
Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus)

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Formulation and Characterization of Omega-3 Nanoparticles
2.3. Experimental Fish and Design
2.4. Formulation of Experimental Diet
2.5. Monitoring Nile Tilapia Performance
2.6. Blood and Tissue Sampling
2.7. Fatty Acid Profile
2.8. Oxidative Stress-Related Biomarkers in Muscle
2.9. Hematological and Serum Biochemical Evaluation
2.10. Immunological Parameters Assessment
2.11. Quantitative Real-Time PCR
2.12. Aeromonas Hydrophila Challenge and Sampling
2.13. Expression Analysis of A. hydrophila Virulence Genes
2.14. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Fatty Acid Composition and Antioxidant Biomarkers in Fish Flesh
3.3. Hematological and Serum Biochemical Indicators
3.4. Relative Expression of Antioxidant-Related Genes before Challenge in Muscle and Intestine
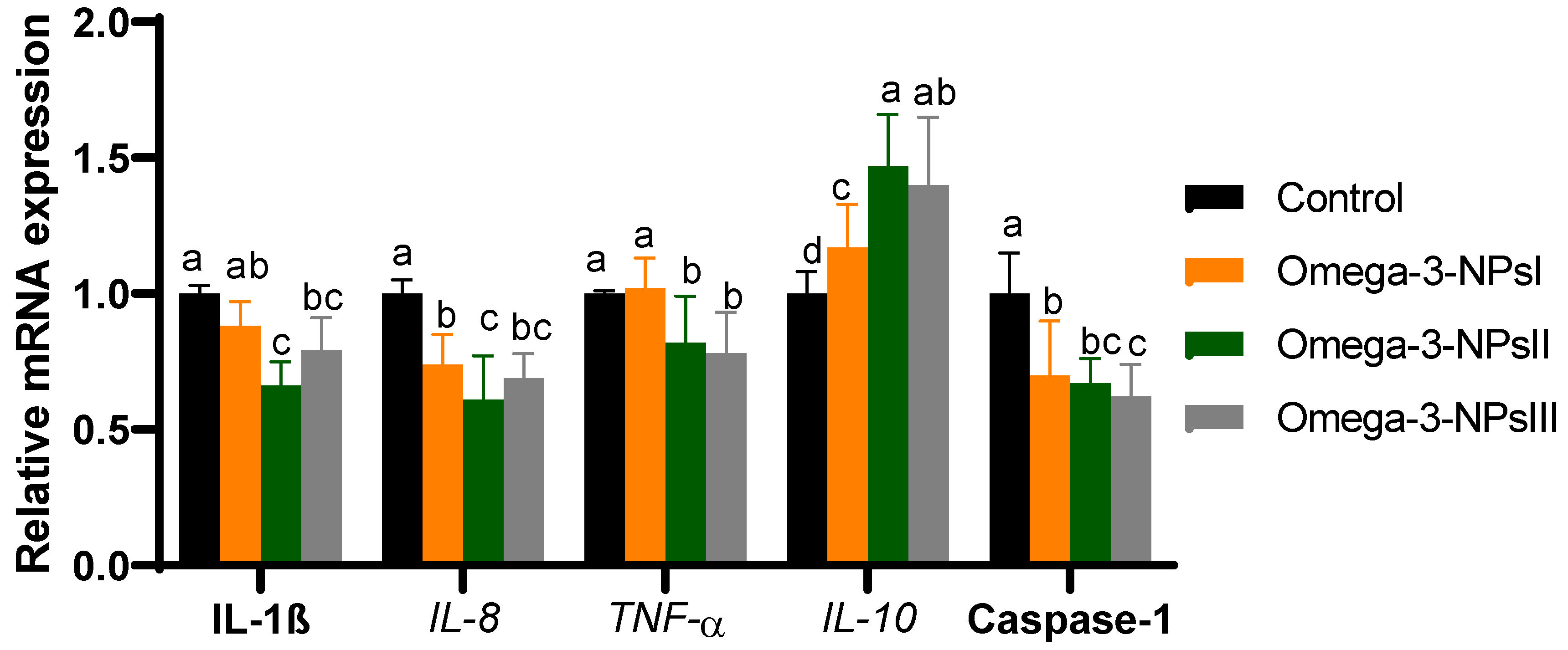
3.5. Relative Expression of Immune-Related Genes Post-Challenge in Splenic Tissue
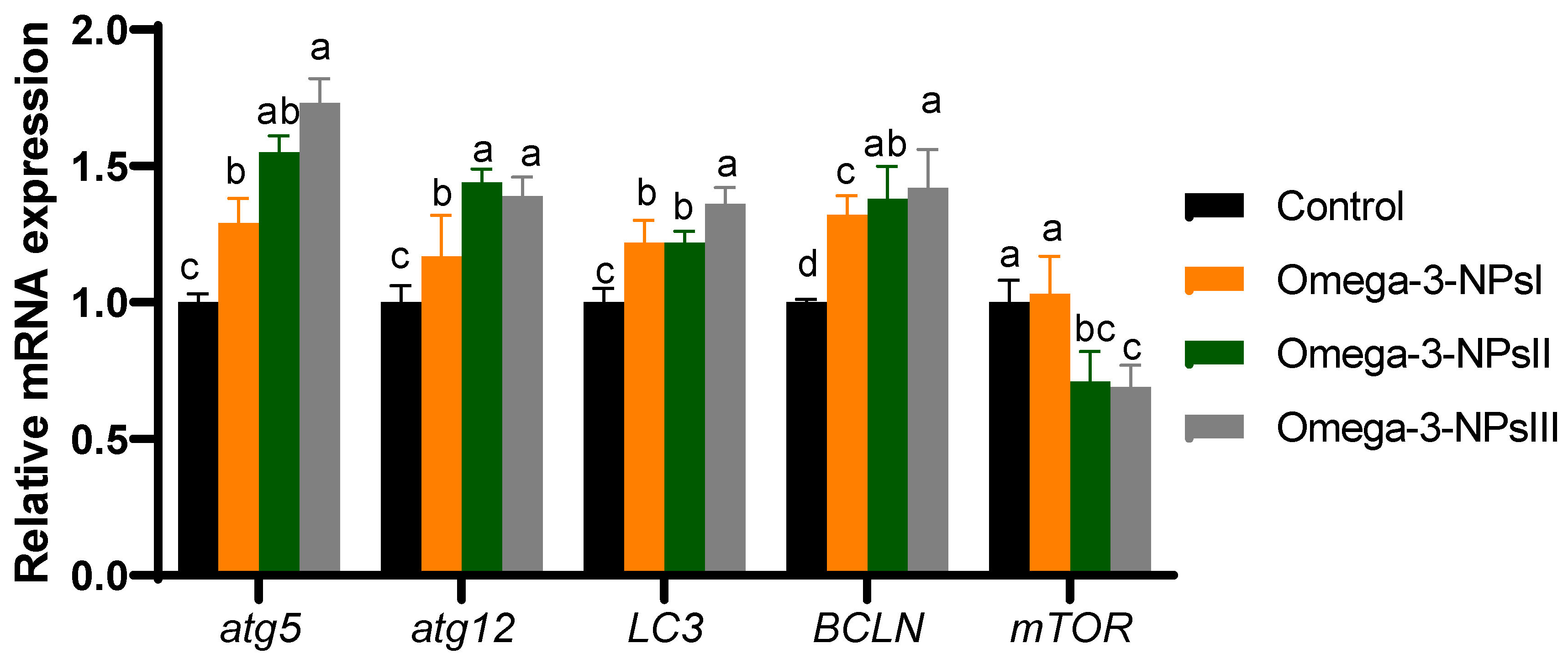
3.6. Relative Expression of Autophagy-Related Genes Post-Challenge in Liver
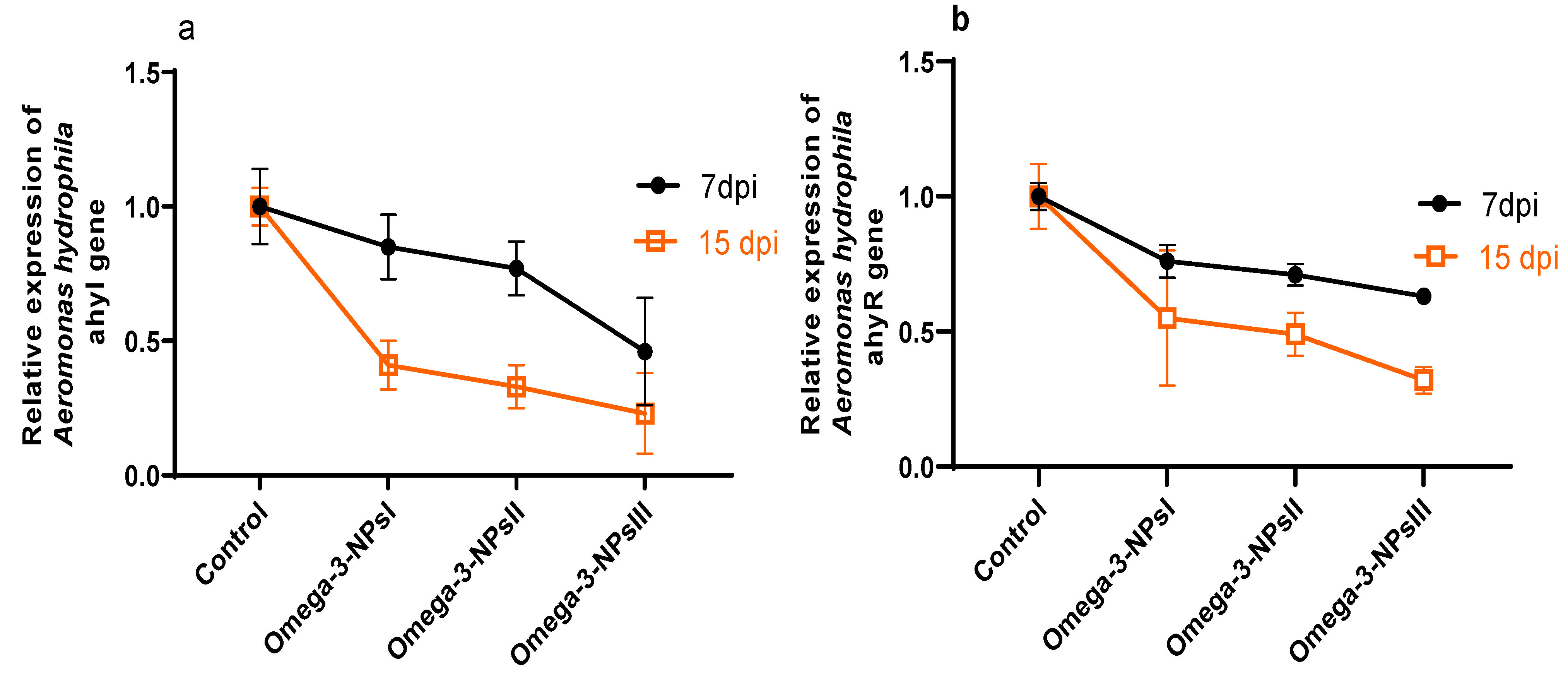
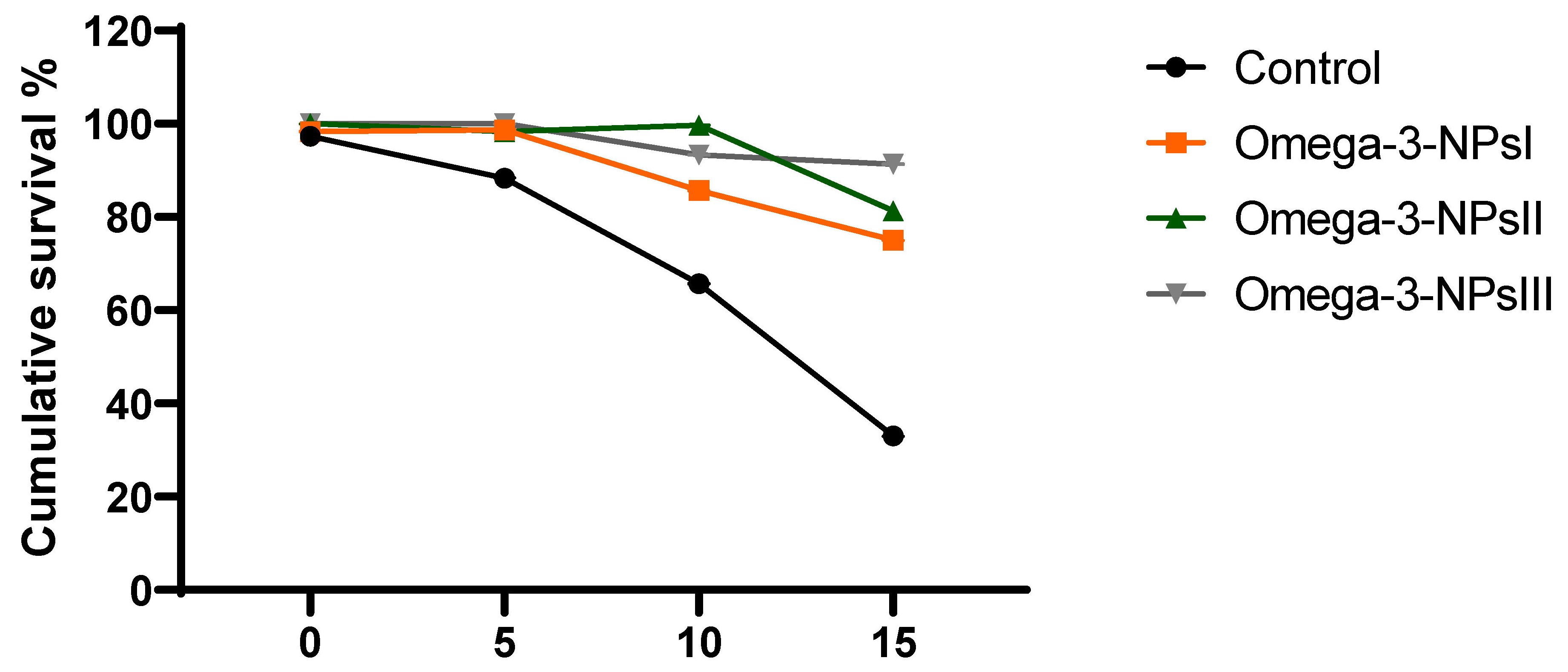
3.7. Effect of Omega-3 NPs on Relative Expression of A. hydrophila Virulence-Related Genes and Survival Percentage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, C.; Beck, B.H.; Peatman, E. Nutritional impacts on gene expression in the surface mucosa of blue catfish (Ictalurus furcatus). Dev. Comp. Immunol. 2014, 44, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Turchini, G.; Francis, D.; Senadheera, S.; Thanuthong, T.; De Silva, S. Fish oil replacement with different vegetable oils in Murray cod: Evidence of an “omega-3 sparing effect” by other dietary fatty acids. Aquaculture 2011, 315, 250–259. [Google Scholar] [CrossRef]
- Heal, R.D.; Hasan, N.A.; Haque, M.M. Increasing disease burden and use of drugs and chemicals in Bangladesh shrimp aquaculture: A potential menace to human health. Mar. Pollut. Bull. 2021, 172, 112796. [Google Scholar] [CrossRef] [PubMed]
- Singha, K.P.; Shamna, N.; Sahu, N.P.; Sardar, P.; Harikrishna, V.; Thirunavukkarasar, R.; Chowdhury, D.K.; Maiti, M.K.; Krishna, G. Optimum dietary crude protein for culture of genetically improved farmed tilapia (GIFT), Oreochromis niloticus (Linnaeus, 1758) juveniles in low inland saline water: Effects on growth, metabolism and gene expression. Anim. Feed Sci. Technol. 2021, 271, 114713. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; Ibrahim, S.M.; Eldemery, F.; El-Mandrawy, S.A.; Metwally, A.S.; Khalifa, E.; Elnahriry, S.S.; Ibrahim, D. Dietary cinnamaldehyde nanoemulsion boosts growth and transcriptomes of antioxidant and immune related genes to fight Streptococcus agalactiae infection in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2021, 113, 96–105. [Google Scholar] [CrossRef]
- Ibrahim, D.; Kishawy, A.T.; Khater, S.I.; Khalifa, E.; Ismail, T.A.; Mohammed, H.A.; Elnahriry, S.S.; Tolba, H.A.; Sherief, W.R.; Farag, M.F. Interactive effects of dietary quercetin nanoparticles on growth, flesh antioxidant capacity and transcription of cytokines and Aeromonas hydrophila quorum sensing orchestrating genes in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2021, 119, 478–489. [Google Scholar] [CrossRef]
- Stoneham, T.R.; Kuhn, D.D.; Taylor, D.P.; Neilson, A.P.; Smith, S.A.; Gatlin, D.M.; Chu, H.S.S.; O’Keefe, S.F. Production of omega-3 enriched tilapia through the dietary use of algae meal or fish oil: Improved nutrient value of fillet and offal. PLoS ONE 2018, 13, e0194241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.A.; Król, E. Nutrigenomics and immune function in fish: New insights from omics technologies. Dev. Comp. Immunol. 2017, 75, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Chandan, N.K.; Gupta, S.K.; Bhushan, S.; Patole, P.B. Omega-3 fatty acids effectively modulate growth performance, immune response, and disease resistance in fish against multiple stresses. Aquaculture 2022, 547, 737506. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Fernandes, D.C.; Eto, S.F.; Funnicelli, M.I.; Fernandes, C.C.; Charlie-Silva, I.; Belo, M.A.; Pizauro, J.M. Immunoglobulin Y in the diagnosis of Aeromonas hydrophila infection in Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 500, 576–585. [Google Scholar] [CrossRef]
- Kirke, D.F.; Swift, S.; Lynch, M.J.; Williams, P. The Aeromonas hydrophila LuxR homologue AhyR regulates the N-acyl homoserine lactone synthase, AhyI positively and negatively in a growth phase-dependent manner. FEMS Microbiol. Lett. 2004, 241, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, R.; Ai, Q.; Mai, K.; Xu, W.; Wang, J.; Xu, H.; Liufu, Z.; Zhang, Y. Effects of dietary n-3 highly unsaturated fatty acids on growth, nonspecific immunity, expression of some immune related genes and disease resistance of large yellow croaker (Larmichthys crocea) following natural infestation of parasites (Cryptocaryon irritans). Fish Shellfish Immunol. 2012, 32, 249–258. [Google Scholar] [PubMed]
- Montero, D.; Mathlouthi, F.; Tort, L.; Afonso, J.; Torrecillas, S.; Fernández-Vaquero, A.; Negrin, D.; Izquierdo, M. Replacement of dietary fish oil by vegetable oils affects humoral immunity and expression of pro-inflammatory cytokines genes in gilthead sea bream Sparus aurata. Fish Shellfish Immunol. 2010, 29, 1073–1081. [Google Scholar] [CrossRef]
- Muchow, M.; Schmitz, E.; Despatova, N.; Maincent, P.; Müller, R. Omega-3 fatty acids-loaded lipid nanoparticles for patient-convenient oral bioavailability enhancement. Die Pharmazie Int. J. Pharm. Sci. 2009, 64, 499–504. [Google Scholar]
- Fajardo, C.; Martinez-Rodriguez, G.; Blasco, J.; Mancera, J.M.; Thomas, B.; De Donato, M. Nanotechnology in aquaculture: Applications, perspectives and regulatory challenges. Aquac. Fish. 2022, 7, 185–200. [Google Scholar] [CrossRef]
- Ibrahim, D.; Nem, A.N.A.; Ibrahim, S.M.; Eissa, H.M.; Fawzey, M.; Mostafa, D.I.; Abd El-Kader, S.A.; Khater, S.; Khater, S.I. Dual effect of Selenium loaded Chitosan Nanoparticles on growth, antioxidant, immune related genes expression, transcriptomics modulation of caspase 1, cytochrome P450 and heat shock protein and Aeromonas hydrophila resistance of Nile Tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2021, 110, 91–99. [Google Scholar]
- Cui, J.; Yu, B.; Zhao, Y.; Zhu, W.; Li, H.; Lou, H.; Zhai, G. Enhancement of oral absorption of curcumin by self-microemulsifying drug delivery systems. Int. J. Pharm. 2009, 371, 148–155. [Google Scholar] [CrossRef]
- Shah, B.R.; Mraz, J. Advances in nanotechnology for sustainable aquaculture and fisheries. Rev. Aquac. 2020, 12, 925–942. [Google Scholar] [CrossRef]
- Ibrahim, D.; Ismail, T.A.; Khalifa, E.; El-Kader, A.; Shaimaa, A.; Mohamed, D.I.; Mohamed, D.T.; Shahin, S.E.; El-Hamid, A.; Marwa, I. Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model. Animals 2021, 11, 2027. [Google Scholar] [CrossRef]
- Vieira, M.d.C.; Bakof, K.K.; Schuch, N.J.; Skupien, J.A.; Boeck, C.R. Os benefícios da nanoencapsulação de ácidos graxos da classe ômega-3 para o enriquecimento de produtos alimentícios: Uma revisão. Rev. De Nutr. 2020, 33. [Google Scholar]
- NRC. Nutrient requirements of fish. In Nutrient Requirements of Domestic Animal Series; National Academy Press: Washigton, DC, USA, 1993. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International; AOAC: Washington, DC, USA, 2002; Volume 13. [Google Scholar]
- Alandiyjany, M.N.; Kishawy, A.T.; Abdelfattah-Hassan, A.; Eldoumani, H.; Elazab, S.T.; El-Mandrawy, S.A.; Saleh, A.A.; ElSawy, N.A.; Attia, Y.A.; Arisha, A.H. Nano-silica and magnetized-silica mitigated lead toxicity: Their efficacy on bioaccumulation risk, performance, and apoptotic targeted genes in Nile tilapia (Oreochromis niloticus). Aquat. Toxicol. 2022, 242, 106054. [Google Scholar] [CrossRef] [PubMed]
- Eleraky, W.; Ibrahim, D.; Mahmoud, R. Effects of rare earth elements and exogenous multienzyme supplementation to plant protein enriched diet on growth performance, digestibility and economic efficiency of Nile tilapia, Oreochromis niloticus. Jpn. J. Vet. Res. 2016, 64, S73–S78. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2′, 7′-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Livingstone, D.; Martinez, P.G.; Michel, X.; Narbonne, J.; O’hara, S.; Ribera, D.; Winston, G. Oxyradical production as a pollution-mediated mechanism of toxicity in the common mussel, Mytilus edulis L., and other molluscs. Funct. Ecol. 1990, 4, 415–424. [Google Scholar] [CrossRef]
- Goldenfarb, P.B.; Bowyer, F.P.; Hall, E.; Brosious, E. Reproducibility in the hematology laboratory: The microhematocrit determination. Am. J. Clin. Pathol. 1971, 56, 35–39. [Google Scholar] [CrossRef]
- Fahey, J.L.; McKelvey, E.M. Quantitative determination of serum immunoglobulins in antibody-agar plates. J. Immunol. 1965, 94, 84–90. [Google Scholar]
- Schultz, L. Methods in Clinical Chemistry; The CV Mosby Co: St Louis, MI, USA, 1987; pp. 742–746. [Google Scholar]
- Yilmaz, S.; Ergün, S.; Yıgıt, M. Effects of dietary FARMARIN® XP supplement on immunological responses and disease resistance of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 211–220. [Google Scholar] [CrossRef]
- Andani, H.; Tukmechi, A.; Meshkini, S.; Sheikhzadeh, N. Antagonistic activity of two potential probiotic bacteria from fish intestines and investigation of their effects on growth performance and immune response in rainbow trout (Oncorhynchus mykiss). J. Appl. Ichthyol. 2012, 28, 728–734. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Algammal, A.M.; Mohamed, M.F.; Tawfiek, B.A.; Hozzein, W.N.; El Kazzaz, W.M.; Mabrok, M. Molecular typing, antibiogram and PCR-RFLP based detection of Aeromonas hydrophila complex isolated from Oreochromis niloticus. Pathogens 2020, 9, 238. [Google Scholar] [CrossRef] [Green Version]
- El-Gohary, F.A.; Zahran, E.; El-Gawad, A.; Eman, A.; El-Gohary, A.H.; Abdelhamid, M.F.; El-Mleeh, A.; Elmahallawy, E.K.; Elsayed, M.M. Investigation of the prevalence, virulence genes, and antibiogram of motile aeromonads Isolated from Nile tilapia fish farms in Egypt and assessment of their water quality. Animals 2020, 10, 1432. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.O.; SANTOS, V.G.; Pezzato, L.E.; Carvalho, P.L.D.; Teixeira, C.P.; Freitas, J.; Padovani, C.R.; Sartori, M.M.; Barros, M.M. Activity of Brazilian propolis against Aeromonas hydrophila and its effect on Nile tilapia growth, hematological and non-specific immune response under bacterial infection. An. Acad. Bras. Cienc. 2017, 89, 1785–1799. [Google Scholar] [CrossRef] [PubMed]
- Tanhay Mangoudehi, H.; Zamani, H.; Shahangian, S.S.; Mirzanejad, L. Effect of curcumin on the expression of ahyI/R quorum sensing genes and some associated phenotypes in pathogenic Aeromonas hydrophila fish isolates. World J. Microbiol. Biotechnol. 2020, 36, 70. [Google Scholar] [CrossRef]
- Nakharuthai, C.; Rodrigues, P.M.; Schrama, D.; Kumkhong, S.; Boonanuntanasarn, S. Effects of different dietary vegetable lipid sources on health status in Nile tilapia (Oreochromis niloticus): Haematological indices, immune response parameters and plasma proteome. Animals 2020, 10, 1377. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef]
- Xu, H.; Wang, J.; Mai, K.; Xu, W.; Zhang, W.; Zhang, Y.; Ai, Q. Dietary docosahexaenoic acid to eicosapentaenoic acid (DHA/EPA) ratio influenced growth performance, immune response, stress resistance and tissue fatty acid composition of juvenile Japanese seabass, Lateolabrax japonicus (Cuvier). Aquac. Res. 2016, 47, 741–757. [Google Scholar] [CrossRef]
- Gingras, A.A.; White, P.J.; Chouinard, P.Y.; Julien, P.; Davis, T.A.; Dombrowski, L.; Couture, Y.; Dubreuil, P.; Myre, A.; Bergeron, K. Long-chain omega-3 fatty acids regulate bovine whole-body protein metabolism by promoting muscle insulin signalling to the Akt–mTOR–S6K1 pathway and insulin sensitivity. J. Physiol. 2007, 579, 269–284. [Google Scholar] [CrossRef]
- Wei, H.-K.; Deng, Z.; Jiang, S.-Z.; Song, T.-X.; Zhou, Y.-F.; Peng, J.; Tao, Y.-X. Eicosapentaenoic acid abolishes inhibition of insulin-induced mTOR phosphorylation by LPS via PTP1B downregulation in skeletal muscle. Mol. Cell. Endocrinol. 2017, 439, 116–125. [Google Scholar] [CrossRef] [PubMed]
- De Souza, N.E.; Matsushita, M.; de Oliveira, C.C.; Franco, M.R.B.; Visentainer, J.V. Manipulation of fatty acid composition of Nile tilapia (Oreochromis niloticus) fillets with flaxseed oil. J. Sci. Food Agric. 2007, 87, 1677–1681. [Google Scholar] [CrossRef]
- Visentainer, J.V.; de Souza, N.E.; Makoto, M.; Hayashi, C.; Franco, M.R.B. Influence of diets enriched with flaxseed oil on the α-linolenic, eicosapentaenoic and docosahexaenoic fatty acid in Nile tilapia (Oreochromis niloticus). Food Chem. 2005, 90, 557–560. [Google Scholar] [CrossRef]
- Olsen, R.E.; Henderson, R.J.; McAndrew, B. The conversion of linoleic acid and linolenic acid to longer chain polyunsaturated fatty acids by Tilapia (Oreochromis) nilotica in vivo. Fish Physiol. Biochem. 1990, 8, 261–270. [Google Scholar] [CrossRef]
- Burdge, G. Metabolism of α-linolenic acid in humans. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 161–168. [Google Scholar] [CrossRef]
- Sinclair, A.J.; Attar-Bashi, N.M.; Li, D. What is the role of α-linolenic acid for mammals? Lipids 2002, 37, 1113–1123. [Google Scholar] [CrossRef]
- Dos Santos, S.K.A.; Schorer, M.; Moura, G.d.S.; Lanna, E.A.T.; Pedreira, M.M. Evaluation of growth and fatty acid profile of Nile tilapia (Oreochromis niloticus) fed with Schizochytrium sp. Aquac. Res. 2019, 50, 1068–1074. [Google Scholar] [CrossRef]
- Hossain, M.; Al-Adul-Elah, K.; Azad, I.; Alzalzalah, A.; Alnuiami, S. High DHA Algae Meal as Cost-effective Alternative to High DHA Fish Oil in Finisher Feed for Sobaity Sea Bream (Sparidentex hasta). Anim. Feed Sci. Technol. 2022, 115209. [Google Scholar] [CrossRef]
- Peng, M.; Xu, W.; Tan, P.; Du, J.; Mai, K.; Zhou, H.; Zhang, Y.; Nian, R.; Macq, B.; Ai, Q. Effect of dietary fatty acid composition on growth, fatty acids composition and hepatic lipid metabolism in juvenile turbot (Scophthalmus maximus L.) fed diets with required n3 LC-PUFAs. Aquaculture 2017, 479, 591–600. [Google Scholar] [CrossRef]
- Winston, G.W.; Di Giulio, R.T. Prooxidant and antioxidant mechanisms in aquatic organisms. Aquat. Toxicol. 1991, 19, 137–161. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant defenses in fish: Biotic and abiotic factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Koruk, M.; Taysi, S.; Savas, M.C.; Yilmaz, O.; Akcay, F.; Karakok, M. Oxidative stress and enzymatic antioxidant status in patients with nonalcoholic steatohepatitis. Ann. Clin. Lab. Sci. 2004, 34, 57–62. [Google Scholar] [PubMed]
- Wu, D.; Lewis, E.D.; Pae, M.; Meydani, S.N. Nutritional modulation of immune function: Analysis of evidence, mechanisms, and clinical relevance. Front. Immunol. 2019, 9, 3160. [Google Scholar] [CrossRef]
- Benítez-Dorta, V.; Caballero, M.J.; Izquierdo, M.; Manchado, M.; Infante, C.; Zamorano, M.J.; Montero, D. Total substitution of fish oil by vegetable oils in Senegalese sole (Solea senegalensis) diets: Effects on fish performance, biochemical composition, and expression of some glucocorticoid receptor-related genes. Fish Physiol. Biochem. 2013, 39, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Kiron, V.; Thawonsuwan, J.; Panigrahi, A.; Scharsack, J.; Satoh, S. Antioxidant and immune defences of rainbow trout (Oncorhynchus mykiss) offered plant oils differing in fatty acid profiles from early stages. Aquacult. Nutr. 2011, 17, 130–140. [Google Scholar] [CrossRef]
- An, W.; Dong, X.; Tan, B.; Yang, Q.; Chi, S.; Zhang, S.; Liu, H.; Yang, Y. Effects of dietary n-3 highly unsaturated fatty acids on growth, non-specific immunity, expression of some immune-related genes and resistance to Vibrio harveyi in hybrid grouper (♀ Epinephelus fuscoguttatus×♂ Epinephelus lanceolatu). Fish Shellfish Immunol. 2020, 96, 86–96. [Google Scholar] [CrossRef]
- Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of omega-3 fatty acids on immune cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Ai, Q.; Mai, K.; Xu, W.; Zheng, Y. A comparative study: In vitro effects of EPA and DHA on immune functions of head-kidney macrophages isolated from large yellow croaker (Larmichthys crocea). Fish Shellfish Immunol. 2013, 35, 933–940. [Google Scholar] [CrossRef]
- Serini, S.; Cassano, R.; Trombino, S.; Calviello, G. Nanomedicine-based formulations containing ω-3 polyunsaturated fatty acids: Potential application in cardiovascular and neoplastic diseases. Int. J. Nanomed. 2019, 14, 2809. [Google Scholar] [CrossRef] [Green Version]
- Nagar, R. Autophagy: A brief overview in perspective of dermatology. Indian J. Dermatol. Venereol. Leprol. 2017, 83, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Jagannath, C.; Liu, X.-D.; Sharafkhaneh, A.; Kolodziejska, K.E.; Eissa, N.T. Toll-like receptor 4 is a sensor for autophagy associated with innate immunity. Immunity 2007, 27, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishawy, A.T.; Al-Khalaifah, H.S.; Nada, H.S.; Roushdy, E.M.; Zaglool, A.W.; Ahmed Ismail, T.; Ibrahim, S.M.; Ibrahim, D. Black Pepper or Radish Seed Oils in a New Combination of Essential Oils Modulated Broiler Chickens’ Performance and Expression of Digestive Enzymes, Lipogenesis, Immunity, and Autophagy-Related Genes. Vet. Sci. 2022, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Spits, H.; Lanier, L.L. Natural killer or dendritic: What’s in a name? Immunity 2007, 26, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, K.; Monsen, V.T.; Pettersen, C.H.H.; Overland, H.B.; Pettersen, G.; Samdal, H.; Tesfahun, A.N.; Lundemo, A.G.; Bjørkøy, G.; Schønberg, S.A. DHA-induced stress response in human colon cancer cells–Focus on oxidative stress and autophagy. Free Radic. Biol. Med. 2016, 90, 158–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Tang, Y.; Wang, S.; Zhou, J.; Zhou, J.; Lu, X.; Bai, X.; Wang, X.-Y.; Chen, Z.; Zuo, D. Endogenous n-3 polyunsaturated fatty acids attenuate T cell-mediated hepatitis via autophagy activation. Front. Immunol. 2016, 7, 350. [Google Scholar] [CrossRef] [Green Version]
- Paquette, M.; El-Houjeiri, L.; Pause, A. mTOR pathways in cancer and autophagy. Cancers 2018, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Levine, B.; Klionsky, D.J. Development by self-digestion: Molecular mechanisms and biological functions of autophagy. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abeliovich, H.; Agostinis, P.; Agrawal, D.K.; Aliev, G.; Askew, D.S.; Baba, M.; Baehrecke, E.H.; Bahr, B.A.; Ballabio, A. Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes. Autophagy 2008, 4, 151–175. [Google Scholar] [CrossRef]
- Liang, X.; Jackson, S.; Seaman, M.; Brown, K.; Kempkes, B.; Hibshoosh, H.; Levine, B. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature 1999, 402, 672–676. [Google Scholar] [CrossRef]
- Kim, Y.C.; Guan, K.-L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.; Jing, K.; Jeong, S.; Kim, N.; Song, K.-S.; Heo, J.-Y.; Park, J.-H.; Seo, K.-S.; Han, J.; Park, J.-I. The omega-3 polyunsaturated fatty acid DHA induces simultaneous apoptosis and autophagy via mitochondrial ROS-mediated Akt-mTOR signaling in prostate cancer cells expressing mutant p53. BioMed Res. Int. 2013, 2013, 568671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams-Bey, Y.; Boularan, C.; Vural, A.; Huang, N.-N.; Hwang, I.-Y.; Shan-Shi, C.; Kehrl, J.H. Omega-3 free fatty acids suppress macrophage inflammasome activation by inhibiting NF-κB activation and enhancing autophagy. PLoS ONE 2014, 9, e97957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, Q.; Mai, K.; Zhang, L.; Tan, B.; Zhang, W.; Xu, W.; Li, H. Effects of dietary β-1, 3 glucan on innate immune response of large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol. 2007, 22, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Sewid, A.H.; Arisha, A.H.; Abd El-Fattah, A.H.; Abdelaziz, A.M.; Al-Jabr, O.A.; Kishawy, A.T. Influence of Glycyrrhiza glabra Extract on Growth, Gene Expression of Gut Integrity, and Campylobacter jejuni Colonization in Broiler Chickens. Front. Vet. Sci 2020, 7, 612063. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-L.; Huang, S.-J.; Wu, C.-L.; Gong, H.-Y.; Ken, C.-F.; Hu, S.-Y.; Wu, J.-L. Transgenic expression of omega-3 PUFA synthesis genes improves zebrafish survival during Vibrio vulnificus infection. J. Biomed. Sci. 2015, 22, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Romero, J.C.; González-Ríos, H.; Borges, A.; Simões, M. Antibacterial effects and mode of action of selected essential oils components against Escherichia coli and Staphylococcus aureus. Evid.-Based Complement. Altern. Med. 2015, 2015, 795435. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, % | Control Diet | Supplemental Omega-3-NPs (g/kg Diet) | ||
|---|---|---|---|---|
| 0.05% | 0.1% | 0.2% | ||
| Fish meal | 16.5 | 16.5 | 16.5 | 16.5 |
| Soybean meal | 33.2 | 33.2 | 33.2 | 33.2 |
| Yellow corn | 28 | 28 | 28 | 28 |
| Corn gluten | 4 | 4 | 4 | 4 |
| Rice bran | 12 | 12 | 12 | 12 |
| Soy oil | 3.5 | 3.45 | 3.4 | 3.3 |
| Lysine | 0.10 | 0.10 | 0.10 | 0.10 |
| DL- Methionine (98%) | 0.20 | 0.20 | 0.20 | 0.20 |
| Threonine | 0.10 | 0.10 | 0.10 | 0.10 |
| Di-calcium phosphate | 1.20 | 1.20 | 1.20 | 1.20 |
| * Vitamins and minerals premix | 1.20 | 1.20 | 1.20 | 1.20 |
| DHA + EPA | 0 | 0.05 | 0.1 | 0.2 |
| Chemical analysis | ||||
| Digestible energy (kcal/kg) | 2926 | 2922 | 2918 | 2909 |
| Crude protein, % | 32.10 | 32.10 | 32.10 | 32.10 |
| Ether extract, % | 4.71 | 4.71 | 4.71 | 4.71 |
| Ca, % | 0.80 | 0.80 | 0.80 | 0.80 |
| Available P, % | 0.43 | 0.43 | 0.43 | 0.43 |
| Lysine, % | 2.00 | 2.00 | 2.00 | 2.00 |
| Methionine, % | 0.81 | 0.81 | 0.81 | 0.81 |
| Fatty Acids | Supplemental Omega-3-NPs (g/kg Diet) | |||
|---|---|---|---|---|
| Control | 1 | 2 | 3 | |
| 18:2n − 6 | 2.053 | 2.053 | 2.053 | 2.053 |
| C18:3n − 6 (Gamma linolenic acid) | 0.002 | 0.002 | 0.002 | 0.002 |
| C18:3n − 3 (Alpha linolenic acid) | 0.074 | 0.074 | 0.074 | 0.074 |
| C18:4n3 (Stearidonic acid) | 0.000 | 0.08 | 0.01 | 0.000 |
| 20:2n − 6 | 0.07 | 0.00 | 0.00 | 0.000 |
| 20:3n − 3 | 0.06 | 0.000 | 0.00 | 0.07 |
| 20.4 n − 6 | 0.004 | 0.000 | 0.000 | 0.06 |
| 20:5n − 3 EPA | 0.201 | 2.01 | 3.81 | 7.41 |
| 22:2 n − 6 | 0.008 | 0.006 | 0.03 | 0.002 |
| C22:5n − 3 Docosapentaenoic acid (DPA) | 0.037 | 0.037 | 0.037 | 0.037 |
| C22:6n − 3 Docosahexaenoic acid (DHA) | 0.095 | 1.30 | 2.50 | 4.90 |
| ∑n − 3 PUFA | 0.41 | 1.41 | 2.41 | 3.42 |
| ∑n − 6 PUFA | 2.01 | 2.01 | 2.01 | 2.01 |
| n − 6/n − 3 | 5.03 | 1.43 | 0.83 | 0.59 |
| Gene | Sequence | Accession No. |
|---|---|---|
| SOD | F-GACGTGACAACACAGGTTGC R-TACAGCCACCGTAACAGCAG | XM_003449940.5 |
| CAT | F-TCAGCACAGAAGACACAGACA R-GACCATTCCTCCACTCCAGAT | XM_031754288.1 |
| GSH-Px | F-CCAAGAGAACTGCAAGAACGA R-CAGGACACGTCATTCCTACAC | NM_001279711.1 |
| IL-1β | F-TGCTGAGCACAGAATTCCAG R-GCTGTGGAGAAGAACCAAGC | XM_019365841.2 |
| IL-6 | ||
| IL-8 | F-GCACTGCCGCTGCATTAAG R-GCAGTGGGAGTTGGGAAGAA | XM_031747075.1 |
| IL-10 | F-CTGCTAGATCAGTCCGTCGAA R-GCAGAACCGTGTCCAGGTAA | XM_013269189.3 |
| TNF-α | F-GAGGTCGGCGTGCCAAGA R-TGGTTTCCGTCCACAGCGT | NM_001279533.1 |
| Caspase 1 | F: GCTGTCTGAGTAAGTGTATCAT R: CCAACACGTTAAAATGGATCTGA | XM_019367762.2 |
| Atg5 | F-ATTGGCGTTTTGTTTGATCTT R-TTTGAGTGCATCCGCCTCTTT | XM_019082404.1 |
| Atg12 | F-ACAGTACAGTCACTCGCTCA R-AAAACACTCGAAAAGCACACC | XM_019125508.1 |
| LC3-II | F-GGAACAGCATCCAAGCAAGA R-TCAGAAATGGCGGTGGACA | NM199604.1 |
| BCLN1 | F-TCTGTTTGATATCATGTCTGG R-TAATTCTGGCACTCATTTTCT | XM_019068185.1 |
| mTOR | F-TGCGGAGTATGTGGAGTT R-CATCTCTTTGGTCTCTCTCTGG | XM_019108641.1 |
| β-actine | F-TGGCATCACACCTTCTATAACGA R-TGGCAGGAGTGTTGAAGGTCT | XM_003455949.2 |
| Parameter | Supplemental Omega-3-NPs (g/kg Diet) | |||||
|---|---|---|---|---|---|---|
| Control | I | II | III | p Value | SEM | |
| Initial body weight (g/fish) | 14.02 | 14.26 | 14.14 | 14.14 | 0.216 | 0.029 |
| Final body weight (g/fish) | 70.14 c | 79.16 b | 96.56 a | 95.00 a | <0.001 | 12.88 |
| Final weight gain (g/fish) | 56.12 c | 64.90 b | 82.42 a | 80.86 a | <0.001 | 12.39 |
| Final weight gain (%) | 400.36 c | 455.21 b | 582.72 a | 571.72 a | <0.001 | 526.22 |
| Total feed intake (g/fish) | 96.52 | 103.08 | 99.41 | 100.26 | 0.08 | 12.85 |
| Feed conversion ratio | 1.71 a | 1.58 b | 1.21c | 1.24c | <0.001 | 0.002 |
| Specific growth rate (%) | 1.92 c | 2.04 b | 2.28 a | 2.27 a | <0.001 | 0.002 |
| Protein efficiency ratio | 1.75 b | 1.89 b | 2.50 a | 2.43 a | <0.001 | 0.016 |
| Condition factor | 2.31 c | 2.54 bc | 3.08 ab | 3.36 a | <0.001 | 0.182 |
| Parameters | Supplemental Omega-3-NPs (g/kg Diet) | |||||
|---|---|---|---|---|---|---|
| Control | I | II | III | p Value | SEM | |
| ΣSFAs | 17.23 a | 16.93 ab | 16.27 b | 15.2 c | <0.02 | 0.25 |
| ΣMUSFAs | 25.23 a | 24.3 ab | 23.2 b | 22.1 c | <0.001 | 0.39 |
| Σn − 3 | 5.9 d | 8.31 c | 11.39 b | 16.33 a | <0.04 | 0.47 |
| Σn − 6 | 53.46 a | 45.36 b | 40.22 c | 35.69 d | <0.02 | 0.29 |
| ΣPUFAs | 59.36 ab | 58.79 b | 61.315 a | 61.58 a | <0.008 | 0.24 |
| n6/n3 | 9.06 a | 5.45 b | 3.53 c | 2.19 d | <0.001 | 0.13 |
| MDA (nmol/g tissue) | 21.30 a | 19.40 ab | 19.60 ab | 17.87 b | 0.045 | 1.40 |
| ROS | 112.80 a | 96.20 b | 67.23 c | 56.57 d | <0.001 | 19.91 |
| T-AOC (U/mg prot) | 0.83 d | 1.62 c | 2.27 b | 3.73 a | <0.001 | 0.03 |
| H2O2 (μmoL/g tissue) | 4.73 a | 3.33 b | 2.23 c | 1.69 c | <0.001 | 0.09 |
| Parameters | Supplemental Omega-3-NPs (g/kg Diet) | |||||
|---|---|---|---|---|---|---|
| Control | I | II | III | p Value | SEM | |
| RBCs (×106/μL) | 2.32 | 2.59 | 2.52 | 2.56 | 0.580 | 0.07 |
| Ht (%) | 28.87 | 28.37 | 29.10 | 29.37 | 0.345 | 0.42 |
| Hb (g/dL) | 9.11 | 9.60 | 9.58 | 9.84 | 0.085 | 0.09 |
| Total protein (g/dL) | 3.07 c | 3.13 c | 4.20 b | 5.33a | <0.001 | 0.10 |
| Albumin (g/dL) | 1.99 b | 2.96 a | 2.38 ab | 2.28 b | 0.012 | 0.07 |
| Globulin (g/dL) | 1.07 d | 1.2 c | 1.71 b | 2.50 a | <0.001 | 0.13 |
| ALT (U/L) | 47.17 | 48.37 | 45.23 | 47.20 | 0.578 | 7.23 |
| AST(U/L) | 29.71 ab | 27.71 b | 31.54 a | 28.54 ab | 0.041 | 1.87 |
| Creatinine (mg/dL) | 0.52 | 0.50 | 0.49 | 0.50 | 0.869 | 0.001 |
| Urea (mg/dL) | 5.00 a | 4.70 ab | 4.65 ab | 4.49 b | 0.054 | 0.04 |
| Cholesterol (mg/dL) | 92.53 a | 87.70 ab | 81.25 b | 69.74 c | 0.001 | 16.95 |
| Triacylglycerol (mg/dL) | 71.63 | 69.96 | 73.63 | 66.96 | 0. 241 | 13.92 |
| HDL-cholesterol (mg/dL) | 33.23 c | 42.66 b | 47.13 ab | 52.44 a | <0.001 | 6.93 |
| LDL-cholesterol (mg/dL) | 44.98 a | 31.05 b | 19.39 b | 13.90 c | <0.001 | 33.61 |
| VLDL-cholesterol (mg/dL) | 14.33 | 13.99 | 14.73 | 13.39 | 0.241 | 0.56 |
| IgM (μg/mL) | 24.90 c | 32.63 b | 36.84 ab | 38.74 a | <0.001 | 6.00 |
| Serum lysozyme (μg/mL) | 0.76 c | 1.21 b | 1.37 b | 1.63 a | <0.001 | 0.01 |
| MPO (μmoL/L, OD 450 nm) | 0.53 c | 0.63 c | 0.81 b | 1.07 a | <0.001 | 0. 004 |
| Serum alternative complementary (u/mL) | 215.10 c | 228.33 b | 239.57 a | 250.29 a | <0.001 | 24.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, D.; Arisha, A.H.; Khater, S.I.; Gad, W.M.; Hassan, Z.; Abou-Khadra, S.H.; Mohamed, D.I.; Ahmed Ismail, T.; Gad, S.A.; Eid, S.A.M.; et al. Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus). Antioxidants 2022, 11, 1523. https://doi.org/10.3390/antiox11081523
Ibrahim D, Arisha AH, Khater SI, Gad WM, Hassan Z, Abou-Khadra SH, Mohamed DI, Ahmed Ismail T, Gad SA, Eid SAM, et al. Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus). Antioxidants. 2022; 11(8):1523. https://doi.org/10.3390/antiox11081523
Chicago/Turabian StyleIbrahim, Doaa, Ahmed H. Arisha, Safaa I. Khater, Wafaa M. Gad, Zeinab Hassan, Sally H. Abou-Khadra, Dalia Ibrahim Mohamed, Tamer Ahmed Ismail, Sara A. Gad, Salwa A. M. Eid, and et al. 2022. "Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus)" Antioxidants 11, no. 8: 1523. https://doi.org/10.3390/antiox11081523
APA StyleIbrahim, D., Arisha, A. H., Khater, S. I., Gad, W. M., Hassan, Z., Abou-Khadra, S. H., Mohamed, D. I., Ahmed Ismail, T., Gad, S. A., Eid, S. A. M., Abd El-Wahab, R. A., & Kishawy, A. T. Y. (2022). Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus). Antioxidants, 11(8), 1523. https://doi.org/10.3390/antiox11081523