
Oleuropein Attenuates Oxidative Stress in Human Trophoblast Cells

 , , , ,
, , , ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line
2.2. Oleuropein Treatment Preparations
2.3. Evaluation of Oleuropein Effects against Cytotoxicity Induced by Hydrogen Peroxide in HTR-8/SVneo Cells
2.4. Investigation of Genotoxic and Antigenotoxic Effects of Oleuropein
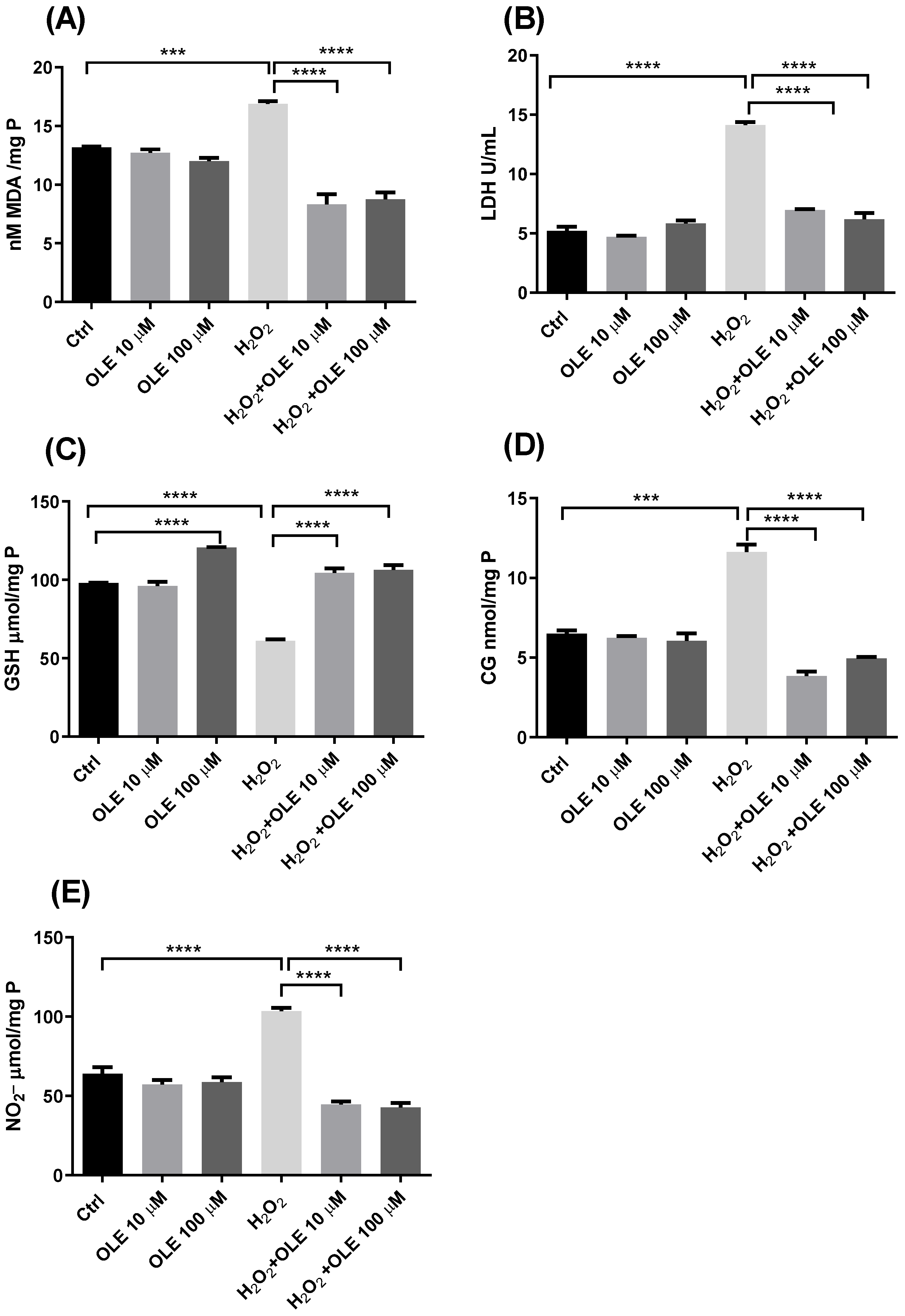
2.5. Oxidative Stress Parameter Determination
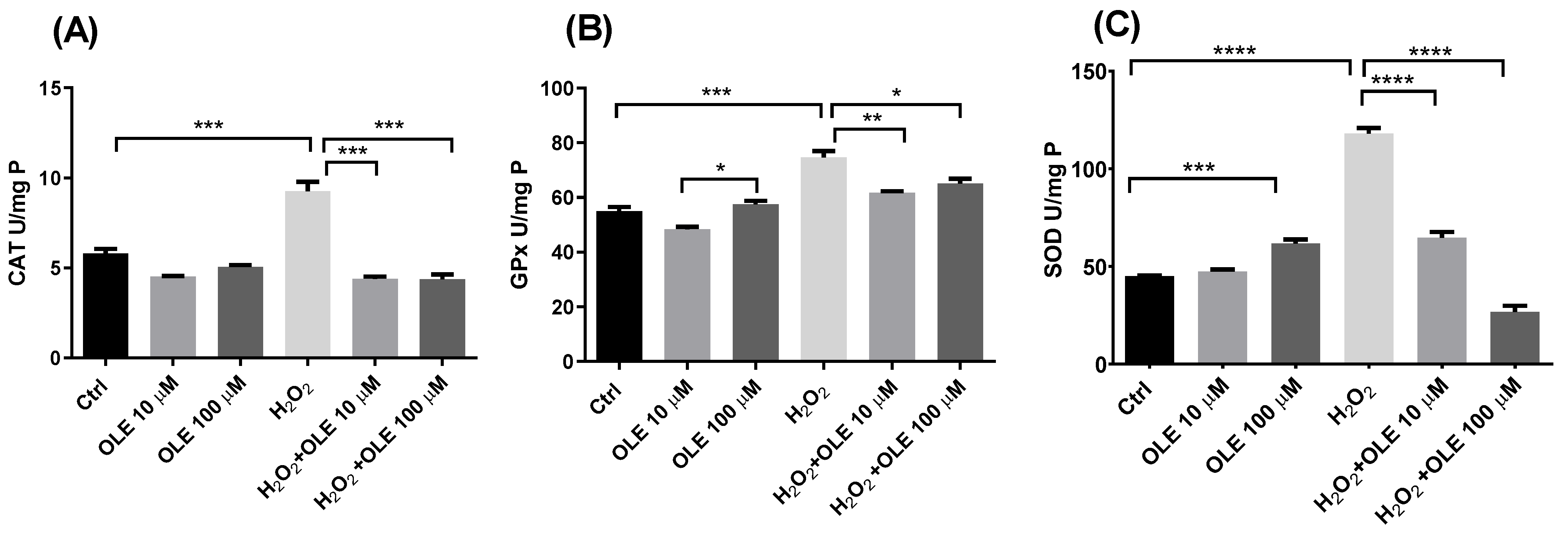
2.6. Antioxidant Enzyme Activity
2.7. Markers of Lipid Peroxidation and Oxidative Protein Damage
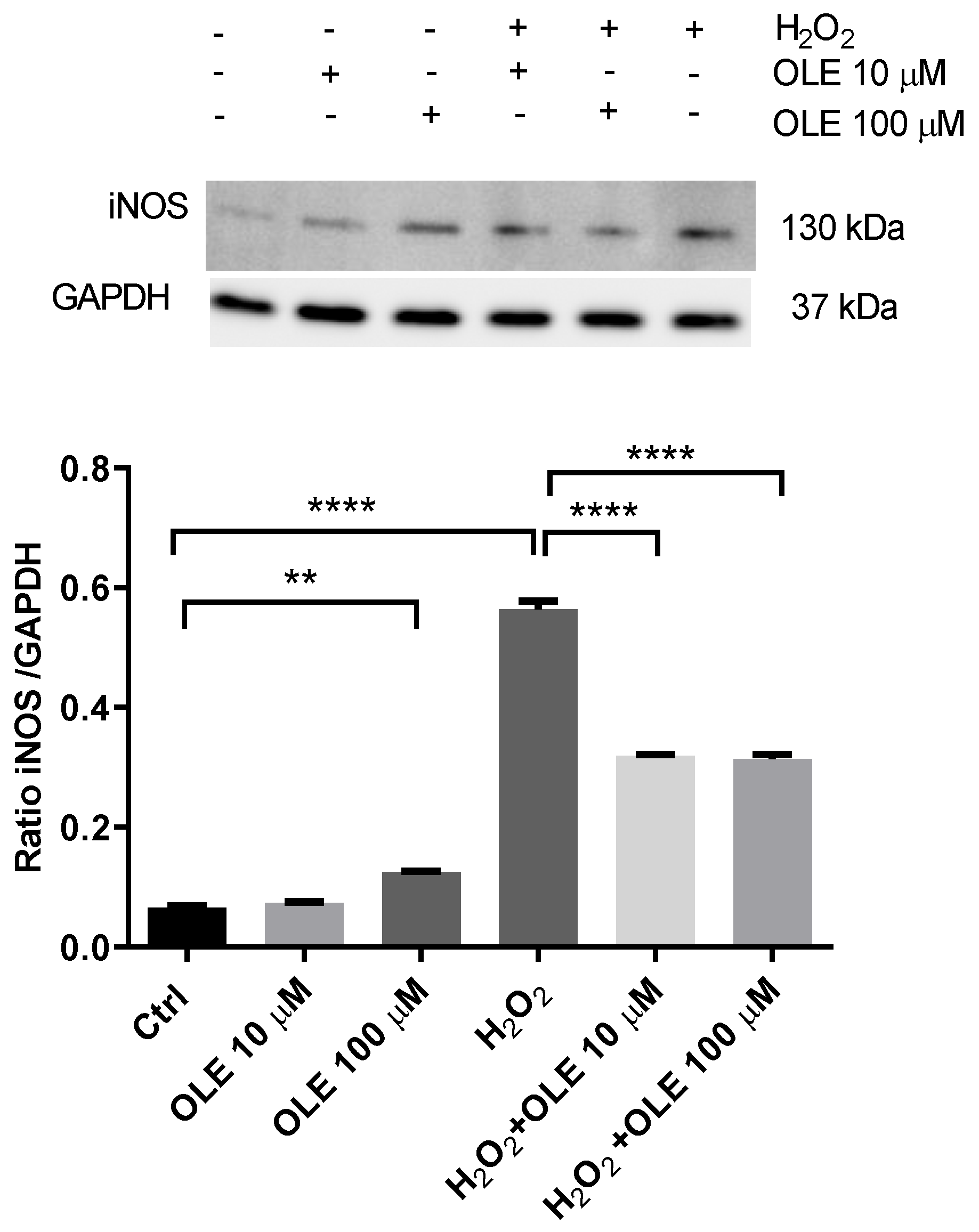
2.8. Western Blot Analysis
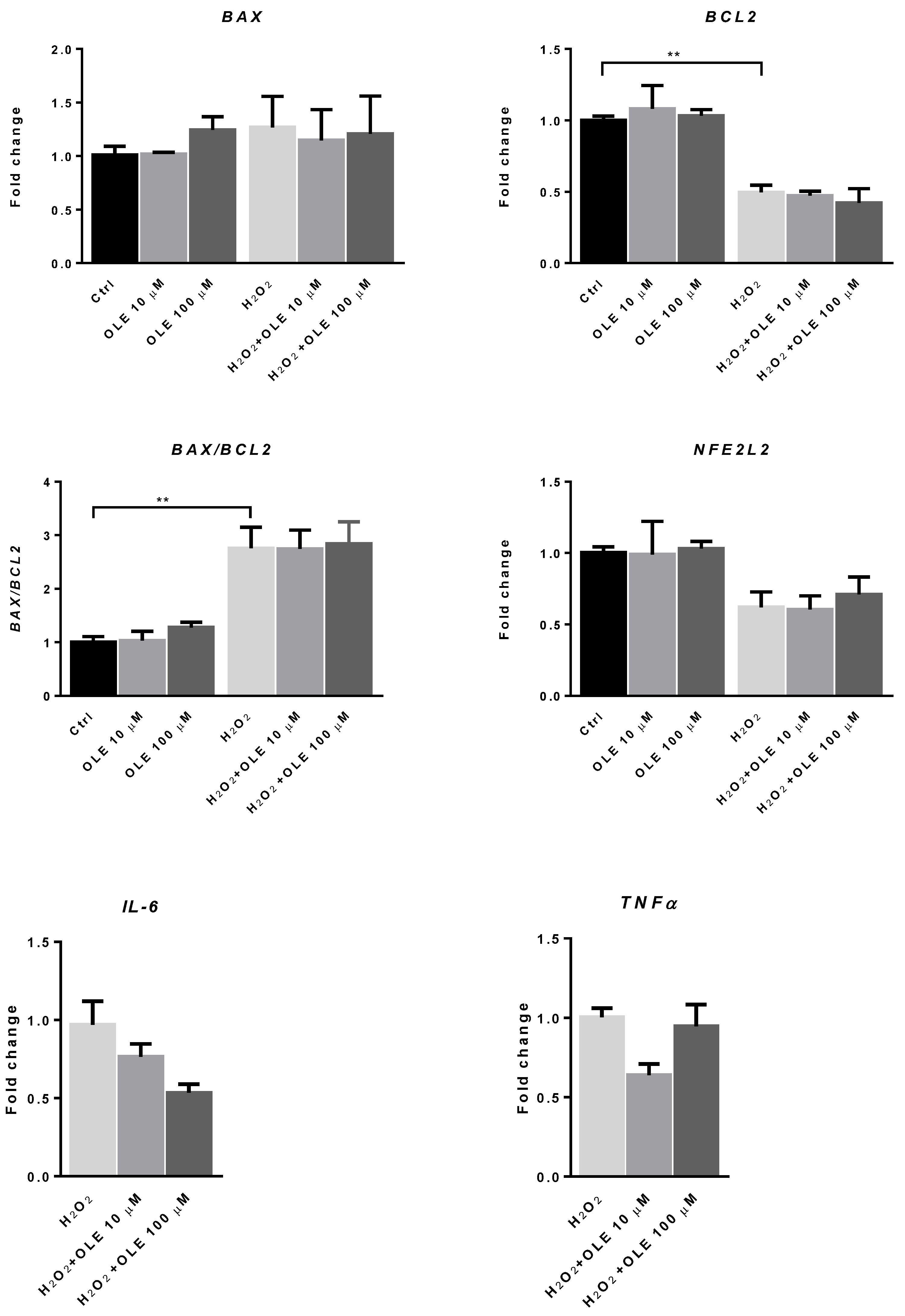
2.9. Quantitative PCR Analysis
2.10. Statistical Analysis
3. Results
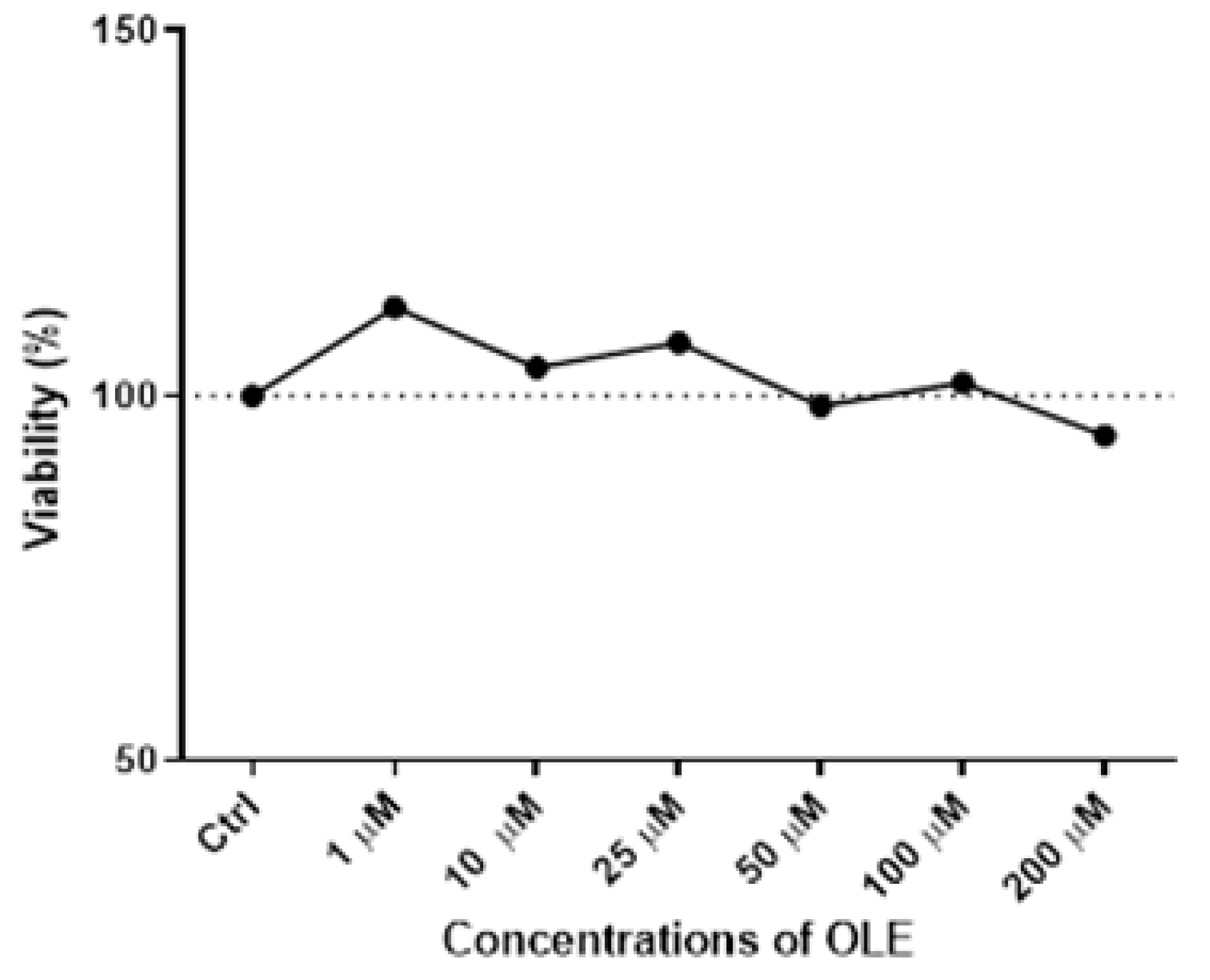
3.1. Cytotoxicity of Oleuropein in HTR-8/SVneo Cells
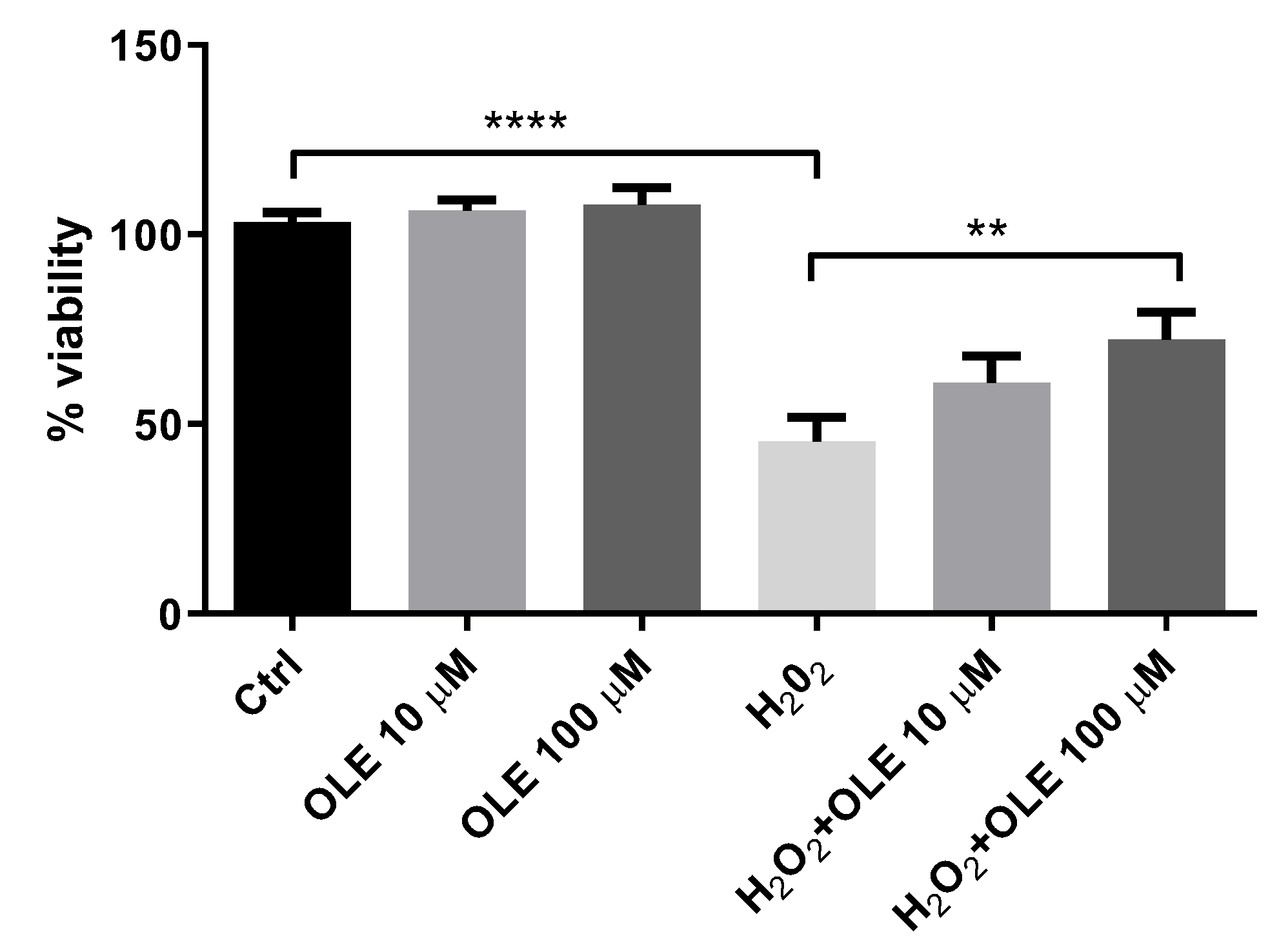
3.2. Oleuropein’s Cytoprotective Effect in H2O2-Treated HTR-8/SVneo Cells
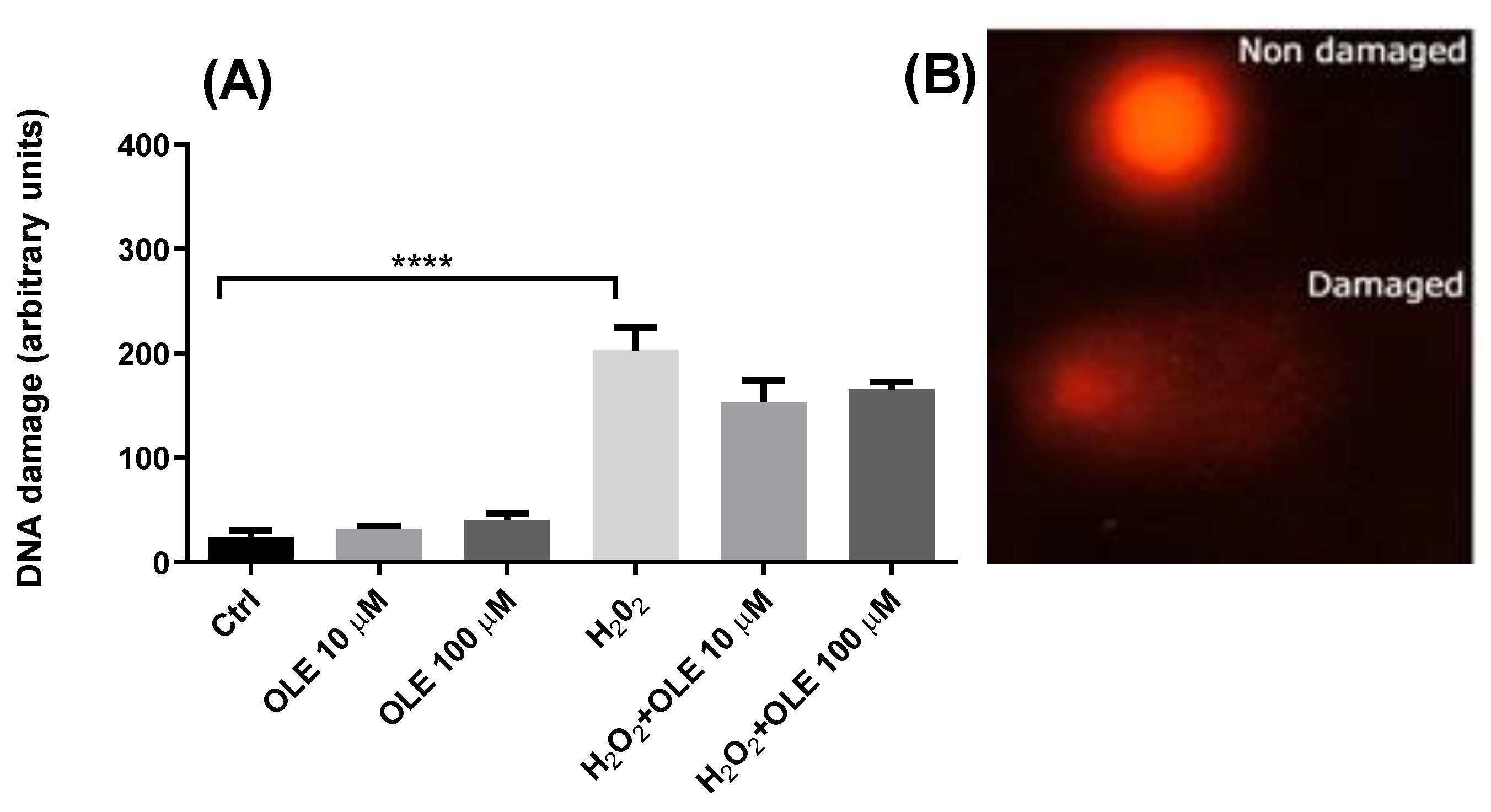
3.3. Genotoxic and Antigenotoxic Properties of Oleuropein in H2O2-Treated HTR-8/SVneo Cells
3.4. The Effects of Oleuropein on Oxidative Stress Parameters in H2O2-Treated HTR-8/SVneo Cells
3.5. The Effect of Oleuropein on Inducible Nitric Oxide Synthase (iNOS) in H2O2-Exposed HTR-8/SVneo Cells
3.6. The Effect of Oleuropein on the mRNA Expression of BAX, BCL2 and NFE2L2 in H2O2-Exposed HTR-8/SVneo Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rufino-Palomares, E.E.; Pérez-Jiménez, A.; García-Salguero, L.; Mokhtari, K.; Reyes-Zurita, F.J.; Peragón-Sánchez, J.; Lupiáñez, J.A. Nutraceutical Role of Polyphenols and Triterpenes Present in the Extracts of Fruits and Leaves of Olea Europaea as Anti-oxidants, Anti-Infectives and Anticancer Agents on Healthy Growth. Molecules 2022, 27, 2341. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Frost, B.; Liu, J. Oleuropein, Unexpected Benefits! Oncotarget 2017, 8, 17409. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, S.; Corona, G.; Yaqoob, P.; Spencer, J.P.E.; Rowland, I. Secoiridoids Delivered as Olive Leaf Extract Induce Acute Improvements in Human Vascular Function and Reduction of an Inflammatory Cytokine: A Randomised, Double-Blind, Placebo-Controlled, Cross-over Trial. Br. J. Nutr. 2015, 114, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Karković Marković, A.; Torić, J.; Barbarić, M.; Jakobušić Brala, C. Hydroxytyrosol, Tyrosol and Derivatives and Their Potential Effects on Human Health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef] [PubMed]
- Hassen, I.; Casabianca, H.; Hosni, K. Biological Activities of the Natural Anti-oxidant Oleuropein: Exceeding the Expectation—A Mini-Review. J. Funct. Foods 2015, 18, 926–940. [Google Scholar] [CrossRef]
- Czerwińska, M.; Kiss, A.K.; Naruszewicz, M. A Comparison of Anti-oxidant Activities of Oleuropein and Its Dialdehydic Derivative from Olive Oil, Oleacein. Food Chem. 2012, 131, 940–947. [Google Scholar] [CrossRef]
- Sun, W.; Wang, X.; Hou, C.; Yang, L.; Li, H.; Guo, J.; Huo, C.; Wang, M.; Miao, Y.; Liu, J.; et al. Oleuropein Improves Mitochondrial Function to Attenuate Oxidative Stress by Activating the Nrf2 Pathway in the Hypothalamic Paraventricular Nucleus of Spontaneously Hypertensive Rats. Neuropharmacology 2017, 113, 556–566. [Google Scholar] [CrossRef]
- Domitrović, R.; Jakovac, H.; Marchesi, V.V.; Šain, I.; Romić, Ž.; Rahelić, D. Preventive and Therapeutic Effects of Oleuropein against Carbon Tetrachloride-Induced Liver Damage in Mice. Pharmacol. Res. 2012, 65, 451–464. [Google Scholar] [CrossRef]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.G.A.; Yin, Y.; Yang, H.; Chughtai, M.I.; et al. The Role of Oxidative Stress and Antioxidant Balance in Pregnancy. Mediators Inflamm. 2021, 2021, 9962860. [Google Scholar] [CrossRef]
- Schoots, M.H.; Gordijn, S.J.; Scherjon, S.A.; van Goor, H.; Hillebrands, J.-L. Oxidative Stress in Placental Pathology. Placenta 2018, 69, 153–161. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, P.; Zheng, Q.; Deka, A.; Choudhury, R.; Rastogi, S. Does a MediDiet With Additional Extra Virgin Olive Oil and Pistachios Reduce the Incidence of Gestational Diabetes? Endocr. Pract. 2022, 28, 135–141. [Google Scholar] [CrossRef]
- Melero, V.; Assaf-Balut, C.; de la Torre, N.G.; Jiménez, I.; Bordiú, E.; del Valle, L.; Valerio, J.; Familiar, C.; Durán, A.; Runkle, I.; et al. Benefits of Adhering to a Mediterranean Diet Supplemented with Extra Virgin Olive Oil and Pistachios in Pregnancy on the Health of Offspring at 2 Years of Age. Results of the San Carlos Gestational Diabetes Mellitus Prevention Study. J. Clin. Med. 2020, 9, 1454. [Google Scholar] [CrossRef]
- García de la Torre, N.; Assaf-Balut, C.; Jiménez Varas, I.; del Valle, L.; Durán, A.; Fuentes, M.; del Prado, N.; Bordiú, E.; Valerio, J.J.; Herraiz, M.A.; et al. Effectiveness of Following Mediterranean Diet Recommendations in the Real World in the Incidence of Gestational Diabetes Mellitus (GDM) and Adverse Maternal-Foetal Outcomes: A Prospective, Universal, Interventional Study with a Single Group. The St Carlos. Nutrients 2019, 11, 1210. [Google Scholar] [CrossRef]
- Al Wattar, H.B.; Dodds, J.; Placzek, A.; Beresford, L.; Spyreli, E.; Moore, A.; Gonzalez Carreras, F.J.; Austin, F.; Murugesu, N.; Roseboom, T.J.; et al. Mediterranean-Style Diet in Pregnant Women with Metabolic Risk Factors (ESTEEM): A Pragmatic Multicentre Randomised Trial. PLoS Med. 2019, 16, e1002857. [Google Scholar] [CrossRef]
- Martínez-Galiano, J.; Olmedo-Requena, R.; Barrios-Rodríguez, R.; Amezcua-Prieto, C.; Bueno-Cavanillas, A.; Salcedo-Bellido, I.; Jimenez-Moleon, J.; Delgado-Rodríguez, M. Effect of Adherence to a Mediterranean Diet and Olive Oil Intake during Pregnancy on Risk of Small for Gestational Age Infants. Nutrients 2018, 10, 1234. [Google Scholar] [CrossRef]
- Minhas, A.S.; Hong, X.; Wang, G.; Rhee, D.K.; Liu, T.; Zhang, M.; Michos, E.D.; Wang, X.; Mueller, N.T. Mediterranean-Style Diet and Risk of Preeclampsia by Race in the Boston Birth Cohort. J. Am. Heart Assoc. 2022, 11, e022589. [Google Scholar] [CrossRef]
- Fournier, S.B.; D’Errico, J.N.; Stapleton, P.A. Uterine Vascular Control Preconception and During Pregnancy. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 2021; pp. 1871–1893. [Google Scholar]
- Mukherjee, I.; Dhar, R.; Singh, S.; Sharma, J.B.; Nag, T.C.; Mridha, A.R.; Jaiswal, P.; Biswas, S.; Karmakar, S. Oxidative Stress-Induced Impairment of Trophoblast Function Causes Preeclampsia through the Unfolded Protein Response Pathway. Sci. Rep. 2021, 11, 18415. [Google Scholar] [CrossRef]
- Di Fabrizio, C.; Giorgione, V.; Khalil, A.; Murdoch, C.E. Anti-oxidants in Pregnancy: Do We Really Need More Trials? Antioxidants 2022, 11, 812. [Google Scholar] [CrossRef]
- Kostić, S.; Vilotić, A.; Pirković, A.; Dekanski, D.; Borozan, S.; Nacka-Aleksić, M.; Vrzić-Petronijević, S.; Jovanović Krivokuća, M. Caffeic Acid Protects Human Trophoblast HTR-8/SVneo Cells from H2O2-Induced Oxidative Stress and Genotoxicity. Food Chem. Toxicol. 2022, 163, 112993. [Google Scholar] [CrossRef]
- Bruić, M.; Pirković, A.; Vilotić, A.; Jovanović-Krivokuća, M.; Spremo-Potparević, B. Cytoprotective and Genoprotective Effects of Taxifolin against Oxidative Damage in HTR-8/SVneo Human Trophoblast Cells. Mutagenesis 2022, geac013. [Google Scholar] [CrossRef]
- Møller, P.; Azqueta, A.; Boutet-Robinet, E.; Koppen, G.; Bonassi, S.; Milić, M.; Gajski, G.; Costa, S.; Teixeira, J.P.; Costa Pereira, C.; et al. Minimum Information for Reporting on the Comet Assay (MIRCA): Recommendations for Describing Comet Assay Procedures and Results. Nat. Protoc. 2020, 15, 3817–3826. [Google Scholar] [CrossRef]
- Collins, A.R.; Dusinská, M.; Gedik, C.M.; Stĕtina, R. Oxidative Damage to DNA: Do We Have a Reliable Biomarker? Environ. Health Perspect. 1996, 104, 465–469. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in Vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Günzler, W.A.; Kremers, H.; Flohé, L. An Improved Coupled Test Procedure for Glutathione Peroxidase (EC 1.11.1.9.) in Blood. Clin. Chem. Lab. Med. 1974, 12, 444–448. [Google Scholar] [CrossRef]
- Sun, M.; Zigman, S. An Improved Spectrophotometric Assay for Superoxide Dismutase Based on Epinephrine Autoxidation. Anal. Biochem. 1978, 90, 81–89. [Google Scholar] [CrossRef]
- Stocks, J.; Dormandy, T.L. The Autoxidation of Human Red Cell Lipids Induced by Hydrogen Peroxide. Br. J. Haematol. 1971, 20, 95–111. [Google Scholar] [CrossRef]
- Bergmayer, H.U. Methods of Enzymatic Analysis. Verlag Chemie G.m.b.H., Weinheim/Bergstr. (Germany) and Academic Press, New York and London. 1963. Starch—Stärke 1963, 15, 272. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.-G.; Ahn, B.-W.; Shaltiel, S.; Stadtman, E.R. [49] Determination of Carbonyl Content in Oxidatively Modified Proteins. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990; pp. 464–478. [Google Scholar]
- Guevara, I.; Iwanejko, J.; Dembińska-Kieć, A.; Pankiewicz, J.; Wanat, A.; Anna, P.; Gołąbek, I.; Bartuś, S.; Malczewska-Malec, M.; Szczudlik, A. Determination of Nitrite/Nitrate in Human Biological Material by the Simple Griess Reaction. Clin. Chim. Acta 1998, 274, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Ćupić Miladinović, D.; Prevendar Crnić, A.; Peković, S.; Dacić, S.; Ivanović, S.; Santibanez, J.F.; Ćupić, V.; Borozan, N.; Antonijević Miljaković, E.; Borozan, S. Recovery of Brain Cholinesterases and Effect on Parameters of Oxidative Stres and Apoptosis in Quails (Coturnix Japonica) after Chlorpyrifos and Vitamin B1 Administration. Chem. Biol. Interact. 2021, 333, 109312. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic-Krivokuca, M.; Stefanoska, I.; Rabi, A.; Vilotic, A.; Petronijevic, M.; Vrzic-Petronijevic, S.; Radojcic, L.; Vicovac, L. MIF Is among the Proinflammatory Cytokines Increased by LPS in the Human Trophoblast Line. Arch. Biol. Sci. 2016, 68, 715–722. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tenório, M.B.; Ferreira, R.C.; Moura, F.A.; Bueno, N.B.; Goulart, M.O.F.; Oliveira, A.C.M. Oral Anti-oxidant Therapy for Prevention and Treatment of Preeclampsia: Meta-Analysis of Randomized Controlled Trials. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 865–876. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.; Qiu, J.; Ni, Y.; Chai, S.; Zhou, L.; Li, J.; Yan, B.; Yang, J.; Liu, Q. The Association between Dietary Vitamin C/E and Gestational Hypertensive Disorder: A Case-Control Study. J. Nutr. Sci. Vitaminol. 2018, 64, 454–465. [Google Scholar] [CrossRef]
- Chappell, L.C.; Seed, P.T.; Kelly, F.J.; Briley, A.; Hunt, B.J.; Charnock-Jones, D.S.; Mallet, A.; Poston, L.; CStat. Vitamin C and E Supplementation in Women at Risk of Preeclampsia Is Associated with Changes in Indices of Oxidative Stress and Placental Function. Am. J. Obstet. Gynecol. 2002, 187, 777–784. [Google Scholar] [CrossRef]
- Poston, L.; Briley, A.; Seed, P.; Kelly, F.; Shennan, A. Vitamin C and Vitamin E in Pregnant Women at Risk for Pre-Eclampsia (VIP Trial): Randomised Placebo-Controlled Trial. Lancet 2006, 367, 1145–1154. [Google Scholar] [CrossRef]
- Poston, L.; Igosheva, N.; Mistry, H.D.; Seed, P.T.; Shennan, A.H.; Rana, S.; Karumanchi, S.A.; Chappell, L.C. Role of Oxidative Stress and Anti-oxidant Supplementation in Pregnancy Disorders. Am. J. Clin. Nutr. 2011, 94, 1980S–1985S. [Google Scholar] [CrossRef]
- Rumbold, A.; Duley, L.; Crowther, C.A.; Haslam, R.R. Anti-oxidants for Preventing Pre-Eclampsia. Cochrane Database Syst. Rev. 2008, 1, 1–86, Art. No.: CD004227. [Google Scholar] [CrossRef]
- Otero, D.M.; Lorini, A.; Oliveira, F.M.; da Fonseca Antunes, B.; Oliveira, R.M.; Zambiazi, R.C. Leaves of Olea europaea L. as a Source of Oleuropein: Characteristics and Biological Aspects. Res. Soc. Dev. 2021, 10, e185101321130. [Google Scholar] [CrossRef]
- Gomez Ribot, D.; Diaz, E.; Fazio, M.V.; Gómez, H.L.; Fornes, D.; Macchi, S.B.; Gresta, C.A.; Capobianco, E.; Jawerbaum, A. An Extra Virgin Olive Oil-enriched Diet Improves Maternal, Placental, and Cord Blood Parameters in GDM Pregnancies. Diabetes. Metab. Res. Rev. 2020, 36, e3349. [Google Scholar] [CrossRef]
- Parisi, F.; Rousian, M.; Huijgen, N.A.; Koning, A.H.J.; Willemsen, S.P.; de Vries, J.H.M.; Cetin, I.; Steegers, E.A.P.; Steegers-Theunissen, R.P.M. Periconceptional Maternal ‘High Fish and Olive Oil, Low Meat’ Dietary Pattern Is Associated with Increased Embryonic Growth: The Rotterdam Periconceptional Cohort (Predict) Study. Ultrasound Obstet. Gynecol. 2017, 50, 709–716. [Google Scholar] [CrossRef]
- Acevedo, N.; Frumento, P.; Harb, H.; Alashkar Alhamwe, B.; Johansson, C.; Eick, L.; Alm, J.; Renz, H.; Scheynius, A.; Potaczek, D. Histone Acetylation of Immune Regulatory Genes in Human Placenta in Association with Maternal Intake of Olive Oil and Fish Consumption. Int. J. Mol. Sci. 2019, 20, 1060. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, H.; Wang, A. Oleuropein Alleviates Gestational Diabetes Mellitus by Activating AMPK Signaling. Endocr. Connect. 2021, 10, 45–53. [Google Scholar] [CrossRef]
- Chiang, Y.-F.; Hung, H.-C.; Chen, H.-Y.; Huang, K.-C.; Lin, P.-H.; Chang, J.-Y.; Huang, T.-C.; Hsia, S.-M. The Inhibitory Effect of Extra Virgin Olive Oil and Its Active Compound Oleocanthal on Prostaglandin-Induced Uterine Hypercontraction and Pain—Ex Vivo and In Vivo Study. Nutrients 2020, 12, 3012. [Google Scholar] [CrossRef]
- Carnevale, R.; Silvestri, R.; Loffredo, L.; Novo, M.; Cammisotto, V.; Castellani, V.; Bartimoccia, S.; Nocella, C.; Violi, F. Oleuropein, a Component of Extra Virgin Olive Oil, Lowers Postprandial Glycaemia in Healthy Subjects. Br. J. Clin. Pharmacol. 2018, 84, 1566–1574. [Google Scholar] [CrossRef]
- Shi, C.; Chen, X.; Liu, Z.; Meng, R.; Zhao, X.; Liu, Z.; Guo, N. Oleuropein Protects L-02 Cells against H2O2-Induced Oxidative Stress by Increasing SOD1, GPx1 and CAT Expression. Biomed. Pharmacother. 2017, 85, 740–748. [Google Scholar] [CrossRef]
- Kucukgul, A.; Isgor, M.M.; Duzguner, V.; Atabay, M.N.; Kucukgul, A. Anti-oxidant Effects of Oleuropein on Hydrogen Peroxide-Induced Neuronal Stress- An In Vitro Study. Antiinflamm. Antiallergy. Agents Med. Chem. 2020, 19, 74–84. [Google Scholar] [CrossRef]
- Maalej, A.; Forte, M.; Bouallagui, Z.; Donato, S.; Mita, L.; Mita, D.G.; Filosa, S.; Crispi, S.; Sayadi, S. Olive Compounds Attenuate Oxidative Damage Induced in HEK-293 Cells via MAPK Signaling Pathway. J. Funct. Foods 2017, 39, 18–27. [Google Scholar] [CrossRef]
- García-Villalba, R.; Larrosa, M.; Possemiers, S.; Tomás-Barberán, F.A.; Espín, J.C. Bioavailability of Phenolics from an Oleuropein-Rich Olive (Olea Europaea) Leaf Extract and Its Acute Effect on Plasma Anti-oxidant Status: Comparison between Pre- and Postmenopausal Women. Eur. J. Nutr. 2014, 53, 1015–1027. [Google Scholar] [CrossRef]
- Harris, L.K.; McCormick, J.; Cartwright, J.E.; Whitley, G.S.; Dash, P.R. S-Nitrosylation of Proteins at the Leading Edge of Migrating Trophoblasts by Inducible Nitric Oxide Synthase Promotes Trophoblast Invasion. Exp. Cell Res. 2008, 314, 1765–1776. [Google Scholar] [CrossRef]
- Shaamash, A.H.; Elsonosy, E.D.; Zakhari, M.M.; Radwan, S.H.; El-Dien, H.M. Placental Nitric Oxide Synthase (NOS) Activity and Nitric Oxide (NO) Production in Normal Pregnancy, Pre-Eclampsia and Eclampsia. Int. J. Gynecol. Obstet. 2001, 72, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Schiessl, B.; Mylonas, I.; Hantschmann, P.; Kuhn, C.; Schulze, S.; Kunze, S.; Friese, K.; Jeschke, U. Expression of Endothelial NO Synthase, Inducible NO Synthase, and Estrogen Receptors Alpha and Beta in Placental Tissue of Normal, Preeclamptic, and Intrauterine Growth-Restricted Pregnancies. J. Histochem. Cytochem. 2005, 53, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, M.; Kim, J.-Y.; Kim, T.; Hwang, J.; Ha, K.-S.; Won, M.-H.; Ryoo, S.; Kwon, Y.-G.; Kim, Y.-M. Circulating MiRNAs Associated with Dysregulated Vascular and Trophoblast Function as Target-Based Diagnostic Biomarkers for Preeclampsia. Cells 2020, 9, 2003. [Google Scholar] [CrossRef] [PubMed]
- Žák, P.; Souček, M. Correlation of Tumor Necrosis Factor Alpha, Interleukin 6 and Interleukin 10 With Blood Pressure, Risk of Preeclampsia and Low Birth Weight in Gestational Diabetes. Physiol. Res. 2019, 68, 395–408. [Google Scholar] [CrossRef]
- Bellos, I.; Karageorgiou, V.; Kapnias, D.; Karamanli, K.-E.; Siristatidis, C. The Role of Interleukins in Preeclampsia: A Comprehensive Review. Am. J. Reprod. Immunol. 2018, 80, e13055. [Google Scholar] [CrossRef]
- Jovanovic, M.; Kovacevic, T.; Stefanoska, I.; Vicovac, L. The Effect of IL-6 on the Trophoblast Cell Line HTR-8/SVneo. Arch. Biol. Sci. 2010, 62, 531–538. [Google Scholar] [CrossRef]
- Haider, S.; Knöfler, M. Human Tumour Necrosis Factor: Physiological and Pathological Roles in Placenta and Endometrium. Placenta 2009, 30, 111–123. [Google Scholar] [CrossRef]
- Fakhr, Y.; Koshti, S.; Habibyan, Y.B.; Webster, K.; Hemmings, D.G. Tumor Necrosis Factor-α Induces a Preeclamptic-like Phenotype in Placental Villi via Sphingosine Kinase 1 Activation. Int. J. Mol. Sci. 2022, 23, 3750. [Google Scholar] [CrossRef]
- Vilotić, A.; Nacka-Aleksić, M.; Pirković, A.; Bojić-Trbojević, Ž.; Dekanski, D.; Jovanović Krivokuća, M. IL-6 and IL-8: An Overview of Their Roles in Healthy and Pathological Pregnancies. Int. J. Mol. Sci. 2022, 23, 14574. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Y.; Alexander, J.S.; Lewis, D.F. Preeclampsia Status Controls Interleukin-6 and Soluble IL-6 Receptor Release from Neutrophils and Endothelial Cells: Relevance to Increased Inflammatory Responses. Pathophysiology 2021, 28, 202–211. [Google Scholar] [CrossRef]
- Romanowska-Próchnicka, K.; Felis-Giemza, A.; Olesińska, M.; Wojdasiewicz, P.; Paradowska-Gorycka, A.; Szukiewicz, D. The Role of TNF-α and Anti-TNF-α Agents during Preconception, Pregnancy, and Breastfeeding. Int. J. Mol. Sci. 2021, 22, 2922. [Google Scholar] [CrossRef]
- Socha, M.W.; Flis, W.; Wartęga, M.; Stankiewicz, M. Impact of Oxidative Stress on Molecular Mechanisms of Cervical Ripening in Pregnant Women. Int. J. Mol. Sci. 2022, 23, 12780. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 System in Development, Oxidative Stress Response and Diseases: An Evolutionarily Conserved Mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Chigusa, Y.; Kawasaki, K.; Kondoh, E.; Mogami, H.; Ujita, M.; Fujita, K.; Tatsumi, K.; Takeda, S.; Konishi, I. Simvastatin Inhibits Oxidative Stress via the Activation of Nuclear Factor Erythroid 2-Related Factor 2 Signaling in Trophoblast Cells. J. Obstet. Gynaecol. Res. 2016, 42, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Chigusa, Y.; Tatsumi, K.; Kondoh, E.; Fujita, K.; Nishimura, F.; Mogami, H.; Konishi, I. Decreased Lectin-Like Oxidized LDL Receptor 1 (LOX-1) and Low Nrf2 Activation in Placenta Are Involved in Preeclampsia. J. Clin. Endocrinol. Metab. 2012, 97, E1862–E1870. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Wu, J.; Li, M.; Wang, L.; Zhu, X.; Chen, Y. Impaction of Factors Associated with Oxidative Stress on the Pathogenesis of Gestational Hypertension and Preeclampsia. Medicine 2021, 100, e23666. [Google Scholar] [CrossRef]
- ALHaithloul, H.A.S.; Alotaibi, M.F.; Bin-Jumah, M.; Elgebaly, H.; Mahmoud, A.M. Olea Europaea Leaf Extract Up-Regulates Nrf2/ARE/HO-1 Signaling and Attenuates Cyclophosphamide-Induced Oxidative Stress, Inflammation and Apoptosis in Rat Kidney. Biomed. Pharmacother. 2019, 111, 676–685. [Google Scholar] [CrossRef]
- Ji, S.T.; Kim, Y.-J.; Jung, S.Y.; Kim, D.Y.; Kang, S.; Park, J.H.; Jang, W.B.; Ha, J.; Yun, J.; Kwon, S.-M. Oleuropein Attenuates Hydrogen Peroxide-Induced Autophagic Cell Death in Human Adipose-Derived Stem Cells. Biochem. Biophys. Res. Commun. 2018, 499, 675–680. [Google Scholar] [CrossRef]
- Castejón, M.L.; Rosillo, M.Á.; Montoya, T.; González-Benjumea, A.; Fernández-Bolaños, J.M.; Alarcón-de-la-Lastra, C. Oleuropein Down-Regulated IL-1β-Induced Inflammation and Oxidative Stress in Human Synovial Fibroblast Cell Line SW982. Food Funct. 2017, 8, 1890–1898. [Google Scholar] [CrossRef]
- Fröhlich, J.D.; Huppertz, B.; Abuja, P.M.; König, J.; Desoye, G. Oxygen Modulates the Response of First-Trimester Trophoblasts to Hyperglycemia. Am. J. Pathol. 2012, 180, 153–164. [Google Scholar] [CrossRef]
- Park, W. MAPK Inhibitors, Particularly the JNK Inhibitor, Increase Cell Death Effects in H2O2-Treated Lung Cancer Cells via Increased Superoxide Anion and Glutathione Depletion. Oncol. Rep. 2018, 39, 860–870. [Google Scholar] [CrossRef]
- Tang, D.; Wu, D.; Hirao, A.; Lahti, J.M.; Liu, L.; Mazza, B.; Kidd, V.J.; Mak, T.W.; Ingram, A.J. ERK Activation Mediates Cell Cycle Arrest and Apoptosis after DNA Damage Independently of P53. J. Biol. Chem. 2002, 277, 12710–12717. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lin, Y.; Huang, X.; Xu, C.; Liu, X.; Wang, L.; Yu, M.; Li, D.; Zhu, Y.; Du, M. SCM-198 Protects Endometrial Stromal Cells from Oxidative Damage through Bax/Bcl-2 and ERK Signaling Pathways. Acta Biochim. Biophys. Sin. 2019, 51, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.-M.; Gipp, J.J.; Mulcahy, R.T.; Jones, D.P. H2O2-Dependent Activation of GCLC-ARE4 Reporter Occurs by Mitogen-Activated Protein Kinase Pathways without Oxidation of Cellular Glutathione or Thioredoxin-1. J. Biol. Chem. 2004, 279, 5837–5845. [Google Scholar] [CrossRef]
- Du, Y.; Cai, Z.; Zhang, H.; Liang, W.; Wang, H.; Man, Q.; Wang, W. Nitric Oxide Mediates Disruption of Human Placental Trophoblast Invasion Induced by Perfluorobutane Sulfonate. Environ. Pollut. 2021, 283, 117137. [Google Scholar] [CrossRef]
- Salakou, S.; Kardamakis, D.; Tsamandas, A.C.; Zolota, V.; Apostolakis, E.; Tzelepi, V.; Papathanasopoulos, P.; Bonikos, D.S.; Papapetropoulos, T.; Petsas, T.; et al. Increased Bax/Bcl-2 ratio up-regulates caspase-3 and increases apoptosis in the thymus of patients with myasthenia gravis. Vivo 2007, 21, 123–132. [Google Scholar] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pirković, A.; Vilotić, A.; Borozan, S.; Nacka-Aleksić, M.; Bojić-Trbojević, Ž.; Krivokuća, M.J.; Battino, M.; Giampieri, F.; Dekanski, D. Oleuropein Attenuates Oxidative Stress in Human Trophoblast Cells. Antioxidants 2023, 12, 197. https://doi.org/10.3390/antiox12010197
Pirković A, Vilotić A, Borozan S, Nacka-Aleksić M, Bojić-Trbojević Ž, Krivokuća MJ, Battino M, Giampieri F, Dekanski D. Oleuropein Attenuates Oxidative Stress in Human Trophoblast Cells. Antioxidants. 2023; 12(1):197. https://doi.org/10.3390/antiox12010197
Chicago/Turabian StylePirković, Andrea, Aleksandra Vilotić, Sunčica Borozan, Mirjana Nacka-Aleksić, Žanka Bojić-Trbojević, Milica Jovanović Krivokuća, Maurizio Battino, Francesca Giampieri, and Dragana Dekanski. 2023. "Oleuropein Attenuates Oxidative Stress in Human Trophoblast Cells" Antioxidants 12, no. 1: 197. https://doi.org/10.3390/antiox12010197
APA StylePirković, A., Vilotić, A., Borozan, S., Nacka-Aleksić, M., Bojić-Trbojević, Ž., Krivokuća, M. J., Battino, M., Giampieri, F., & Dekanski, D. (2023). Oleuropein Attenuates Oxidative Stress in Human Trophoblast Cells. Antioxidants, 12(1), 197. https://doi.org/10.3390/antiox12010197