


Effects of Low Temperature on Antioxidant and Heat Shock Protein Expression Profiles and Transcriptomic Responses in Crayfish (Cherax destructor)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Organisms
2.2. Experimental Procedures
2.3. Antioxidant Index Detection
2.4. Detection of Anti-Stress-Gene Expression
2.5. Transcriptomic Analysis
2.5.1. Transcriptome Sequencing
2.5.2. RNA-Seq Read Processing and Mapping
2.5.3. Differentially Expressed Genes (DEGs) and Functional Enrichment
2.6. Transcriptome Data Validation
2.7. Statistical Analysis
3. Results
3.1. Effects of Temperature on the Growth Indicators of C. destructor
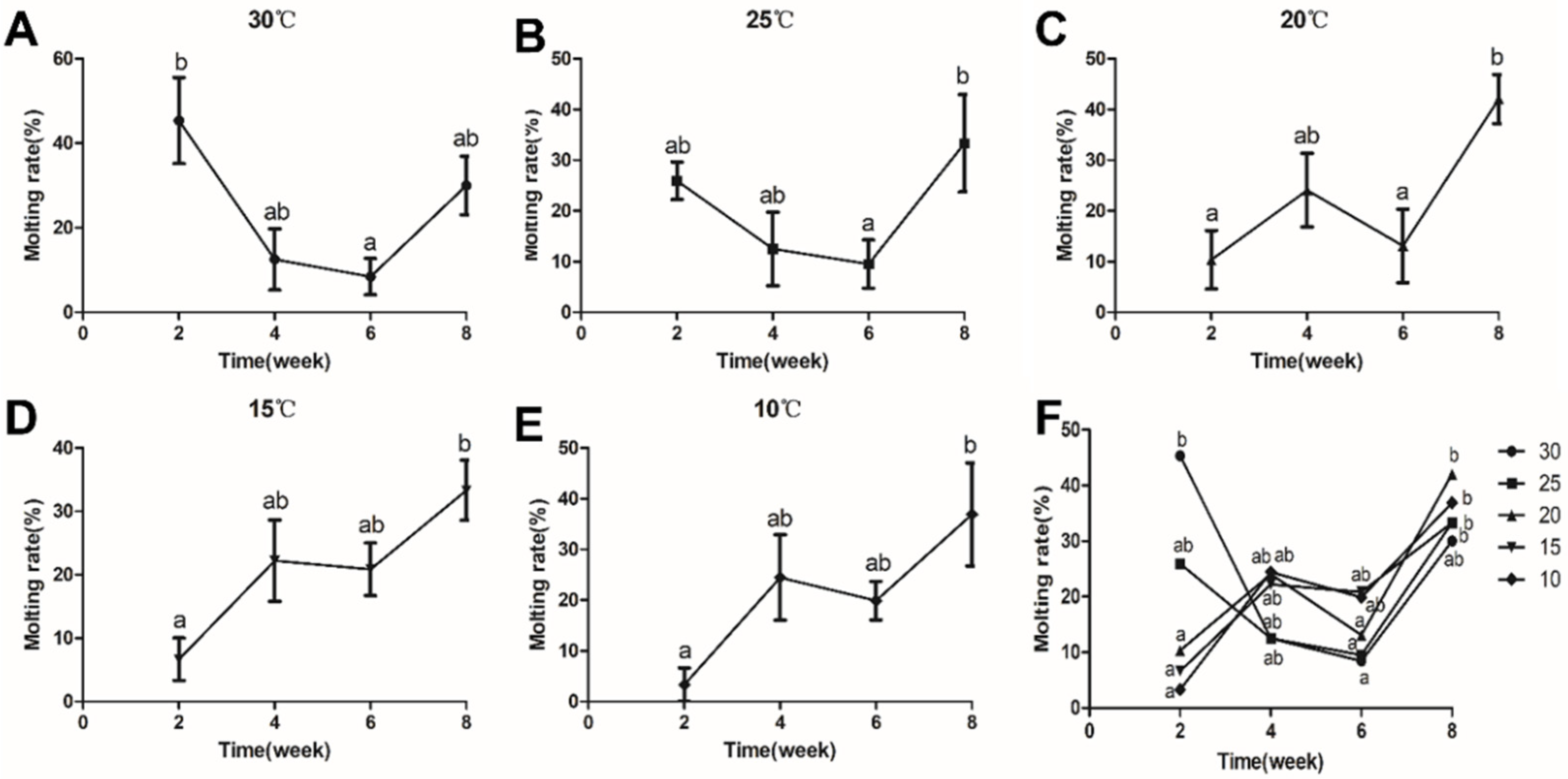
3.2. Effects of Temperature on the Molting Rate of C. destructor
3.3. Effects of Temperature on Antioxidant Indicators of C. destructor
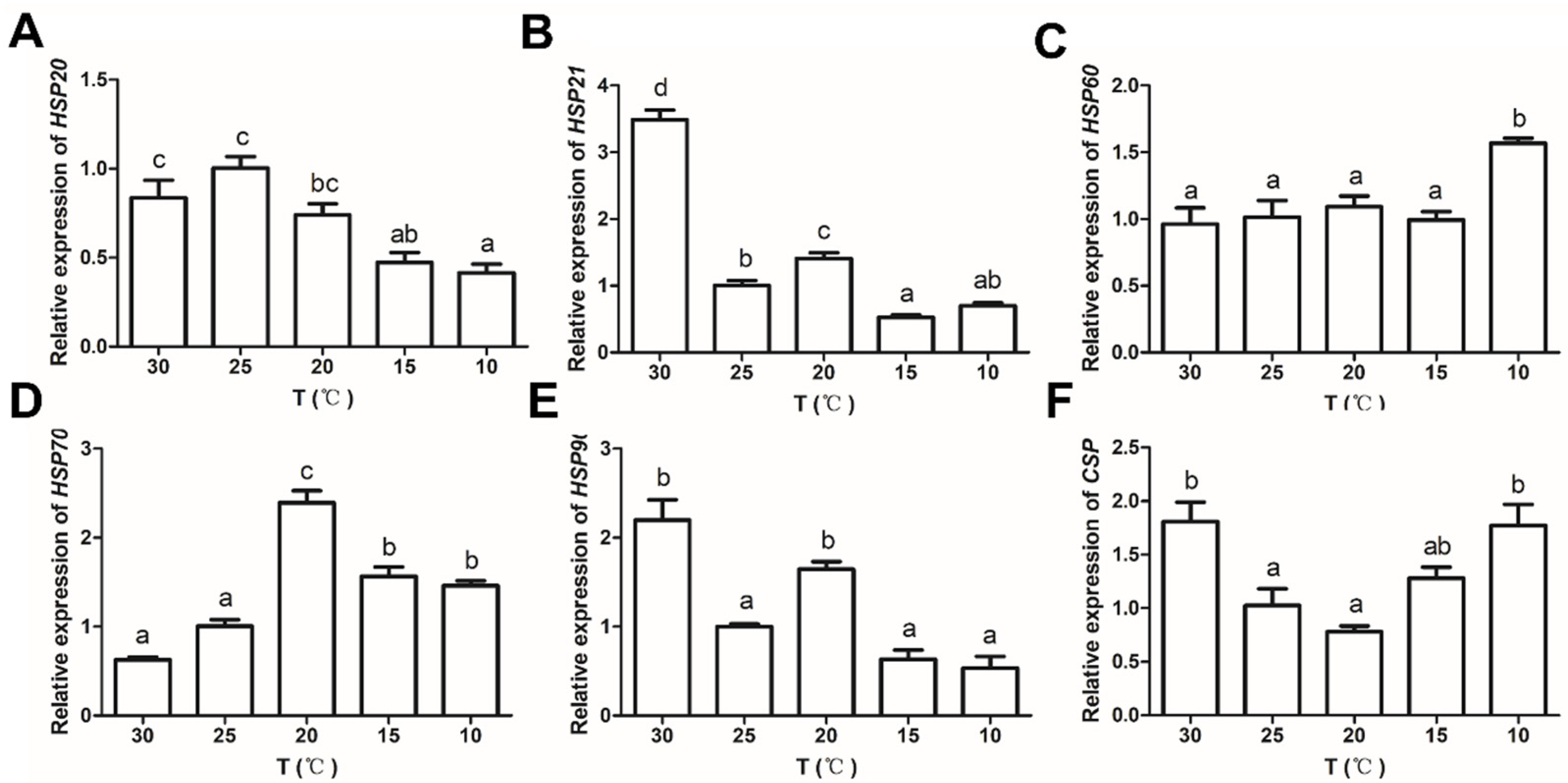
3.4. Effects of Temperature on the Expression of Anti-Stress Genes in C. destructor
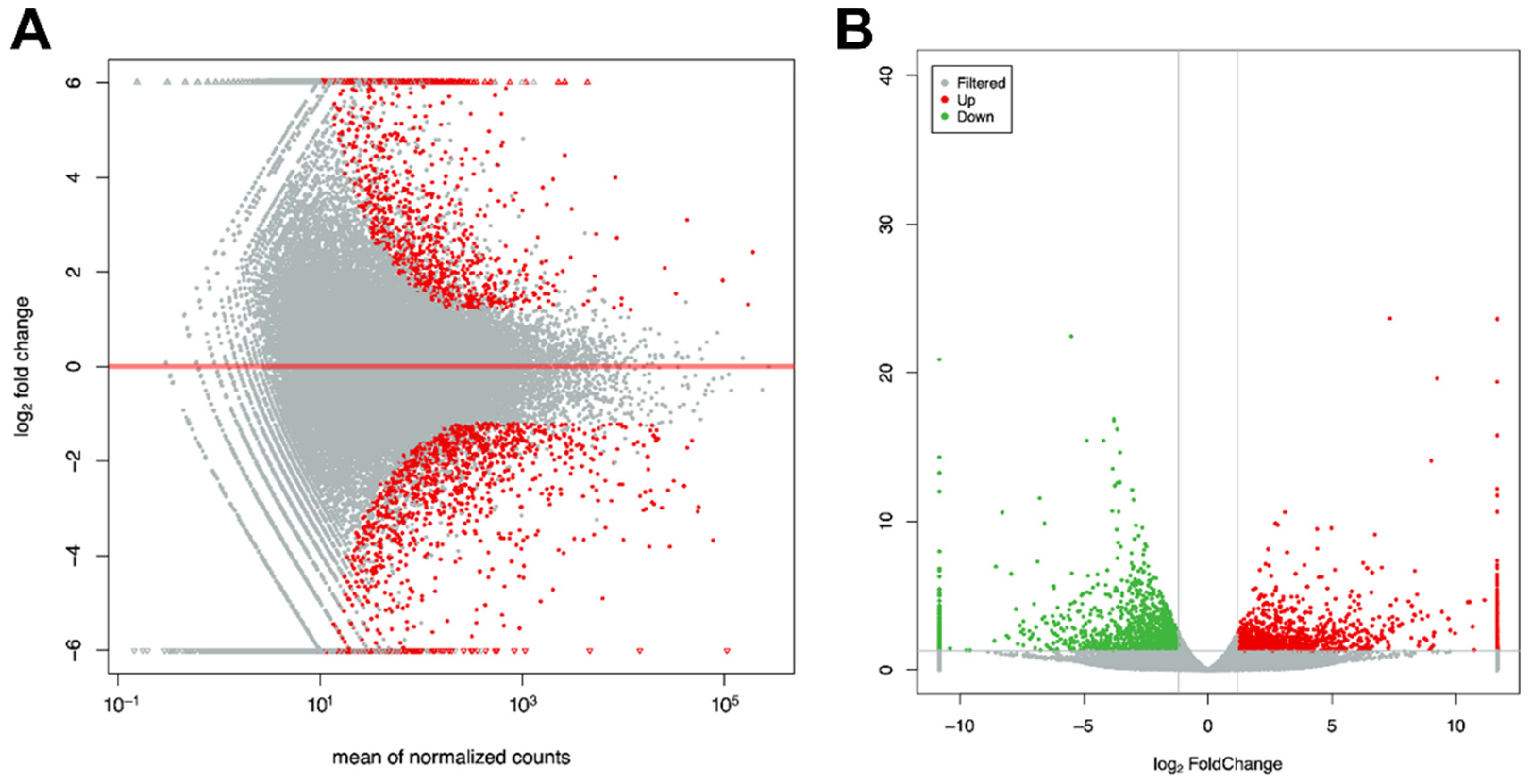
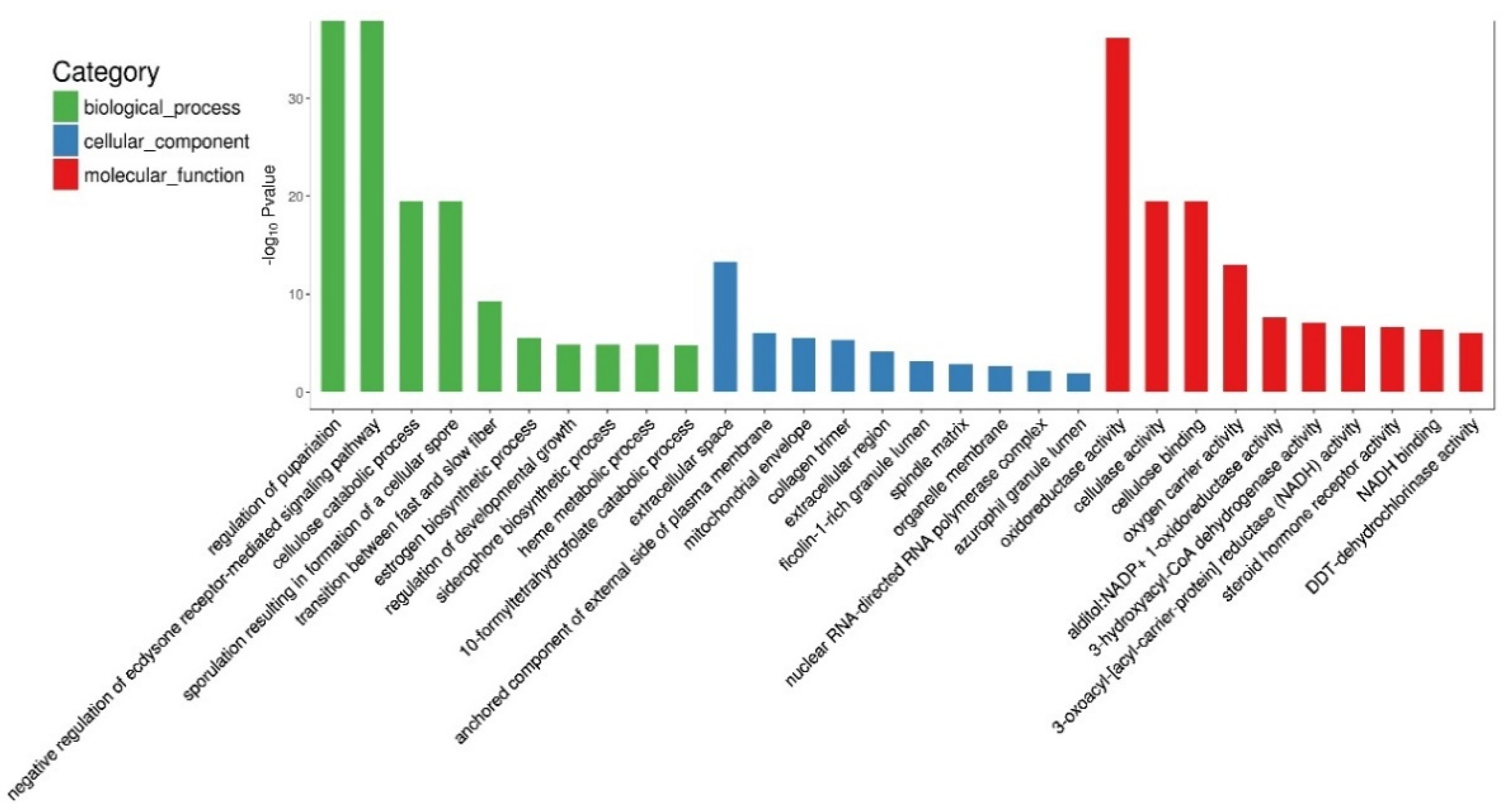
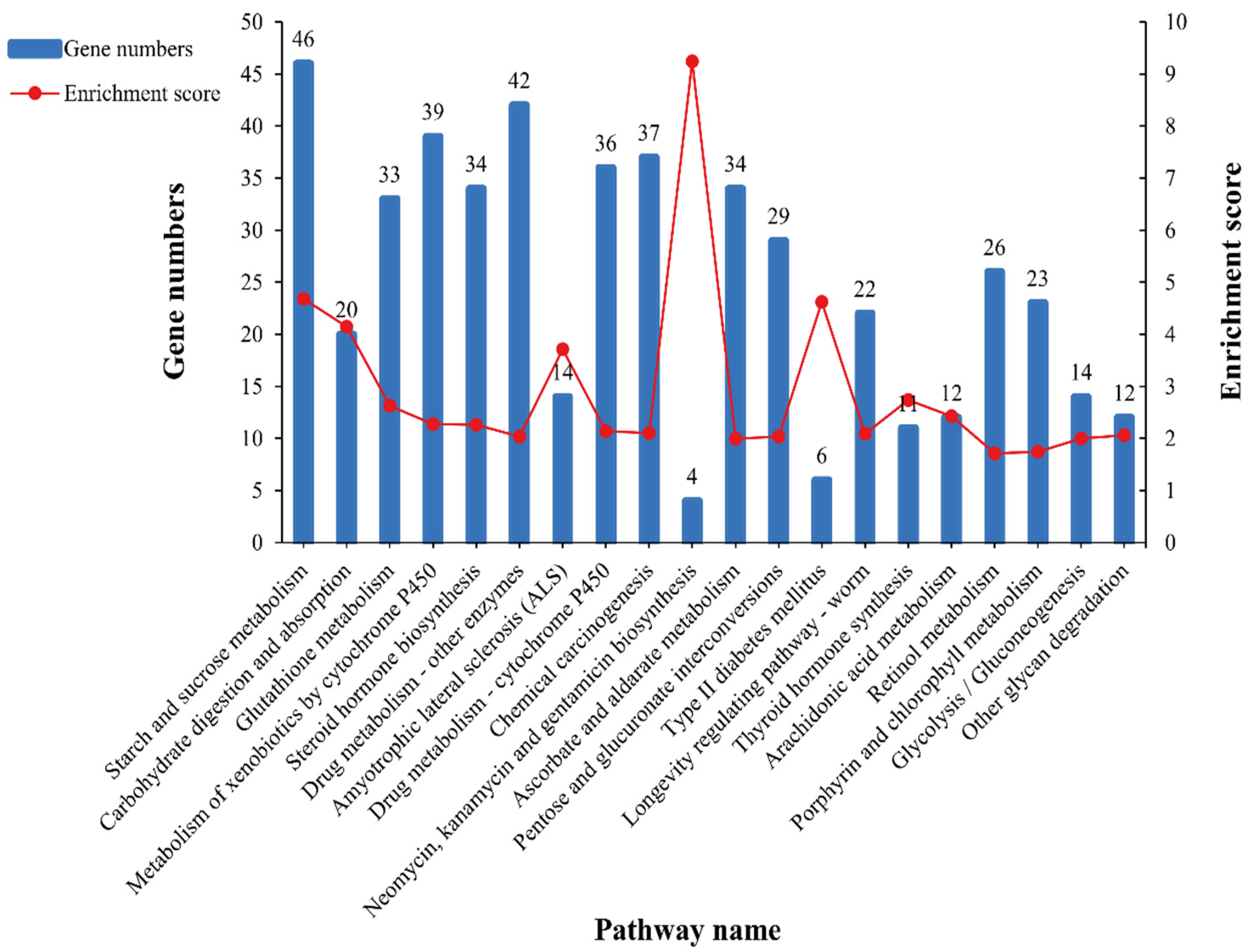
3.5. GO and KEGG Pathway Enrichment Analysis
3.6. Data Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, D.; Wu, J.; Sun, L.; Qin, X.; Fan, X.; Zheng, X. Combined stress of acute cold exposure and waterless duration at low temperature induces mortality of shrimp Litopenaeus vannamei through injuring antioxidative and immunological response in hepatopancreas tissue. J. Therm. Biol. 2021, 100, 103080. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Wang, Q.; Shao, H.; Xu, Y.; Liu, P.; Li, J. Effects of Low Temperature on Shrimp and Crab Physiology, Behavior, and Growth: A Review. Front. Mar. Sci. 2021, 8, 746177. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, W.-N.; Wang, L.-j.; Liu, Y.-F.; Wang, A.-L. Oxidative stress, DNA damage and osmolality in the Pacific white shrimp, Litopenaeus vannamei exposed to acute low temperature stress. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 154, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Cárdenas, P.; Cruz-Moreno, D.G.; González-Ruiz, R.; Peregrino-Uriarte, A.B.; Leyva-Carrillo, L.; Camacho-Jiménez, L.; Quintero-Reyes, I.; Yepiz-Plascencia, G. Combined hypoxia and high temperature affect differentially the response of antioxidant enzymes, glutathione and hydrogen peroxide in the white shrimp Litopenaeus vannamei. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 254, 110909. [Google Scholar] [CrossRef]
- Wang, Z.; Qu, Y.; Yan, M.; Li, J.; Zou, J.; Fan, L. Physiological responses of Pacific white shrimp Litopenaeus vannamei to temperature fluctuation in low-salinity water. Front. Physiol. 2019, 10, 1025. [Google Scholar] [CrossRef]
- Wu, D.-L.; Huang, Y.-H.; Liu, Z.-Q.; Yu, P.; Gu, P.-H.; Fan, B.; Zhao, Y.-L. Molecular cloning, tissue expression and regulation of nutrition and temperature on Δ6 fatty acyl desaturase-like gene in the red claw crayfish (Cherax quadricarinatus). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 225, 58–66. [Google Scholar] [CrossRef]
- Wu, D.-L.; Rao, Q.-X.; Cheng, L.; Lv, W.-W.; Zhao, Y.-L.; Song, W.-G. Cloning and characterisation of a Δ9 fatty acyl desaturase-like gene from the red claw crayfish (Cherax quadricarinatus) and its expression analysis under cold stress. J. Therm. Biol. 2021, 102, 103122. [Google Scholar] [CrossRef]
- Sun, Z.; Tan, X.; Liu, Q.; Ye, H.; Zou, C.; Xu, M.; Zhang, Y.; Ye, C. Physiological, immune responses and liver lipid metabolism of orange-spotted grouper (Epinephelus coioides) under cold stress. Aquaculture 2019, 498, 545–555. [Google Scholar] [CrossRef]
- Chen, L.; Gómez, R.; Weiss, L.C. Distinct gene expression patterns of two heat shock protein 70 members during development, diapause, and temperature stress in the freshwater crustacean Daphnia magna. Front. Cell Dev. Biol. 2021, 9, 1531. [Google Scholar] [CrossRef]
- Meng, X.-l.; Liu, P.; Li, J.; Gao, B.-Q.; Chen, P. Physiological responses of swimming crab Portunus trituberculatus under cold acclimation: Antioxidant defense and heat shock proteins. Aquaculture 2014, 434, 11–17. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, H.J.; Kim, D.H.; Oh, C.W.; Lee, J.S.; Kang, J.C. Changes in hematological parameters and heat shock proteins in juvenile sablefish depending on water temperature stress. J. Aquat. Anim. Health 2019, 31, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, L.; Xin, Y.; Wang, W.-N.; He, W.-Y.; Wang, A.-L.; Liu, Y. Effect of temperature on antioxidant enzyme gene expression and stress protein response in white shrimp, Litopenaeus vannamei. J. Therm. Biol. 2010, 35, 284–289. [Google Scholar] [CrossRef]
- Ju-Ngam, T.; McMillan, N.; Yoshimizu, M.; Kasai, H.; Wongpanya, R.; Srisapoome, P. Functional and Stress Response Analysis of Heat Shock Proteins 40 and 90 of Giant River Prawn (Macrobrachium rosenbergii) under Temperature and Pathogenic Bacterial Exposure Stimuli. Biomolecules 2021, 11, 1034. [Google Scholar] [CrossRef] [PubMed]
- Padmini, E.; Rani, M.U. Impact of seasonal variation on HSP70 expression quantitated in stressed fish hepatocytes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Pelham, H.R. Speculations on the functions of the major heat shock and glucose-regulated proteins. Cell 1986, 46, 959–961. [Google Scholar] [CrossRef]
- de Souza, D.M.; Borges, V.D.; Furtado, P.; Romano, L.A.; Wasielesky, W., Jr.; Monserrat, J.M.; de Oliveira Garcia, L. Antioxidant enzyme activities and immunological system analysis of Litopenaeus vannamei reared in biofloc technology (BFT) at different water temperatures. Aquaculture 2016, 451, 436–443. [Google Scholar] [CrossRef]
- Ren, X.; Yu, Z.; Xu, Y.; Zhang, Y.; Mu, C.; Liu, P.; Li, J. Integrated transcriptomic and metabolomic responses in the hepatopancreas of kuruma shrimp (Marsupenaeus japonicus) under cold stress. Ecotoxicol. Environ. Saf. 2020, 206, 111360. [Google Scholar] [CrossRef]
- Wu, D.-L.; Liu, Z.-Q.; Huang, Y.-H.; Lv, W.-W.; Chen, M.-H.; Li, Y.-M.; Zhao, Y.-L. Effects of cold acclimation on the survival, feeding rate, and non-specific immune responses of the freshwater red claw crayfish (Cherax quadricarinatus). Aquac. Int. 2018, 26, 557–567. [Google Scholar] [CrossRef]
- Wu, D.; Huang, Y.; Chen, Q.; Jiang, Q.; Li, Y.; Zhao, Y. Effects and transcriptional responses in the hepatopancreas of red claw crayfish Cherax quadricarinatus under cold stress. J. Therm. Biol. 2019, 85, 102404. [Google Scholar] [CrossRef]
- McCormack, R.B. New records and review of the translocation of the yabby Cherax destructor into eastern drainages of New South Wales, Australia. Aust. Zool. 2014, 37, 85–94. [Google Scholar] [CrossRef]
- Mauro, M.; Arizza, V.; Arculeo, M.; Attanzio, A.; Pinto, P.; Chirco, P.; Badalamenti, G.; Tesoriere, L.; Vazzana, M. Haemolymphatic Parameters in Two Aquaculture Crustacean Species Cherax destructor (Clark, 1836) and Cherax quadricarinatus (Von Martens, 1868). Animals 2022, 12, 543. [Google Scholar] [CrossRef]
- Mills, B.; Geddes, M. Salinty tolerance and osmoregulation of the Australian freshwater crayfish Cherax destructor Clark (Decapoda: Parastacidae). Mar. Freshw. Res. 1980, 31, 667–676. [Google Scholar] [CrossRef]
- Morris, S.; Callaghan, J. The emersion response of the Australian Yabby Cherax destructor to environmental hypoxia and the respiratory and metabolic responses to consequent air-breathing. J. Comp. Physiol. B 1998, 168, 389–398. [Google Scholar] [CrossRef]
- Ellis, B.; Morris, S. Effects of extreme pH on the physiology of the Australian’yabby’Cherax destructor: Acute and chronic changes in haemolymph oxygen levels, oxygen consumption and metabolic levels. J. Exp. Biol. 1995, 198, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, E.; Li, T.; Xu, C.; Wang, X.; Lin, H.; Qin, J.G.; Chen, L. Transcriptome and molecular pathway analysis of the hepatopancreas in the Pacific white shrimp Litopenaeus vannamei under chronic low-salinity stress. PLoS ONE 2015, 10, e0131503. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, F.; Huang, J.; Yang, L.; Jiang, S.; Yang, Q.; He, J.; Jiang, S. Transcriptome reveals involvement of immune defense, oxidative imbalance, and apoptosis in ammonia-stress response of the black tiger shrimp (Penaeus monodon). Fish Shellfish Immunol. 2018, 83, 162–170. [Google Scholar] [CrossRef]
- Zhong, S.; Mao, Y.; Wang, J.; Liu, M.; Zhang, M.; Su, Y. Transcriptome analysis of Kuruma shrimp (Marsupenaeus japonicus) hepatopancreas in response to white spot syndrome virus (WSSV) under experimental infection. Fish Shellfish Immunol. 2017, 70, 710–719. [Google Scholar] [CrossRef]
- Huang, W.; Li, H.; Cheng, C.; Ren, C.; Chen, T.; Jiang, X.; Cheng, K.; Luo, P.; Hu, C. Analysis of the transcriptome data in Litopenaeus vannamei reveals the immune basis and predicts the hub regulation-genes in response to high-pH stress. PLoS ONE 2018, 13, e0207771. [Google Scholar] [CrossRef]
- Lou, F.; Gao, T.; Han, Z. Transcriptome analyses reveal alterations in muscle metabolism, immune responses and reproductive behavior of Japanese mantis shrimp (Oratosquilla oratoria) at different cold temperature. Comp. Biochem. Physiol. D-Genom. Proteom. 2019, 32, 100615. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Jin, Y.; Ji, G.; Zhang, X. Significant genes in response to low temperature in Penaeus chinensis screened from multiple groups of transcriptome comparison. J. Therm. Biol. 2022, 107, 103198. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-N.; Wang, A.-L.; Liu, Y.; Xiu, J.; Liu, Z.-B.; Sun, R.-Y. Effects of temperature on growth, adenosine phosphates, ATPase and cellular defense response of juvenile shrimp Macrobrachium nipponense. Aquaculture 2006, 256, 624–630. [Google Scholar] [CrossRef]
- García-Guerrero, M.; Hernández-Sandoval, P.; Orduña-Rojas, J.; Cortés-Jacinto, E. Effect of temperature on weight increase, survival, and thermal preference of juvenile redclaw crayfish Cherax quadricarinatus. Hidrobiológica 2013, 23, 73–81. [Google Scholar]
- Nagasawa, H. The crustacean cuticle: Structure, composition and mineralization. Front. Biosci. 2012, 4, 711–720. [Google Scholar] [CrossRef]
- Gong, J.; Yu, K.; Shu, L.; Ye, H.; Li, S.; Zeng, C. Evaluating the effects of temperature, salinity, starvation and autotomy on molting success, molting interval and expression of ecdysone receptor in early juvenile mud crabs, Scylla paramamosain. J. Exp. Mar. Biol. Ecol. 2015, 464, 11–17. [Google Scholar] [CrossRef]
- Travis, D.F. The molting cycle of the spiny lobster, Panulirus argus Latreille. I. Molting and growth in laboratory-maintained individuals. Biol. Bull. 1954, 107, 433–450. [Google Scholar] [CrossRef]
- Hsieh, S.; Chen, S.; Yang, Y.; Kuo, C. Involvement of norepinephrine in the hyperglycemic responses of the freshwater giant prawn, Macrobrachium rosenbergii, under cold shock. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 143, 254–263. [Google Scholar] [CrossRef]
- Pan, L.-Q.; Hu, F.-W.; Jing, F.-T.; Liu, H.-J. The effect of different acclimation temperatures on the prophenoloxidase system and other defence parameters in Litopenaeus vannamei. Fish Shellfish Immunol. 2008, 25, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Subramoniam, T. Crustacean ecdysteriods in reproduction and embryogenesis. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 2000, 125, 135–156. [Google Scholar] [CrossRef]
- Nelson, E.R.; Habibi, H.R. Estrogen receptor function and regulation in fish and other vertebrates. Gen. Comp. Endocrinol. 2013, 192, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Huang, H.; Li, S.; Wang, G. Immunorecognition of estrogen and androgen receptors in the brain and thoracic ganglion mass of mud crab, Scylla paramamosain. Prog. Nat. Sci. 2008, 18, 691–695. [Google Scholar] [CrossRef]
- Lago-Lestón, A.; Ponce, E.; Muñoz, M.E. Cloning and expression of hyperglycemic (CHH) and molt-inhibiting (MIH) hormones mRNAs from the eyestalk of shrimps of Litopenaeus vannamei grown in different temperature and salinity conditions. Aquaculture 2007, 270, 343–357. [Google Scholar] [CrossRef]
- Qiao, H.; Xiong, Y.; Zhang, W.; Fu, H.; Jiang, S.; Sun, S.; Bai, H.; Jin, S.; Gong, Y. Characterization, expression, and function analysis of gonad-inhibiting hormone in Oriental River prawn, Macrobrachium nipponense and its induced expression by temperature. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 185, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef]
- Little, A.G.; Seebacher, F. The evolution of endothermy is explained by thyroid hormone-mediated responses to cold in early vertebrates. J. Exp. Biol. 2014, 217, 1642–1648. [Google Scholar] [CrossRef]
- Moriya, T. The effect of temperature on the action of thyroid hormone and prolactin in larvae of the salamander Hynobius retardatus. Gen. Comp. Endocrinol. 1983, 49, 1–7. [Google Scholar] [CrossRef]
- Politis, S.N.; Servili, A.; Mazurais, D.; Zambonino-Infante, J.-L.; Miest, J.J.; Tomkiewicz, J.; Butts, I. Temperature induced variation in gene expression of thyroid hormone receptors and deiodinases of European eel (Anguilla anguilla) larvae. Gen. Comp. Endocrinol. 2018, 259, 54–65. [Google Scholar] [CrossRef]
- Hammond, S.A.; Veldhoen, N.; Helbing, C.C. Influence of temperature on thyroid hormone signaling and endocrine disruptor action in Rana (Lithobates) catesbeiana tadpoles. Gen. Comp. Endocrinol. 2015, 219, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Bizhanova, A.; Kopp, P. The sodium-iodide symporter NIS and pendrin in iodide homeostasis of the thyroid. Endocrinology 2009, 150, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.C.; Klootwijk, W.; van Toor, H.; Pinto, G.; D’Alessandro, M.; Lèger, A.; Goudie, D.; Polak, M.; Grüters, A.; Visser, T.J. Mutations in the iodotyrosine deiodinase gene and hypothyroidism. N. Engl. J. Med. 2008, 358, 1811–1818. [Google Scholar] [CrossRef]
- Little, A.G.; Kunisue, T.; Kannan, K.; Seebacher, F. Thyroid hormone actions are temperature-specific and regulate thermal acclimation in zebrafish (Danio rerio). BMC Biol. 2013, 11, 26. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, A.-L.; Xian, J.-A. Variation of free amino acid and carbohydrate concentrations in white shrimp, Litopenaeus vannamei: Effects of continuous cold stress. Aquaculture 2011, 317, 182–186. [Google Scholar] [CrossRef]
- Ighodaro, O.; Akinloye, O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef] [PubMed]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Gálvez, S.; Gadal, P. On the function of the NADP-dependent isocitrate dehydrogenase isoenzymes in living organisms. Plant Sci. 1995, 105, 1–14. [Google Scholar] [CrossRef]
- Frei, B.; England, L.; Ames, B.N. Ascorbate is an outstanding antioxidant in human blood plasma. Proc. Natl. Acad. Sci. USA 1989, 86, 6377–6381. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerriero, G.; Di Finizio, A.; Ciarcia, G. Stress-induced changes of plasma antioxidants in aquacultured sea bass, Dicentrarchus labrax. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 205–211. [Google Scholar] [CrossRef]
- Han, B.; Kaur, V.I.; Baruah, K.; Nguyen, V.D.; Bossier, P. High doses of sodium ascorbate act as a prooxidant and protect gnotobiotic brine shrimp larvae (Artemia franciscana) against Vibrio harveyi infection coinciding with heat shock protein 70 activation. Dev. Comp. Immunol. 2019, 92, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Bullock, T.H. Compensation for temperature in the metabolism and activity of poikilotherms. Biol. Rev. 1955, 30, 311–342. [Google Scholar] [CrossRef]
- Sasseville, D. Neomycin. Dermatitis 2010, 21, 3–7. [Google Scholar] [CrossRef]
- Umezawa, H.; Ueda, M.; Maeda, K.; Yagishita, K.; Kondō, S.; Okami, Y.; Utahara, R.; Ōsato, Y.; Nitta, K.; Takeuchi, T. Production and isolation of a new antibiotic, kanamycin. J. Antibiot. Ser. A 1957, 10, 181–188. [Google Scholar]
- Yoshizawa, S.; Fourmy, D.; Puglisi, J.D. Structural origins of gentamicin antibiotic action. EMBO J. 1998, 17, 6437–6448. [Google Scholar] [CrossRef]
- Beard, E.L., Jr. The American Society of Health System Pharmacists. JONA’S Healthc. Law Ethics Regul. 2001, 3, 78–79. [Google Scholar] [CrossRef]
- Emanuele, E.; Bertona, M.; Altabas, K.; Altabas, V.; Alessandrini, G. Anti-inflammatory effects of a topical preparation containing nicotinamide, retinol, and 7-dehydrocholesterol in patients with acne: A gene expression study. Clin. Cosmet. Investig. Dermatol. 2012, 5, 33. [Google Scholar] [CrossRef]
- Ruamrak, C.; Lourith, N.; Natakankitkul, S. Comparison of clinical efficacies of sodium ascorbyl phosphate, retinol and their combination in acne treatment. Int. J. Cosmet. Sci. 2009, 31, 41–46. [Google Scholar] [CrossRef]
- Fenzl, A.; Kulterer, O.C.; Spirk, K.; Mitulović, G.; Marculescu, R.; Bilban, M.; Baumgartner-Parzer, S.; Kautzky-Willer, A.; Kenner, L.; Plutzky, J. Cold-mediated regulation of systemic retinol transport controls adipose tissue browning. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sundaresan, P.; Winters, V.G.; Therriault, D.G. Effect of low environmental temperature on the metabolism of vitamin A (retinol) in the rat. J. Nutr. 1967, 92, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yu, J.; Wang, H.; Liu, B.; Yue, X. p38 MAPK is involved in the immune response to pathogenic Vibrio in the clam Meretrix petechialis. Fish Shellfish Immunol. 2019, 95, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wen, H.; Qi, X.; Zhang, X.; Li, Y. Identification of mapk gene family in Lateolabrax maculatus and their expression profiles in response to hypoxia and salinity challenges. Gene 2019, 684, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P. Biology, environmental and nutritional modulation of skin mucus alkaline phosphatase in fish: A review. Fish Shellfish Immunol. 2019, 89, 179–186. [Google Scholar] [CrossRef] [PubMed]
- van Beek, J.H.; de Moor, M.H.; de Geus, E.J.; Lubke, G.H.; Vink, J.M.; Willemsen, G.; Boomsma, D.I. The genetic architecture of liver enzyme levels: GGT, ALT and AST. Behav. Genet. 2013, 43, 329–339. [Google Scholar] [CrossRef]
- Penning, T.M. The aldo-keto reductases (AKRs): Overview. Chem.-Biol. Interact. 2015, 234, 236–246. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Santoro, M.G. Stress–inducible responses and heat shock proteins: New pharmacologic targets for cytoprotection. Nat. Biotechnol. 1998, 16, 833–838. [Google Scholar] [CrossRef]
- Ahn, Y.-J.; Im, E. Heterologous expression of heat shock proteins confers stress tolerance in Escherichia coli, an industrial cell factory: A short review. Biocatal. Agric. Biotechnol. 2020, 29, 101833. [Google Scholar] [CrossRef]
- Chen, T.; Lin, T.; Li, H.; Lu, T.; Li, J.; Huang, W.; Sun, H.; Jiang, X.; Zhang, J.; Yan, A. Heat shock protein 40 (HSP40) in pacific white shrimp (Litopenaeus vannamei): Molecular cloning, tissue distribution and ontogeny, response to temperature, acidity/alkalinity and salinity stresses, and potential role in ovarian development. Front. Physiol. 2018, 9, 1784. [Google Scholar] [CrossRef]
- Qian, Z.; Liu, X.; Wang, L.; Wang, X.; Li, Y.; Xiang, J.; Wang, P. Gene expression profiles of four heat shock proteins in response to different acute stresses in shrimp, Litopenaeus vannamei. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 156, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Mallouk, Y.; Vayssier-Taussat, M.; Bonventre, J.V.; Polla, B.S. Heat shock protein 70 and ATP as partners in cell homeostasis. Int. J. Mol. Med. 1999, 4, 463–537. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.E. The ATP synthase: The understood, the uncertain and the unknown. Biochem. Soc. Transact. 2013, 41, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P. Roles of heat-shock proteins in innate and adaptive immunity. Nat. Rev. Immunol. 2002, 2, 185–194. [Google Scholar] [CrossRef]
- Pockley, A.G.; Henderson, B. Extracellular cell stress (heat shock) proteins—Immune responses and disease: An overview. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160522. [Google Scholar] [CrossRef]
- Beere, H.M.; Green, D.R. Stress management–heat shock protein-70 and the regulation of apoptosis. Trends Cell Biol. 2001, 11, 6–10. [Google Scholar] [CrossRef]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Denis-Larose, C.; Massie, B. Role of the human heat shock protein hsp70 in protection against stress-induced apoptosis. Mol. Cell. Biol. 1997, 17, 5317–5327. [Google Scholar] [CrossRef]
- Ravagnan, L.; Gurbuxani, S.; Susin, S.A.; Maisse, C.; Daugas, E.; Zamzami, N.; Mak, T.; Jäättelä, M.; Penninger, J.M.; Garrido, C. Heat-shock protein 70 antagonizes apoptosis-inducing factor. Nat. Cell Biol. 2001, 3, 839–843. [Google Scholar] [CrossRef]
- Luckinbill, L. Selection for longevity confers resistance to low-temperature stress in Drosophila melanogaster. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 1998, 53, B147–B153. [Google Scholar] [CrossRef]
- Lee, M.-C.; Yoon, D.-S.; Lee, Y.; Choi, H.; Shin, K.-H.; Park, H.G.; Lee, J.-S. Effects of low temperature on longevity and lipid metabolism in the marine rotifer Brachionus koreanus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 250, 110803. [Google Scholar] [CrossRef]
- Zhang, B.; Xiao, R.; Ronan, E.A.; He, Y.; Hsu, A.-L.; Liu, J.; Xu, X.S. Environmental temperature differentially modulates C. elegans longevity through a thermosensitive TRP channel. Cell Rep. 2015, 11, 1414–1424. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.; Ma, H.; Burch, D.; Maciel, G.A.; Hunter, T.; Dillin, A. SMK-1, an essential regulator of DAF-16-mediated longevity. Cell 2006, 124, 1039–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazi, A.; Henis-Korenblit, S.; Kenyon, C. A transcription elongation factor that links signals from the reproductive system to lifespan extension in Caenorhabditis elegans. PLoS Genet. 2009, 5, e1000639. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, J.A.; Mertens, P.R. Cold shock proteins: From cellular mechanisms to pathophysiology and disease. Cell Commun. Signal. 2018, 16, 63. [Google Scholar] [CrossRef] [PubMed]
- Phadtare, S.; Alsina, J.; Inouye, M. Cold-shock response and cold-shock proteins. Curr. Opin. Microbiol. 1999, 2, 175–180. [Google Scholar] [CrossRef]
- Sun, S.; Fu, H.; Ge, X.; Zhu, J.; Qiao, H.; Jin, S.; Zhang, W. Molecular cloning and expression analysis of cold shock protein Y-box gene from oriental river pawn (Macrobrachium nipponense). J. Fish. China 2017, 41, 1345–1354. [Google Scholar]
- Meng, Q.; Chen, J.; Huang, Y.; Jin, M.; Wei, G.; Wang, W. Molecular cloning and expression analysis of the cold shock protein Y-box coding gene of red claw crayfish, Cherax quadricarinatus. Freshw. Fish. 2012, 42, 14–20. [Google Scholar]
- Hui, M.; Cheng, J.; Sha, Z. Adaptation to the deep-sea hydrothermal vents and cold seeps: Insights from the transcriptomes of Alvinocaris longirostris in both environments. Deep Sea Res. Part I Oceanogr. Res. Pap. 2018, 135, 23–33. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Manfrin, C.; Pallavicini, A.; Battistella, S.; Lorenzon, S.; Giulianini, P.G. Crustacean immunity: The modulation of stress responses. In Lessons in Immunity; Elsevier: Amsterdam, The Netherlands, 2016; pp. 107–116. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 30 | 25 | 20 | 15 | 10 | |
|---|---|---|---|---|---|
| Initial crayfish wet weight (g) | 3.40 ± 0.25 | 3.61 ± 0.32 | 3.47 ± 0.25 | 3.41 ± 0.41 | 3.39 ± 0.43 |
| Initial crayfish body length (cm) | 3.31 ± 0.45 | 3.29 ± 0.26 | 3.06 ± 0.40 | 3.32 ± 0.48 | 3.22 ± 0.35 |
| Final crayfish wet weight (g) | 10.86 ± 1.39 | 10.44 ± 0.74 | 9.95 ± 0.95 | 4.64 ± 0.26 | 4.46 ± 0.23 |
| Final crayfish body length (cm) | 7.95 ± 0.29 | 7.67 ± 0.37 | 7.62 ± 0.36 | 4.67 ± 0.53 | 4.63 ± 0.29 |
| Weight gain rate (WG, %) | 68.67% b | 65.41% b | 65.14% b | 26.40% a | 24.05% a |
| Length gain rate (LG, %) | 58.37% b | 57.08% b | 59.86% b | 28.82% a | 30.55% a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Xu, W.; Jiang, Q.; Ye, Y.; Tian, J.; Huang, Y.; Du, X.; Li, Y.; Zhao, Y.; Liu, Z. Effects of Low Temperature on Antioxidant and Heat Shock Protein Expression Profiles and Transcriptomic Responses in Crayfish (Cherax destructor). Antioxidants 2022, 11, 1779. https://doi.org/10.3390/antiox11091779
Yang Y, Xu W, Jiang Q, Ye Y, Tian J, Huang Y, Du X, Li Y, Zhao Y, Liu Z. Effects of Low Temperature on Antioxidant and Heat Shock Protein Expression Profiles and Transcriptomic Responses in Crayfish (Cherax destructor). Antioxidants. 2022; 11(9):1779. https://doi.org/10.3390/antiox11091779
Chicago/Turabian StyleYang, Ying, Wenyue Xu, Qichen Jiang, Yucong Ye, Jiangtao Tian, Yingying Huang, Xinglin Du, Yiming Li, Yunlong Zhao, and Zhiquan Liu. 2022. "Effects of Low Temperature on Antioxidant and Heat Shock Protein Expression Profiles and Transcriptomic Responses in Crayfish (Cherax destructor)" Antioxidants 11, no. 9: 1779. https://doi.org/10.3390/antiox11091779
APA StyleYang, Y., Xu, W., Jiang, Q., Ye, Y., Tian, J., Huang, Y., Du, X., Li, Y., Zhao, Y., & Liu, Z. (2022). Effects of Low Temperature on Antioxidant and Heat Shock Protein Expression Profiles and Transcriptomic Responses in Crayfish (Cherax destructor). Antioxidants, 11(9), 1779. https://doi.org/10.3390/antiox11091779