
Role of Oxidative Stress in Vascular Low-Grade Inflammation Initiation Due to Acute Salt Loading in Young Healthy Individuals

 ,
,  ,
,  , , ,
, , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Study Protocol
2.3. 24-h Urine Samples Analysis
2.4. Anthropometric and Blood Pressure Measurement
2.5. Venous Blood Sampling and Analysis
2.6. Serum Protein Concentration of Pro- and Anti-Inflammatory Cytokines, C3a Complement Component, Soluble Cell Adhesion Molecules, and Endoglin Assay
2.7. Peripheral Blood Mononuclear Cells (PBMCs) Isolation, Cryopreservation, Thawing, and Cultivation
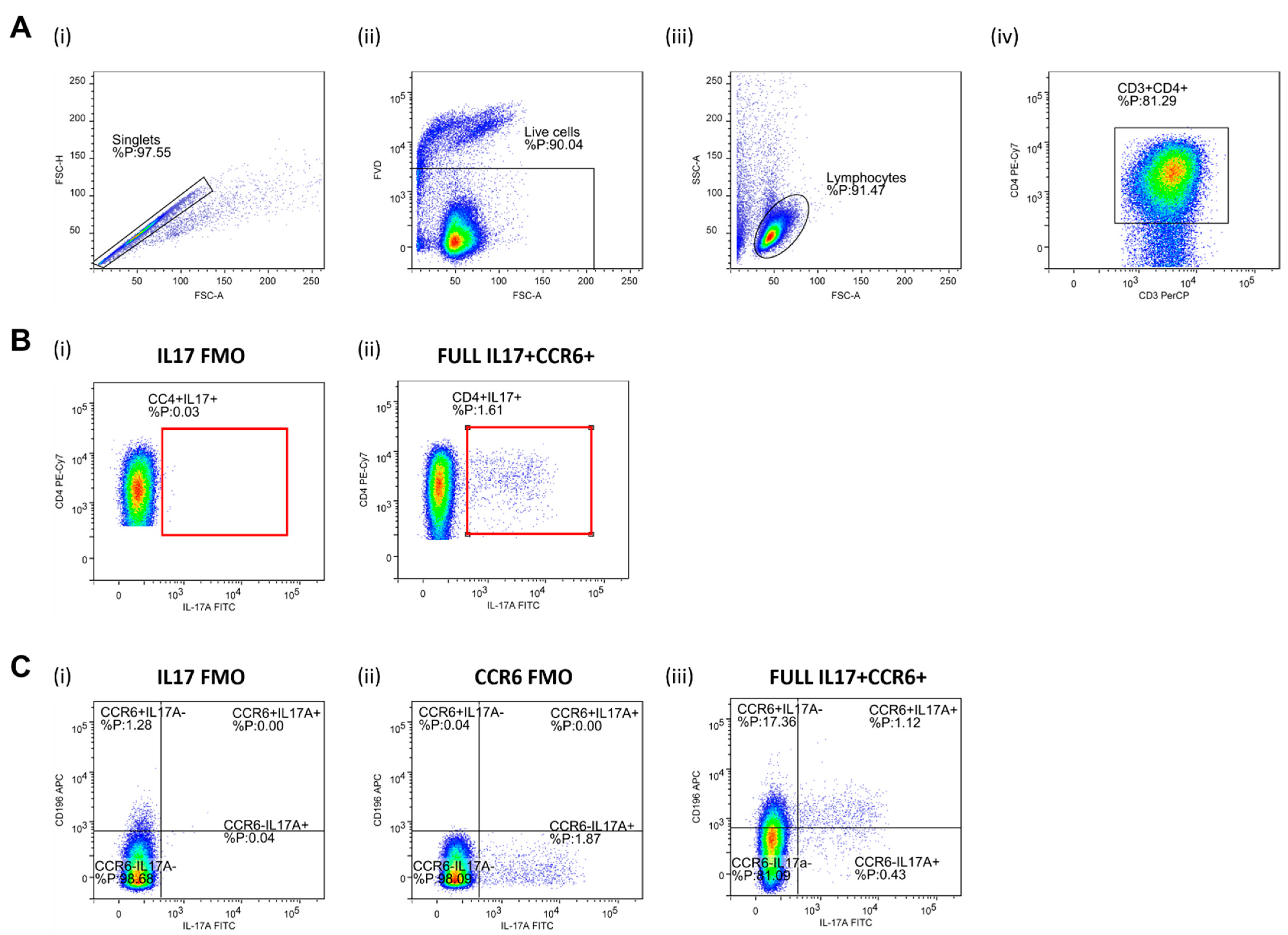
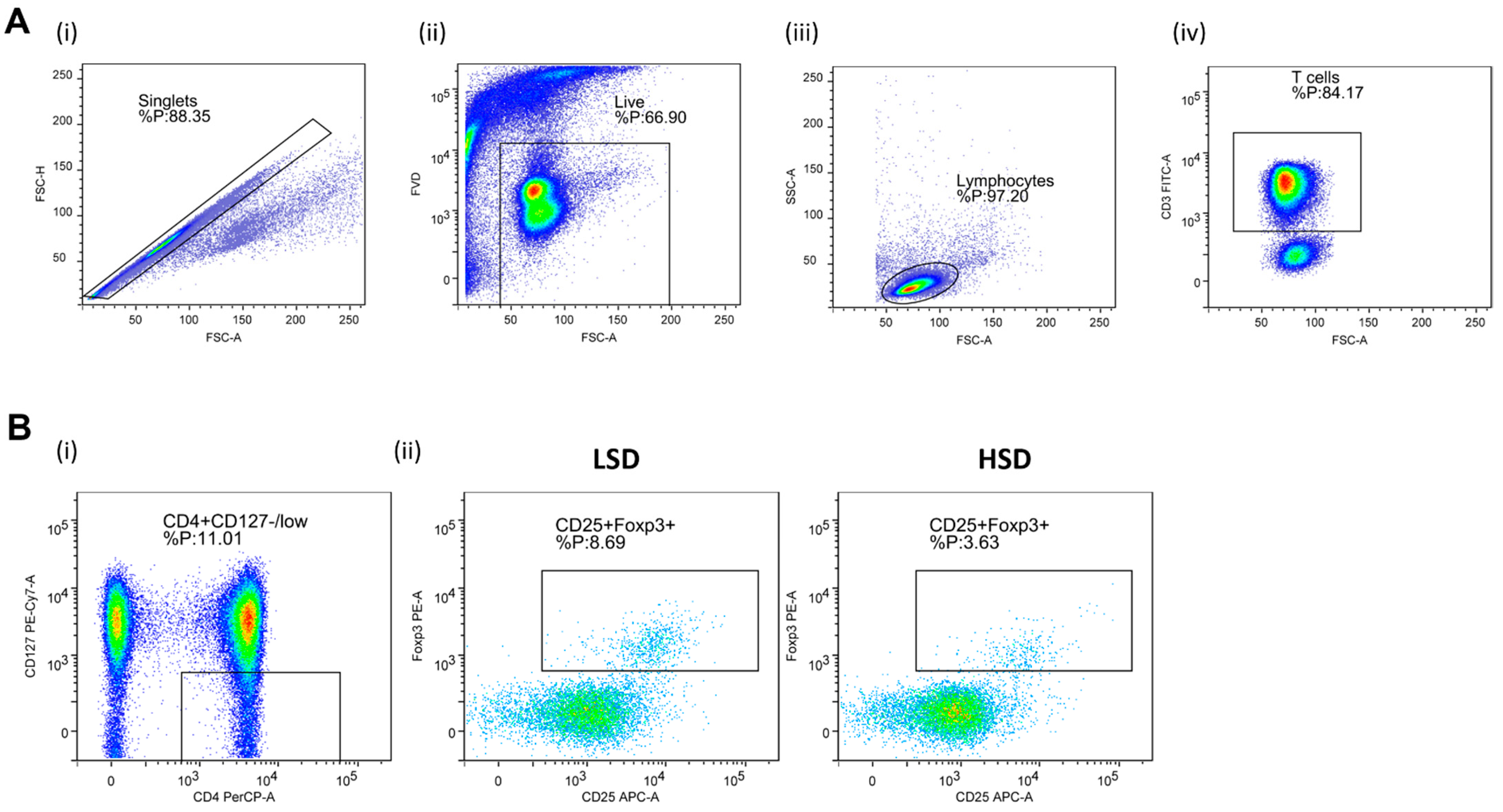
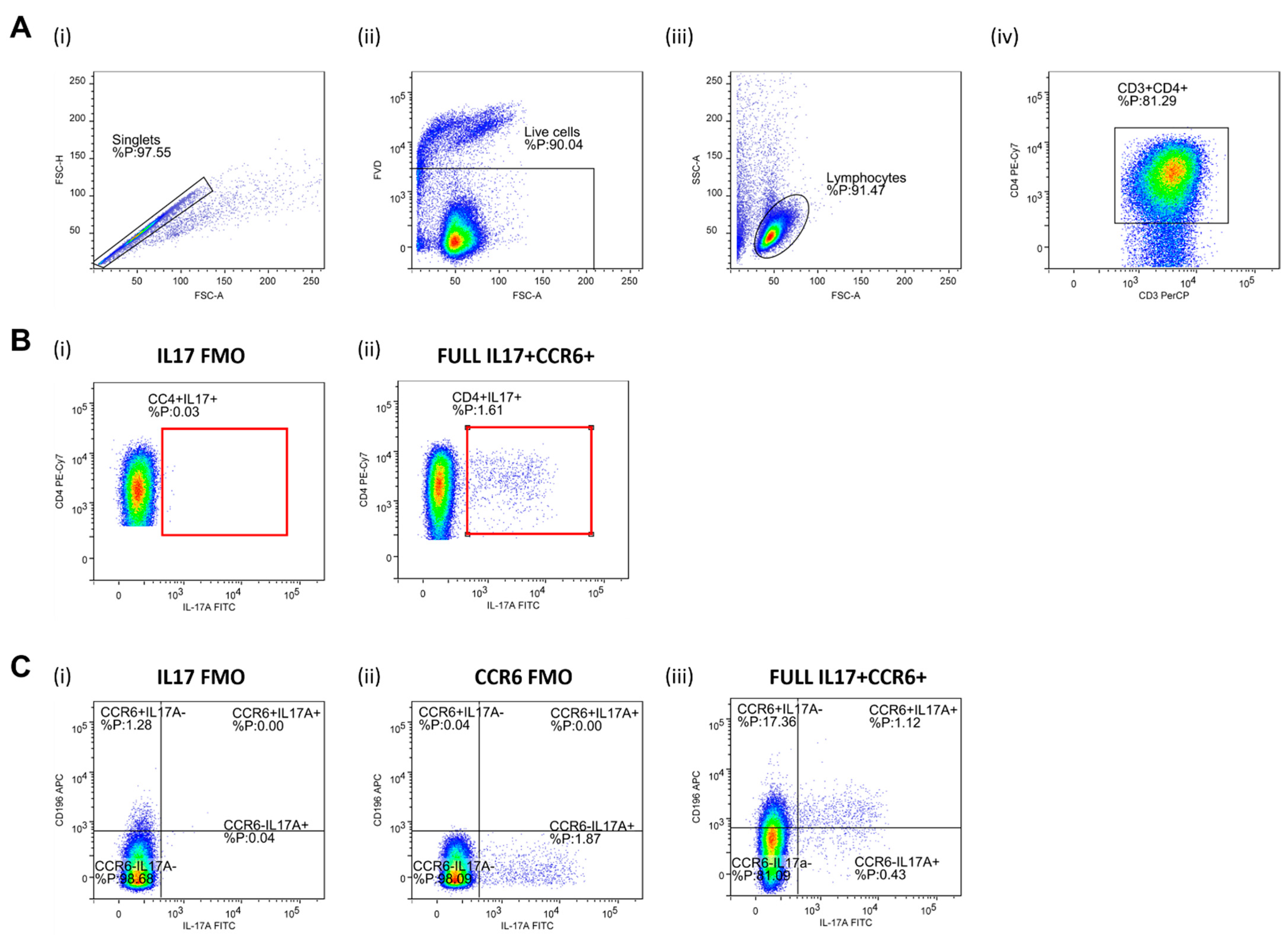
2.8. Regulatory T Lymphocytes (Treg) and Helper T Lymphocytes (Th17) Frequencies Assay and Th17/Treg Ratio
2.9. Serum- and Glucocorticoid Regulated Kinase 1 (SGK1) Assay
2.10. p38 Mitogen-Activated Protein (MAP) Kinase Assay
2.11. Biomarkers of Oxidative Stress and Antioxidant Defense Assay: Thiobarbituric Acid Reactive Substances (TBARS), Ferric-Reducing Ability of Plasma (FRAP), and Serum 8-Iso Prostaglandin F2 (8-Iso-PGF2α) Protein Concentration
2.12. Statistical Analysis
3. Results
3.1. Serum Pro- and Anti-Inflammatory Cytokines, C3a Complement Component, Soluble Cell Adhesion Molecules, and Endoglin Protein Concentration
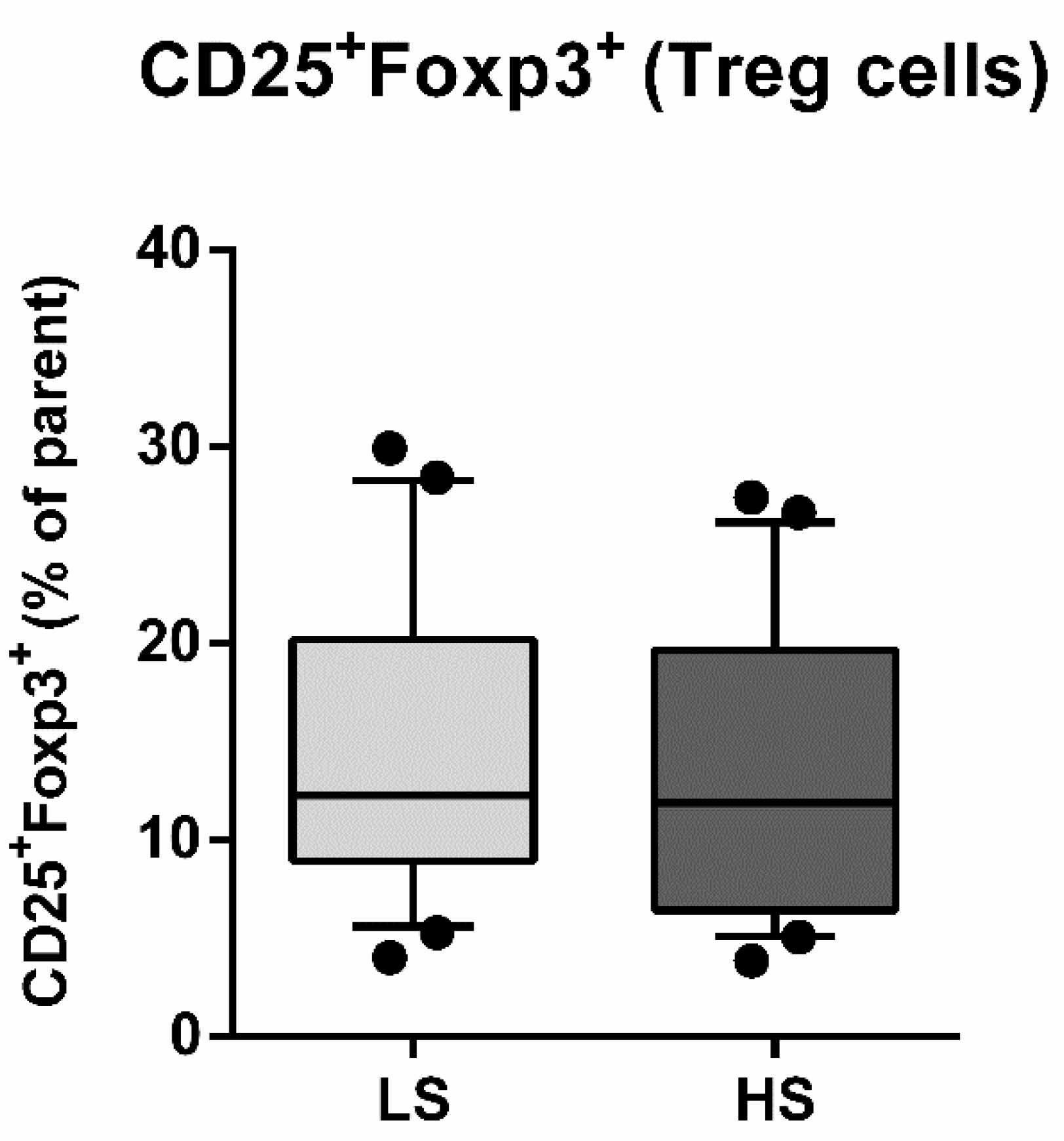
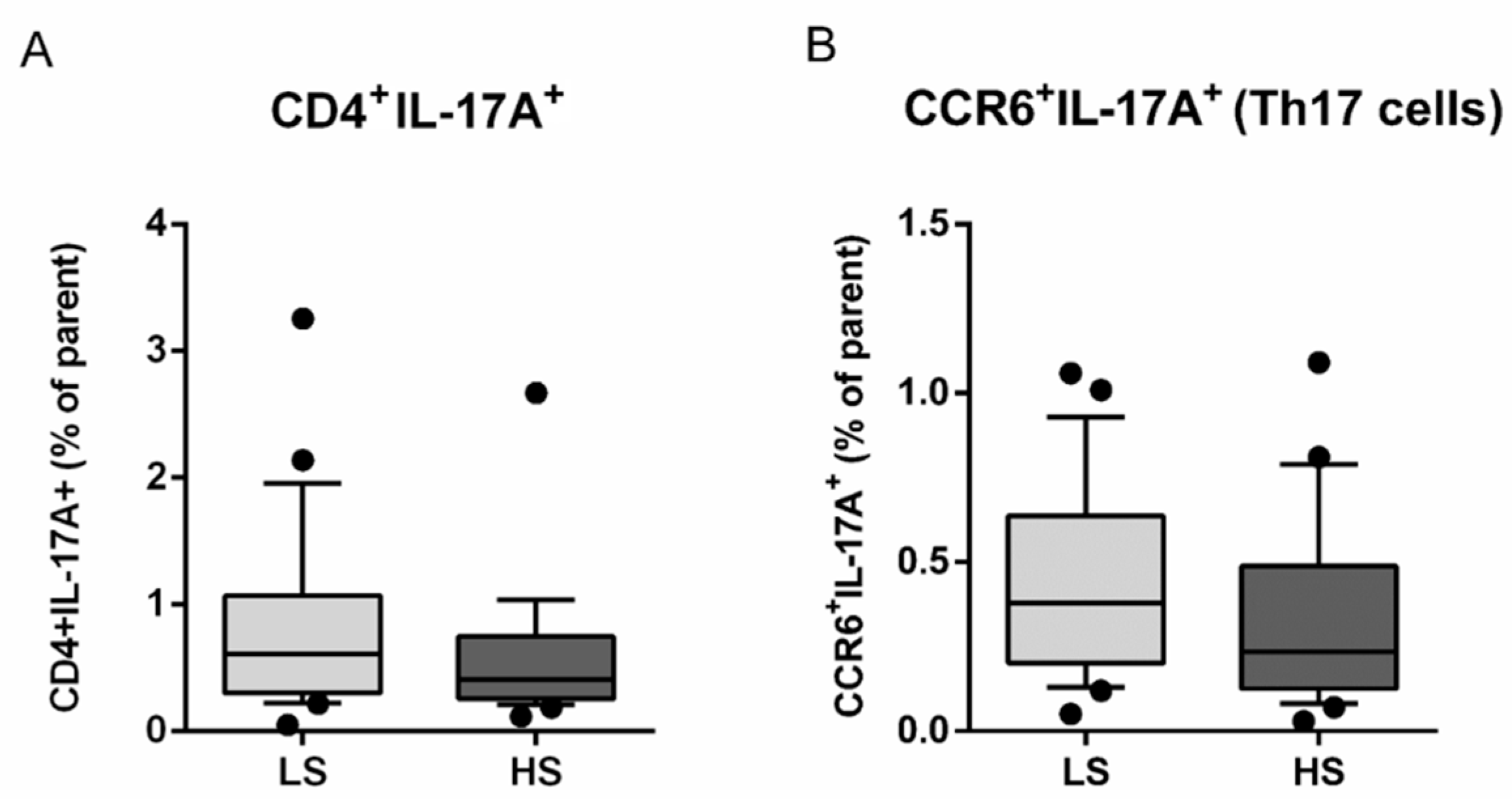
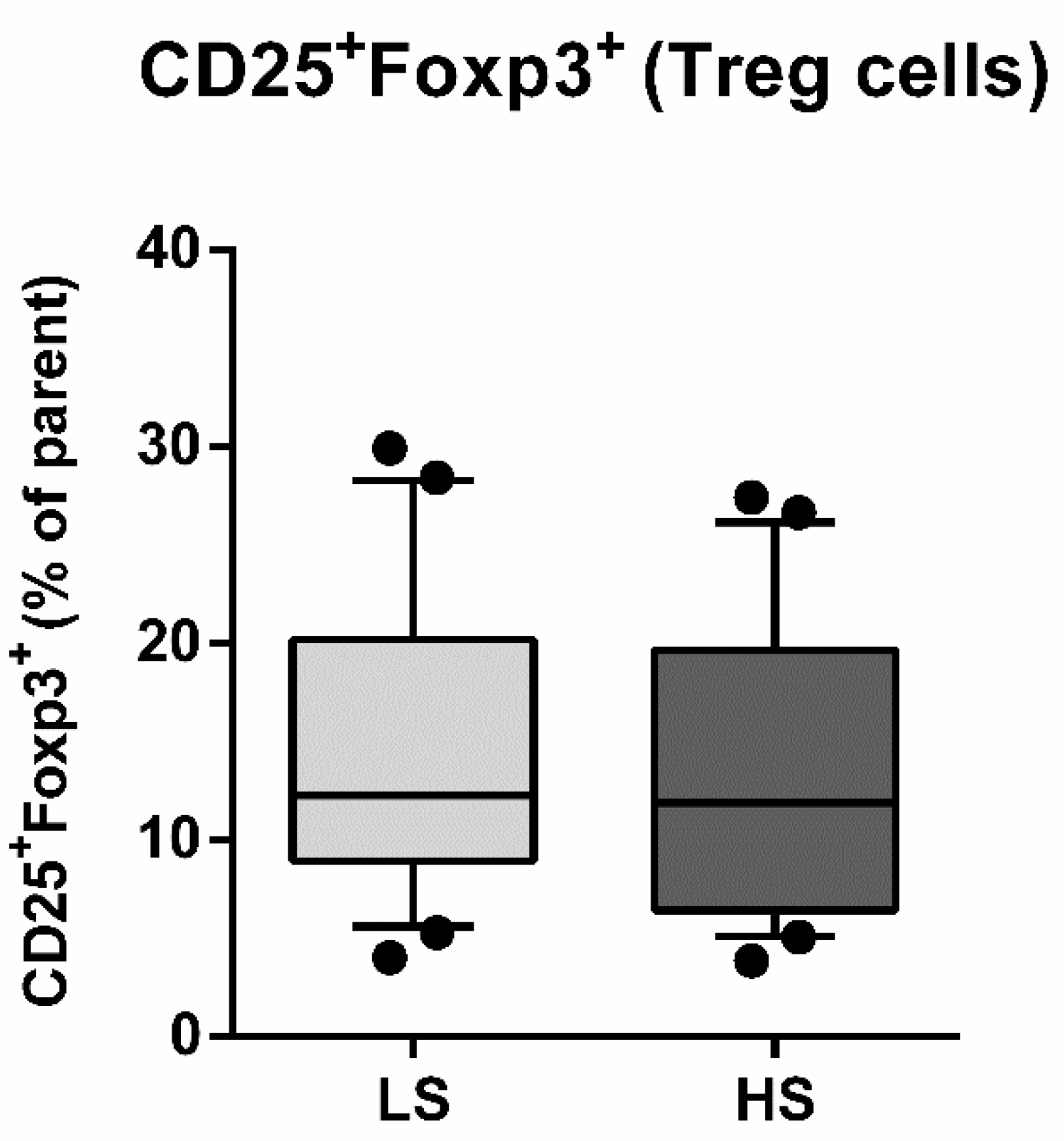
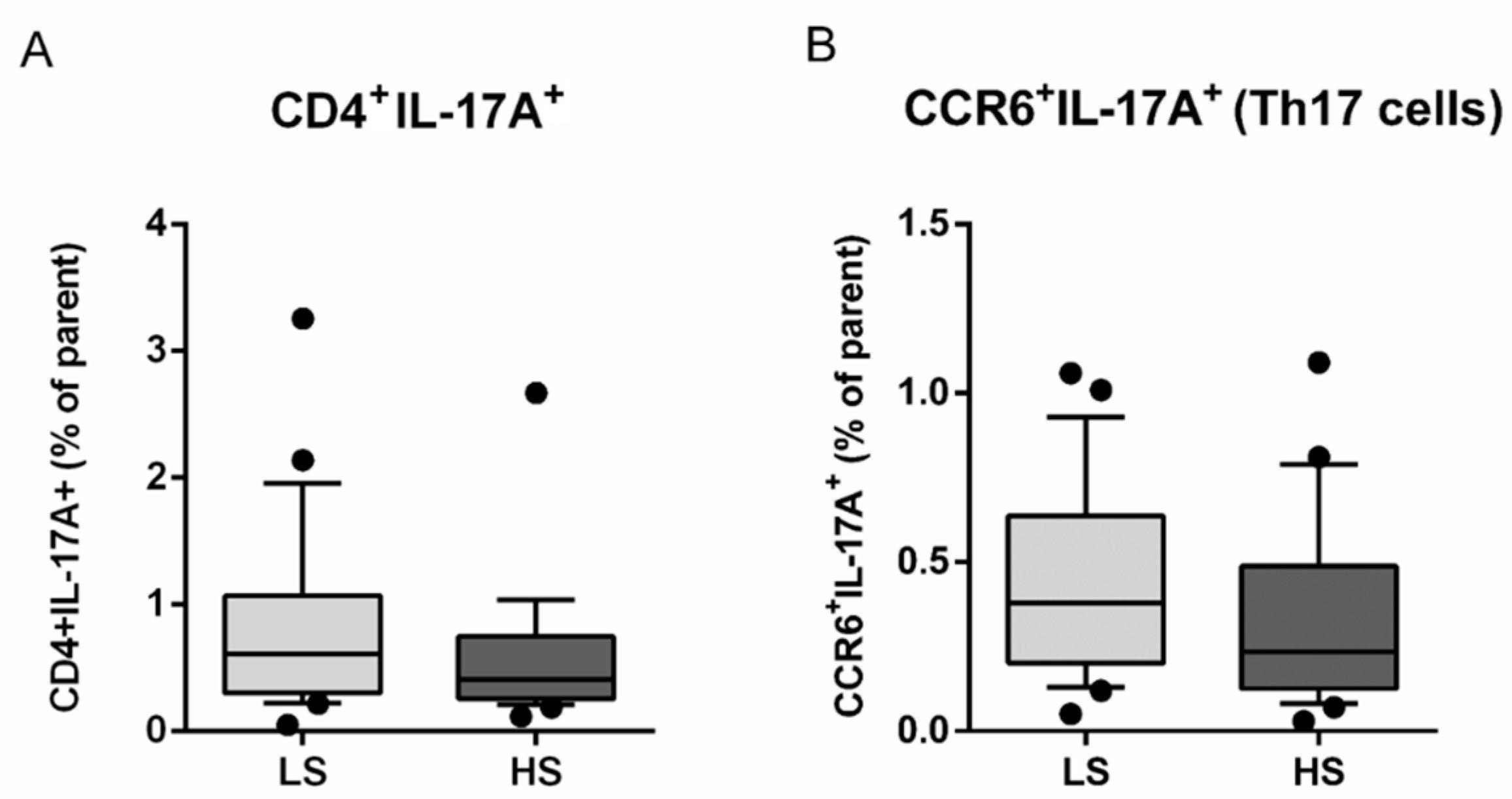
3.2. Frequencies of Regulatory T Lymphocytes (Treg) and Helper T Lymphocytes (Th17) and Th17/Treg Ratio
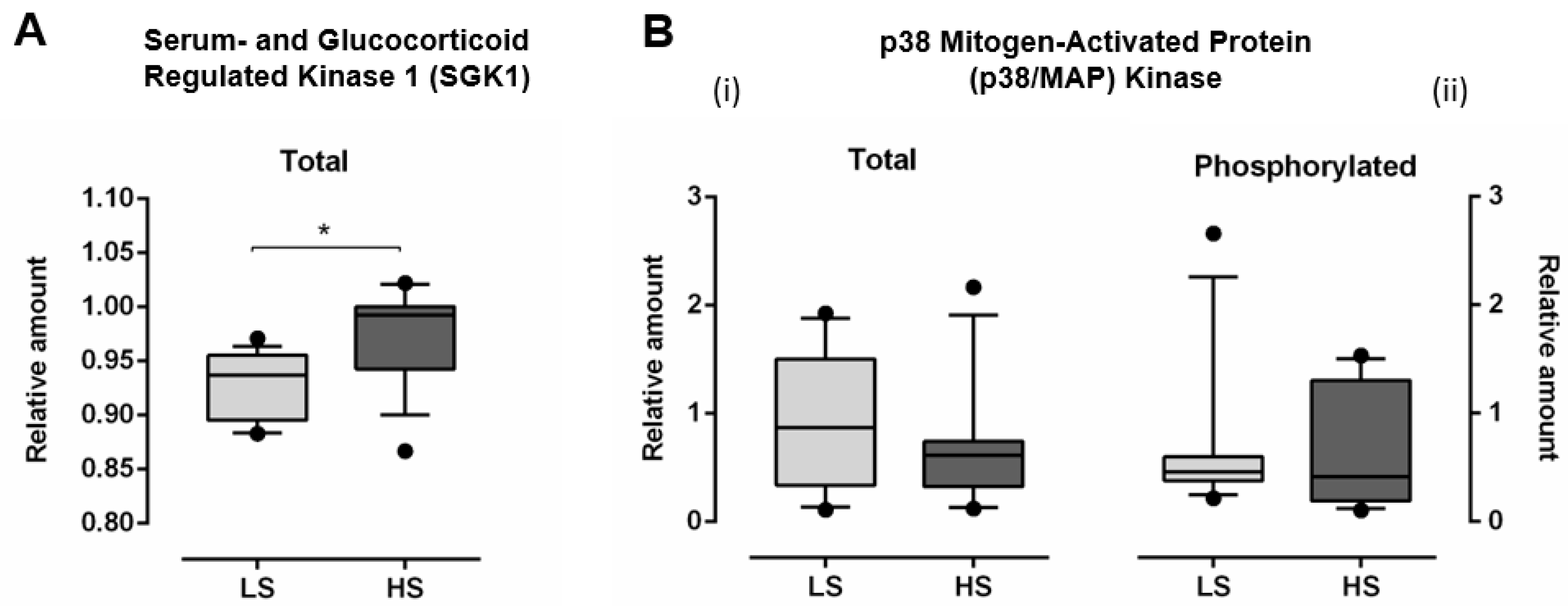
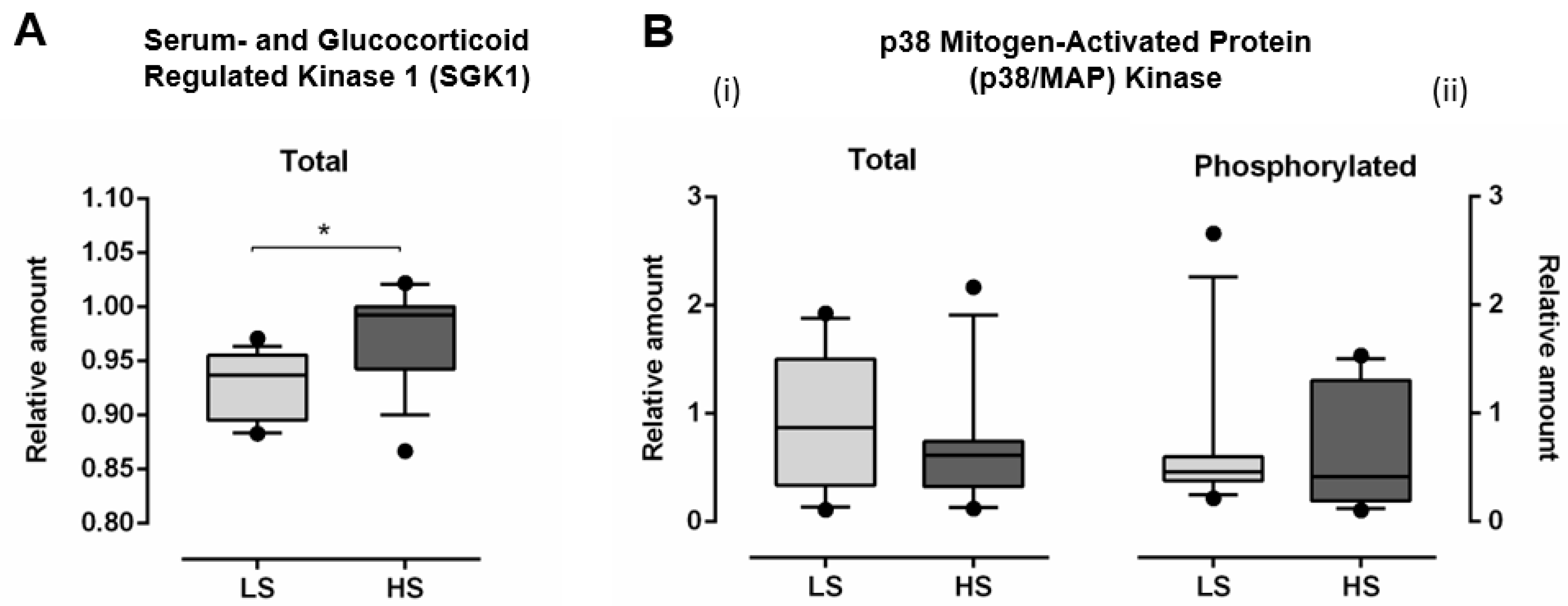
3.3. Serum- and Glucocorticoid Regulated Kinase 1 (SGK1) and p38 Mitogen-Activated Protein (MAP) Kinase
3.4. Markers of Oxidative Stress and Antioxidant Defense
3.5. Correlations
4. Discussion
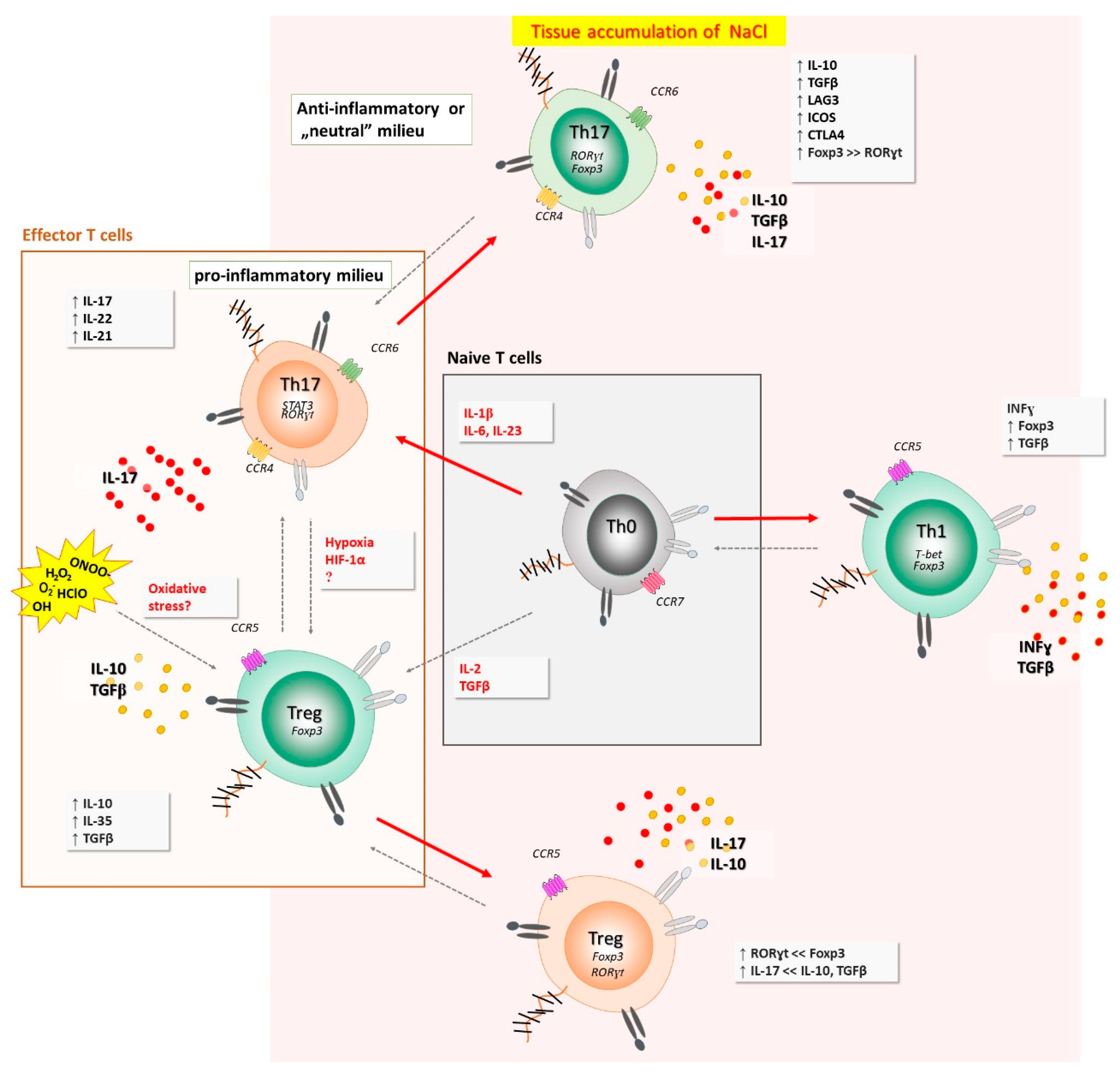
4.1. Salt Intake and Modulation of Early Inflammation
4.2. Salt Intake and Endothelial Activation
4.3. Increased Oxidative Stress Due to High-Salt Intake and Early Inflammation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pilic, L.; Pedlar, C.R.; Mavrommatis, Y. Salt-sensitive hypertension: Mechanisms and effects of dietary and other lifestyle factors. Nutr. Rev. 2016, 74, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boegehold, M.A.; Drenjancevic, I.; Lombard, J.H. Salt, Angiotensin II, superoxide, and endothelial function. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 215–254. [Google Scholar]
- Imaizumi, Y.; Eguchi, K.; Murakami, T.; Arakawa, K.; Tsuchihashi, T.; Kario, K. High salt intake is independently associated with hypertensive target organ damage. J. Clin. Hypertens. 2016, 18, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patik, J.C.; Lennon, S.L.; Farquhar, W.B.; Edwards, D.G. Mechanisms of dietary sodium-induced impairments in endothelial function and potential countermeasures. Nutrients 2021, 13, 270. [Google Scholar] [CrossRef] [PubMed]
- Barić, L.; Drenjančević, I.; Matić, A.; Stupin, M.; Kolar, L.; Mihaljević, Z.; Lenasi, H.; Šerić, V.; Stupin, A. Seven-Day salt loading impairs microvascular endothelium-dependent vasodilation without changes in blood pressure, body composition and fluid status in healthy young humans. Kidney Blood Press. Res. 2019, 44, 835–847. [Google Scholar] [CrossRef] [Green Version]
- Cosic, A.; Jukic, I.; Stupin, A.; Mihalj, M.; Mihaljevic, Z.; Novak, S.; Vukovic, R.; Drenjancevic, I. Attenuated flow-induced dilatation of middle cerebral arteries is related to increased vascular oxidative stress in rats on a short-term high salt diet. J. Physiol. 2016, 594, 4917–4931. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Drenjancevic-Peric, I.; McEwen, S.; Friesema, J.; Schulta, D.; Yu, M.; Roman, R.J.; Lombard, J.H. Role of superoxide and angiotensin II suppression in salt-induced changes in endothelial Ca2+ signaling and NO production in rat aorta. Am. J. Physiol. Circ. Physiol. 2006, 291, H929–H938. [Google Scholar] [CrossRef] [Green Version]
- Greaney, J.L.; DuPont, J.J.; Lennon-Edwards, S.L.; Sanders, P.W.; Edwards, D.G.; Farquhar, W.B. Dietary sodium loading impairs microvascular function independent of blood pressure in humans: Role of oxidative stress. J. Physiol. 2012, 590, 5519–5528. [Google Scholar] [CrossRef]
- Barić, L.; Drenjančević, I.; Mihalj, M.; Matić, A.; Stupin, M.; Kolar, L.; Mihaljević, Z.; Mrakovčić-Šutić, I.; Šerić, V.; Stupin, A. Enhanced antioxidative defense by vitamins C and E consumption prevents 7-day high-salt diet-induced microvascular endothelial function impairment in young healthy individuals. J. Clin. Med. 2020, 9, 843. [Google Scholar] [CrossRef] [Green Version]
- Afsar, B.; Kuwabara, M.; Ortiz, A.; Yerlikaya, A.; Siriopol, D.; Covic, A.; Rodriguez-Iturbe, B.; Johnson, R.J.; Kanbay, M. Salt Intake and Immunity. Hypertension 2018, 72, 19–23. [Google Scholar] [CrossRef]
- Benigni, A.; Cassis, P.; Remuzzi, G. Angiotensin II revisited: New roles in inflammation, immunology and aging. EMBO Mol. Med. 2010, 2, 247–257. [Google Scholar] [CrossRef]
- Takahashi, H.; Nakagawa, S.; Wu, Y.; Kawabata, Y.; Numabe, A.; Yanagi, Y.; Tamaki, Y.; Uehara, Y.; Araie, M. A high-salt diet enhances leukocyte adhesion in association with kidney injury in young dahl salt-sensitive rats. Hypertens. Res. 2017, 40, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wei, X.; Sun, R.; Cai, Y.; Lou, H.; Wang, J.-W.; Chen, A.F.; Zhang, X.-M. Angiotensin II stimulates intercellular adhesion molecule-1 via an AT1 receptor/nuclear factor-κB pathway in brain microvascular endothelial cells. Life Sci. 2006, 78, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Waki, H.; Liu, B.; Miyake, M.; Katahira, K.; Murphy, D.; Kasparov, S.; Paton, J.F.R. Junctional Adhesion Molecule-1 Is Upregulated in Spontaneously Hypertensive Rats. Hypertension 2007, 49, 1321–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrotra, P.; Patel, J.B.; Ivancic, C.M.; Collett, J.A.; Basile, D.P. Th-17 cell activation in response to high salt following acute kidney injury is associated with progressive fibrosis and attenuated by AT-1R antagonism. Kidney Int. 2015, 88, 776–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Yosef, N.; Thalhamer, T.; Zhu, C.; Xiao, S.; Kishi, Y.; Regev, A.; Kuchroo, V.K. Induction of pathogenic TH17 cells by inducible salt-sensing kinase SGK1. Nature 2013, 496, 513–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinewietfeld, M.; Manzel, A.; Titze, J.; Kvakan, H.; Yosef, N.; Linker, R.A.; Muller, D.N.; Hafler, D.A. Sodium chloride drives autoimmune disease by the induction of pathogenic TH17 cells. Nature 2013, 496, 518–522. [Google Scholar] [CrossRef]
- Ferri, C.; Bellini, C.; Desideri, G.; Giuliani, E.; De Siati, L.; Cicogna, S.; Santucci, A. Clustering of endothelial markers of vascular damage in human salt-sensitive hypertension. Hypertension 1998, 32, 862–868. [Google Scholar] [CrossRef] [Green Version]
- Yi, B.; Titze, J.; Rykova, M.; Feuerecker, M.; Vassilieva, G.; Nichiporuk, I.; Schelling, G.; Morukov, B.; Choukèr, A. Effects of dietary salt levels on monocytic cells and immune responses in healthy human subjects: A longitudinal study. Transl. Res. 2015, 166, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Luo, T.; Ji, W.; Yuan, F.; Guo, Z.; Li, Y.; Dong, Y.; Ma, Y.; Zhou, X.; Li, Y. Th17/treg imbalance induced by dietary salt variation indicates inflammation of target organs in humans. Sci. Rep. 2016, 6, 26767. [Google Scholar] [CrossRef]
- Mihalj, M.; Matić, A.; Mihaljević, Z.; Barić, L.; Stupin, A.; Drenjančević, I. Short-term high-nacl dietary intake changes leukocyte expression of VLA-4, LFA-1, and Mac-1 integrins in both healthy humans and sprague-dawley rats: A comparative study. Mediators Inflamm. 2019, 2019, 1–18. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, L.; Ji, W.-J.; Yuan, F.; Guo, Z.-Z.; Pang, B.; Luo, T.; Liu, X.; Zhang, W.-C.; Jiang, T.-M.; et al. Variation in dietary salt intake induces coordinated dynamics of monocyte subsets and monocyte-platelet aggregates in humans: Implications in end organ inflammation. PLoS ONE 2013, 8, e60332. [Google Scholar]
- Stupin, A.; Mihalj, M.; Kolobarić, N.; Šušnjara, P.; Kolar, L.; Mihaljević, Z.; Matić, A.; Stupin, M.; Jukić, I.; Kralik, Z.; et al. Anti-inflammatory potential of n-3 Polyunsaturated fatty acids enriched hen eggs consumption in improving microvascular endothelial function of healthy individuals—Clinical trial. Int. J. Mol. Sci. 2020, 21, 4149. [Google Scholar] [CrossRef] [PubMed]
- Kolobarić, N.; Drenjančević, I.; Matić, A.; Šušnjara, P.; Mihaljević, Z.; Mihalj, M. Dietary intake of n-3 PUFA-Enriched hen eggs changes inflammatory markers concentration and Treg/Th17 cells distribution in blood of young healthy adults—A randomised study. Nutrients 2021, 13, 1851. [Google Scholar] [CrossRef] [PubMed]
- Hashmat, S.; Rudemiller, N.; Lund, H.; Abais-Battad, J.M.; Van Why, S.; Mattson, D.L. Interleukin-6 inhibition attenuates hypertension and associated renal damage in Dahl salt-sensitive rats. Am. J. Physiol. Physiol. 2016, 311, F555–F561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostyk, A.G.; Dahl, K.M.; Wynes, M.W.; Whittaker, L.A.; Weiss, D.J.; Loi, R.; Riches, D.W.H. Regulation of Chemokine expression by NaCl occurs independently of cystic fibrosis transmembrane conductance regulator in macrophages. Am. J. Pathol. 2006, 169, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, F.; Ito, Y.; Mizuno, M.; Sawai, A.; Suzuki, Y.; Tomita, T.; Tawada, M.; Tanaka, A.; Hirayama, A.; Sagara, A.; et al. Sodium chloride promotes tissue inflammation via osmotic stimuli in subtotal-nephrectomized mice. Lab. Investig. 2017, 97, 432–446. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, A.L.; Kitz, A.; Wu, C.; Lowther, D.E.; Rodriguez, D.M.; Vudattu, N.; Deng, S.; Herold, K.C.; Kuchroo, V.K.; Kleinewietfeld, M.; et al. Sodium chloride inhibits the suppressive function of FOXP3+ regulatory T cells. J. Clin. Investig. 2015, 125, 4212–4222. [Google Scholar] [CrossRef] [Green Version]
- Sharif, K.; Amital, H.; Shoenfeld, Y. The role of dietary sodium in autoimmune diseases: The salty truth. Autoimmun. Rev. 2018, 17, 1069–1073. [Google Scholar] [CrossRef]
- Matthias, J.; Maul, J.; Noster, R.; Meinl, H.; Chao, Y.-Y.; Gerstenberg, H.; Jeschke, F.; Gasparoni, G.; Welle, A.; Walter, J.; et al. Sodium chloride is an ionic checkpoint for human T H 2 cells and shapes the atopic skin microenvironment. Sci. Transl. Med. 2019, 11, eaau0683. [Google Scholar] [CrossRef]
- Müller, D.N.; Wilck, N.; Haase, S.; Kleinewietfeld, M.; Linker, R.A. Sodium in the microenvironment regulates immune responses and tissue homeostasis. Nat. Rev. Immunol. 2019, 19, 243–254. [Google Scholar] [CrossRef]
- Wu, C.; Chen, Z.; Xiao, S.; Thalhamer, T.; Madi, A.; Han, T.; Kuchroo, V. SGK1 Governs the reciprocal development of Th17 and regulatory T cells. Cell Rep. 2018, 22, 653–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Ivanov, I.I.; Spolski, R.; Min, R.; Shenderov, K.; Egawa, T.; Levy, D.E.; Leonard, W.J.; Littman, D.R. IL-6 programs TH-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nat. Immunol. 2007, 8, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, D.A.; Zheng, S.G.; Wang, J.; Gray, J.D. Critical role of IL-2 and TGF-β in generation, function and stabilization of Foxp3+CD4+ Treg. Eur. J. Immunol. 2008, 38, 912–915. [Google Scholar] [CrossRef]
- Leong, M.L.L.; Maiyar, A.C.; Kim, B.; O’Keeffe, B.A.; Firestone, G.L. Expression of the serum- and glucocorticoid-inducible protein kinase, Sgk, is a cell survival response to multiple types of environmental stress stimuli in mammary epithelial cells. J. Biol. Chem. 2003, 278, 5871–5882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Rodríguez, C.; Aramburu, J.; Jin, L.; Rakeman, A.S.; Michino, M.; Rao, A. Bridging the NFAT and NF-κB families. Immunity 2001, 15, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Lang, F.; Stournaras, C.; Alesutan, I. Regulation of transport across cell membranes by the serum- and glucocorticoid-inducible kinase SGK1. Mol. Membr. Biol. 2014, 31, 29–36. [Google Scholar] [CrossRef]
- Lang, F.; Böhmer, C.; Palmada, M.; Seebohm, G.; Strutz-Seebohm, N.; Vallon, V. (Patho)physiological Significance of the Serum- and Glucocorticoid-Inducible Kinase Isoforms. Physiol. Rev. 2006, 86, 1151–1178. [Google Scholar] [CrossRef]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-Activated protein kinases and reactive oxygen species: How can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 1–6. [Google Scholar] [CrossRef]
- Wilck, N.; Matus, M.G.; Kearney, S.M.; Olesen, S.W.; Forslund, K.; Bartolomaeus, H.; Haase, S.; Mähler, A.; Balogh, A.; Markó, L.; et al. Salt-responsive gut commensal modulates TH17 axis and disease. Nature 2017, 551, 585–589. [Google Scholar] [CrossRef]
- Vitales-Noyola, M.; Layseca-Espinosa, E.; Baranda, L.; Abud-Mendoza, C.; Niño-Moreno, P.; Monsiváis-Urenda, A.; Rosenstein, Y.; González-Amaro, R. Analysis of sodium chloride intake and Treg/Th17 lymphocytes in healthy individuals and patients with rheumatoid arthritis or systemic lupus erythematosus. J. Immunol. Res. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Matthias, J.; Heink, S.; Picard, F.; Zeiträg, J.; Kolz, A.; Chao, Y.-Y.; Soll, D.; de Almeida, G.P.; Glasmacher, E.; Jacobsen, I.D.; et al. Salt generates antiinflammatory Th17 cells but amplifies pathogenicity in proinflammatory cytokine microenvironments. J. Clin. Investig. 2020, 130, 4587–4600. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Kim, N.; Kang, J.; Yoon, S.; Lee, H.-A.; Jung, H.; Kim, S.-H.; Kim, I. Activated pathogenic Th17 lymphocytes induce hypertension following high-fructose intake in Dahl salt-sensitive (SS) but not Dahl salt-resistant (SR) rats. Dis. Model. Mech. 2020, 13, dmm044107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golias, C.; Tsoutsi, E.; Matziridis, A.; Makridis, P.; Batistatou, A.; Charalabopoulos, K. Review. Leukocyte and endothelial cell adhesion molecules in inflammation focusing on inflammatory heart disease. In Vivo 2007, 21, 757–769. [Google Scholar]
- Abram, C.L.; Lowell, C.A. The ins and outs of leukocyte integrin signaling. Annu. Rev. Immunol. 2009, 27, 339–362. [Google Scholar] [CrossRef] [Green Version]
- Tadzic, R.; Mihalj, M.; Vcev, A.; Ennen, J.; Tadzic, A.; Drenjancevic, I. The effects of arterial blood pressure reduction on endocan and soluble endothelial cell adhesion molecules (CAMs) and CAMs ligands expression in hypertensive patients on ca-channel blocker therapy. Kidney Blood Press. Res. 2013, 37, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Cavka, A.; Cosic, A.; Jukic, I.; Jelakovic, B.; Lombard, J.H.; Phillips, S.A.; Seric, V.; Mihaljevic, I.; Drenjancevic, I. The role of cyclo-oxygenase-1 in high-salt diet-induced microvascular dysfunction in humans. J. Physiol. 2015, 593, 5313–5324. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, Y.; Xie, L.; Jin, K.; Sheibani, N.; Greenberg, D.A. Hypoxic induction of endoglin via mitogen-activated protein kinases in mouse brain microvascular endothelial cells. Stroke 2003, 34, 2483–2488. [Google Scholar] [CrossRef]
- Schoonderwoerd, M.J.A.; Goumans, M.-J.T.H.; Hawinkels, L.J.A.C. Endoglin: Beyond the endothelium. Biomolecules 2020, 10, 289. [Google Scholar] [CrossRef] [Green Version]
- Lenda, D.M.; Sauls, B.A.; Boegehold, M.A. Reactive oxygen species may contribute to reduced endothelium-dependent dilation in rats fed high salt. Am. J. Physiol. Circ. Physiol. 2000, 279, H7–H14. [Google Scholar] [CrossRef]
- Drenjancevic-Peric, I.; Lombard, J.H. Reduced angiotensin II and oxidative stress contribute to impaired vasodilation in dahl salt-sensitive rats on low-salt diet. Hypertension 2005, 45, 687–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, V.; Cornelius, C.; Maiolino, L.; Luca, M.; Chiaramonte, R.; Toscano, M.A.; Serra, A. Oxidative stress, redox homeostasis and cellular stress response in Ménière’s disease: Role of vitagenes. Neurochem. Res. 2010, 35, 2208–2217. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, C.; Perrotta, R.; Graziano, A.; Calabrese, E.J.; Calabrese, V. Stress responses, vitagenes and hormesis as critical determinants in aging and longevity: Mitochondria as a “chi”. Immun. Ageing 2013, 10, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelius, C.; Graziano, A.; Calabrese, E.J.; Calabrese, V. Hormesis and vitagenes in aging and longevity: Mitochondrial control and hormonal regulation. Horm. Mol. Biol. Clin. Investig. 2013, 16, 73–89. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | LS | HS |
|---|---|---|
| N (F/M) | 30 (15/15) | |
| Age (Years) | 21 [19–23] | |
| Anthropometric Parameters | ||
| BMI (kg/m2) | 24.03 [22.14–25.91] | 24.03 [22.46–25.91] |
| WHR | 0.83 [0.76–0.87] | 0.82 [0.77–0.84] |
| Hemodynamic Parameters | ||
| SBP (mmHg) | 116 ± 11 | 116 ± 13 |
| DBP (mmHg) | 75 [70–80] | 73 [71–78] |
| MAP (mmHg) | 88 ± 7 | 88 ± 6 |
| 24-h Urine Biochemical Parameters | ||
| 24 h urine volume (mL) | 1573 ± 760 | 1645 ± 740 |
| 24 h creatinine coefficient (µmol/24 h/kg) | 153.5 ± 62.6 | 157.0 ± 52.5 |
| 24 h urine urea (mmol/dU) | 234.3 ± 132.1 | 238.5 ± 100.6 |
| 24 h urine protein (mg/dU) | 91.5 [50.0–107.8] | 81.0 [66.0–107.5] |
| 24 h urine albumin (mg/dU) | 5.3 [2.5–9.4] | 5.6 [4.0–8.0] |
| 24 h sodium (mmol/dU) | 66.0 [46.0–114.8] | 211.5 [162.5–336.0] * |
| 24 h potassium (mmol/dU) | 40.5 ± 17.1 | 48.8 ± 23.3 |
| calculated salt intake (g/day) | 3.9 [2.7–6.7] | 12.4 [9.5–19.6] * |
| Serum Biochemical Parameters | ||
| leukocytes (×10−9/L) | 6.2 [5.5–7.4] | 6.0 [5.2–7.3] |
| erythrocytes (×10−12/L) | 5.02 ± 0.55 | 4.89 ± 0.55 |
| hemoglobin (g/L) | 145.3 ± 15.7 | 142.0 ± 15.7 |
| hematocrit (%) | 42.2 ± 4.4 | 41.4 ± 4.3 |
| thrombocytes (×10−9/L) | 242.3 ± 38.4 | 229.1 ± 42.3 |
| urea (mmol/L) | 4.3 ± 1.1 | 4.4 ± 0.9 |
| creatinine (µmol/L) | 76.7 ± 12.6 | 71.1 ± 9.4 |
| sodium (mmol/L) | 138.0 [137.0–139.0] | 140.0 [139.0–141.0] * |
| potassium (mmol/L) | 4.0 [3.9–4.2] | 4.1 [4.0–4.3] * |
| calcium (mmol/L) | 2.48 ± 0.07 | 2.43 ± 0.10 * |
| hsCRP (mg/L) | 0.55 [0.32–1.13] | 0.56 [0.34–0.80] |
| Parameter (pg/mL) | LS | HS |
|---|---|---|
| N (F/M) | 30 (15/15) | |
| Pro-inflammatory cytokines | ||
| IFNγ | 1.26 [0.76–1.58] | 1.80 [1.31–4.85] * |
| TNF-α | 7.55 ± 12.27 | 14.49 ± 16.23 * |
| IL-6 | 2.24 [1.35–3.18] | 2.82 [1.93–3.53] |
| IL-9 | 0.09 [0.05–0.23] | 0.19 [0.11–3.49] * |
| IL-17A | 1.52 [0.89–4.67] | 2.02 [1.56–12.2] * |
| IL-23 | 0.05 ± 0.02 | 0.05 ± 0.02 |
| Anti-inflammatory (immunomodulatory) cytokines | ||
| IL-10 | 0.44 [0.27–0.66] | 0.68 [0.44–3.60] * |
| IL-21 | 168.2 ± 210.5 | 281.6 ± 296.8 |
| IL-22 | 1.39 [1.10–2.18] | 1.88 [1.29–3.21] |
| TGF-β1 | 36.6 ± 33.0 | 49.7 ± 31.6 * |
| C3a Complement Component | 202.3 ± 159.4 | 204.5 ± 178.0 |
| Soluble Cell Adhesion Molecules | ||
| sICAM-1 | 1108 [444–1623] | 968 [537–1558] |
| sVCAM-1 | 3342 [2195–4808] | 3230 [2524–4024] |
| E-selectin (CD-62E) | 132.4 ± 47.5 | 130.5 ± 39.3 |
| Endoglin | 1753 [1118–2679] | 1709 [1337–2599] |
| Parameter (pg/mL) | LS | HS |
|---|---|---|
| N (F/M) | 30 (15/15) | |
| Biomarkers of Oxidative Stress Level | ||
| TBARS (μM/MDA) | 21.6 [16.8–24.1] | 24.3 [19.5–26.4] * |
| 8-iso-PGF2α (pg/mL) | 692.5 ± 112.3 | 813.7 ± 78.7 † |
| Biomarker of Antioxidant Defense | ||
| FRAP (mM/L TE) | 0.33 [0.30–0.39] | 0.21 [0.18–0.28] ‡ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knezović, A.; Kolobarić, N.; Drenjančević, I.; Mihaljević, Z.; Šušnjara, P.; Jukić, I.; Stupin, M.; Kibel, A.; Marczi, S.; Mihalj, M.; et al. Role of Oxidative Stress in Vascular Low-Grade Inflammation Initiation Due to Acute Salt Loading in Young Healthy Individuals. Antioxidants 2022, 11, 444. https://doi.org/10.3390/antiox11030444
Knezović A, Kolobarić N, Drenjančević I, Mihaljević Z, Šušnjara P, Jukić I, Stupin M, Kibel A, Marczi S, Mihalj M, et al. Role of Oxidative Stress in Vascular Low-Grade Inflammation Initiation Due to Acute Salt Loading in Young Healthy Individuals. Antioxidants. 2022; 11(3):444. https://doi.org/10.3390/antiox11030444
Chicago/Turabian StyleKnezović, Ana, Nikolina Kolobarić, Ines Drenjančević, Zrinka Mihaljević, Petar Šušnjara, Ivana Jukić, Marko Stupin, Aleksandar Kibel, Saška Marczi, Martina Mihalj, and et al. 2022. "Role of Oxidative Stress in Vascular Low-Grade Inflammation Initiation Due to Acute Salt Loading in Young Healthy Individuals" Antioxidants 11, no. 3: 444. https://doi.org/10.3390/antiox11030444
APA StyleKnezović, A., Kolobarić, N., Drenjančević, I., Mihaljević, Z., Šušnjara, P., Jukić, I., Stupin, M., Kibel, A., Marczi, S., Mihalj, M., & Stupin, A. (2022). Role of Oxidative Stress in Vascular Low-Grade Inflammation Initiation Due to Acute Salt Loading in Young Healthy Individuals. Antioxidants, 11(3), 444. https://doi.org/10.3390/antiox11030444