
Alanyl-Glutamine Protects against Lipopolysaccharide-Induced Liver Injury in Mice via Alleviating Oxidative Stress, Inhibiting Inflammation, and Regulating Autophagy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animal Experiments
2.3. Histological Studies and IHC Analysis
2.4. TUNEL Assay
2.5. Dihydroethidium (DHE) Staining
2.6. Plasma Biochemistry and ELISAs
2.7. Western Blot Analysis
2.8. Real-Time PCR
2.9. Cell Culture and Treatment
2.10. Statistical Analysis
3. Results
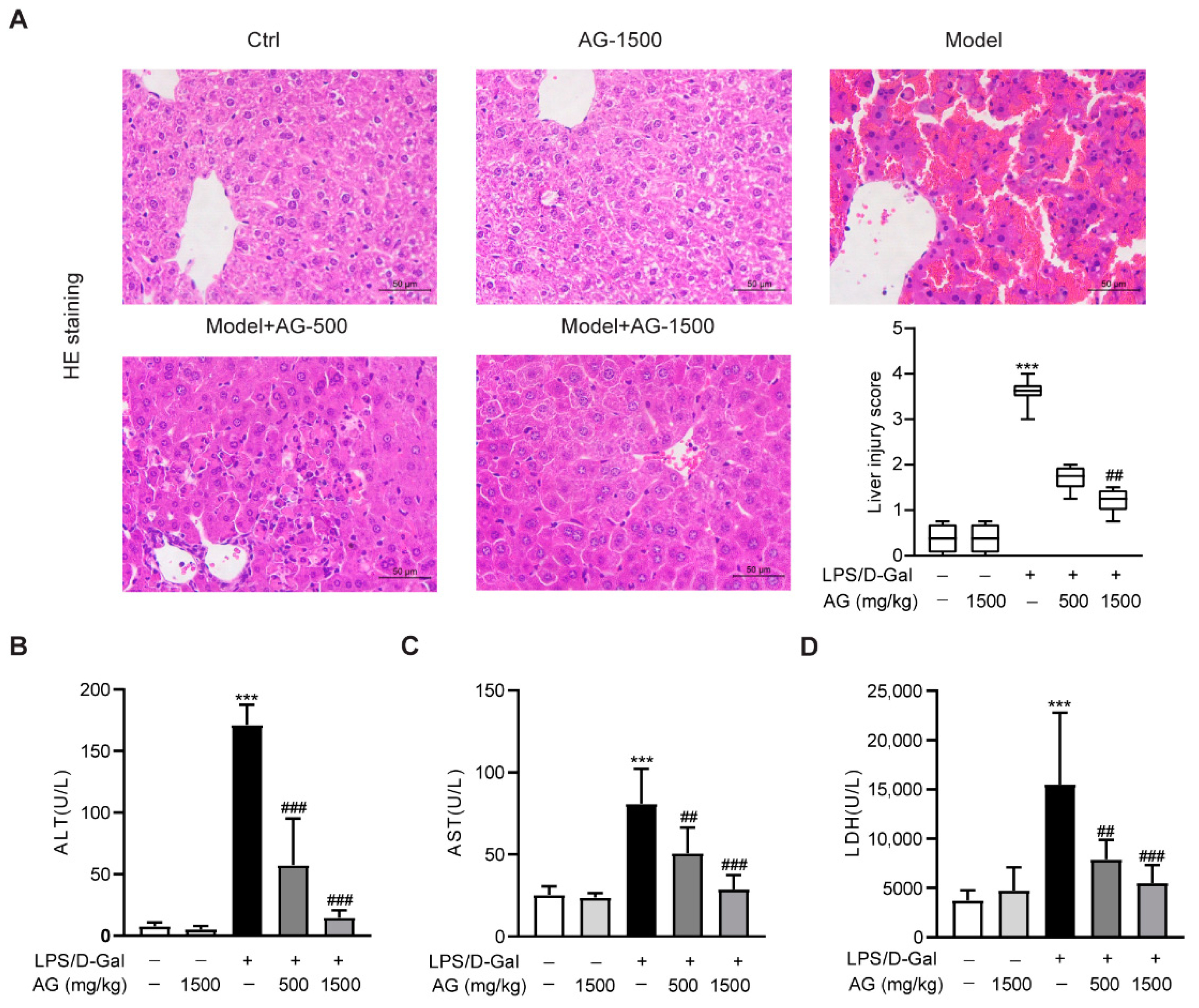
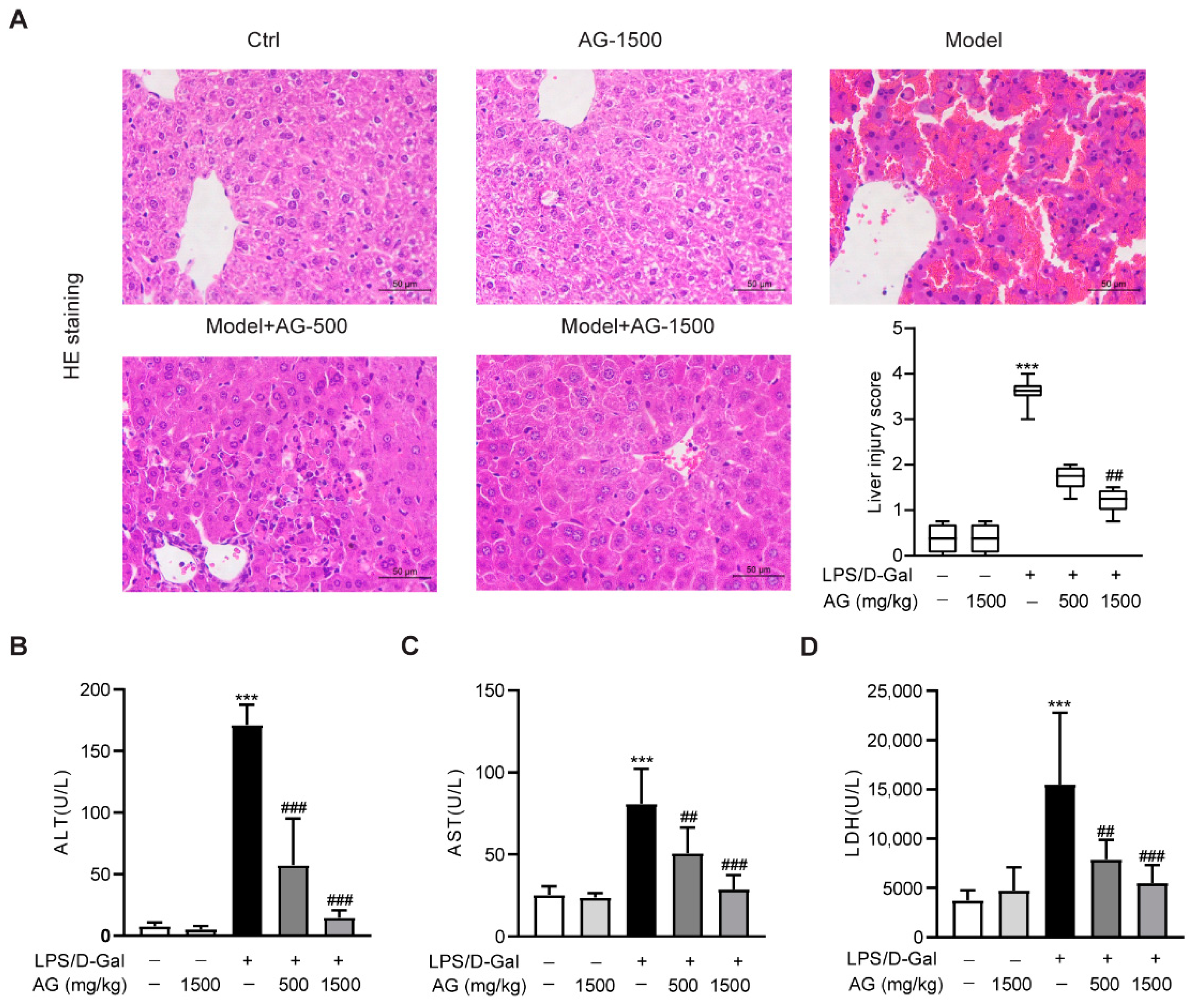
3.1. Ala-Gln Pretreatment Alleviates Acute Liver Injury in LPS/D-Gal-Treated Mice
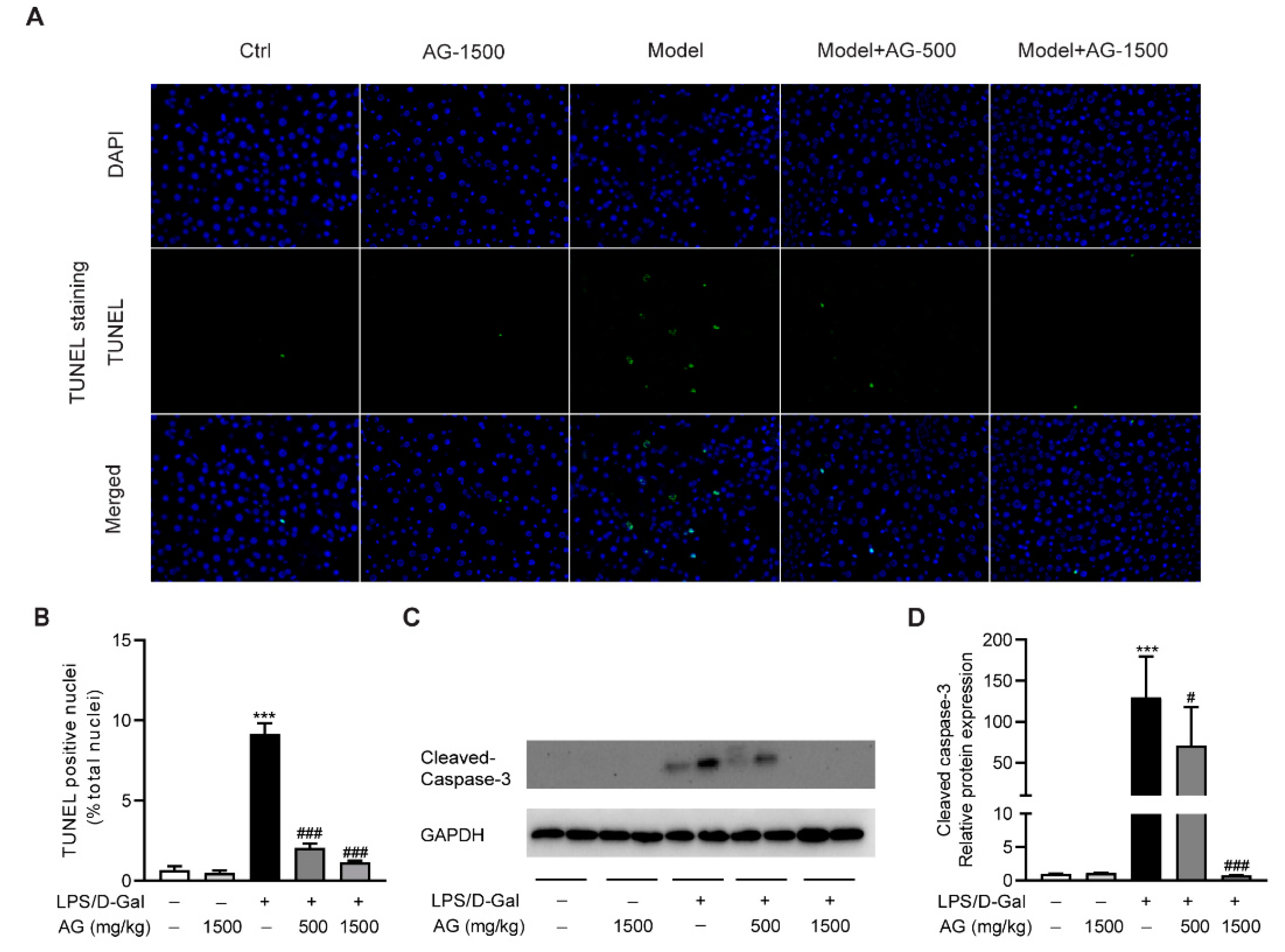
3.2. The Protective Effect of Ala-Gln on LPS/D-Gal-Induced Apoptosis in Mice
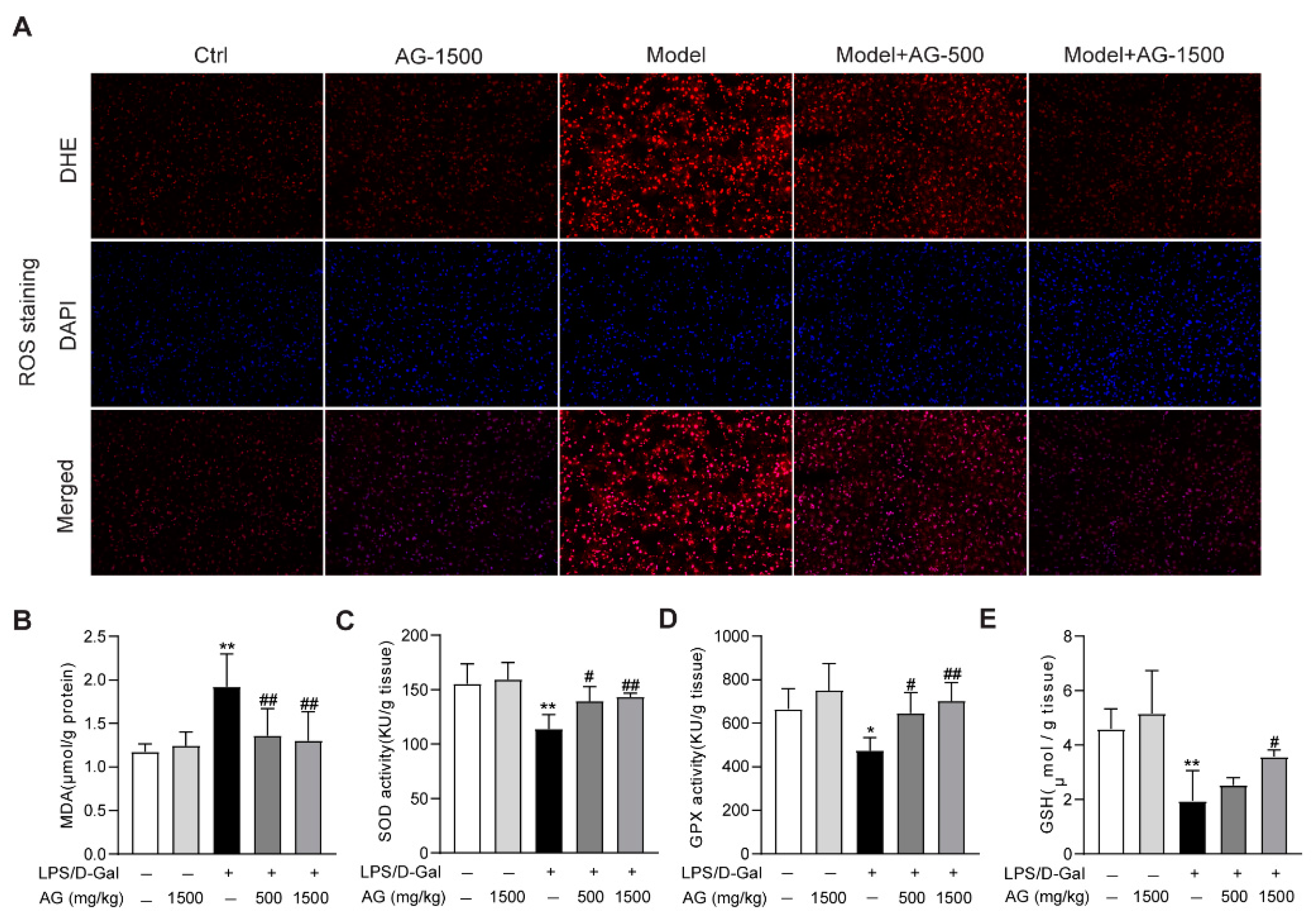
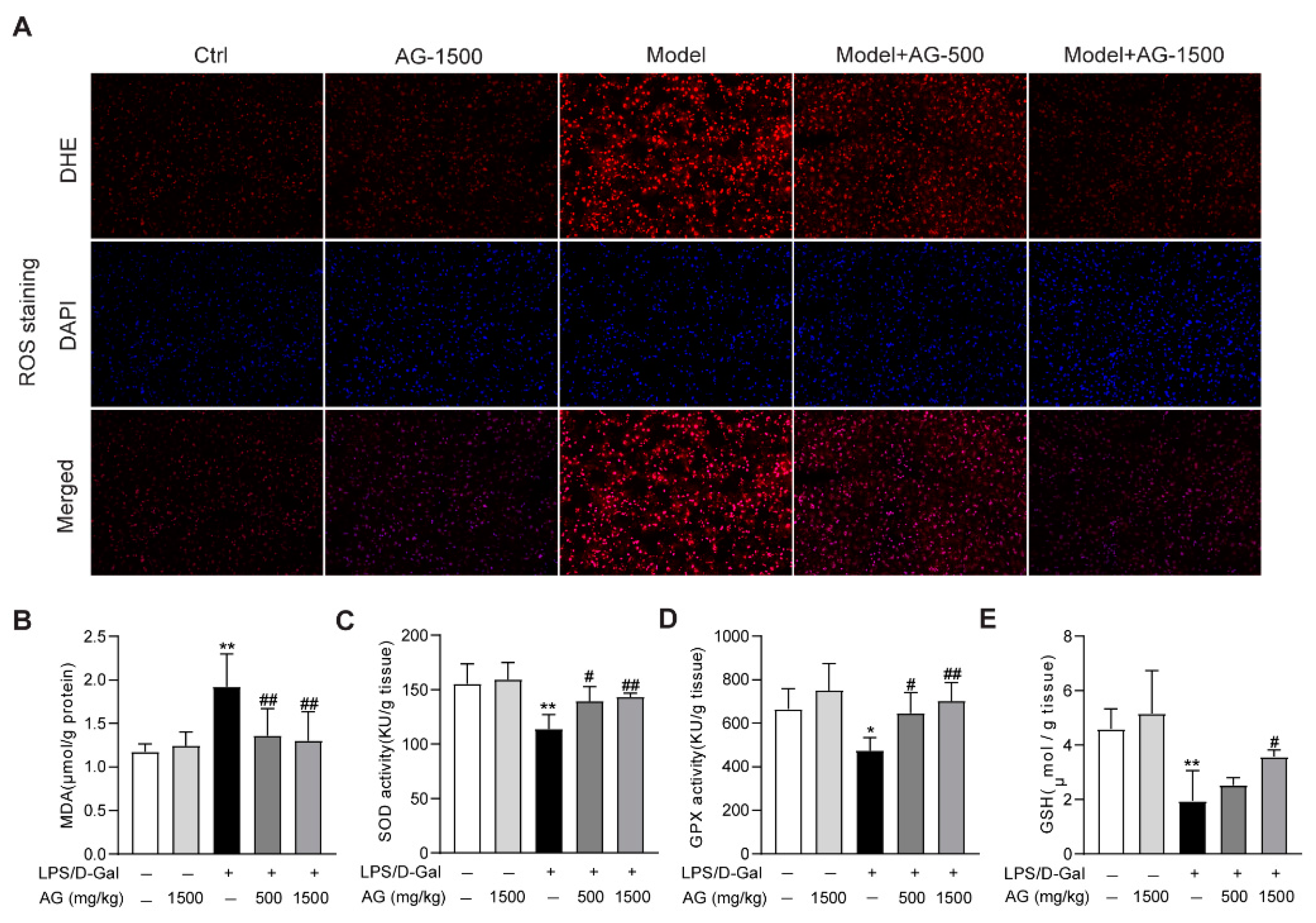
3.3. Effect of Ala-Gln on Liver Antioxidant Capacity
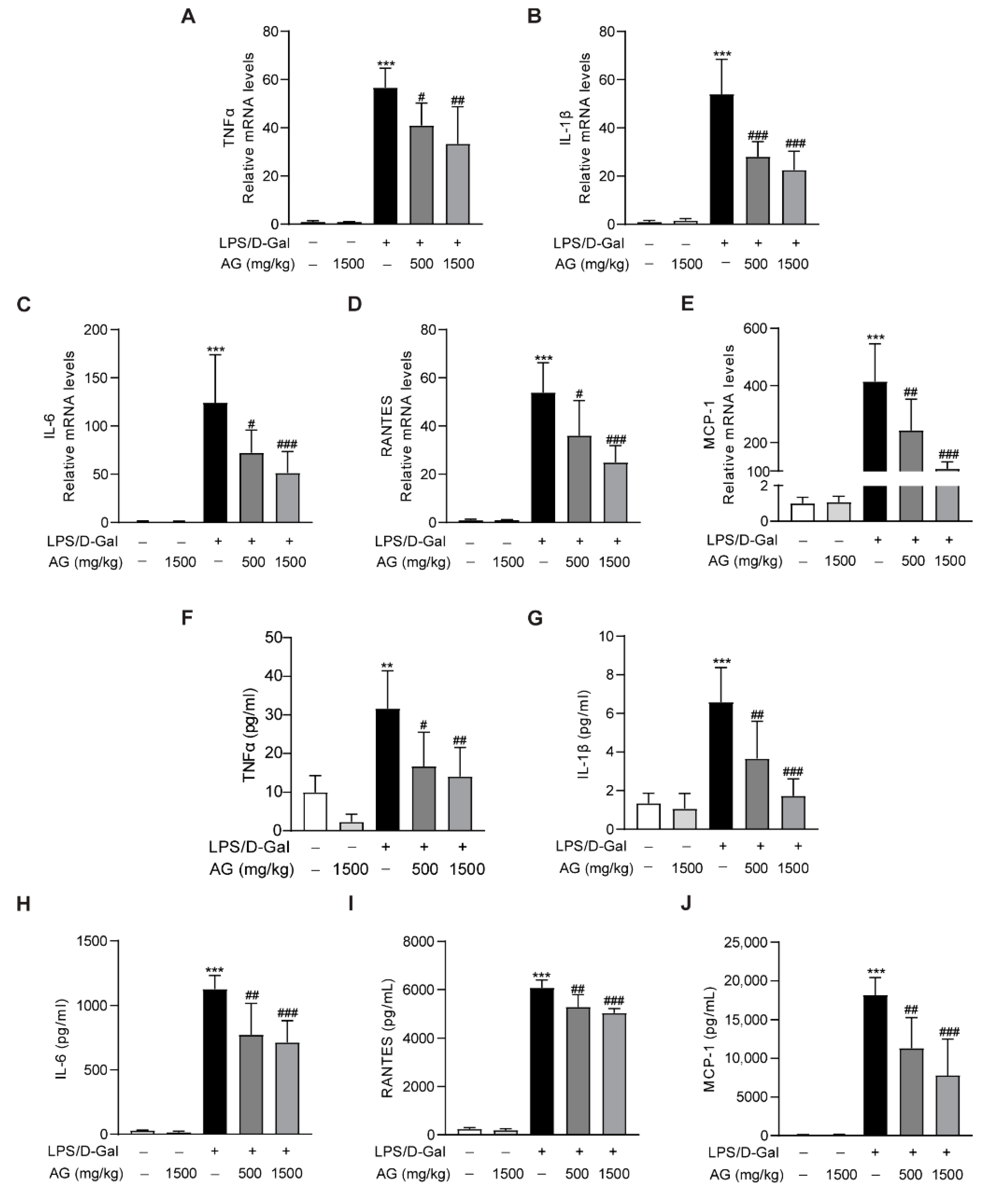
3.4. Ala-Gln Regulates the Distribution of Hepatic F4/80- and CD68-Positive Immune Cells in the Liver and Attenuates Inflammation
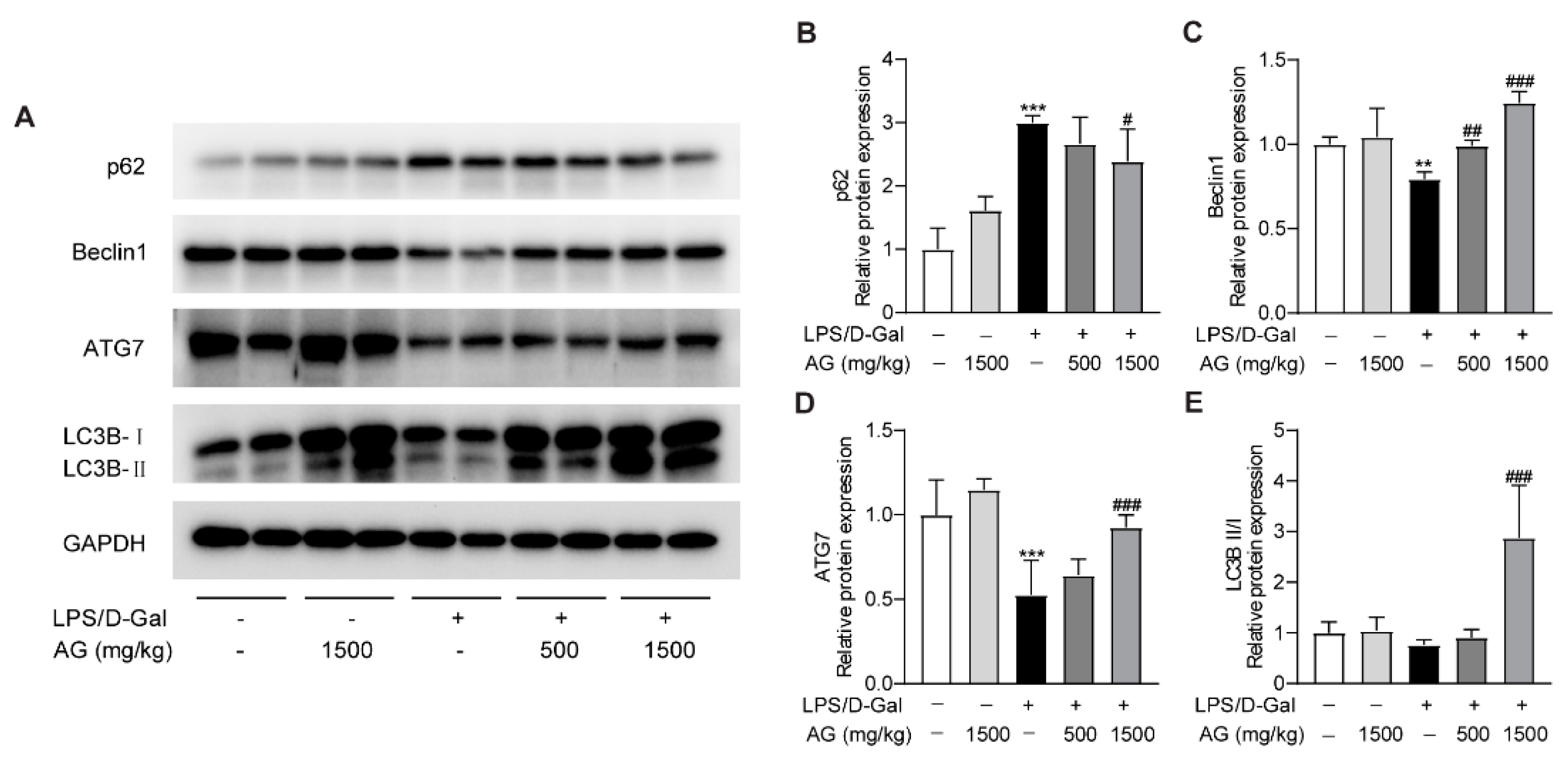
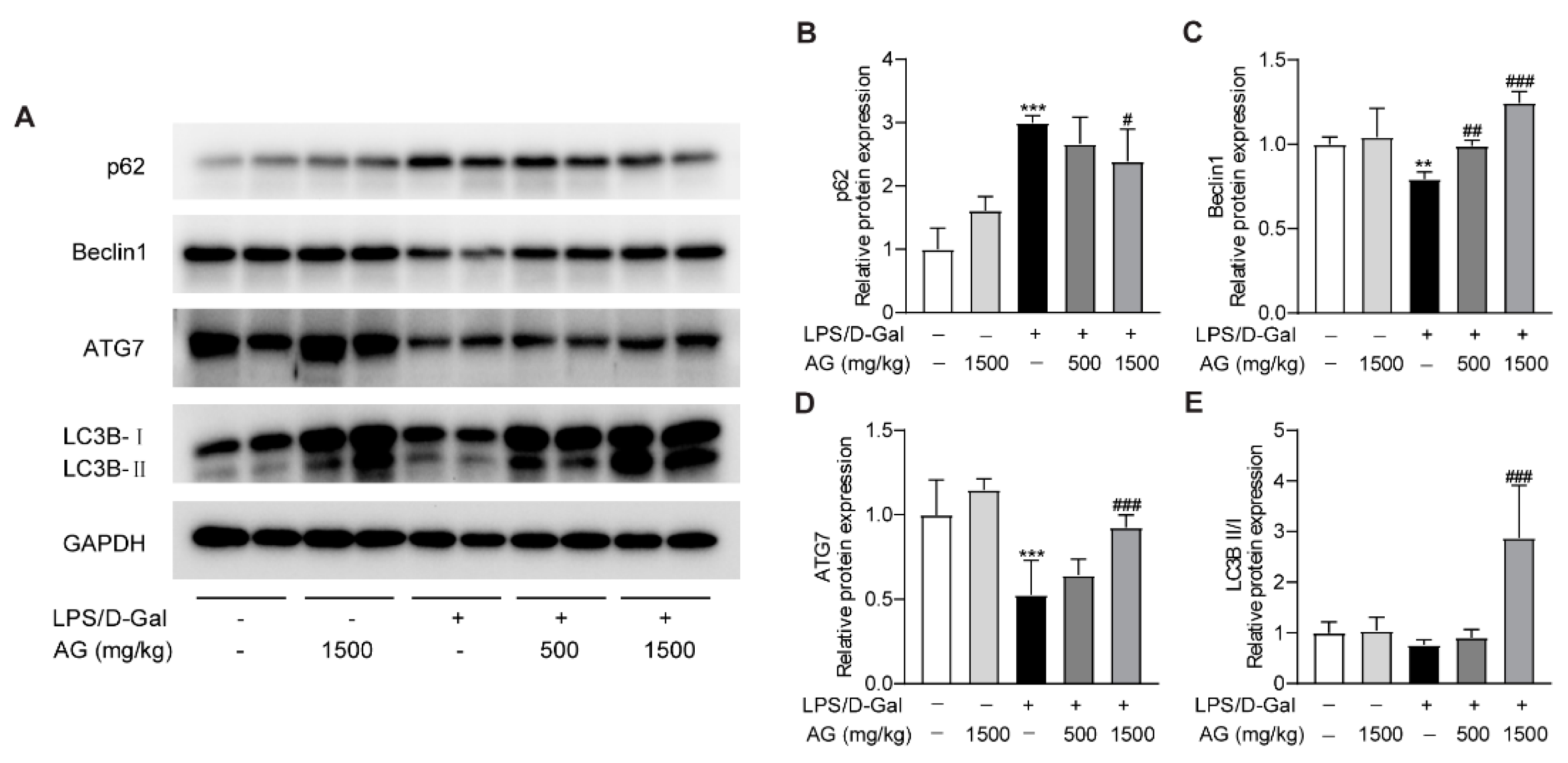
3.5. Ala-Gln Promotes Autophagy in the Livers of LPS/D-Gal-Treated Mice
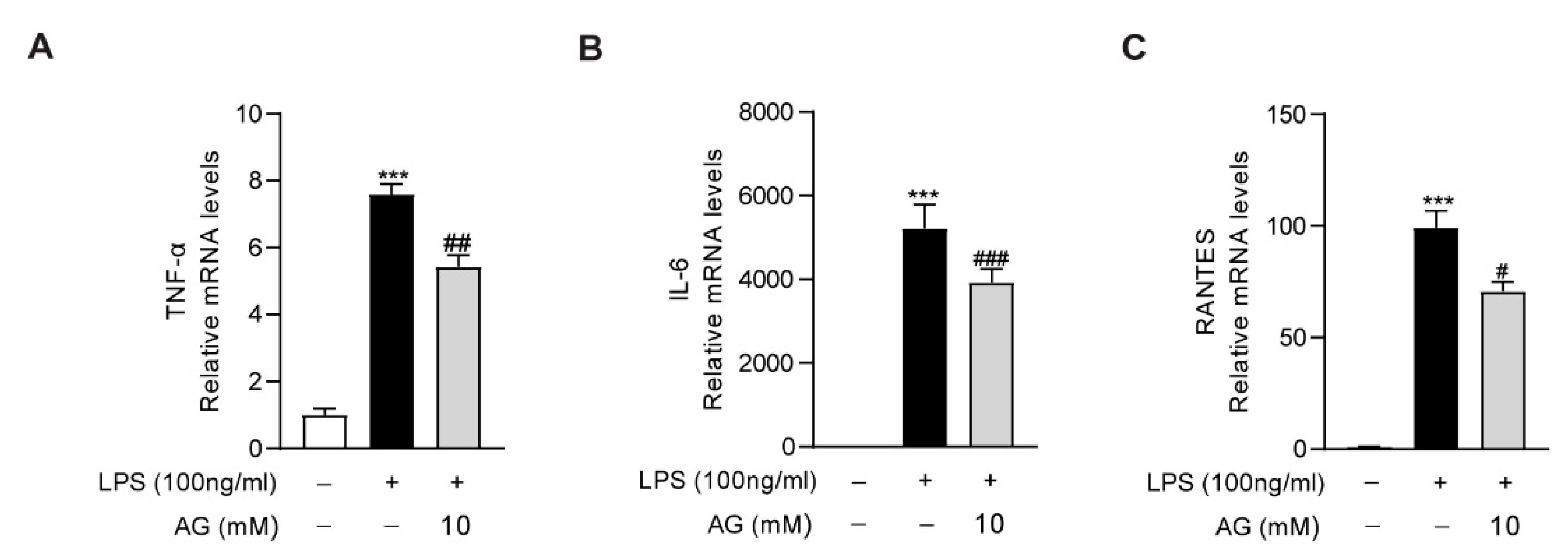
3.6. Ala-Gln Inhibits LPS-Induced Inflammatory Response in RAW264.7 Cells
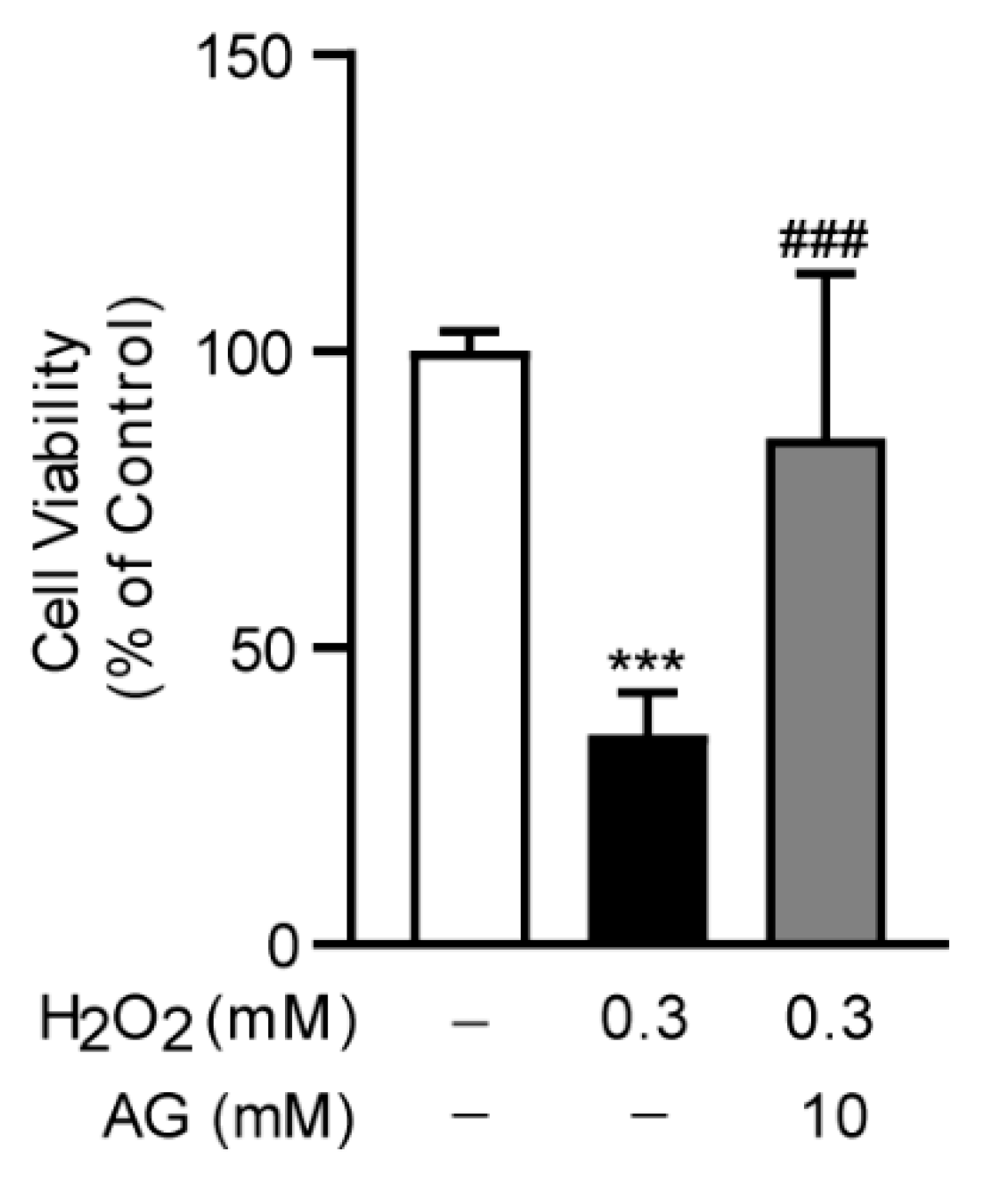
3.7. Ala-Gln Suppresses H2O2-Induced Oxidative Stress in AML-12 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, X.; Davis, A.M.; Parker, W.F. Managing Adult Acute and Acute-on-Chronic Liver Failure in the ICU. JAMA 2021, 326, 1964–1965. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yang, Y.; Wen, Y.; Jeong, J.M.; Emontzpohl, C.; Atkins, C.L.; Sun, Z.; Poulsen, K.L.; Hall, D.R.; Steve Bynon, J.; et al. Hepatic recruitment of eosinophils and their protective function during acute liver injury. J. Hepatol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Jaeschke, H. Oxidative Stress and Acute Hepatic Injury. Curr. Opin. Toxicol. 2018, 7, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Ji, X.; Chen, G.; An, B. Atractylodin ameliorates lipopolysaccharide and d-galactosamine-induced acute liver failure via the suppression of inflammation and oxidative stress. Int. Immunopharmacol. 2019, 72, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Zhou, Z.; Lim, C.W.; Kim, J.W.; Kim, B. Amlexanox ameliorates acetaminophen-induced acute liver injury by reducing oxidative stress in mice. Toxicol. Appl. Pharmacol. 2019, 385, 114767. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Liu, D.; Chen, S.; Li, X.; Ge, M.; Huang, W. Sevoflurane preconditioning activates HGF/Met-mediated autophagy to attenuate hepatic ischemia-reperfusion injury in mice. Cell Signal. 2021, 82, 109966. [Google Scholar] [CrossRef]
- Allaire, M.; Rautou, P.E.; Codogno, P.; Lotersztajn, S. Autophagy in liver diseases: Time for translation? J. Hepatol. 2019, 70, 985–998. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Yang, H.; Wang, Z.; Feng, H.; Deng, X.; Cheng, G.; Ci, X. Nrf2 signaling and autophagy are complementary in protecting lipopolysaccharide/d-galactosamine-induced acute liver injury by licochalcone A. Cell Death Dis. 2019, 10, 313. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.N.; Kim, S.H.; Dey, D.K.; Park, S.M.; Nasif, O.; Bajpai, V.K.; Kang, S.C.; Lee, J.; Park, J.G. 5-O-Demethylnobiletin Alleviates CCl4-Induced Acute Liver Injury by Equilibrating ROS-Mediated Apoptosis and Autophagy Induction. Int. J. Mol. Sci. 2021, 22, 1083. [Google Scholar] [CrossRef]
- Han, Y.H.; Onufer, E.J.; Huang, L.H.; Sprung, R.W.; Davidson, W.S.; Czepielewski, R.S.; Wohltmann, M.; Sorci-Thomas, M.G.; Warner, B.W.; Randolph, G.J. Enterically derived high-density lipoprotein restrains liver injury through the portal vein. Science 2021, 373, eabe6729. [Google Scholar] [CrossRef]
- Hu, J.; Zhu, Z.; Ying, H.; Yao, J.; Ma, H.; Li, L.; Zhao, Y. Oleoylethanolamide Protects Against Acute Liver Injury by Regulating Nrf-2/HO-1 and NLRP3 Pathways in Mice. Front. Pharmacol. 2020, 11, 605065. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.N.; Tan, Y.; Xiao, X.C.; Li, Q.; Wu, Q.; Peng, Y.Y.; Ren, J.; Dong, M.L. Deletion of TLR4 attenuates lipopolysaccharide-induced acute liver injury by inhibiting inflammation and apoptosis. Acta Pharmacol. Sin. 2021, 42, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Stegen, S.; Rinaldi, G.; Loopmans, S.; Stockmans, I.; Moermans, K.; Thienpont, B.; Fendt, S.M.; Carmeliet, P.; Carmeliet, G. Glutamine Metabolism Controls Chondrocyte Identity and Function. Dev. Cell 2020, 53, 530–544.e538. [Google Scholar] [CrossRef] [PubMed]
- Hasani, M.; Mansour, A.; Asayesh, H.; Djalalinia, S.; Mahdavi Gorabi, A.; Ochi, F.; Qorbani, M. Effect of glutamine supplementation on cardiometabolic risk factors and inflammatory markers: A systematic review and meta-analysis. BMC Cardiovasc. Disord. 2021, 21, 190. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, W.; Wang, B.; Liu, Q.; Zhong, X.; Gao, Q.; Liu, J.; Huang, J.; Lin, B.; Tao, Y. Metabolic engineering of Escherichia coli for efficient production of L-alanyl-L-glutamine. Microb. Cell Factories 2020, 19, 129. [Google Scholar] [CrossRef]
- Jin, M.; Wang, Y.; Wang, Y.; Li, Y.; Wang, G.; Liu, X.; Xue, Y.; Liu, Z.; Li, C. Protective Effects Oncorneal Endothelium During Intracameral Irrigation Using N-(2)-l-alanyl-l-Glutamine. Front. Pharmacol. 2020, 11, 369. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Santos, R.; da Silva Cardoso, G.; da Costa Lima, L.; de Sousa Cavalcante, M.L.; Silva, M.S.; Cavalcante, A.K.M.; Severo, J.S.; de Melo Sousa, F.B.; Pacheco, G.; Alves, E.H.P.; et al. L-Glutamine and Physical Exercise Prevent Intestinal Inflammation and Oxidative Stress Without Improving Gastric Dysmotility in Rats with Ulcerative Colitis. Inflammation 2021, 44, 617–632. [Google Scholar] [CrossRef]
- Luo, L.L.; Li, Y.F.; Shan, H.M.; Wang, L.P.; Yuan, F.; Ma, Y.Y.; Li, W.L.; He, T.T.; Wang, Y.Y.; Qu, M.J.; et al. L-glutamine protects mouse brain from ischemic injury via up-regulating heat shock protein 70. CNS Neurosci. Ther. 2019, 25, 1030–1041. [Google Scholar] [CrossRef] [Green Version]
- Araujo Junior, R.J.; Silva Junior, R.G.; Vasconcelos, M.P.; Guimaraes, S.B.; Vasconcelos, P.R.; Garcia, J.H. Preconditioning with L-alanyl-glutamine reduces hepatic ischemia-reperfusion injury in rats. Acta Cir. Bras. 2011, 26 (Suppl. 1), 8–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yang, Y.; Song, Y.Q.; Geng, J.; Chen, Q.L. Protective effects of N(2)LalanylLglutamine mediated by the JAK2/STAT3 signaling pathway on myocardial ischemia reperfusion. Mol. Med. Rep. 2018, 17, 5102–5108. [Google Scholar] [CrossRef]
- Jia, C.J.; Dai, C.L.; Zhang, X.; Cui, K.; Xu, F.; Xu, Y.Q. Alanyl-glutamine dipeptide inhibits hepatic ischemia-reperfusion injury in rats. World J. Gastroenterol. 2006, 12, 1373–1378. [Google Scholar] [CrossRef] [PubMed]
- Szijarto, A.; Hahn, O.; Batmunkh, E.; Stangl, R.; Kiss, A.; Lotz, G.; Schaff, Z.; Vali, L.; Blazovics, A.; Gero, D.; et al. Short-term alanyl-glutamine dipeptide pretreatment in liver ischemia-reperfusion model: Effects on microcirculation and antioxidant status in rats. Clin. Nutr. 2007, 26, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Schuster, H.; Blanc, M.C.; Bonnefont-Rousselot, D.; Nakib, S.; Le Tourneau, A.; Furst, P.; Cynober, L.; De Bandt, J.P. Protective effects of glutamine dipeptide and alpha-tocopherol against ischemia-reperfusion injury in the isolated rat liver. Clin. Nutr. 2009, 28, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Chuang, I.C.; Huang, M.S.; Huang, L.J.; Chou, S.H.; Tsai, T.N.; Chen, Y.C.; Yang, R.C. Prophylactic inhalation of L-alanyl-L-glutamine enhances heat shock protein 72 and attenuates endotoxin-induced lung injury in rats. Physiol. Res. 2015, 64, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Huang, C.; Liu, Y.; Lin, D.; Zhao, Y. NMR-based metabolomic analysis of the effects of alanyl-glutamine supplementation on C2C12 myoblasts injured by energy deprivation. RSC Adv. 2018, 8, 16114–16125. [Google Scholar] [CrossRef] [Green Version]
- Pisano, C.A.; Brugnoli, A.; Novello, S.; Caccia, C.; Keywood, C.; Melloni, E.; Vailati, S.; Padoani, G.; Morari, M. Safinamide inhibits in vivo glutamate release in a rat model of Parkinson’s disease. Neuropharmacology 2020, 167, 108006. [Google Scholar] [CrossRef]
- Pengrattanachot, N.; Cherngwelling, R.; Jaikumkao, K.; Pongchaidecha, A.; Thongnak, L.; Swe, M.T.; Chatsudthipong, V.; Lungkaphin, A. Atorvastatin attenuates obese-induced kidney injury and impaired renal organic anion transporter 3 function through inhibition of oxidative stress and inflammation. Biochim. Biophys. Acta Mol. Basis. Dis. 2020, 1866, 165741. [Google Scholar] [CrossRef]
- Yang, F.; Wang, S.; Liu, Y.; Zhou, Y.; Shang, L.; Feng, M.; Yuan, X.; Zhu, W.; Shi, X. IRE1alpha aggravates ischemia reperfusion injury of fatty liver by regulating phenotypic transformation of kupffer cells. Free Radic. Biol. Med. 2018, 124, 395–407. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, B.; Tang, L.; Zhou, Y.; Wang, Q.; Gong, L.; Ni, J.; Li, W. Probiotic Bacillus Alleviates Oxidative Stress-Induced Liver Injury by Modulating Gut-Liver Axis in a Rat Model. Antioxidants 2022, 11, 291. [Google Scholar] [CrossRef]
- Hu, J.; Ying, H.; Yao, J.; Yang, L.; Jin, W.; Ma, H.; Li, L.; Zhao, Y. Micronized Palmitoylethanolamide Ameliorates Methionine- and Choline-Deficient Diet-Induced Nonalcoholic Steatohepatitis via Inhibiting Inflammation and Restoring Autophagy. Front. Pharmacol. 2021, 12, 744483. [Google Scholar] [CrossRef]
- Ge, J.; Hao, R.; Rong, X.; Dou, Q.P.; Tan, X.; Li, G.; Li, F.; Li, D. Secoisolariciresinol diglucoside mitigates benzo[a]pyrene-induced liver and kidney toxicity in mice via miR-101a/MKP-1-mediated p38 and ERK pathway. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2022, 159, 112733. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Yang, L.; Yi, Z.; Fang, H.; Chen, W.; Hong, Z.; Zhang, Y.; Zhang, G.; Li, L. Anti-Inflammatory Effects of Fucoxanthinol in LPS-Induced RAW264.7 Cells through the NAAA-PEA Pathway. Mar. Drugs 2020, 18, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K. Molecular mechanisms of hepatic apoptosis. Cell Death Dis. 2014, 5, e996. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Ju, C. Hepatic Macrophages in Liver Injury. Front. Immunol. 2020, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, G.; Zhao, E.; Liu, K.; Lin, Y.; Tesfa, L.; Tanaka, K.E.; Czaja, M.J. Macrophage autophagy limits acute toxic liver injury in mice through down regulation of interleukin-1beta. J. Hepatol. 2016, 64, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yin, H.; Zhao, Y.; Zhang, X.; Duan, C.; Liu, J.; Huang, C.; Liu, S.; Yang, S.; Li, X. Protective role of puerarin on LPS/D-Gal induced acute liver injury via restoring autophagy. Am. J. Transl. Res. 2018, 10, 957–965. [Google Scholar]
- Schwabe, R.F.; Luedde, T. Apoptosis and necroptosis in the liver: A matter of life and death. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 738–752. [Google Scholar] [CrossRef]
- Huang, S.; Wang, Y.; Xie, S.; Lai, Y.; Mo, C.; Zeng, T.; Kuang, S.; Deng, G.; Zhou, C.; Chen, Y.; et al. Hepatic TGFbetar1 Deficiency Attenuates Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure Through Inhibiting GSK3beta-Nrf2-Mediated Hepatocyte Apoptosis and Ferroptosis. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1649–1672. [Google Scholar] [CrossRef]
- El-Lekawy, A.M.; Abdallah, D.M.; El-Abhar, H.S. Alanyl-glutamine Heals Indomethacin-induced Gastric Ulceration in Rats Via Antisecretory and Anti-apoptotic Mechanisms. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 710–718. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Petry, E.R.; Dresch, D.F.; Carvalho, C.; Medeiros, P.C.; Rosa, T.G.; de Oliveira, C.M.; Martins, L.A.M.; Schemitt, E.; Bona, S.; Guma, F.C.R.; et al. Oral glutamine supplementation attenuates inflammation and oxidative stress-mediated skeletal muscle protein content degradation in immobilized rats: Role of 70kDa heat shock protein. Free Radic. Biol. Med. 2019, 145, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Je, J.Y.; Cho, Y.S.; Gong, M.; Udenigwe, C.C. Dipeptide Phe-Cys derived from in silico thermolysin-hydrolysed RuBisCO large subunit suppresses oxidative stress in cultured human hepatocytes. Food Chem. 2015, 171, 287–291. [Google Scholar] [CrossRef]
- Barros, M.A.; Vasconcelos, P.R.; Souza, C.M.; Andrade, G.M.; Moraes, M.O.; Costa, P.E.; Coelho, G.R.; Garcia, J.H. L-Alanyl-Glutamine Attenuates Oxidative Stress in Liver Transplantation Patients. Transplant. Proc. 2015, 47, 2478–2482. [Google Scholar] [CrossRef] [PubMed]
- Wiesenhofer, F.M.; Herzog, R.; Boehm, M.; Wagner, A.; Unterwurzacher, M.; Kasper, D.C.; Alper, S.L.; Vychytil, A.; Aufricht, C.; Kratochwill, K. Targeted Metabolomic Profiling of Peritoneal Dialysis Effluents Shows Anti-oxidative Capacity of Alanyl-Glutamine. Front. Physiol. 2018, 9, 1961. [Google Scholar] [CrossRef] [Green Version]
- Dworzanski, J.; Strycharz-Dudziak, M.; Kliszczewska, E.; Kielczykowska, M.; Dworzanska, A.; Drop, B.; Polz-Dacewicz, M. Glutathione peroxidase (GPx) and superoxide dismutase (SOD) activity in patients with diabetes mellitus type 2 infected with Epstein-Barr virus. PLoS ONE 2020, 15, e0230374. [Google Scholar] [CrossRef] [PubMed]
- Petry, E.R.; Cruzat, V.F.; Heck, T.G.; de Bittencourt, P.I.H., Jr.; Tirapegui, J. L-glutamine supplementations enhance liver glutamine-glutathione axis and heat shock factor-1 expression in endurance-exercise trained rats. Int. J. Sport Nutr. Exerc. Metab. 2015, 25, 188–197. [Google Scholar] [CrossRef]
- Wen, Y.; Lambrecht, J.; Ju, C.; Tacke, F. Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell. Mol. Immunol. 2021, 18, 45–56. [Google Scholar] [CrossRef]
- Li, R.; Yang, W.; Yin, Y.; Zhang, P.; Wang, Y.; Tao, K. Protective Role of 4-Octyl Itaconate in Murine LPS/D-GalN-Induced Acute Liver Failure via Inhibiting Inflammation, Oxidative Stress, and Apoptosis. Oxid. Med. Cell. Longev. 2021, 2021, 9932099. [Google Scholar] [CrossRef]
- Oliveira, L.R.A.; Albuquerque, A.O.; Silva, C.; Silva, J.M.; Casadevall, M.; Azevedo, O.G.R.; Albuquerque, V.; Vasconcelos, P.R.L. Preconditioning with L-Ala-Gln reduces the expression of inflammatory markers (TNF-alpha, NF-kappaB, IL-6 and HO-1) in an injury animal model of cerebrovascular ischemia in Meriones unguiculatus (gerbils). Acta Cir. Bras. 2020, 35, e202000601. [Google Scholar] [CrossRef]
- Cruzat, V.F.; Keane, K.N.; Scheinpflug, A.L.; Cordeiro, R.; Soares, M.J.; Newsholme, P. Alanyl-glutamine improves pancreatic beta-cell function following ex vivo inflammatory challenge. J. Endocrinol. 2015, 224, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Tan, X.; Liu, Y.; You, W.; Liu, G.; Liu, X.; Jin, Q.; Wei, C.; Wan, F.; Zhao, H. Alanyl-glutamine ameliorates lipopolysaccharide-induced inflammation and barrier function injury in bovine jejunum epithelial cells. Biochem. Cell Biol. 2019, 97, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Mo, R.; Lai, R.; Lu, J.; Zhuang, Y.; Zhou, T.; Jiang, S.; Ren, P.; Li, Z.; Cao, Z.; Liu, Y.; et al. Enhanced autophagy contributes to protective effects of IL-22 against acetaminophen-induced liver injury. Theranostics 2018, 8, 4170–4180. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Huang, L.; Guo, E.; Li, R.; Yang, J.; Li, A.; Yang, Y.; Liu, S.; Hu, J.; Jiang, X.; et al. Baicalein pretreatment reduces liver ischemia/reperfusion injury via induction of autophagy in rats. Sci. Rep. 2016, 6, 25042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushima, N.; Yoshimori, T. How to interpret LC3 immunoblotting. Autophagy 2007, 3, 542–545. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene Accession Number | Forward Primer | Reverse Primer |
|---|---|---|---|
| TNF-α | NM_013693 | CAGGCGGTGCCTATGTCTC | CGATCACCCCGAAGTTCAGTAG |
| IL-6 | NM_031168 | AATTAAGCCTCCGACTTGTGAAG | CTTCCATCCAGTTGCCTTCTTG |
| IL-1β | NM_008361 | GAAATGCCACCTTTTGACAGTG | TGGATGCTCTCATCAGGACAG |
| MCP-1 | NM_011333 | TTAAAAACCTGGATCGGAACCAA | GCATTAGCTTCAGATTTACGGGT |
| RANTES | NM_013653 | GCTGCTTTGCCTACCTCTCC | TCGAGTGACAAACACGACTGC |
| GAPDH | NM_008084 | CGGTTCCGATGCCCTGAGGCTCTT | CGTCACACTTCATGATGGAATTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Ying, H.; Zheng, Y.; Ma, H.; Li, L.; Zhao, Y. Alanyl-Glutamine Protects against Lipopolysaccharide-Induced Liver Injury in Mice via Alleviating Oxidative Stress, Inhibiting Inflammation, and Regulating Autophagy. Antioxidants 2022, 11, 1070. https://doi.org/10.3390/antiox11061070
Hu J, Ying H, Zheng Y, Ma H, Li L, Zhao Y. Alanyl-Glutamine Protects against Lipopolysaccharide-Induced Liver Injury in Mice via Alleviating Oxidative Stress, Inhibiting Inflammation, and Regulating Autophagy. Antioxidants. 2022; 11(6):1070. https://doi.org/10.3390/antiox11061070
Chicago/Turabian StyleHu, Jiaji, Hanglu Ying, Yigang Zheng, Huabin Ma, Long Li, and Yufen Zhao. 2022. "Alanyl-Glutamine Protects against Lipopolysaccharide-Induced Liver Injury in Mice via Alleviating Oxidative Stress, Inhibiting Inflammation, and Regulating Autophagy" Antioxidants 11, no. 6: 1070. https://doi.org/10.3390/antiox11061070
APA StyleHu, J., Ying, H., Zheng, Y., Ma, H., Li, L., & Zhao, Y. (2022). Alanyl-Glutamine Protects against Lipopolysaccharide-Induced Liver Injury in Mice via Alleviating Oxidative Stress, Inhibiting Inflammation, and Regulating Autophagy. Antioxidants, 11(6), 1070. https://doi.org/10.3390/antiox11061070