

Effect of In Ovo Administration of a Multi-Strain Probiotic and Zinc Glycine Chelate on Antioxidant Capacity and Selected Immune Parameters in Newly Hatched Chicks

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Incubation Period
2.3. In Ovo Inoculations
2.4. Birds and Housing
2.5. Collection of Blood and Tissue Samples
2.6. Assay of IL-10, IFN-γ, TNF-α, Corticosterone, Hsp 70, Catalase and Superoxide Dismutase (SOD) in Chicken Serum and Tissues
2.7. Total Antioxidant Capacity (T-AOC)
2.8. Protein Content
2.9. Content of Sulfhydryl Groups
2.10. Content of Bityrosine Bridges
2.11. Content of Formylkynurenine
2.12. Content of Hydroperoxides
2.13. Statistical Analysis
3. Results
3.1. Mean Activity of Superoxide Dismutase (SOD) and Catalase (CAT) in the Serum and Tissues of Chicks
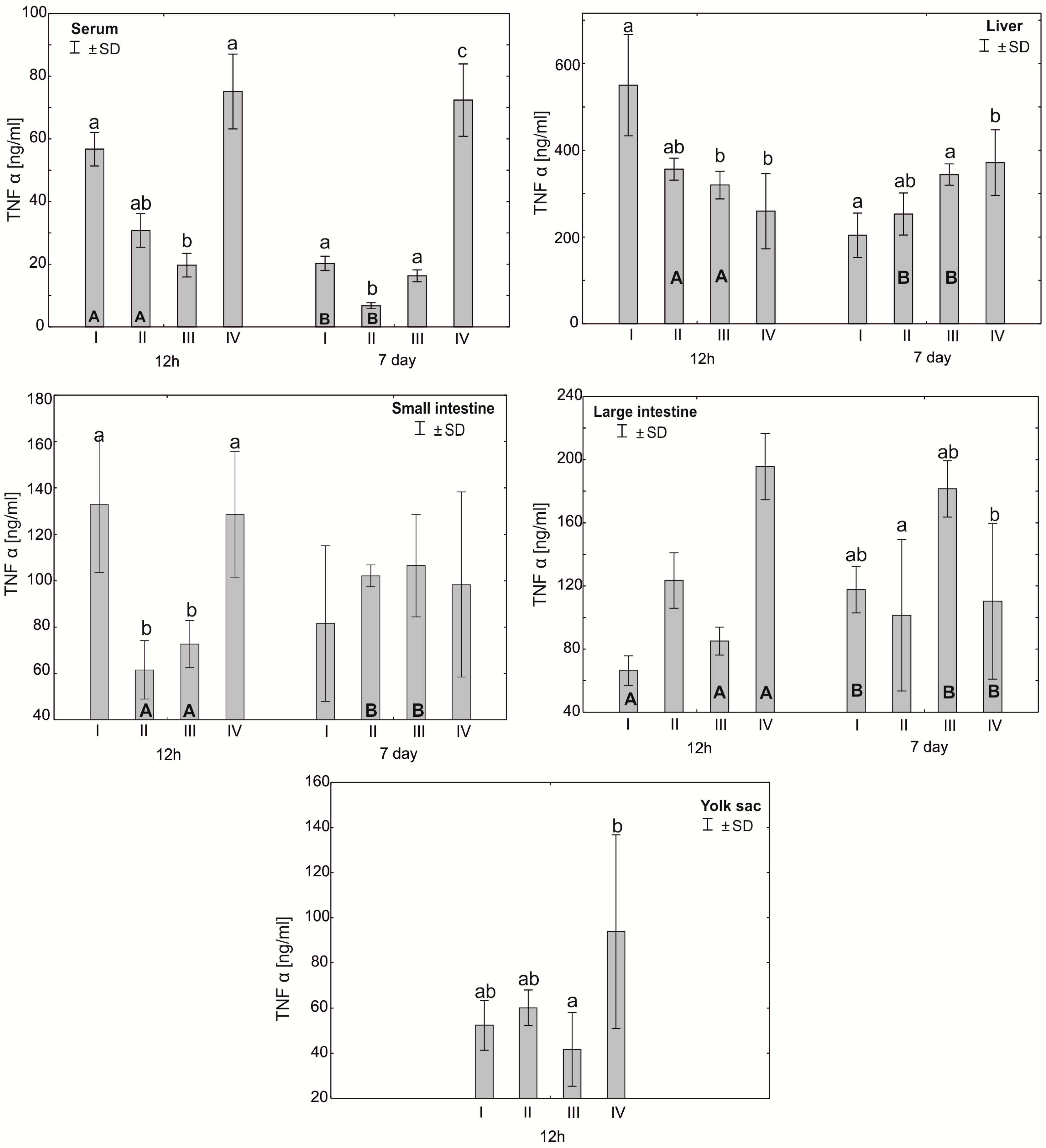
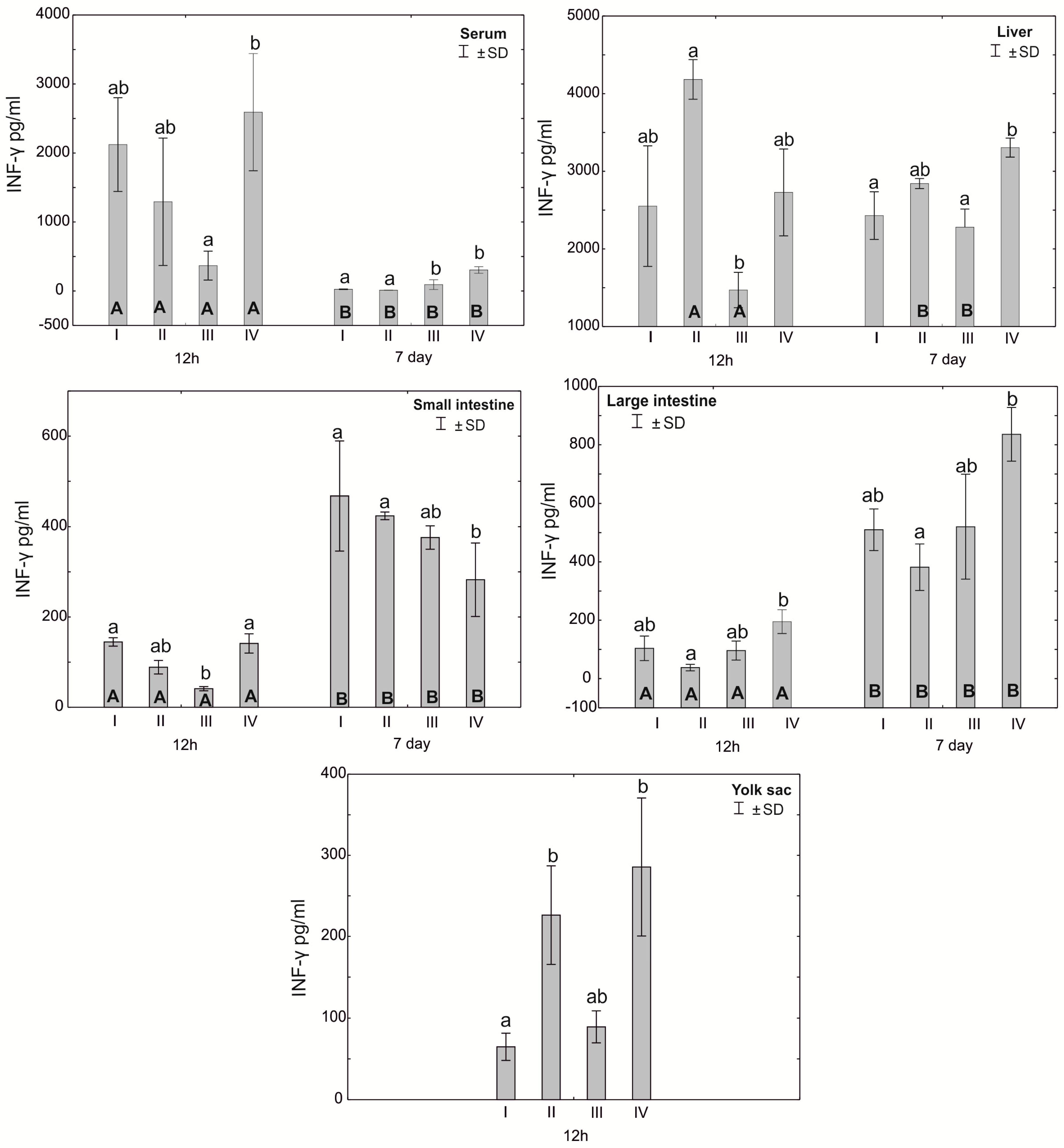
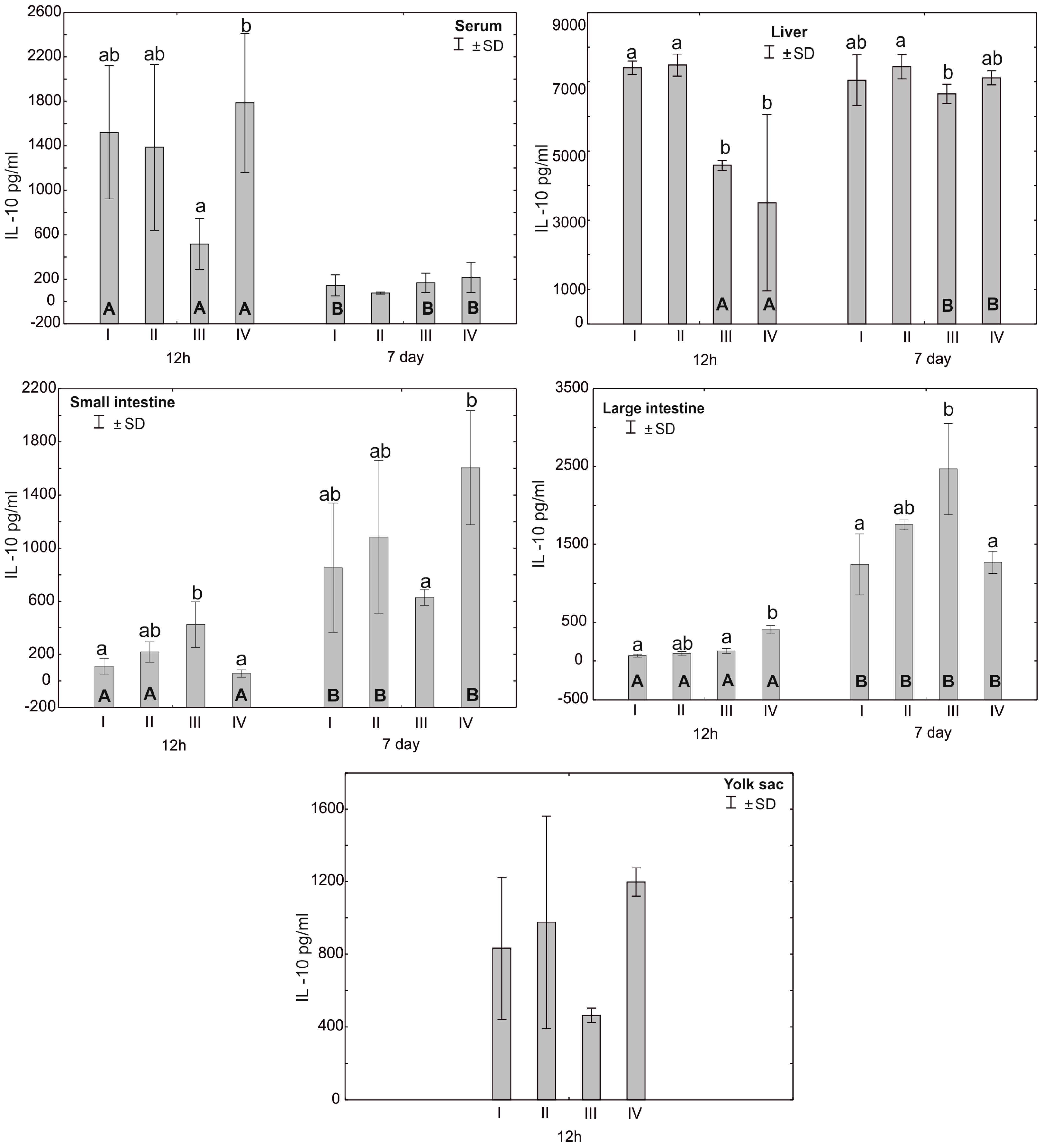
3.2. Concentration of Cytokines TNF-α, IFN-γ and IL-10 in the Serum, Liver, Intestines and Yolk Sacs of Chickens
3.3. Corticosterone (CORT) Concentration in the Serum and Tissues of Chicks
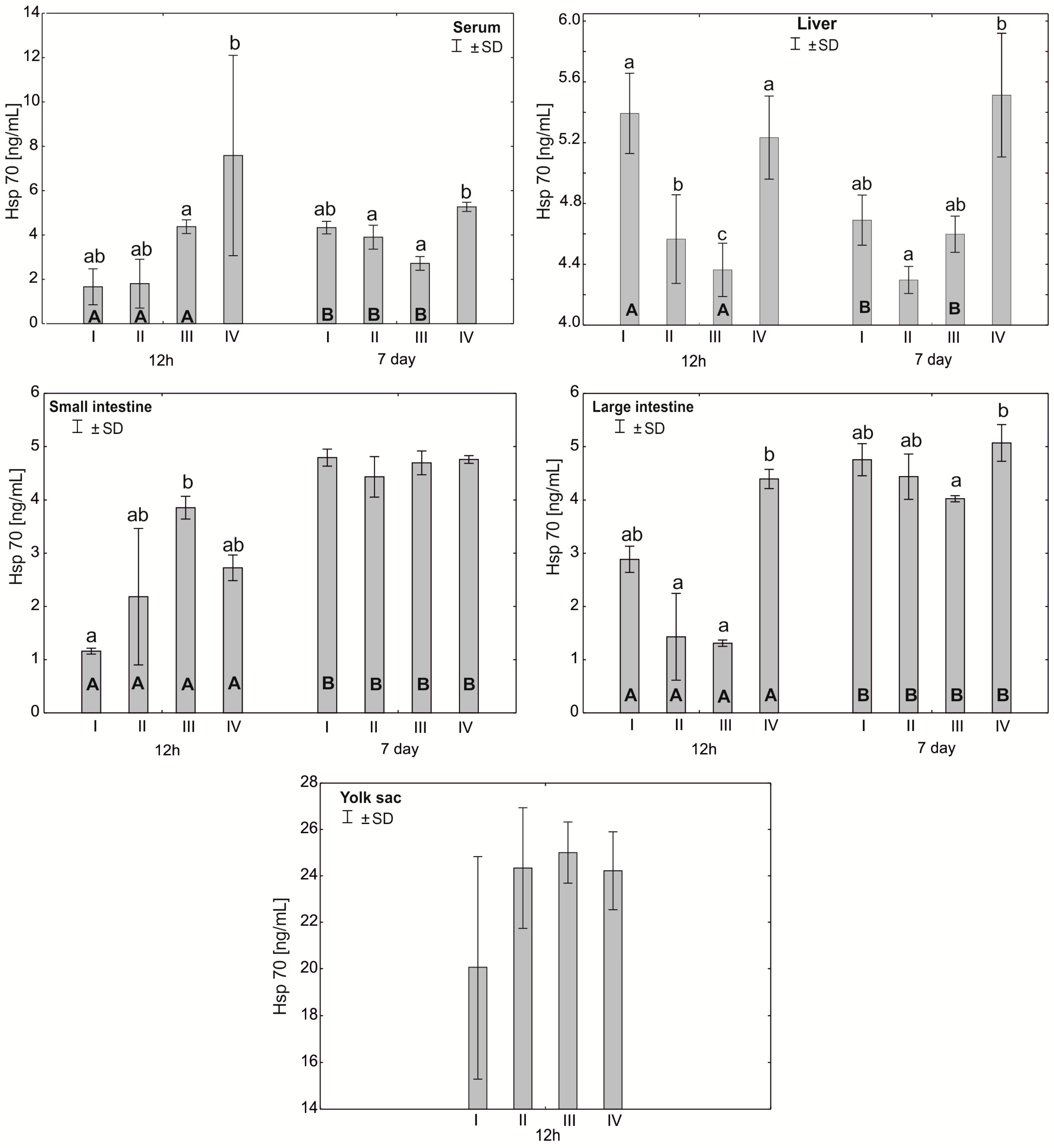
3.4. Heat Shock Protein (Hsp70) Concentration in the Serum and Tissues of Chicks
3.5. Levels of Total Antioxidant Capacity (T-AOC), Protein, Sulfhydryl (SH) Groups, Bityrosine Bridges, Formylkynurenine and Hydroperoxides/DEPPD in the Serum and Tissues of Chicks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elson, H. Poultry welfare in intensive and extensive production systems. World’s Poult. Sci. J. 2015, 71, 449–460. [Google Scholar] [CrossRef]
- Lukić, M.; Petričević, V.; Delić, N.; Tolimir, N.; Dosković, V.; Rakonjac, S.; Škrbić, Z. How does the choice of genotype and feed in the local market affect broiler performance and the farm economy? A case study in Serbia. Agriculture 2022, 12, 843. [Google Scholar] [CrossRef]
- Deeming, D.C.; Pike, T.W. Embryonic growth and antioxidant provision in avian eggs. Biol. Lett. 2013, 9, 20130757. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lian, G. ROS and diseases: Role in metabolism and energy supply. Mol. Cell. Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef]
- Surai, P.F.; Fisinin, V.I.; Karadas, F. Antioxidant systems in chick embryo development. Part 1. Vitamin E, carotenoids and selenium. Anim. Nutr. 2016, 2, 1–11. [Google Scholar] [CrossRef]
- Surai, P.F.; Speake, B.K.; Noble, R.C.; Sparks, N.H. Tissue-specific antioxidant profiles and susceptibility to lipid peroxidation of the newly hatched chick. Biol. Trace Elem. Res. 1999, 68, 63–78. [Google Scholar] [CrossRef]
- Surai, P.F. Tissue-specific changes in the activities of antioxidant enzymes during the development of the chicken embryo. Br. Poult. Sci. 1999, 40, 397–405. [Google Scholar] [CrossRef]
- Surai, P.F. Vitamin E in avian reproduction. Poult. Avian Biol. Rev. 1999, 10, 1–60. [Google Scholar]
- Surai, P.F. Effect of the selenium and vitamin E content of the maternal diet on the antioxidant system of the yolk and the developing chick. Br. Poult. Sci. 2000, 41, 235–243. [Google Scholar] [CrossRef]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidant action of carnitine: Molecular mechanisms and practical applications. EC Vet. Sci. 2015, 2, 66–84. [Google Scholar]
- Karadas, F.; Surai, P.F.; Sparks, N.H. Changes in broiler chick tissue concentrations of lipid-soluble antioxidants immediately post-hatch. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 160, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Speake, B.K.; Murray, A.M.; Noble, R.C. Transport and transformations of yolk lipids during development of the avian embryo. Prog. Lipid Res. 1998, 37, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ding, X.; Bai, S.; Wang, J.; Zeng, Q.; Peng, H.; Xuan, Y.; Su, Z.; Zhang, K. The effects of broiler breeder dietary vitamin E and egg storage time on the quality of eggs and newly hatched chicks. Animals 2020, 10, 1409. [Google Scholar] [CrossRef]
- Matsue, H.; Edelbaum, D.; Shalhevert, D.; Mizumoto, N.; Yang, C.; Mummert, M.E.; Oeda, J.; Masayasu, H.; Takashima, A. Generation and function of reactive oxygen species in dendritic cells during antigen presentation. J. Immunol. 2003, 171, 3010–3018. [Google Scholar] [CrossRef]
- Arai, T.; Yamada, H.; Namba, T.; Mori, H.; Ishii, H.; Yamashita, K.; Sasada, M.; Makino, K.; Fukuda, K. Effects of intracellular reactive oxygen species generated by 6-formylptrin on T cell functions. Biochem. Pharmacol. 2004, 67, 1185–1193. [Google Scholar] [CrossRef]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef]
- Malheiro, R.D.; Ferket, P.R.; Goncalves, F.M. Oxidative stress protection of embryos by “In ovo” supplementation. In Proceedings of the World’s Poultry Congress Salvador, Bahia, Brazil, 5–9 August 2012. [Google Scholar]
- Lee, S.H.; Lillehoj, H.S.; Jang, S.I.; Jeong, M.S.; Xu, S.Z.; Kim, J.B.; Park, H.J.; Kim, H.R.; Lillehoj, E.P.; Bravo, D.M. Effects of in ovo injection with selenium on immune and antioxidant responses during experimental necrotic enteritis in broiler chickens. Poult. Sci. 2014, 93, 1113–1121. [Google Scholar] [CrossRef]
- Yigit, A.; Panda, A.; Cherian, G. The avian embryo and its antioxidant defense system. World’s Poult. Sci. J. 2014, 70, 563–574. [Google Scholar] [CrossRef]
- Wilson, K.M.; Rodrigues, D.R.; Briggs, W.N.; Duff, A.F.; Chasser, K.M.; Bielke, L.R. Evaluation of the impact of in ovo administered bacteria on microbiome of chicks through 10 days of age. Poult. Sci. 2019, 98, 5949–5960. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant properties of probiotic bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Mishra, P.; Jha, R. In ovo feeding as a tool for improving performance and gut health of poultry: A review. Front. Vet. Sci. 2021, 8, 754246. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.M.; Paswan, V.K.; Attia, Y.A.; Abdel-Moneim, A.E.; Abougabal, M.S.; Sharaf, M.; Elmazoudy, R.; Alghafari, W.T.; Osman, M.A.; Farag, M.R.; et al. Managing gut microbiota through in ovo nutrition influences early-life programming in broiler chickens. Animals 2021, 11, 3491. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef]
- Atsbeha, A.T.; Hailu, T.G. The impact of effective microorganisms (EM) on egg quality and laying performance of chickens. Int. J. Food Sci. 2021, 2021, 8895717. [Google Scholar] [CrossRef]
- Esatu, W.; Melesse, A.; Dessie, T. Effect of effective microorganisms on growth parameters and serum cholesterol levels in broilers. Afr. J. Agric. Res. 2011, 6, 3841–3846. [Google Scholar]
- Chen, X.; He, C.; Zhang, K.; Wang, J.; Ding, X.; Zeng, Q.; Peng, H.; Bai, J.; Lv, L.; Xuan, Y.; et al. Comparison of zinc bioavailability in zinc-glycine and zinc-methionine chelates for broilers fed with a corn-soybean meal diet. Front. Physiol. 2022, 13, 983954. [Google Scholar] [CrossRef]
- Huang, Y.L.; Lu, L.; Li, S.F.; Luo, X.G.; Liu, B. Relative bioavailabilities of organic zinc sources with different chelation strengths for broilers fed a conventional corn-soybean meal diet. J. Anim. Sci. 2009, 87, 2038–2046. [Google Scholar] [CrossRef]
- Star, L.; van der Klis, J.D.; Rapp, C.; Ward, T.L. Bioavailability of organic and inorganic zinc sources in male broilers. Poult. Sci. 2012, 91, 3115–3120. [Google Scholar] [CrossRef]
- Jarosz, Ł.; Marek, A.; Grądzki, Z.; Kwiecień, M.; Żylinska, B.; Kaczmarek, B. Effect of feed supplementation with zinc glycine chelate and zinc sulfate on cytokine and immunoglobulin gene expression profiles in chicken intestinal tissue. Poult. Sci. 2017, 96, 4224–4235. [Google Scholar] [CrossRef]
- Jarosz, Ł.; Marek, A.; Grądzki, Z.; Kwiecień, M.; Kalinowski, M. The effect of feed supplementation with zinc chelate and zinc sulphate on selected humoral and cell-mediated immune parameters and cytokine concentration in broiler chickens. Res. Vet. Sci. 2017, 112, 59–65. [Google Scholar] [CrossRef]
- Jarosz, Ł.; Marek, A.; Grądzki, Z.; Laskowska, E.; Kwiecień, M. Effect of zinc sulfate and zinc glycine chelate on concentrations of acute phase proteins in chicken serum and liver tissue. Biol. Trace Elem. Res. 2019, 187, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Ciszewski, A.; Jarosz, Ł.; Marek, A.; Michalak, K.; Grądzki, Z.; Kaczmarek, B.; Rysiak, A. Effect of combined in ovo administration of zinc glycine chelate (Zn-Gly) and a multistrain probiotic on the modulation of cellular and humoral immune responses in broiler chickens. Poult. Sci. 2023, 102, 102823. [Google Scholar] [CrossRef] [PubMed]
- Weese, J.S.; Martin, H. Assessment of commercial probiotic bacterial contents and label accuracy. Can. Vet. J. 2011, 52, 43–46. [Google Scholar] [PubMed]
- Alizadeh, M.; Bavananthasivam, J.; Shojadoost, B.; Astill, J.; Taha-Abdelaziz, K.; Alqazlan, N.; Boodhoo, N.; Doost, J.S.; Sharif, S. In ovo and oral administration of probiotic Lactobacilli modulate cell- and antibody-mediated immune responses in newly hatched chicks. Front. Immunol. 2021, 12, 664387. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Astill, J.; Alqazlan, N.; Shojadoost, B.; Taha-Abdelaziz, K.; Bavananthasivam, J.; Doost, J.S.; Sedeghiisfahani, N.; Sharif, S. In ovo co-administration of vitamins (A and D) and probiotic lactobacilli modulates immune responses in broiler chickens. Poult. Sci. 2022, 101, 101717. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Katalinic, V.; Modun, D.; Music, I.; Boban, M. Gender differences in antioxidant capacity of rat tissues determined by 2,2′-azinobis (3-ethylbenzothiazoline 6-sulfonate; ABTS) and ferric reducing antioxidant power (FRAP) assays. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 140, 47–52. [Google Scholar] [CrossRef]
- Gornal, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret method reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Diplock, A.T.; Symons, M.C.R. Techniques in Free Radical Research, 1st ed.; Elsevier: Amsterdam, The Netherlands, 1991. [Google Scholar]
- Alberti, A.; Bolognini, L.; Macciantelli, D.; Caratelli, M. The radical cation of N,N-diethyl-para-phenylendiamine: A possibile indicator of oxidative stress in biological samples. Res. Chem. Intermed. 2000, 26, 253–267. [Google Scholar] [CrossRef]
- Givisiez, P.E.N.; Moreira Filho, A.L.B.; Santos, M.R.B.; Oliveira, H.B.; Ferket, P.R.; Oliveira, C.J.B.; Malheiros, R.D. Chicken embryo development: Metabolic and morphological basis for in ovo feeding technology. Poult. Sci. 2020, 99, 6774–6782. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth Defects Res. C Embryo Today 2007, 81, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Cherian, G. Nutrition and metabolism in poultry: Role of lipids in early diet. J. Anim. Sci. Biotechnol. 2015, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Shokraneh, M.; Sadeghi, A.A.; Mousavi, S.N.; Esmaeilkhanian, S.; Chamani, M. Effects of in ovo injection of nano-selenium and nano-zinc oxide and high eggshell temperature during late incubation on antioxidant activity, thyroid and gluco-corticoid hormones and some blood metabolites in broiler hatchlings. Acta Sci.-Anim. Sci. 2020, 42, e46029. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.F.; Wang, S.X.; Zhang, D.Y.; Liu, H.; Shan, D.C.; Wang, Y.M. Lactobacillus plantarum ZLP001: In vitro assessment of antioxidant capacity and effect on growth performance and antioxidant status in weaning piglets. Asian-Australas. J. Anim. Sci. 2012, 25, 1153–1158. [Google Scholar] [CrossRef]
- Hou, C.L.; Zhang, J.; Liu, X.T.; Liu, H.; Zeng, X.F.; Qiao, S.Y. Superoxide dismutase recombinant Lactobacillus fermentum ameliorates intestinal oxidative stress through inhibiting NF-κB activation in a trinitrobenzene sulphonic acid-induced colitis mouse model. J. Appl. Microbiol. 2014, 116, 1621–1631. [Google Scholar] [CrossRef]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic acid bacteria antimicrobial compounds: Characteristics and applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Hojyo, S.; Fukada, T. Roles of zinc signaling in the immune system. J. Immunol. Res. 2016, 2016, 6762343. [Google Scholar] [CrossRef]
- Pender, C.M.; Kim, S.; Potter, T.D.; Ritzi, M.M.; Young, M.; Dalloul, R.A. In ovo supplementation of probiotics and its effectson performance and immune-related gene expression in broiler chicks. Poult. Sci. 2017, 96, 1052–1062. [Google Scholar] [CrossRef]
- Shehata, A.M.; Paswan, V.K.; Attia, Y.A.; Abougabal, M.S.; Khamis, T.; Alqosaibi, A.I.; Alnamshan, M.M.; Elmazoudy, R.; Abaza, M.A.; Salama, E.A.A.; et al. In ovo inoculation of Bacillus subtilis and raffinose affects growth performance, cecal microbiota, volatile fatty acid, ileal morphology and gene expression, and sustainability of broiler chickens (Gallus gallus). Front. Nutr. 2022, 9, 903847. [Google Scholar] [CrossRef]
- Ghiselli, A.; Serafini, M.; Natella, F.; Scaccini, C. Total antioxidant capacity as a tool to assess redox status: Critical view and experimental data. Free Radic. Biol. Med. 2000, 29, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Hegde, M.N.; Hegde, N.D.; Ashok, A.; Shetty, S. Evaluation of total antioxidant capacity of saliva and serum in caries-free and caries-active adults: An in-vivo study. Indian J. Dent. Res. 2013, 24, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Peluso, I.; Roguzzini, A. Salivary and urinary total antioxidant capacity as biomarkers of oxidative stress in humans. Pathol. Res. Int. 2016, 2016, 5480267. [Google Scholar] [CrossRef] [PubMed]
- Kucuk, O.; Sahin, N.; Sahin, K. Supplemental zinc and vitamin A can alleviate negative effects of heat stress in broiler chickens. Biol. Trace Elem. Res. 2003, 94, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, K.Z.; Edens, F.W.; Eisen, E.J.; Havenstein, G.B. Ascorbic acid decreases heat shock protein 70 and plasma corticosterone response in broilers (Gallus gallus domesticus) subjected to cyclic heat stress. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 137, 35–42. [Google Scholar] [CrossRef]
- Miao, Q.; Si, X.; Xie, Y.; Chen, L.; Liu, Z.; Liu, L.; Tang, X.; Zhang, H. Effects of acute heat stress at different ambient temperature on hepatic redox status in broilers. Poult. Sci. 2020, 99, 4113–4122. [Google Scholar] [CrossRef]
- Li, T.; He, W.; Liao, X.; Lin, X.; Zhang, L.; Lu, L.; Guo, Y.; Liu, Z.; Luo, X. Zinc alleviates the heat stress of primary cultured hepatocytes of broiler embryos via enhancing the antioxidant ability and attenuating the heat shock responses. Anim. Nutr. 2021, 7, 621–630. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, L.; Ding, X.; Wang, Y.; Xu, Y.; Li, D.; Liang, S.; Wang, Y.; Zhang, L.; Fu, A.; et al. Mechanisms underlying the protective effect of maternal zinc (ZnSO4 or Zn-Gly) against heat stress-induced oxidative stress in chicken embryo. Antioxidants 2022, 11, 1699. [Google Scholar] [CrossRef]
- Yu, Y.; Lu, L.; Li, S.F.; Zhang, L.Y.; Luo, X.G. Organic zinc absorption by the intestine of broilers in vivo. Br. J. Nutr. 2017, 117, 1086–1094. [Google Scholar] [CrossRef]
- Yang, P.C.; Tu, Y.H.; Perdue, M.H.; Oluwole, C.; Struiksma, S. Regulatory effect of heat shock protein 70 in stress-induced rat intestinal epithelial barrier dysfunction. N. Am. J. Med. Sci. 2009, 1, 9–15. [Google Scholar]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Li, D.L.; Wang, J.S.; Liu, L.J.; Li, K.; Xu, Y.B.; Ding, X.Q.; Wang, Y.Y.; Zhang, Y.F.; Xie, L.Y.; Liang, S.; et al. Effects of early post-hatch feeding on the growth performance, hormone secretion, intestinal morphology, and intestinal microbiota structure in broilers. Poult. Sci. 2022, 101, 102133. [Google Scholar] [CrossRef] [PubMed]
- Gatin, A.; Billault, I.; Duchambon, P.; van der Rest, G.; Sicard-Roselli, C. Oxidative radicals (HO• or N3•) induce several di-tyrosine bridge isomers at the protein scale. Free Radic. Biol. Med. 2021, 162, 461–470. [Google Scholar] [CrossRef]
- Hajsl, M.; Hlavackova, A.; Broulikova, K.; Sramek, M.; Maly, M.; Dyr, J.E.; Suttnar, J. Tryptophan metabolism, inflammation, and oxidative stress in patients with neurovascular disease. Metabolites 2020, 10, 208. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, A.; Jamioł, M.; Kankofer, M. Antioxidative and oxidative profiles in plasma and saliva in dairy cows during pregnancy. Animals 2021, 11, 3204. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Replicates/Number of Eggs per Replicate N= | Volume and Solution of Bioactive Compounds Injected In Ovo at 17 DOI * |
|---|---|---|
| I—control group | 10/35 350 | 500 µL 0.9% NaCl |
| II | 10/35 350 | 100 µL multi-strain probiotic + 400 µL MQ water |
| III | 10/35 350 | 100 µL multi-strain probiotic + 100 µL Zn-Gly + 300 µL MQ water |
| IV | 10/35 350 | 100 µL Zn-Gly + 400 µL MQ water |
| Time | Tissue | Formylkynurenine (μg/mg Protein in Homogenate Tissue) | Sulfhydryl Groups/SH (mmol/g Protein in Homogenate Tissue) | |||||||
| N= | Groups | |||||||||
| I | II | III | IV | I | II | III | IV | |||
| 12 h | Serum | 30 | 0.21 ± 0.02 | 0.26 ± 0.05 | 0.23 ± 0.06 | 0.21 ± 0.01 | 10.41 ± 0.43 a *** | 11.27 ± 0.49 a *** | 8.74 ± 0.37 b *** | 9.59 ± 0.46 b *** |
| Liver | 1.10 ± 0.44 a *** | 0.76 ± 0.02 a *** | 0.51 ± 0.03 ab *** | 0.44 ± 0.01 b *** | 62.82 ± 1.34 a *** | 89.14 ± 0.20 b *** | 72.0 ± 2.15 ab *** | 80.33 ± 2.92 b *** | ||
| Pectoral muscle | 0.42 ± 0.04 | 0.41 ± 0.14 | 0.29 ± 0.12 | 0.42 ± 0.07 | 83.5 ± 6.70 a ** | 77.74 ± 0.49 a ** | 94.27 ± 5.15 ab ** | 98.08 ± 3.46 b ** | ||
| Heart | 0.35 ± 0.09 | 0.45 ± 0.08 | 0.34 ± 0.05 | 0.38 ± 0.03 | 66.49 ± 12.36 | 73.08 ± 5.69 | 67.49 ± 2.21 | 73.97 ± 2.81 | ||
| Small intestine | 0.63 ± 0.09 ab *** | 1.27 ± 0.05 a *** | 0.47 ± 0.14 b *** | 0.30 ± 0.01 b *** | 67.98 ± 0.57 a *** | 83.65 ± 4.29 ab *** | 94.81 ± 5.62 b *** | 86.78 ± 5.06 b *** | ||
| Yolk sac | 1.23 ± 0.35 b ** | 1.64 ± 0.50 ab ** | 2.59 ± 0.21 a ** | 2.0 ± 0.66 ab ** | 20.33 ± 3.49 a ** | 22.62 ± 0.51 a ** | 13.55 ± 2.8 b ** | 25.10 ± 6.59 ab ** | ||
| After 7 days | Serum | 30 | 0.13 ± 0.03 a * | 0.16 ± 0.04 abc * | 0.17 ± 0.02 * c | 0.19 ± 0.03 b * | 11.8 ± 0.68 a * | 10.31 ± 0.93 b * | 10.72 ± 0.34 b * | 11.91 ± 1.28 a * |
| Liver | 0.78 ± 0.03 abc *** | 0.56 ± 0.09 b *** | 0.87 ± 0.08 a *** | 0.53 ± 0.20 c *** | 67.58 ± 5.06 a ** | 71.91 ± 12.33 ab ** | 63.61 ± 10.57 a ** | 86.61 ± 1.82 b ** | ||
| Pectoral muscle | 0.22 ± 0.05 | 0.18 ± 0.01 | 0.3 ± 0.18 | 0.24 ± 0.08 | 101.27 ± 2.88 | 102.26 ± 3.24 | 101.85 ± 5.27 | 103.29 ± 6.64 | ||
| Heart | 0.52 ± 0.06 a *** | 0.37 ± 0.03 abc *** | 0.34 ± 0.00 c *** | 0.5 ± 0.04 b *** | 95.56 ± 1.07 a *** | 75.04 ± 8.77 b *** | 88.84 ± 1.88 ab *** | 88.76 ± 1.66 ab *** | ||
| Small intestine | 0.58 ± 0.11 a *** | 0.43 ± 0.05 ab *** | 0.34 ± 0.02 b *** | 0.53 ± 0.03 a *** | 70.5 ± 3.73 | 71.05 ± 6.56 | 70.75 ± 2.24 | 74.06 ± 0.15 | ||
| Time | Tissue | Bityrosine bridges (µg/mg protein in homogenate tissue) | T-AOC (μmol/g protein in homogenate tissue) | |||||||
| N= | Groups | |||||||||
| I | II | III | IV | I | II | III | IV | |||
| 12 h | Serum | 30 | 0.73 ± 0.10 ab * | 0.86 ± 0.13 ab * | 0.61 ± 0.15 a * | 0.87 ± 0.08 b * | 88.37 ± 7.00 ab *** | 94.65 ± 2.72 ab *** | 111.8 ± 7.60 a *** | 69.79 ± 1.38 b *** |
| Liver | 6.87 ± 2.24 a *** | 6.88 ± 0.19 a *** | 3.77 ± 0.15 b *** | 3.87 ± 0.05 b *** | 56.07 ± 22.62 | 42.65 ± 5.20 | 40.76 ± 5.30 | 57.83 ± 5.48 | ||
| Pectoral muscle | 3.79 ± 1.43 | 4.31 ± 1.90 | 3.26 ± 2.03 | 3.19 ± 0.43 | 31.38 ± 1.11 ab *** | 23.66 ± 2.46 b *** | 23.45 ± 1.61 b *** | 32.40 ± 0.61 a *** | ||
| Heart | 2.68 ± 0.21 a * | 3.72 ± 0.64 b * | 3.19 ± 0.75 ab * | 3.62 ± 0.57 ab * | 26.47 ± 3.16 ab * | 25.12 ± 2.44 ab * | 24.20 ± 1.12 b * | 30.83 ± 2.68 a * | ||
| Small intestine | 5.58 ± 1.90 ab *** | 9.21 ± 2.99 a *** | 4.14 ± 1.07 ab *** | 2.75 ± 0.29 b *** | 57.31 ± 6.56 a *** | 99.75 ± 13.51 b *** | 97.53 ± 14.11 b *** | 54.48 ± 1.98 a *** | ||
| Yolk sac | 4.44 ± 0.39 a ** | 5.94 ± 1.54 ab ** | 7.31 ± 0.40 b ** | 6.79 ± 0.22 ab ** | 21.38 ± 1.01 a ** | 45.10 ± 11.07 b ** | 44.19 ± 7.84 ab ** | 49.96 ± 6.81 b ** | ||
| After 7 days | Serum | 30 | 0.32 ± 0.16 | 0.46 ± 0.02 | 0.5 ± 0.11 | 0.46 ± 0.06 | 30.05 ± 2.18 a * | 44.13 ± 0.55 b * | 37.14 ± 2.99 ab * | 44.73 ± 14.51 ab * |
| Liver | 5.38 ± 0.15 a *** | 3.30 ± 0.70 b *** | 4.02 ± 0.43 ab *** | 1.92 ± 0.50 b *** | 77.95 ± 5.96 a ** | 82.30 ± 16.71 ab *** | 78.61 ± 9.84 a *** | 108.71 ± 6.29 b *** | ||
| Pectoral muscle | 1.92 ± 0.11 a * | 2.12 ± 0.20 ab * | 2.48 ± 0.30 b * | 2.19 ± 0.33 ab * | 18.16 ± 1.85 ab *** | 15.71 ± 0.17 b *** | 19.04 ± 0.26 ab *** | 24.76 ± 2.66 a *** | ||
| Heart | 2.30 ± 0.23 ab *** | 1.59 ± 0.70 a *** | 1.53 ± 0.12 a *** | 3.99 ± 0.24 b *** | 26.61 ± 0.66 a * | 24.97 ± 1.55 ab * | 22.30 ± 0.71 b * | 24.37 ± 3.16 ab * | ||
| Small intestine | 3.85 ± 0.45 | 3.00 ± 0.34 | 2.39 ± 0.10 | 3.88 ± 0.24 | 55.63 ± 8.46 ab * | 61.96 ± 8.85 a * | 58.57 ± 1.65 ab * | 48.38 ± 1.69 b * | ||
| Time | Tissue | Protein (g/L in homogenate tissue) | Hydroperoxides/DEPPD (µmol/g protein in homogenate tissue) | |||||||
| N= | Groups | |||||||||
| I | II | III | IV | I | II | III | IV | |||
| 12 h | Serum | 30 | 25.71 ± 0.68 a ** | 25.93 ± 1.03 a ** | 29.28 ± 1.57 b ** | 27.9 ± 0.72 ab ** | 0.04 ± 0.01 a ** | 0.04 ± 0.01 a ** | 0.05 ± 0.02 ab ** | 0.10 ± 0.01 b ** |
| Liver | 51.41 ± 2.00 a *** | 47.34 ± 7.90 a *** | 65.52 ± 4.98 b *** | 63.46 ± 2.33 b *** | 0.28 ± 0.01 | 0.27 ± 0.01 | 0.27 ± 0.03 | 0.29 ± 0.01 | ||
| Pectoral muscle | 6.36 ± 0.18 a *** | 8.57 ± 1.08 b *** | 11.56 ± 0.87 b *** | 7.06 ± 0.18 a *** | 0.15 ± 0.03 a *** | 0.18 ± 0.01 a *** | 0.25 ± 0.02 b *** | 0.23 ± 0.02 b *** | ||
| Heart | 12.49 ± 2.59 a ** | 8.36 ± 0.55 b ** | 12.37 ± 0.12 a ** | 9.76 ± 0.88 ab ** | 0.18 ± 0.00 a *** | 0.20 ± 0.01 ab *** | 0.22 ± 0.03 ab *** | 0.26 ± 0.01 b *** | ||
| Small intestine | 7.51 ± 1.83 ab *** | 5.49 ± 1.12 a *** | 15.45 ± 1.21 b *** | 15.37 ± 0.25 b *** | 0.21 ± 0.001 | 0.20 ± 0.01 | 0.20 ± 0.01 | 0.21 ± 0.01 | ||
| Yolk sac | 41.26 ± 0.17 a *** | 26.45 ± 9.67 ab *** | 19.05 ± 0.35 ab *** | 10.95 ± 3.67 b *** | 0.20 ± 0.001 a *** | 0.19 ± 0.01 ab *** | 0.18 ± 0.003 b *** | 0.18 ± 0.01 b *** | ||
| After 7 days | Serum | 30 | 27.33 ± 1.07 a * | 26.51 ± 1.86 ab * | 26.94 ± 0.45 ab * | 24.76 ± 0.75 b * | 0.07 ± 0.03 * | 0.09 ± 0.04 * | 0.05 ± 0.006 * | 0.06 ± 0.02 * |
| Liver | 37.09 ± 7.99 | 39.25 ± 7.06 | 38.77 ± 10.37 | 28.84 ± 2.00 | 0.37 ± 0.01 a *** | 0.32 ± 0.002 a *** | 0.30 ± 0.01 b *** | 0.31 ± 0.01 b *** | ||
| Pectoral muscle | 22.08 ± 2.29 a *** | 24.07 ± 1.49 a *** | 14.38 ± 0.27 b *** | 20.51 ± 0.37 ab *** | 0.23 ± 0.02 | 0.23 ± 0.01 | 0.22 ± 0.03 | 0.21 ± 0.005 | ||
| Heart | 10.38 ± 0.19 a ** | 10.31 ± 1.53 a ** | 12.62 ± 0.46 b ** | 11.90 ± 1.06 ab ** | 0.30 ± 0.01 ab ** | 0.32 ± 0.01 a ** | 0.30 ± 0.002 ab ** | 0.28 ± 0.01 b ** | ||
| Small intestine | 23.32 ± 1.64 ab ** | 29.73 ± 6.81 a ** | 25.98 ± 1.26 a ** | 20.94 ± 1.48 b ** | 0.24 ± 0.00 a *** | 0.23 ± 0.00 ab *** | 0.22 ± 0.01 b *** | 0.25 ± 0.003 a *** | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciszewski, A.; Jarosz, Ł.S.; Bielecka, A.; Marek, A.; Szymczak, B.; Grądzki, Z.; Rysiak, A. Effect of In Ovo Administration of a Multi-Strain Probiotic and Zinc Glycine Chelate on Antioxidant Capacity and Selected Immune Parameters in Newly Hatched Chicks. Antioxidants 2023, 12, 1905. https://doi.org/10.3390/antiox12111905
Ciszewski A, Jarosz ŁS, Bielecka A, Marek A, Szymczak B, Grądzki Z, Rysiak A. Effect of In Ovo Administration of a Multi-Strain Probiotic and Zinc Glycine Chelate on Antioxidant Capacity and Selected Immune Parameters in Newly Hatched Chicks. Antioxidants. 2023; 12(11):1905. https://doi.org/10.3390/antiox12111905
Chicago/Turabian StyleCiszewski, Artur, Łukasz S. Jarosz, Arletta Bielecka, Agnieszka Marek, Bartłomiej Szymczak, Zbigniew Grądzki, and Anna Rysiak. 2023. "Effect of In Ovo Administration of a Multi-Strain Probiotic and Zinc Glycine Chelate on Antioxidant Capacity and Selected Immune Parameters in Newly Hatched Chicks" Antioxidants 12, no. 11: 1905. https://doi.org/10.3390/antiox12111905
APA StyleCiszewski, A., Jarosz, Ł. S., Bielecka, A., Marek, A., Szymczak, B., Grądzki, Z., & Rysiak, A. (2023). Effect of In Ovo Administration of a Multi-Strain Probiotic and Zinc Glycine Chelate on Antioxidant Capacity and Selected Immune Parameters in Newly Hatched Chicks. Antioxidants, 12(11), 1905. https://doi.org/10.3390/antiox12111905