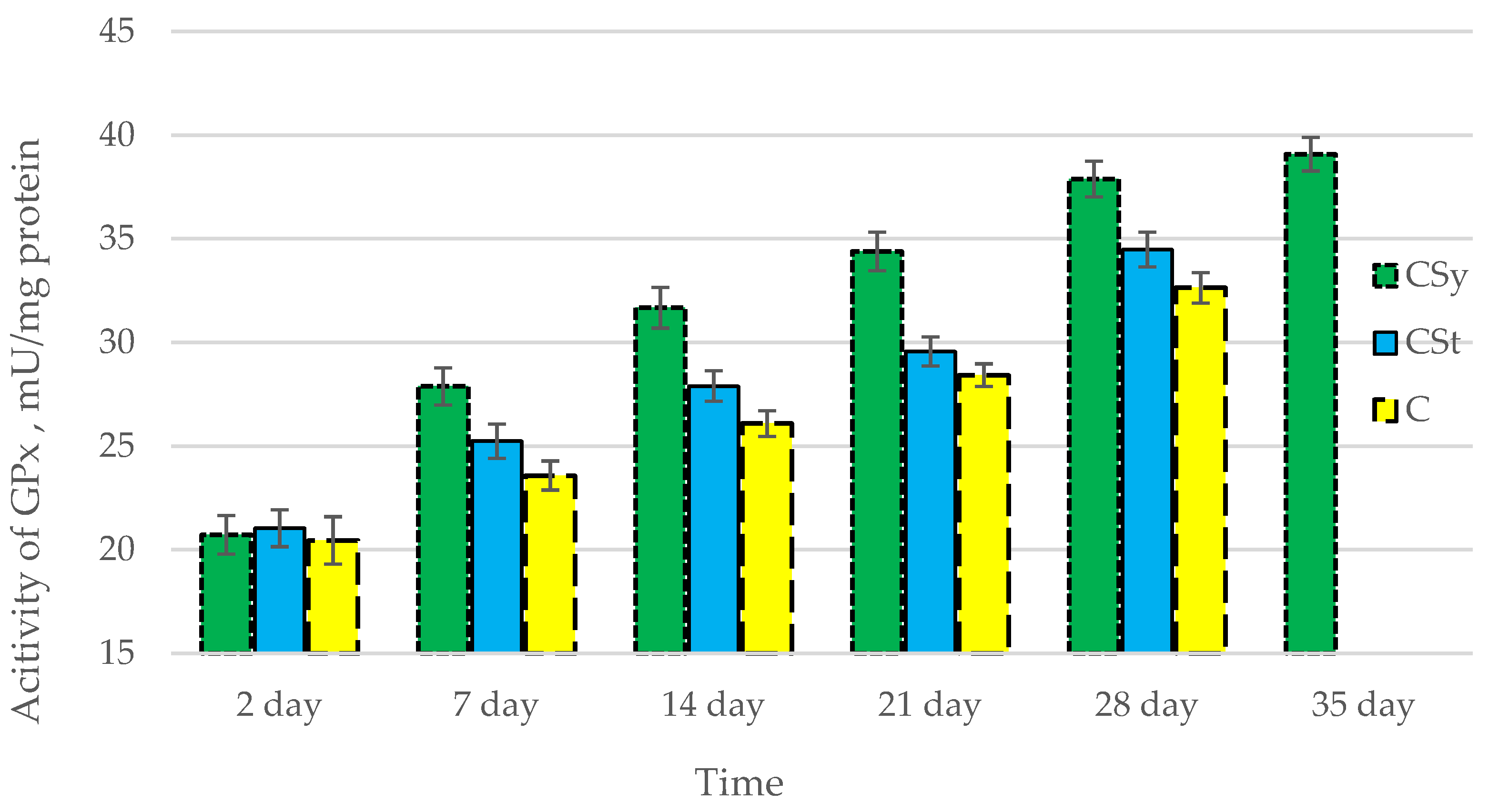
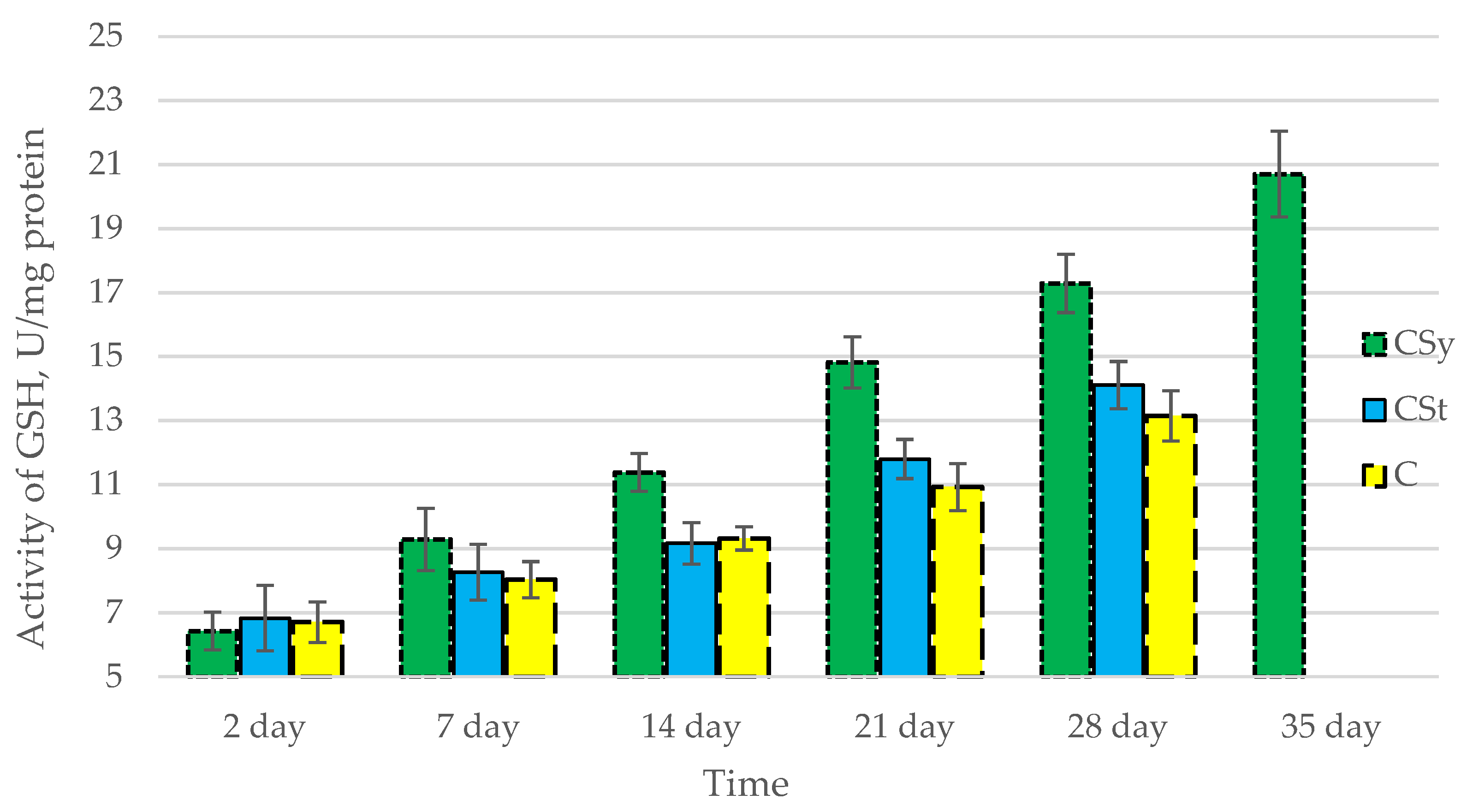
3.1. Antioxidant Enzymes
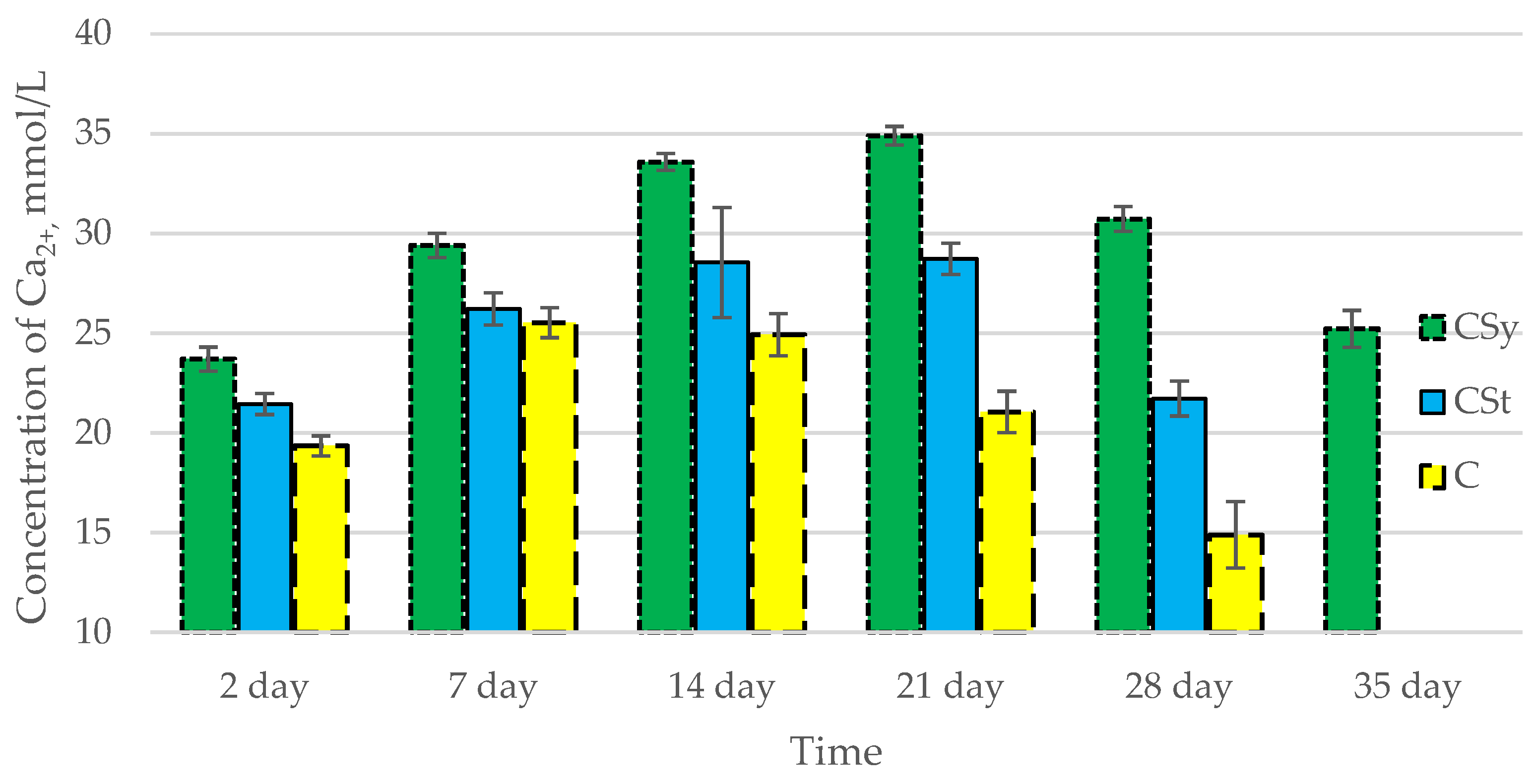
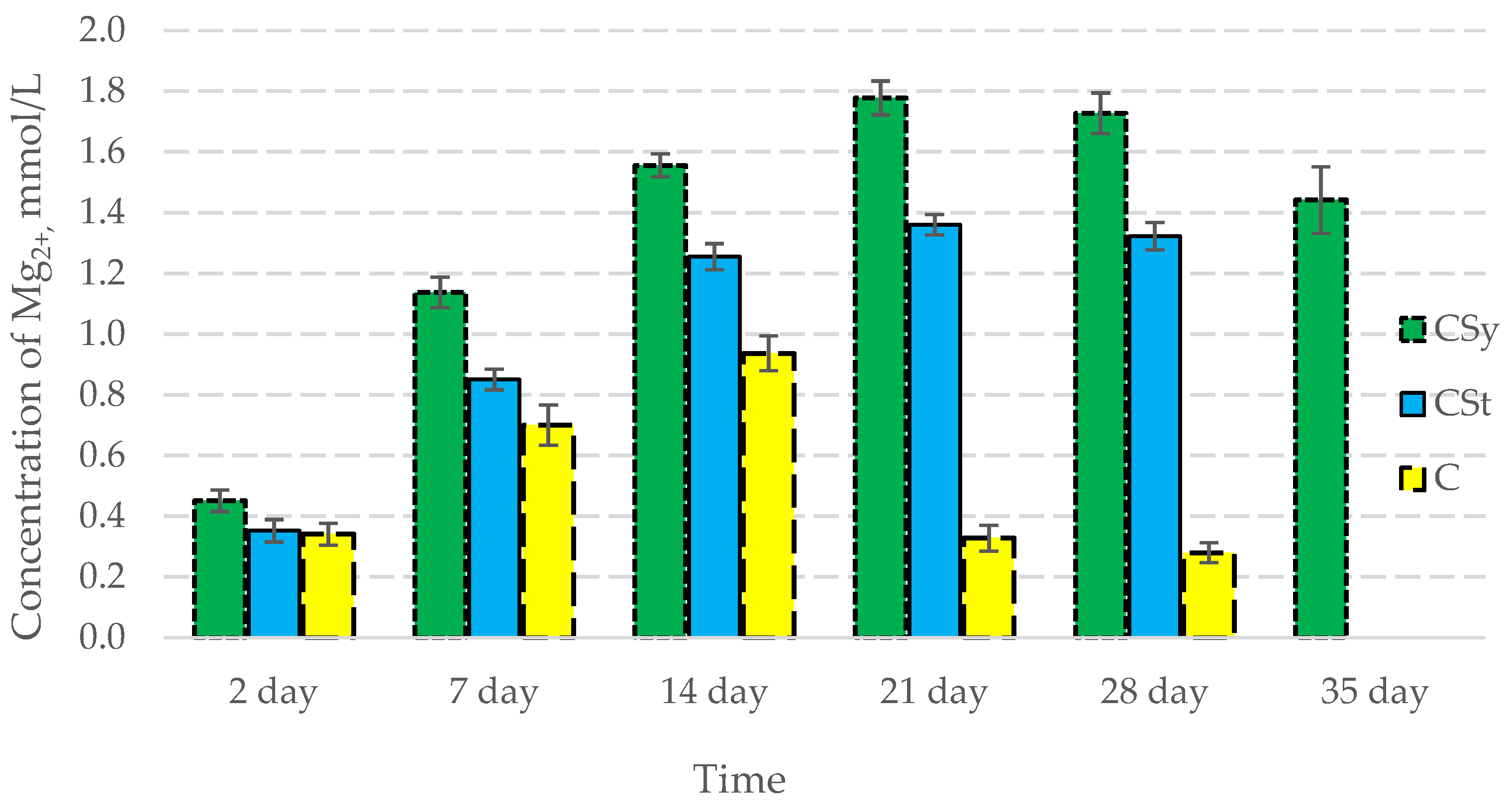
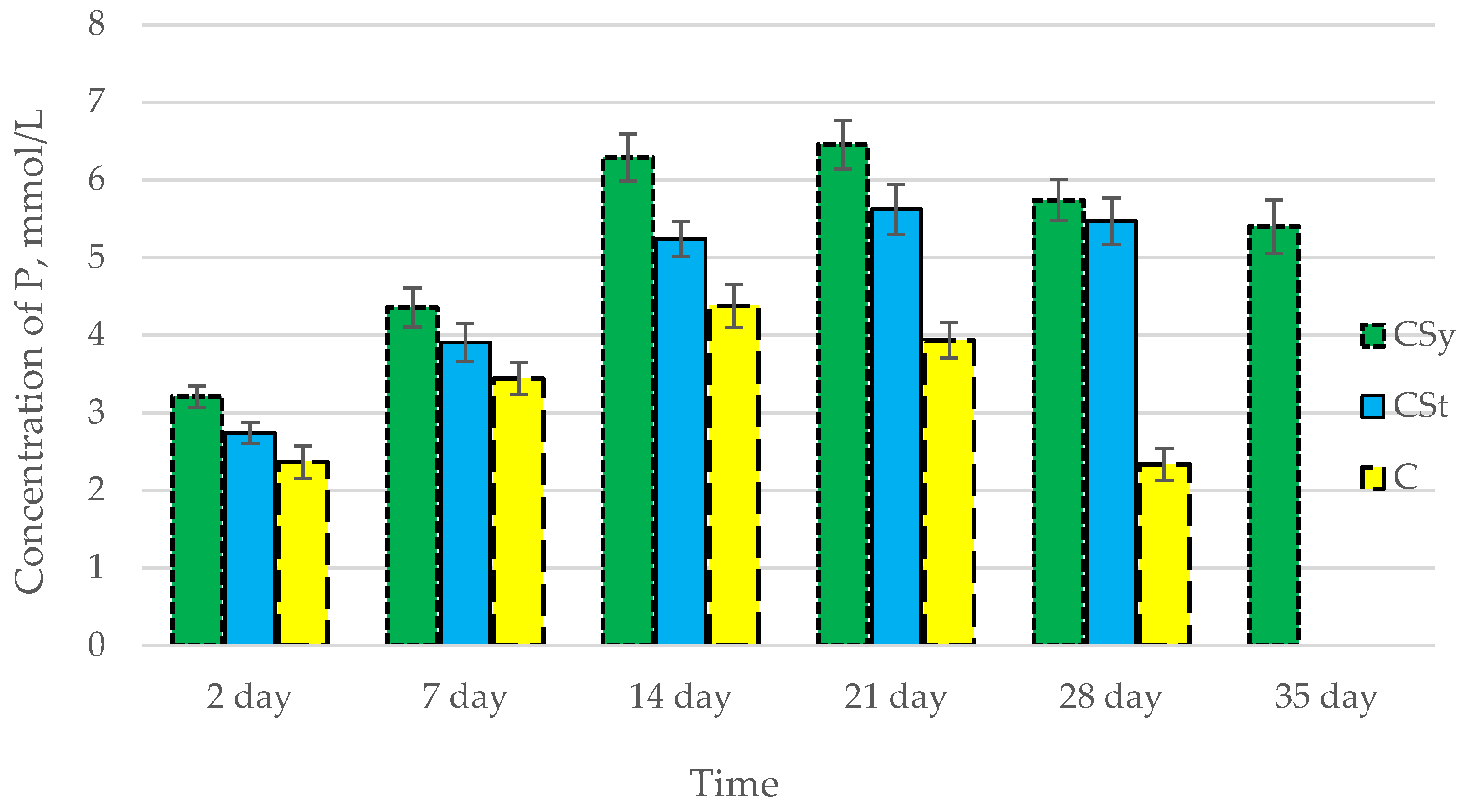
3.2. Ion Concentrations

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Connor, B.; Bojinski, S.; Röösli, C.; Schaepman, M.E. Monitoring global changes in biodiversity and climate essential as ecological crisis intensifies. Ecol. Inform. 2020, 55, 101033. [Google Scholar] [CrossRef]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. OIE Rev. Sci. Tech. 2008, 27, 485–510. [Google Scholar] [CrossRef]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors Associated with Honey Bee Colony Losses: A Mini-Review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; Martín-Hernández, R.; Garrido-Bailón, E.; González-Porto, A.V.; García-Palencia, P.; Meana, A.; Del Nozal, M.J.; Mayo, R.; Bernal, J.L. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 2009, 1, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Lupi, D.; Mesiano, M.P.; Adani, A.; Benocci, R.; Giacchini, R.; Parenti, P.; Zambon, G.; Lavazza, A.; Boniotti, M.B.; Bassi, S.; et al. Combined Effects of Pesticides and Electromagnetic-Fields on Honeybees: Multi-Stress Exposure. Insects 2021, 12, 716. [Google Scholar] [CrossRef] [PubMed]
- Aizen, M.A.; Harder, L.D. The Global Stock of Domesticated Honey Bees Is Growing Slower Than Agricultural Demand for Pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; He, Q.; Liu, Q.; Wang, Z.; Yin, F.; Chai, Y.; Yang, Q.; Jiang, X.; Liao, M.; Yu, L.; et al. Analysis of honey bee exposure to multiple pesticide residues in the hive environment. Sci. Total Environ. 2022, 805, 150292. [Google Scholar] [CrossRef]
- Jankielsohn, A.; Jankielsohn, A. The Importance of Insects in Agricultural Ecosystems. Adv. Entomol. 2018, 6, 62–73. [Google Scholar] [CrossRef]
- van der Steen, J. Effect of a home-made pollen substitute on honey bee colony development. J. Apic. Res. 2007, 46, 114–119. [Google Scholar] [CrossRef]
- Skowronek, P.; Wójcik, Ł.; Strachecka, A. Cannabis Extract Has a Positive–Immunostimulating Effect through Proteolytic System and Metabolic Compounds of Honey Bee (Apis mellifera) Workers. Animals 2021, 11, 2190. [Google Scholar] [CrossRef]
- Rascón, B.; Hubbard, B.P.; Sinclair, D.A.; Amdam, G.V. The lifespan extension effects of resveratrol are conserved in the honey bee and may be driven by a mechanism related to caloric restriction. Aging 2012, 4, 499–508. [Google Scholar] [CrossRef]
- Schulz, M.; Łoś, A.; Grzybek, M.; Ścibior, R.; Strachecka, A. Piperine as a new natural supplement with beneficial effects on the life-span and defence system of honeybees. J. Agric. Sci. 2019, 157, 140–149. [Google Scholar] [CrossRef]
- Suwannapong, G.; Maksong, S.; Phainchajoen, M.; Benbow, M.E.; Mayack, C. Survival and health improvement of Nosema infected Apis florea (Hymenoptera: Apidae) bees after treatment with propolis extract. J. Asia. Pac. Entomol. 2018, 21, 437–444. [Google Scholar] [CrossRef]
- Malassigné, S.; Minard, G.; Vallon, L.; Martin, E.; Moro, C.V.; Luis, P. Diversity and Functions of Yeast Communities Associated with Insects. Microorganisms 2021, 9, 1552. [Google Scholar] [CrossRef] [PubMed]
- Strachecka, A.; Olszewski, K.; Paleolog, J. Curcumin stimulates biochemical mechanisms of Apis mellifera resistance and extends the apian life-span. J. Apic. Sci. 2015, 59, 129–141. [Google Scholar] [CrossRef]
- Strachecka, A.; Krauze, M.; Olszewski, K.; Borsuk, G.; Paleolog, J.; Merska, M.; Chobotow, J.; Bajda, M.; Grzywnowicz, K. Unexpectedly strong effect of caffeine on the vitality of western honeybees (Apis mellifera). Biochem. 2014, 79, 1192–1201. [Google Scholar] [CrossRef]
- Farjan, M.; Dmitryjuk, M.; LipiŃski, Z.; Biernat-łOpieŃska, E.; Zółtowska, K. Supplementation of the honey bee diet with vitamin C: The effect on the antioxidative system of Apis mellifera carnica brood at different stages. J. Apic. Res. 2012, 51, 263–270. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, K.A.; Khan, S.A.; Ghramh, H.A.; Gul, A. Comparative assessment of various supplementary diets on commercial honey bee (Apis mellifera) health and colony performance. PLoS ONE 2021, 16, e0258430. [Google Scholar] [CrossRef] [PubMed]
- Stanimirović, Z.; Glavinić, U.; Ristanić, M.; Jelisić, S.; Vejnović, B.; Niketić, M.; Stevanović, J. Diet Supplementation Helps Honey Bee Colonies in Combat Infections by Enhancing their Hygienic Behaviour. Acta Vet. Brno. 2022, 72, 145–166. [Google Scholar] [CrossRef]
- Strachecka, A.; Olszewski, K.; Paleolog, J.; Borsuk, G.; Bajda, M.; Krauze, M.; Merska, M.; Chobotow, J. Coenzyme Q10 treatments influence the lifespan and key biochemical resistance systems in the honeybee, apis mellifera. Arch. Insect Biochem. Physiol. 2014, 86, 165–179. [Google Scholar] [CrossRef]
- Geldert, C.; Abdo, Z.; Stewart, J.E.; Arathi, H.S. Dietary supplementation with phytochemicals improves diversity and abundance of honey bee gut microbiota. J. Appl. Microbiol. 2021, 130, 1705–1720. [Google Scholar] [CrossRef] [PubMed]
- Ricigliano, V.A.; Simone-Finstrom, M. Nutritional and prebiotic efficacy of the microalga Arthrospira platensis (spirulina) in honey bees. Apidologie 2020, 51, 898–910. [Google Scholar] [CrossRef]
- Skowronek, P.; Wójcik, Ł.; Strachecka, A. CBD Supplementation Has a Positive Effect on the Activity of the Proteolytic System and Biochemical Markers of Honey Bees (Apis mellifera) in the Apiary. Animals 2022, 12, 2313. [Google Scholar] [CrossRef]
- Manning, R.; Rutkay, A.; Eaton, L.; Dell, B. Lipid-enhanced pollen and lipid-reduced flour diets and their effect on the longevity of honey bees (Apis mellifera L.). Aust. J. Entomol. 2007, 46, 251–257. [Google Scholar] [CrossRef]
- Safian Boldaji, M.; Jalali Dehkordi, K.; Taghian, F.; Author, C. Effect of eight weeks of swimming training and CBD oil consumption on PI3K and ERK gene expression in the heart tissue of rats with myocardial infarction. J. Shahrekord Univ. Med. Sci. 2022, 24, 54–59. [Google Scholar] [CrossRef]
- Majewski, M.; Jurgoński, A. The Effect of Hemp (Cannabis sativa L.) Seeds and Hemp Seed Oil on Vascular Dysfunction in Obese Male Zucker Rats. Nutrients 2021, 13, 2575. [Google Scholar] [CrossRef]
- Isenmann, E.; Veit, S.; Starke, L.; Flenker, U.; Diel, P. Effects of Cannabidiol Supplementation on Skeletal Muscle Regeneration after Intensive Resistance Training. Nutrients 2021, 13, 3028. [Google Scholar] [CrossRef] [PubMed]
- Draeger, A.L.; Thomas, E.P.; Jones, K.A.; Davis, A.J.; Porr, C.A.S. The effects of pelleted cannabidiol supplementation on heart rate and reaction scores in horses. J. Vet. Behav. 2021, 46, 97–100. [Google Scholar] [CrossRef]
- Xin, G.; Yang, J.; Li, R.; Gao, Q.; Li, R.; Wang, J.; Zhang, J.; Wang, J. Dietary supplementation of hemp oil in teddy dogs: Effect on apparent nutrient digestibility, blood biochemistry and metabolomics. Bioengineered 2022, 13, 6173–6187. [Google Scholar] [CrossRef]
- Skowronek, P.; Wójcik, Ł.; Strachecka, A. Impressive Impact of Hemp Extract on Antioxidant System in Honey Bee (Apis mellifera) Organism. Antioxidants 2022, 11, 707. [Google Scholar] [CrossRef]
- Birch-Machin, M.A.; Bowman, A. Oxidative stress and ageing. Br. J. Dermatol. 2016, 175, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Siuda, M.; Wilde, J.; Woyke, J.; Jasiński, Z.; Madras-Majewska, B. Wintering reserve queens in mini-plus and 3-comb nuclei. J. Apic. Sci. 2014, 58, 61–68. [Google Scholar] [CrossRef]
- Scheiner, R.; Abramson, C.I.; Brodschneider, R.; Crailsheim, K.; Farina, W.M.; Fuchs, S.; Grünewald, B.; Hahshold, S.; Karrer, M.; Koeniger, G.; et al. Standard methods for behavioural studies of Apis mellifera. J. Apic. Res. 2013, 52, 1–58. [Google Scholar] [CrossRef]
- Łoś, A.; Strachecka, A. Fast and cost-effective biochemical spectrophotometric analysis of solution of insect “blood” and body surface elution. Sensors 2018, 18, 1494. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, B.; Zheng, X.; Ji, H.; Feng, K.; Hu, X.; Gul, I.; Abbas, M.N.; Cui, H.; Zhu, Y. Molecular Characterization of Two Genes Encoding Novel Ca2+-Independent Phospholipase A2s from the Silkworm, Bombyx mori. Curr. Issues Mol. Biol. 2022, 44, 777–790. [Google Scholar] [CrossRef]
- Kim, Y.; Stanley, D. Eicosanoid Signaling in Insect Immunology: New Genes and Unresolved Issues. Genes 2021, 12, 211. [Google Scholar] [CrossRef]
- Büyükgüzel, E.; Erdem, M.; Tunaz, H.; Küçük, C.; Atılgan, U.C.; Stanley, D.; Büyükgüzel, K. Inhibition of eicosanoid signaling leads to increased lipid peroxidation in a host/parasitoid system. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 204, 121–128. [Google Scholar] [CrossRef]
- Park, Y.; Stanley, D.W.; Kim, Y. Eicosanoids up-regulate production of reactive oxygen species by NADPH-dependent oxidase in Spodoptera exigua phagocytic hemocytes. J. Insect Physiol. 2015, 79, 63–72. [Google Scholar] [CrossRef]
- Park, Y.; Kumar, S.; Kanumuri, R.; Stanley, D.; Kim, Y. A novel calcium-independent cellular PLA2 acts in insect immunity and larval growth. Insect Biochem. Mol. Biol. 2015, 66, 13–23. [Google Scholar] [CrossRef]
- Büyükgüzel, E.; Hyršl, P.; Büyükgüzel, K. Eicosanoids mediate hemolymph oxidative and antioxidative response in larvae of Galleria mellonella L. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Poovaiah, B.W.; Ryan, C.A. Hydrogen peroxide homeostasis: Activation of plant catalase by calciumcalmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [PubMed]
- Watkins, A.R. Cannabinoid interactions with ion channels and receptors. Channels 2019, 13, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Corona, M.; Hughes, K.A.; Weaver, D.B.; Robinson, G.E. Gene expression patterns associated with queen honey bee longevity. Mech. Ageing Dev. 2005, 126, 1230–1238. [Google Scholar] [CrossRef]
- Mittapalli, O.; Neal, J.J.; Shukle, R.H. Antioxidant defense response in a galling insect. Proc. Natl. Acad. Sci. USA 2007, 104, 1889–1894. [Google Scholar] [CrossRef]
- Tunaz, H.; Park, Y.; Büyükgüzel, K.; Bedick, J.G.; Aliza, A.R.N.; Stanley, D.W. Eicosanoids in insect immunity: Bacterial infection stimulates hemocytic phospholipase A2 activity in tobacco hornworms. Arch. Insect Biochem. Physiol. 2003, 52, 1–6. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Jin, S.; Lu, Y.; Peng, Y.; Zhao, L.; Wang, X. Cannabidiol protects against Alzheimer’s disease in C. elegans via ROS scavenging activity of its phenolic hydroxyl groups. Eur. J. Pharmacol. 2022, 919, 174829. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Day of Supplementation | se± | ||||
|---|---|---|---|---|---|---|
| TAC | SOD | CAT | GPx | GSH | ||
| CSt | 2 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 |
| 7 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 14 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 21 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 28 days | 0.367377 | 0.05739 | 0.192164 | 0.279045 | 0.264402 | |
| 35 days | - | - | - | - | - | |
| CSy | 2 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 |
| 7 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 14 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 21 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 28 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 35 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| C | 2 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 |
| 7 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 14 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 21 days | 0.348525 | 0.054445 | 0.182303 | 0.264725 | 0.250834 | |
| 28 days | 0.492888 | 0.076997 | 0.257815 | 0.374378 | 0.354732 | |
| 35 days | - | - | - | - | - | |
| Groups | Day of Supplementation | se± | ||
|---|---|---|---|---|
| Ca2+ | Mg2+ | P | ||
| C | 2 days | 0.336983 | 0.016605 | 0.080677 |
| 7 days | 0.336983 | 0.016605 | 0.080677 | |
| 14 days | 0.336983 | 0.016605 | 0.080677 | |
| 21 days | 0.336983 | 0.016605 | 0.080677 | |
| 28 days | 0.336983 | 0.016605 | 0.080677 | |
| 35 days | - | - | - | |
| CSt | 2 days | 0.336983 | 0.016605 | 0.080677 |
| 7 days | 0.336983 | 0.016605 | 0.080677 | |
| 14 days | 0.336983 | 0.016605 | 0.080677 | |
| 21 days | 0.336983 | 0.016605 | 0.080677 | |
| 28 days | 0.355211 | 0.017503 | 0.085041 | |
| 35 days | - | - | - | |
| CSy | 2 days | 0.336983 | 0.016605 | 0.080677 |
| 7 days | 0.336983 | 0.016605 | 0.080677 | |
| 14 days | 0.336983 | 0.016605 | 0.080677 | |
| 21 days | 0.336983 | 0.016605 | 0.080677 | |
| 28 days | 0.336983 | 0.016605 | 0.080677 | |
| 35 days | 0.336983 | 0.016605 | 0.080677 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skowronek, P.; Strachecka, A. Cannabidiol (CBD) Supports the Honeybee Worker Organism by Activating the Antioxidant System. Antioxidants 2023, 12, 279. https://doi.org/10.3390/antiox12020279
Skowronek P, Strachecka A. Cannabidiol (CBD) Supports the Honeybee Worker Organism by Activating the Antioxidant System. Antioxidants. 2023; 12(2):279. https://doi.org/10.3390/antiox12020279
Chicago/Turabian StyleSkowronek, Patrycja, and Aneta Strachecka. 2023. "Cannabidiol (CBD) Supports the Honeybee Worker Organism by Activating the Antioxidant System" Antioxidants 12, no. 2: 279. https://doi.org/10.3390/antiox12020279
APA StyleSkowronek, P., & Strachecka, A. (2023). Cannabidiol (CBD) Supports the Honeybee Worker Organism by Activating the Antioxidant System. Antioxidants, 12(2), 279. https://doi.org/10.3390/antiox12020279