
Oxidative Stress and Fetal Growth Restriction Set Up Earlier in Undernourished Sheep Twin Pregnancies: Prevention with Antioxidant and Nutritional Supplementation

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Procedure
2.2. Assessment of Maternal and Fetal Oxygenation Status
2.3. Assessment of Oxidative Stress Biomarkers
2.4. Statistical Analysis
3. Results
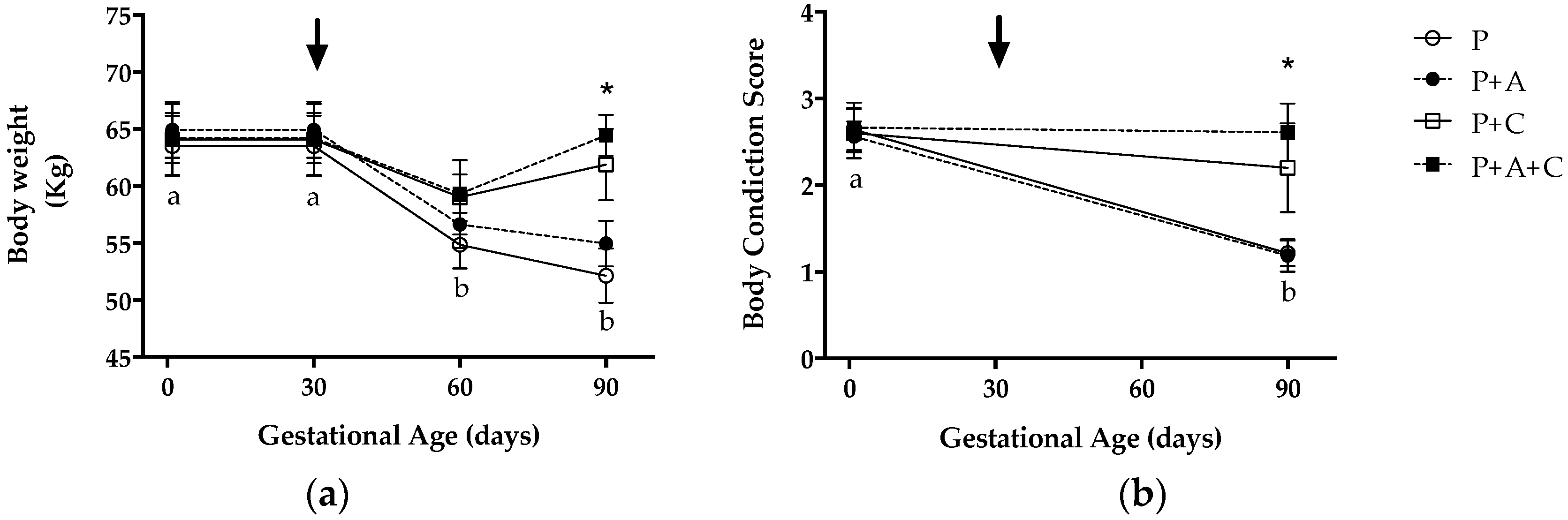
3.1. Maternal Weight and Body Condition
3.2. Effects of Maternal Antioxidant and Nutritional Supplementation on Maternal and Fetal Blood Gases
3.3. Effects of Maternal Antioxidant and Nutritional Supplementation on Fetal Traits
3.3.1. Effects on Fetal Oxidative Status
3.3.2. Effects on Fetal Body Weight and Biometry
3.3.3. Effects on Fetal Semitendinosus Muscle and Main Organs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- González-Bulnes, A.; Parraguez, V.H.; Berlinger, F.; Barbero, A.; García-Contreras, C.; Lopez-Tello, J.; Pesantez-Pacheco, J.L.; Martínez-Ros, P. The impact of prenatal environment on postnatal life and performance: Future perspectives for prevention and treatment. Theriogenology 2020, 150, 15–19. [Google Scholar] [CrossRef] [PubMed]
- McCoard, S.A.; Sales, F.A.; Sciascia, Q.L. Invited review: Impact of specific nutrient interventions during mid-to-late gestation on physiological traits important for survival of multiple-born lambs. Animal 2017, 11, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- McCoard, S. Issues and opportunities to capitalize on increased litter size in hill country sheep farming systems—A New Zealand perspective. Anim. Front. 2017, 7, 32–37. [Google Scholar] [CrossRef]
- Hinch, G.N.; Brien, F. Lamb survival in Australian flocks: A review. Anim. Prod. Sci. 2014, 54, 656–666. [Google Scholar] [CrossRef]
- Skapetas, B.; Kalaitzidou, M. Current Status and Perspectives of Sheep Sector in the World. Livestock Res. Rur. Dev. 2017, 29, 21. Available online: https://www.lrrd.org/lrrd29/2/cont2902.htm (accessed on 28 January 2022).
- Tibbo, M.; Philipsson, J.; Ayalew, W. Sustainable Sheep Breeding Programmes in the Tropics: A Framework for Ethiopia. In Proceedings of the Conference on International Agricultural Research for Development, University of Bonn, Bonn, Germany, 11–13 October 2006; Available online: https://www.researchgate.net/publication/228669959 (accessed on 28 January 2022).
- Sales, F.; Strauch, O. Efecto del tipo de preñez sobre la variación de peso invernal en ovejas Corriedale. In Proceedings of the XXXI Reunión Anual de la Sociedad Chilena de Producción Animal, Centro Regional de Investigación, INIA, Quilamapu, Chillán, Chile, 20 April 2006; pp. 151–152. [Google Scholar]
- Sales, F.; Peralta, O.A.; Narbona, E.; McCoard, S.; De los Reyes, M.; González-Bulnes, A.; Parraguez, V.H. Hypoxia and oxidative stress are associated with reduced fetal growth in twin and undernourished sheep pregnancies. Animals 2018, 8, 217. [Google Scholar] [CrossRef]
- Zambrano, E.; Guzmán, C.; Rodríguez-González, G.L.; Durand-Carvajal, M.; Nathanielsz, P.W. Fetal programming of sexual development and reproductive function. Mol. Cell. Endocrinol. 2014, 382, 538–549. [Google Scholar] [CrossRef]
- McCoard, S.; Koolaard, J.; Charteris, A.; Luo, D. Brief communication: Effect of twinning and sex on carcass weight and composition in lambs. In Proceedings of the New Zealand Society of Animal Production, Leipzig, Germany, 1–6 August 2010; pp. 133–136. [Google Scholar]
- Nash, M.L.; Hungerford, L.L.; Nash, T.G.; Zinn, G.M. Risk factors for perinatal and postnatal mortality in lambs. Vet. Rec. 1996, 139, 64–67. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Cafe, L.M.; Hearnshaw, H.; Hennessy, D.W.; Thompson, J.M.; Morris, S.G. Long-term consequences of birth weight and growth to weaning on carcass, yield and beef quality characteristics of Piedmontese- and Wagyu-sired cattle. Aust. J. Exp. Agric. 2006, 46, 257–269. [Google Scholar] [CrossRef]
- Abbate, J.M.; Macrì, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of protein hydrolysates from anchovy (Engraulis encrasicolus) waste for twelve weeks decreases metabolic dysfunction-associated fatty liver disease severity in ApoE-/-mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef]
- Vazzana, I.; Rizzo, M.; Giambelluca, S.; Zumbo, A.; Piccione, G.; Monteverde, V. The response of some blood constituents after administration of two different diets in goats. Comp. Clin. Pathol. 2014, 23, 1587–1591. [Google Scholar] [CrossRef]
- Avondo, M.; Pagano, R.; Guastella, A.; Criscione, A.; Di Gloria, M.; Valenti, B.; Piccione, G.; Pennisi, P. Diet selection and milk production and composition in Girgentana goats with different αs1-casein genotype. J. Dairy Res. 2009, 76, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Armato, L.; Gianesella, M.; Morgante, M.; Fiore, E.; Rizzo, M.; Giudice, E.; Piccione, G. Rumen volatile fatty acids × dietary supplementation with live yeast and yeast cell wall in feedlot beef cattle. Acta Agric. Scand. 2016, 66, 119–124. [Google Scholar] [CrossRef]
- Monteverde, V.; Congiu, F.; Vazzana, I.; Dara, S.; Di Pietro, S.; Piccione, G. Serum lipid profile modification related to polyunsaturated fatty acid supplementation in thoroughbred horses. J. Appl. Anim. Res. 2017, 45, 615–618. [Google Scholar] [CrossRef]
- Sales, F.; Peralta, O.A.; Narbona, E.; McCoard, S.; Lira, R.; De los Reyes, M.; González-Bulnes, A.; Parraguez, V.H. Maternal supplementation with antioxidant vitamins in sheep results in increased transfer to the fetus and improvement of fetal antioxidant status and development. Antioxidants 2019, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Parraguez, V.H.; Sales, F.; Peralta, O.A.; Narbona, E.; Lira, R.; De los Reyes, M.; González-Bulnes, A. Supplementation of underfed twin-bearing ewes with herbal vitamins C and E: Impacts on birth weight, postnatal growth, and pre-weaning survival of the lambs. Animals 2020, 10, 652. [Google Scholar] [CrossRef]
- Jefferies, B.C. Body condition scoring and its use in management. Tasm. J. Agric. 1961, 32, 19–21. [Google Scholar]
- Van der Linden, D.; Kenyon, P.; Jenkinson, C.M.C.; Peterson, S.; Blair, H.T. Carry-over effects of ewe nutrition and birth rank during the previous pregnancy on the milking performance during the subsequent lactation of Romney ewes. Anim. Prod. Sci. 2011, 51, 102–110. [Google Scholar] [CrossRef]
- Merck Veterinary Manual. 2018. Available online: http://www.merckvetmanual.com (accessed on 20 February 2022).
- Cofré, E.; Peralta, O.A.; Raggi, A.; De los Reyes, M.; Sales, F.; González-Bulnes, A.; Parraguez, V.H. Ram semen deterioration by short-term exposure to high altitude is prevented by improvement of antioxidant status. Animal 2017, 12, 1007–1014. [Google Scholar] [CrossRef]
- Cranston, L.; Kenyon, P.R.; Corner-Thomas, R.A.; Morris, S.T. The potential interaction between ewe body condition score and nutrition during very late pregnancy and lactation on the performance of twin-bearing ewes and their lambs. Asian-Australas. J. Anim. Sci. 2017, 30, 1270–1277. [Google Scholar] [CrossRef]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Narang, M.; Banerjee, B.D.; Basu, S. Oxidative stress in term small for gestational age neonates born to undernourished mothers: A case control study. BMC Pediatr. 2004, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Kamath, U.; Rao, G.; Kamath, S.U.; Rai, L. Maternal and fetal indicators of oxidative stress during intrauterine growth retardation (IUGR). Indian J. Clin. Biochem. 2006, 21, 111–115. [Google Scholar] [CrossRef]
- Li, H.P.; Chen, X.; Li, M.Q. Gestational diabetes induces chronic hypoxia stress and excessive inflammatory response in murine placenta. Int. J. Clin. Exp. Pathol. 2013, 6, 650–659. [Google Scholar] [PubMed]
- Parraguez, V.H.; Atlagich, M.; Araneda, O.; Garcia, C.; Muñoz, A.; De los Reyes, M.; Urquieta, B. Effects of antioxidant vitamins on newborn and placental traits in gestations at high altitude: Comparative study in high and low altitude native sheep. Reprod. Fertil. Dev. 2011, 23, 285–296. [Google Scholar] [CrossRef]
- Parraguez, V.H.; Urquieta, B.; De los Reyes, M.; Gonzalez-Bulnes, A.; Astiz, S.; Muñoz, A. Steroidogenesis in sheep pregnancy with intrauterine growth retardation by high-altitude hypoxia: Effects of maternal altitudinal status and antioxidant treatment. Reprod. Fertil. Dev. 2013, 25, 639–645. [Google Scholar] [CrossRef]
- Symonds, M.E.; Budge, H.; Stephenson, T.; McMillen, I.C. Fetal endocrinology and development-manipulation and adaptation to long-term nutritional and environmental challenges. Reproduction 2001, 121, 853–862. [Google Scholar] [CrossRef]
- Gootwine, E.; Spencer, T.E.; Bazer, F.W. Litter-size-dependent intrauterine growth restriction in sheep. Animal 2007, 1, 547–564. [Google Scholar] [CrossRef]
- Barbero, A.; Porcu, C.; Spezzigu, A.; Succu, S.; Dattena, M.; Gallus, M.; Molle, G.; Naitana, S.; Gonzalez-Bulnes, A.; Berlinguer, F. Changes in renal hemodynamics of undernourished fetuses appear earlier than IUGR evidences. J. Dev. Orig. Health Dis. 2018, 9, 338–343. [Google Scholar] [CrossRef]
- McCoard, S.A.; Peterson, S.W.; McNabb, W.C.; Harris, P.M.; McCutcheon, S.N. Maternal constraint influences muscle fiber development in fetal lambs. Reprod. Fertil. Dev. 1997, 9, 675–681. [Google Scholar] [CrossRef]
- McCoard, S.A.; McNabb, W.C.; Peterson, S.W.; McCutcheon, S.N.; Harris, P.M. Muscle growth, cell number, type and morphometry in single and twin fetal lambs during mid to late gestation. Reprod. Fertil. Dev. 2000, 12, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Thakor, A.S.; Herrera, E.; Giussani, D.A.; Serón-Ferré, M. Melatonin and vitamin C increase umbilical blood flow via nitric oxide-dependent mechanisms. J. Pineal Res. 2010, 49, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, R.; Kasimanickam, V.; Rodriguez, J.S.; Pelzer, K.; Sponenberg, P.; Thatcher, C.D. Tocopherol induced angiogenesis in placental vascular network in late pregnant ewes. Reprod. Biol. Endocrinol. 2010, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Richter, H.G.; Hansell, J.A.; Raut, S.; Giussani, D.A. Melatonin improves placental efficiency and birth weight and increases the placental expression of antioxidant enzymes in undernourished pregnancy. J. Pineal Res. 2009, 46, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Osgerby, J.C.; Wathes, D.C.; Howard, D.; Gadd, T.S. The effect of maternal undernutrition on the placental growth trajectory and the uterine insulin-like growth factor axis in the pregnant ewe. J. Endocrinol. 2004, 182, 89–103. [Google Scholar] [CrossRef][Green Version]



{kind=link}
{kind=link}
| Group | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Blood Trait | P | P+A | P+C | P+A+C | AOX | FP | AOXxNP |
| PO2 (mm Hg) | 17.4 ± 0.9 | 22.5 ± 2.3 | 21.2 ± 2.1 | 21.7 ± 1.7 | 0.071 | ns | ns |
| PCO2 (mm Hg) | 57.9 ± 1.4 | 55.5 ± 2.2 | 45.6 ± 2.4 | 48.8 ± 1.6 | ns | <0.001 | ns |
| SatHb (%) | 18.8 ± 1.8 | 29.6 ± 4.4 | 28.3 ± 4.8 | 29.7 ± 4.2 | 0.066 | ns | ns |
| Ht (%) | 36.3 ± 1.2 | 37.8 ± 1.0 | 35.9 ± 0.8 | 37.2 ± 0.8 | 0.060 | ns | ns |
| pH | 7.33 ± 0.02 | 7.31 ± 0.02 | 7.34 ± 0.02 | 7.36 ± 0.02 | ns | 0.043 | ns |
| Group | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Blood Trait | P | P+A | P+C | P+A+C | AOX | FP | AOXxNP |
| BW (g) | 869 ± 28 c | 915 ± 26 bc | 976 ± 33 ab | 1050 ± 35 a | 0.068 | <0.001 | ns |
| CRL (cm) | 26.2 ± 0.4 b | 26.6 ± 0.3 b | 27.0 ± 0.5 ab | 27.8 ± 0.4 a | ns | 0.016 | ns |
| TP (cm) | 20.1 ± 0.3 b | 20.4 ± 0.2 b | 20.8 ± 0.3 ab | 21.3 ± 0.3 a | ns | 0.008 | ns |
| FL (cm) | 16.3 ± 0.3 b | 16.4 ± 0.3 b | 17.2 ± 0.4 a | 17.2 ± 0.3 a | ns | 0.055 | ns |
| HL (cm) | 18.7 ± 0.3 c | 18.9 ± 0.2 bc | 19.7 ± 0.5 ab | 19.8 ± 0.3 a | ns | 0.003 | ns |
| Group | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Fetal Trait | P | P+A | P+C | P+A+C | AOX | FP | AOXxNP |
| SM (g) | 1.80 ± 0.06 c | 1.91 ± 0.1 bc | 2.08 ± 0.07 ab | 2.19 ± 0.08 a | ns | 0.003 | ns |
| Kidneys (g) | 4.80 ± 0.13 b | 4.96 ± 0.14 b | 5.14 ± 0.14 ab | 5.43 ± 0.16 a | ns | 0.009 | ns |
| BAT (g) | 2.33 ± 0.20 bc | 2.09 ± 0.15 c | 2.72 ± 0.20 b | 3.34 ± 0.23 a | ns | <0.001 | ns |
| Heart (g) | 7.98 ± 0.32 ab | 7.58 ± 0.32 b | 8.78 ± 0.53 a | 8.69 ± 0.23 a | ns | 0.007 | ns |
| Brain (g) | 23.5 ± 0.76 | 23.2 ± 0.53 | 22.3 ± 0.70 | 23.5 ± 0.66 | ns | ns | ns |
| Liver (g) | 45.1 ± 1.74 bc | 42.8 ± 1.61 c | 50.1 ± 1.88 b | 56.4 ± 1.79 a | ns | <0.001 | ns |
| Brain/Liver | 0.53 ± 0.02 a | 0.55 ± 0.03 a | 0.45 ± 0.02 b | 0.42 ± 0.01 b | ns | <0.001 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parraguez, V.H.; Sales, F.; Peralta, O.; De los Reyes, M.; Gonzalez-Bulnes, A. Oxidative Stress and Fetal Growth Restriction Set Up Earlier in Undernourished Sheep Twin Pregnancies: Prevention with Antioxidant and Nutritional Supplementation. Antioxidants 2022, 11, 1287. https://doi.org/10.3390/antiox11071287
Parraguez VH, Sales F, Peralta O, De los Reyes M, Gonzalez-Bulnes A. Oxidative Stress and Fetal Growth Restriction Set Up Earlier in Undernourished Sheep Twin Pregnancies: Prevention with Antioxidant and Nutritional Supplementation. Antioxidants. 2022; 11(7):1287. https://doi.org/10.3390/antiox11071287
Chicago/Turabian StyleParraguez, Víctor H., Francisco Sales, Oscar Peralta, Monica De los Reyes, and Antonio Gonzalez-Bulnes. 2022. "Oxidative Stress and Fetal Growth Restriction Set Up Earlier in Undernourished Sheep Twin Pregnancies: Prevention with Antioxidant and Nutritional Supplementation" Antioxidants 11, no. 7: 1287. https://doi.org/10.3390/antiox11071287
APA StyleParraguez, V. H., Sales, F., Peralta, O., De los Reyes, M., & Gonzalez-Bulnes, A. (2022). Oxidative Stress and Fetal Growth Restriction Set Up Earlier in Undernourished Sheep Twin Pregnancies: Prevention with Antioxidant and Nutritional Supplementation. Antioxidants, 11(7), 1287. https://doi.org/10.3390/antiox11071287