
Ursolic Acid Protects Sodium Dodecyl Sulfate-Induced Drosophila Ulcerative Colitis Model by Inhibiting the JNK Signaling



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Fly Husbandry
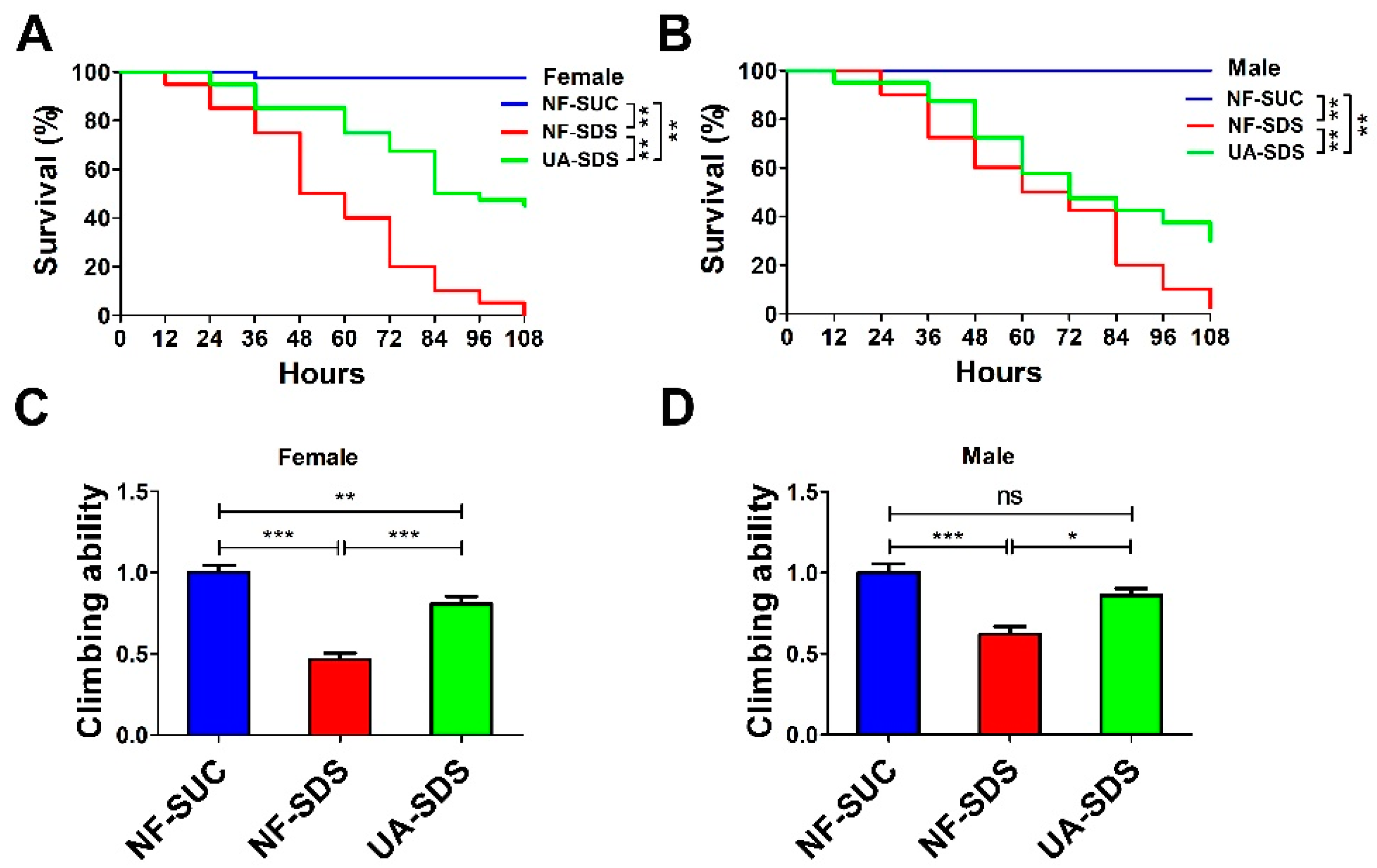
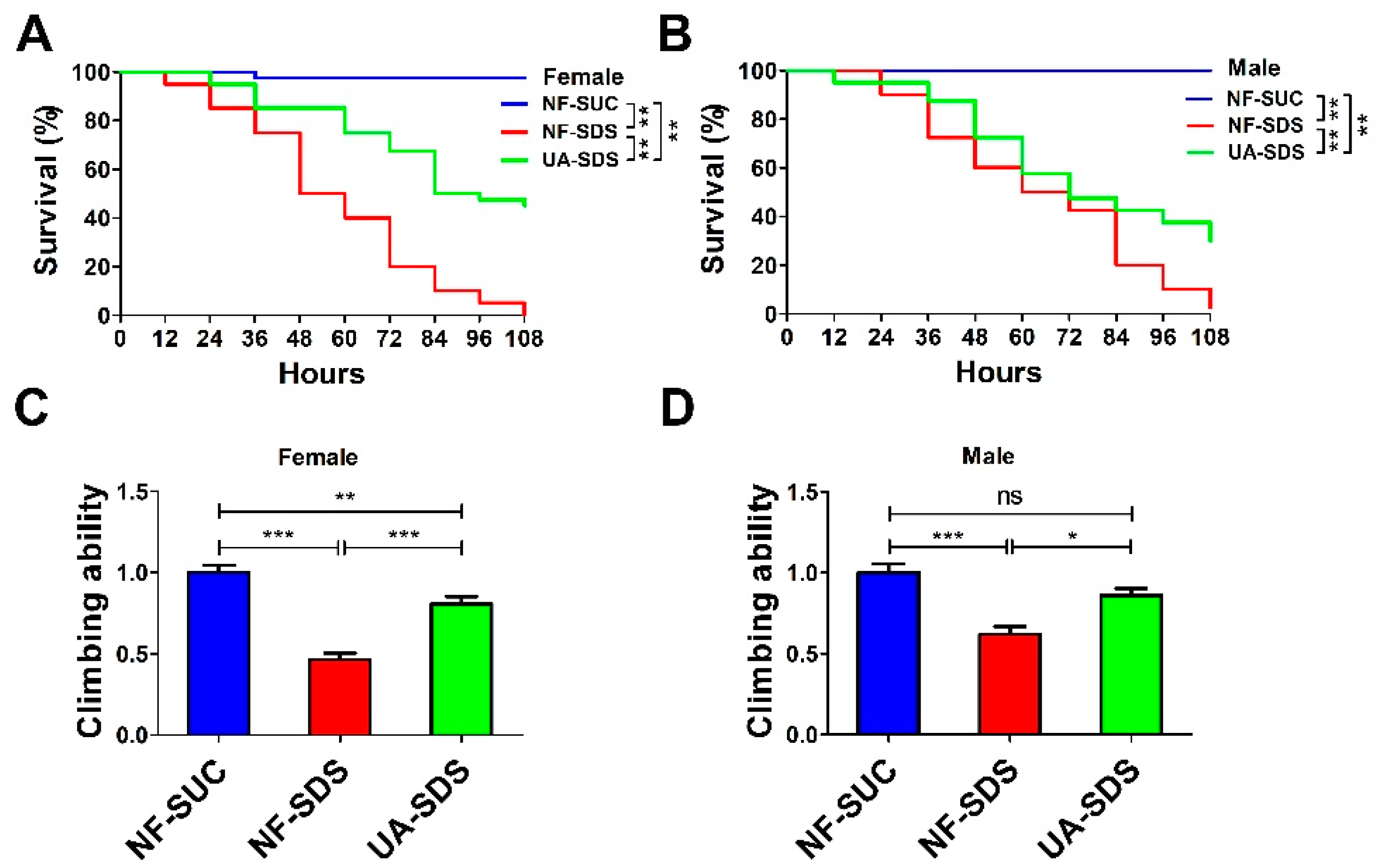
2.3. Survival Assays
2.4. Climbing Ability Assay
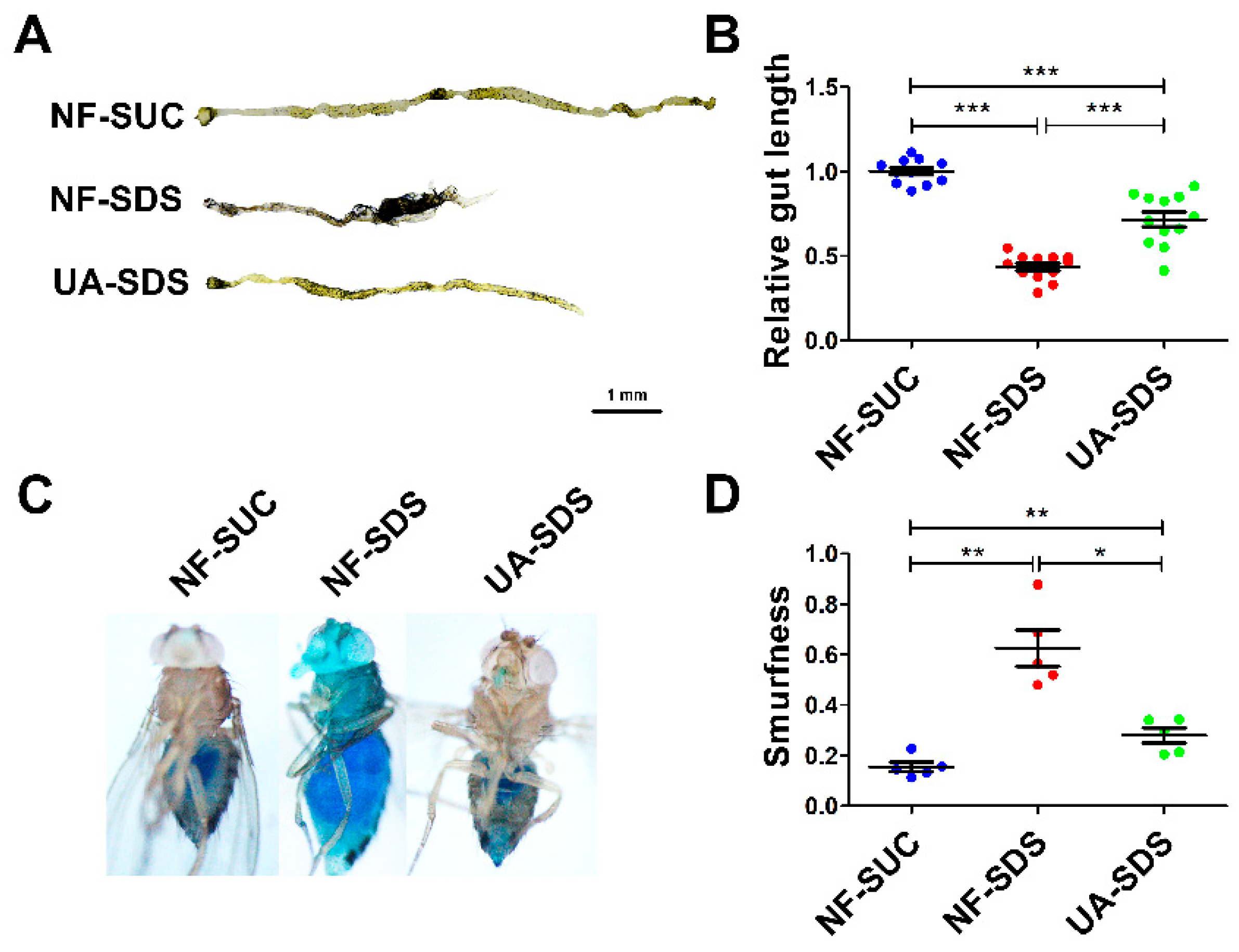
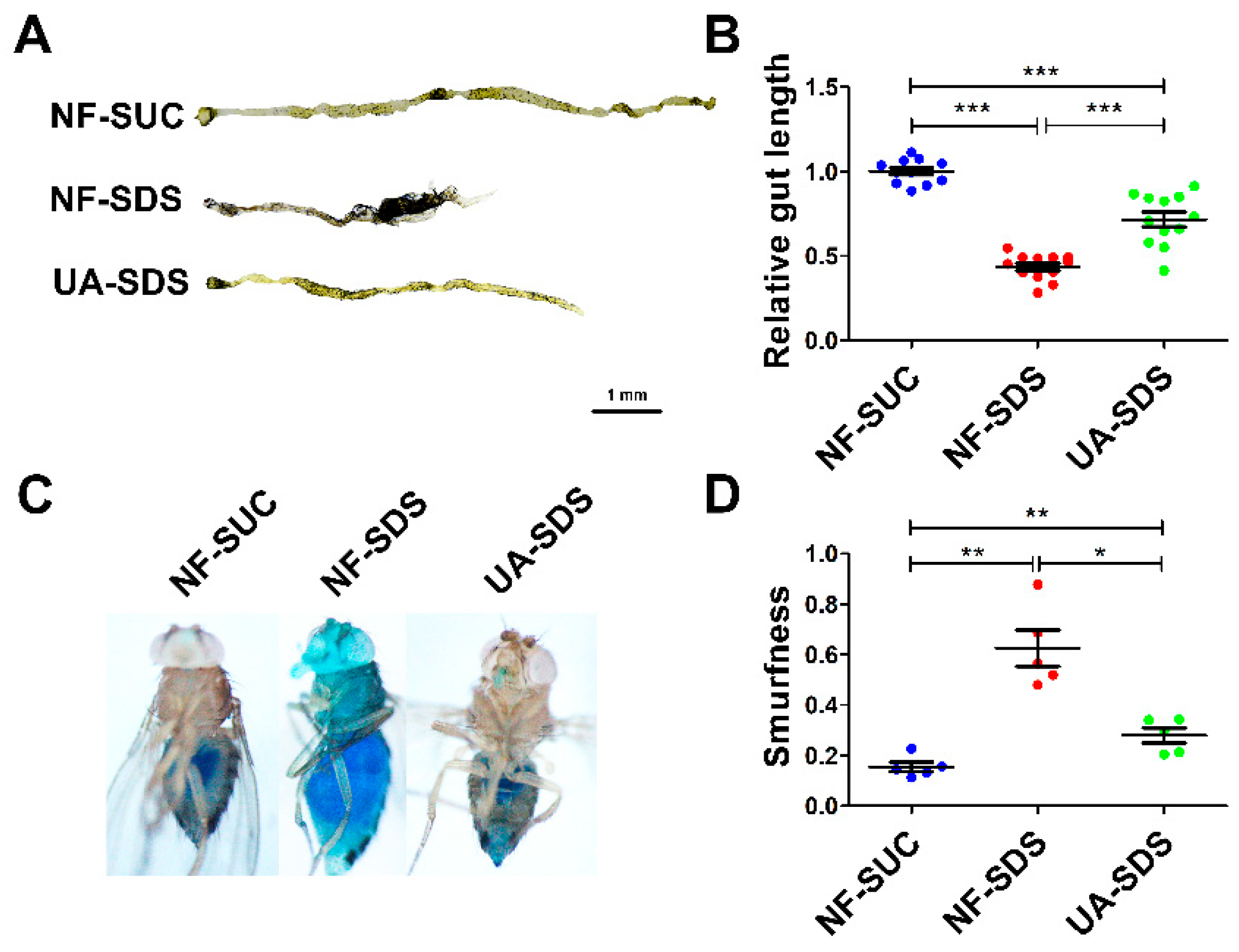
2.5. Intestinal Morphology Analysis
2.6. Smurf Assay
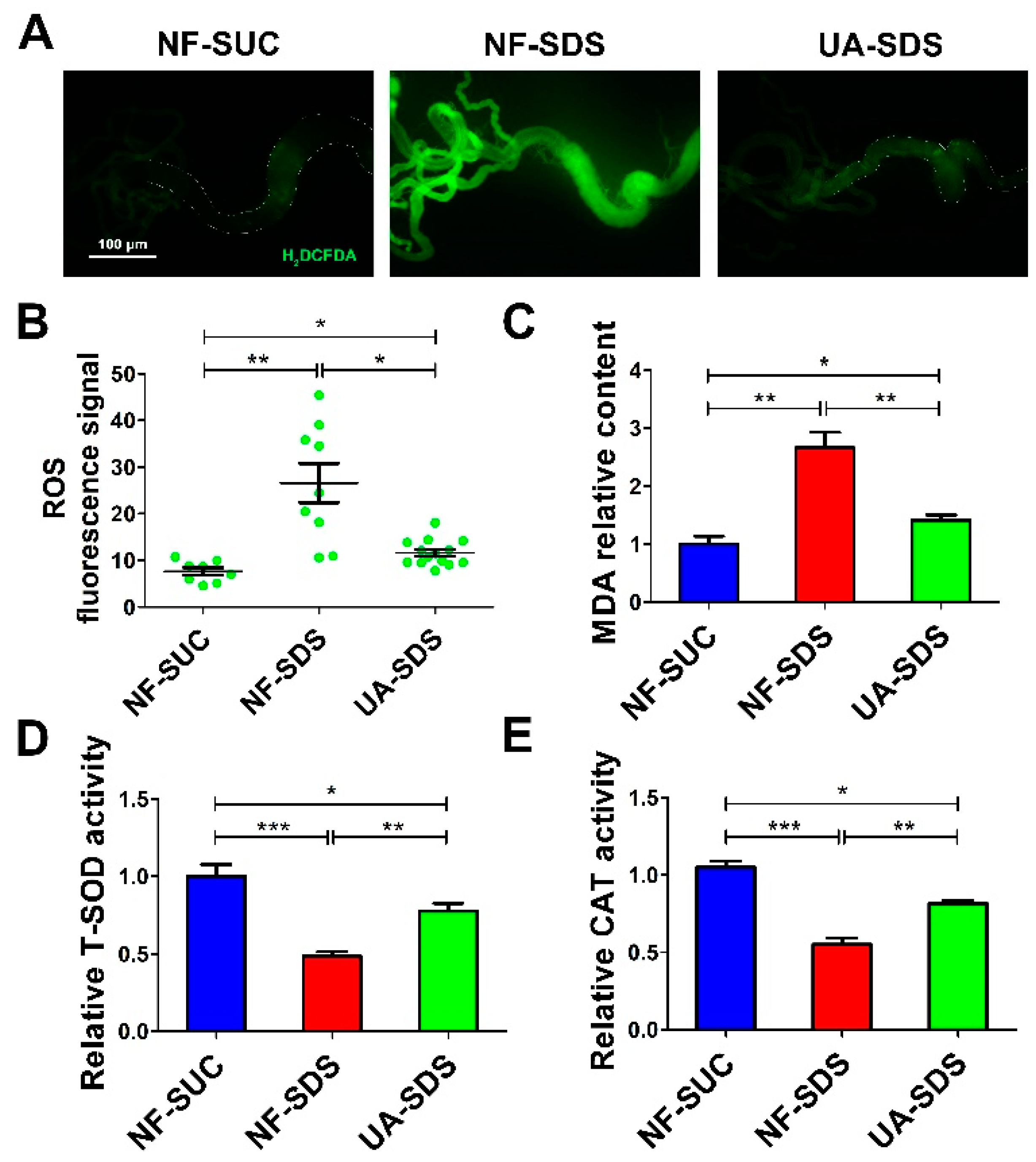
2.7. Reactive Oxygen Species Assay
2.8. Immunohistochemistry
2.9. Fluorescence Microscopy
2.10. Statistical Analysis
3. Results
3.1. The Intestinal Disruption Induced by SDS in Drosophila Could Be Rescued by UA
3.2. Protective Effect of UA against Morphological Changes in the Drosophila Intestine following Treatment with SDS
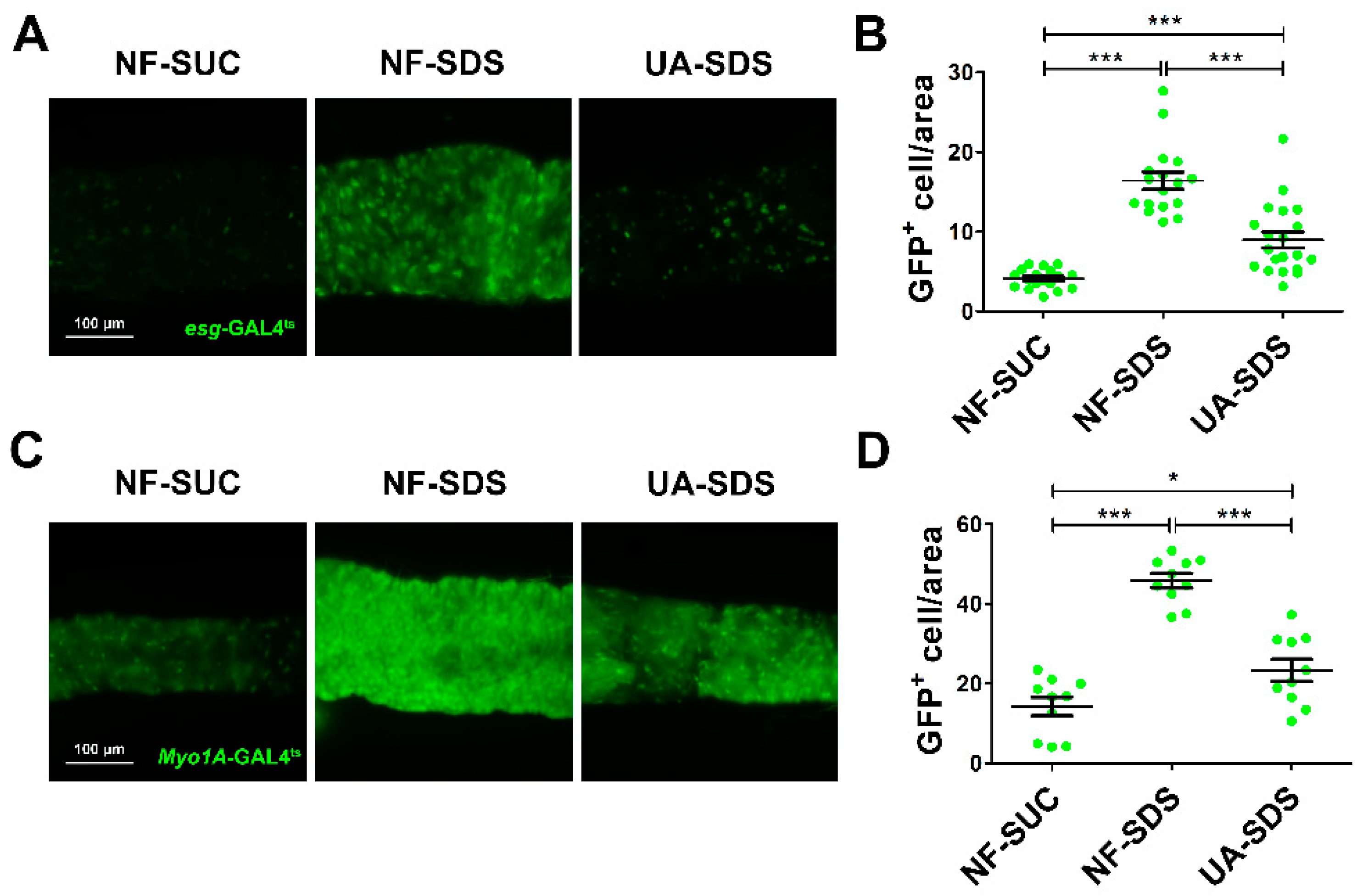
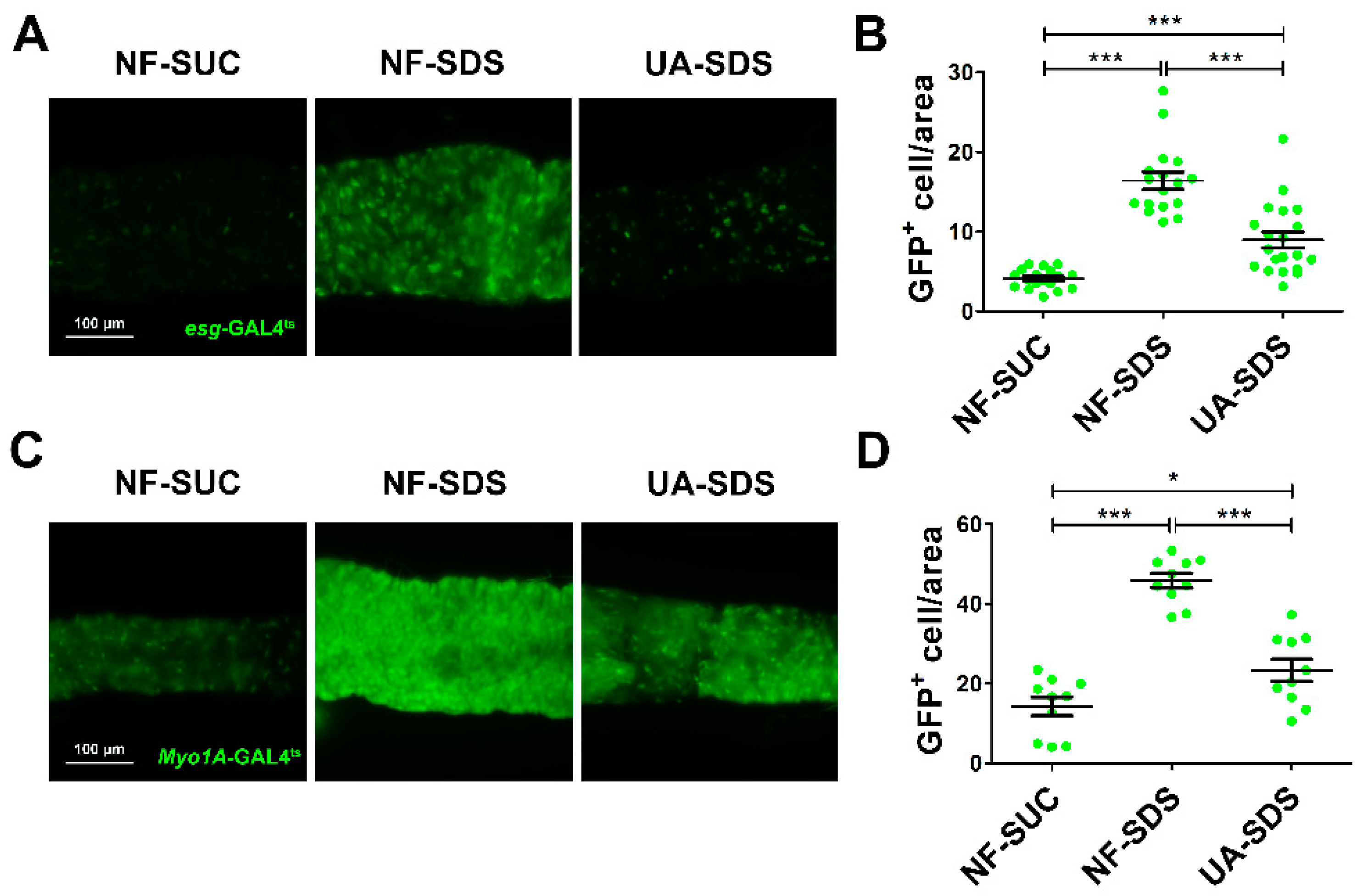
3.3. UA Protected Intestinal Stem Cells from SDS-Induced Proliferation and Differentiation
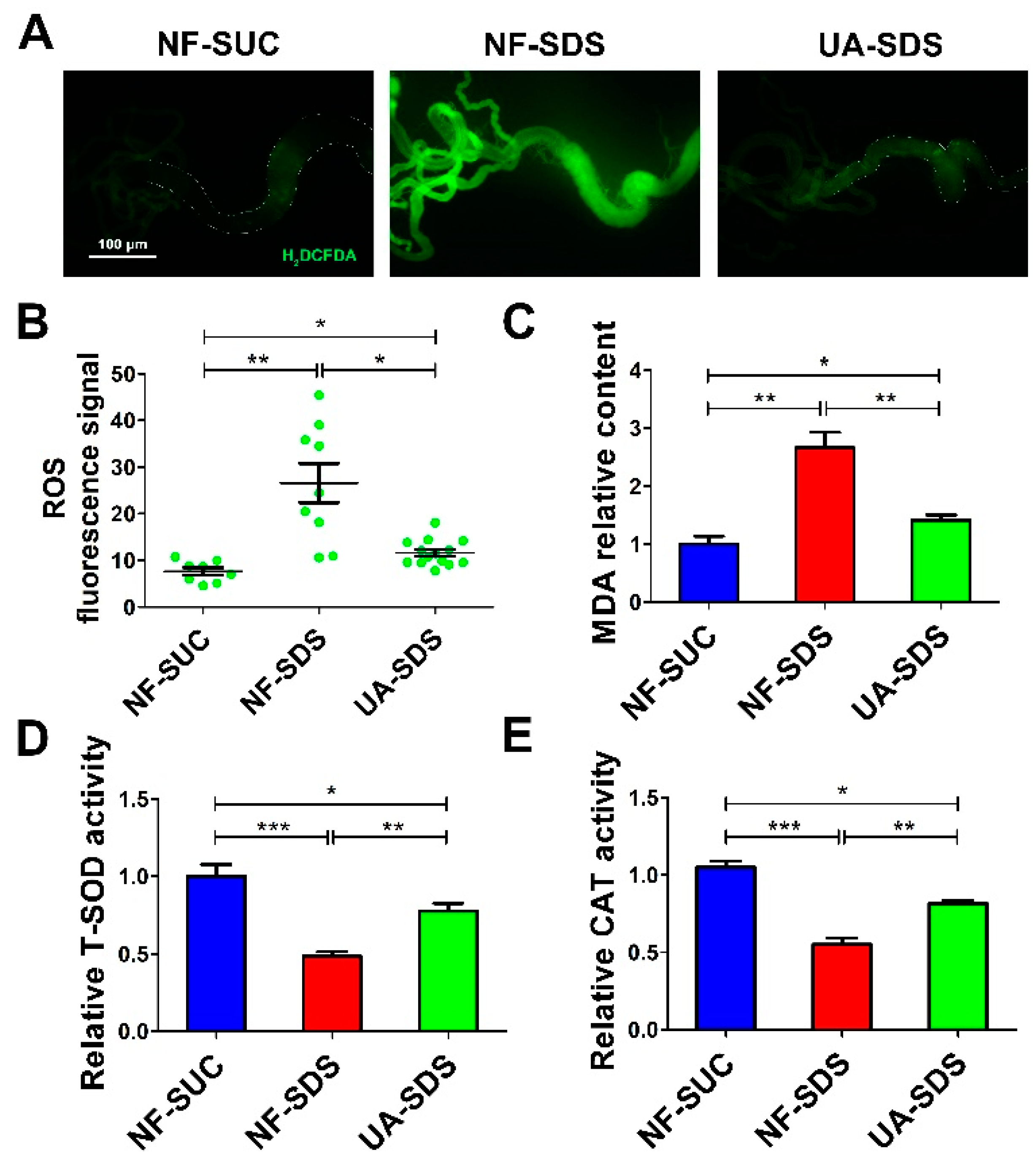
3.4. UA Protects Intestine against SDS-Induced Oxidative Damage
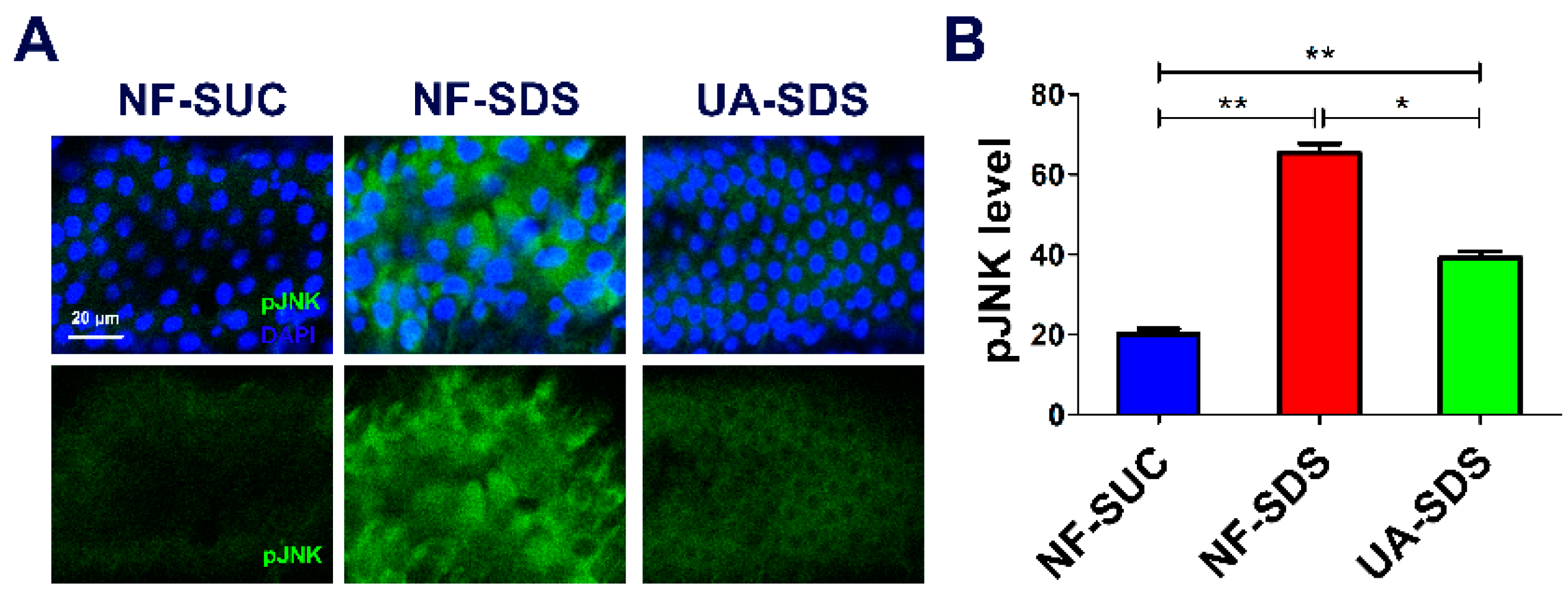
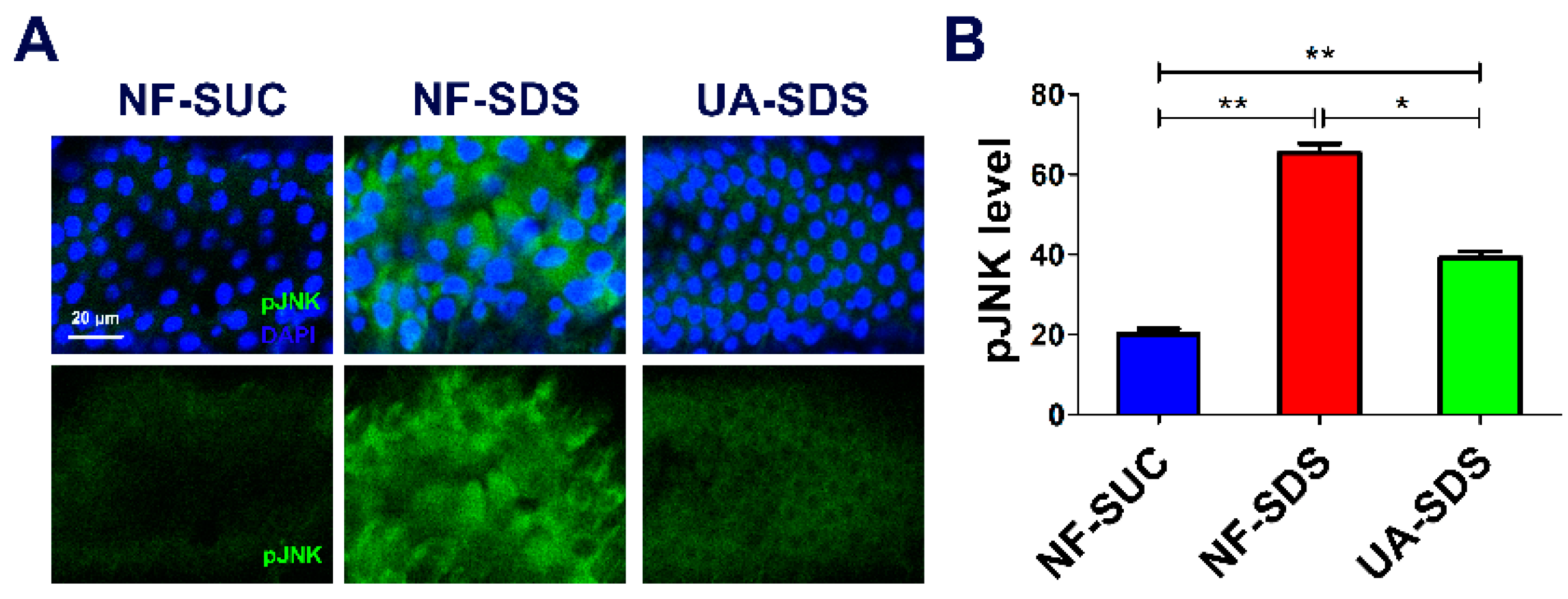
3.5. UA Protects against SDS-Induced Intestine Damage by Regulating JNK Signaling Pathway
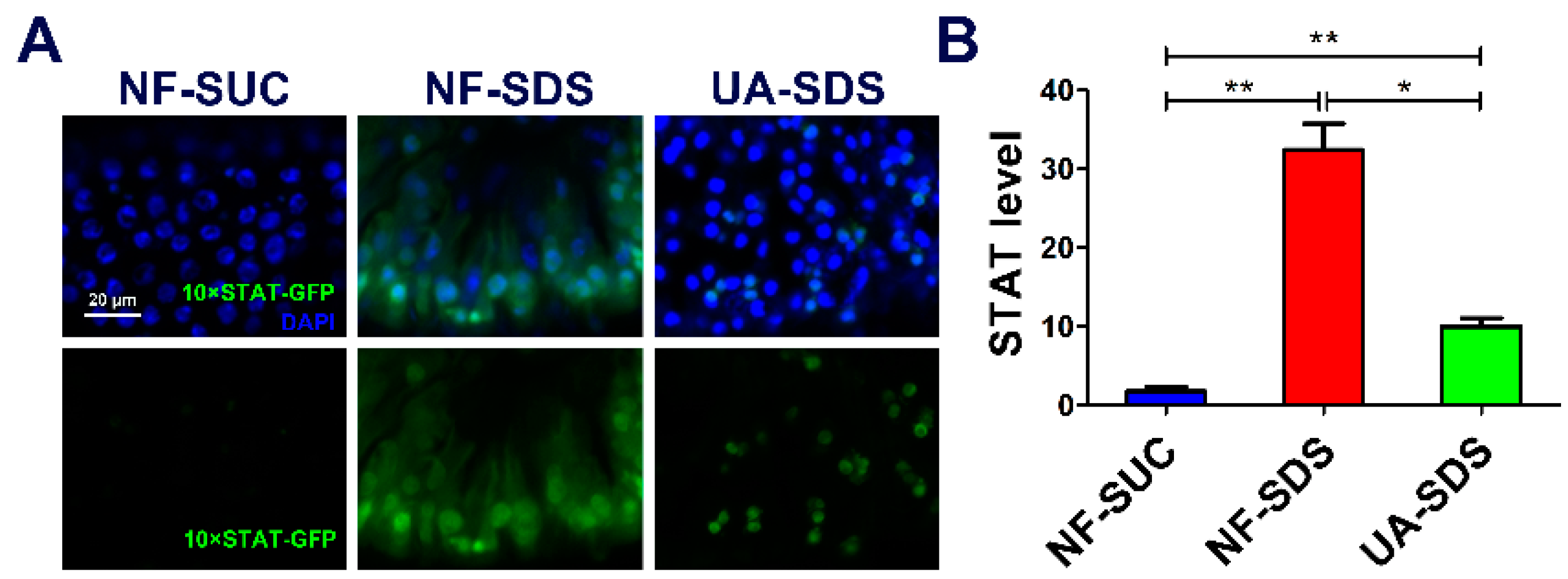
3.6. JAK/STAT Pathway Is Involved in the Protection of UA on Drosophila Intestine Homeostasis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cosnes, J.; Gower-Rousseau, C.; Seksik, P.; Cortot, A. Epidemiology and natural history of inflammatory bowel diseases. Gastroenterology 2011, 140, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Shou, J.; Wang, J.; Hu, W.; Hannan, F.; Mwamba, T.M.; Farooq, M.A.; Zhou, W.; Islam, F. Ursolic acid limits salt-induced oxidative damage by interfering with nitric oxide production and oxidative defense machinery in rice. Front. Plant Sci. 2020, 11, 697. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Facciorusso, A.; Dulai, P.S.; Jairath, V.; Sandborn, W.J. Comparative risk of serious infections with biologic and/or immunosuppressive therapy in patients with inflammatory bowel diseases: A systematic review and meta-analysis. Clin. Gastroenterol. Hepatol. 2020, 18, 69–81.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, W.; Garud, N.; Joshi, R. Role of inulin as prebiotics on inflammatory bowel disease. Drug Discov. Ther. 2019, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Radtke, F.; Clevers, H. Self-renewal and cancer of the gut: Two sides of a coin. Science 2005, 307, 1904–1909. [Google Scholar] [CrossRef] [Green Version]
- Raphael, T.J.; Kuttan, G. Effect of naturally occurring triterpenoids glycyrrhizic acid, ursolic acid, oleanolic acid and nomilin on the immune system. Phytomedicine 2003, 10, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, Y.; Murakami, A.; Ohigashi, H. Ursolic acid: An anti- and pro-inflammatory, triterpenoid. Mol. Nutr. Food Res. 2008, 52, 26–42. [Google Scholar] [CrossRef]
- Checker, R.; Sandur, S.K.; Sharma, D.; Patwardhan, R.S.; Jayakumar, S.; Kohli, V.; Sethi, G.; Aggarwal, B.B.; Sainis, K.B. Potent anti-inflammatory activity of ursolic acid, a triterpenoid antioxidant, is mediated through suppression of NF-kappaB, AP-1 and NF-AT. PLoS ONE 2012, 7, e31318. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.S.; Ibrahim, S.A.; Jalil, S.; Choudhary, M.I. Ursolic acid: A potent inhibitor of superoxides produced in the cellular system. Phytother. Res. 2007, 21, 558–561. [Google Scholar] [CrossRef]
- Yeh, C.T.; Wu, C.H.; Yen, G.C. Ursolic acid, a naturally occurring triterpenoid, suppresses migration and invasion of human breast cancer cells by modulating c-Jun N-terminal kinase, Akt and mammalian target of rapamycin signaling. Mol. Nutr. Food Res. 2010, 54, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.M.; Lin, J.Y. Anti-inflammatory effects of 27 selected terpenoid compounds tested through modulating Th1/Th2 cytokine secretion profiles using murine primary splenocytes. Food Chem. 2013, 141, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, P.G.G.; Lemos, T.L.G.; Bizerra, A.M.C.; Arriaga, A.M.C.; Ferreira, D.A.; Santiago, G.M.P.; Braz Filho, R.; Costa, J.G.M. Antibacterial and antioxidant activities of ursolic acid and derivatives. Molecules 2014, 19, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Leng, Y.; Liu, D.; Liu, X.; Ren, Y.; Zhang, J.; Chen, F. Research advances in protective effects of ursolic acid and oleanolic acid against gastrointestinal diseases. Am. J. Chin. Med. 2021, 49, 413–435. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.Z.; Liu, C.; Huang, C.K.; Luo, F.Y.; Zhu, X. Ursolic acid improves intestinal damage and bacterial dysbiosis in liver fibrosis mice. Front. Pharmacol. 2019, 10, 1321. [Google Scholar] [CrossRef]
- Amcheslavsky, A.; Jiang, J.; Ip, Y.T. Tissue damage-induced intestinal stem cell division in Drosophila. Cell Stem Cell 2009, 4, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liang, X.; Yang, X. Ursolic acid inhibits growth and induces apoptosis in gemcitabine-resistant human pancreatic cancer via the JNK and PI3K/Akt/NF-kappaB pathways. Oncol. Rep. 2012, 28, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Staats, S.; Wagner, A.E.; Luersen, K.; Kunstner, A.; Meyer, T.; Kahns, A.K.; Derer, S.; Graspeuntner, S.; Rupp, J.; Busch, H.; et al. Dietary ursolic acid improves health span and life span in male Drosophila melanogaster. Biofactors 2019, 45, 169–186. [Google Scholar] [CrossRef]
- Wang, E.M.; Fan, Q.L.; Yue, Y.; Xu, L. Ursolic acid attenuates high glucose-mediated mesangial cell injury by inhibiting the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin (PI3K/Akt/mTOR) signaling pathway. Med. Sci. Monit. 2018, 24, 846–854. [Google Scholar] [CrossRef]
- Liu, B.H.; Piao, X.H.; Guo, L.Y.; Liu, S.S.; Chai, F.; Gao, L.M. Ursolic acid protects against ulcerative colitis via anti-inflammatory and antioxidant effects in mice. Mol. Med. Rep. 2016, 13, 4779–4785. [Google Scholar] [CrossRef] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.; Zheng, P.Y.; Xia, L.; Yong, Y.; Lu, G.F.; Tang, F.; Zhao, Z.G. Calycosin attenuates dextran sulfate sodium (DSS)-induced experimental colitis. Iran J. Basic Med. Sci. 2017, 20, 1056–1062. [Google Scholar] [PubMed]
- Redhai, S.; Boutros, M. The role of organelles in intestinal function, physiology, and disease. Trends Cell Biol. 2021, 31, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, X.; Xi, R. EGFR and Notch signaling respectively regulate proliferative activity and multiple cell lineage differentiation of Drosophila gastric stem cells. Cell Res. 2014, 24, 610–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.S.; Ma, H.Y.; Shi, W.; Duan, J.L.; Wang, Y.; Zhang, C.T.; Li, C.L.; Lin, J.; Li, S.; Lv, J.G.; et al. Inhibition of STAT3 signaling pathway by ursolic acid suppresses growth of hepatocellular carcinoma. Int. J. Oncol. 2017, 51, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Model. Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Xiao, G.; Zhou, B. What can flies tell us about zinc homeostasis? Arch. Biochem. Biophys. 2016, 611, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Lv, J.; Xi, R. The specification and function of enteroendocrine cells in Drosophila and mammals: A comparative review. FEBS J. 2021. Epub ahead of print. [Google Scholar] [CrossRef]
- Hung, R.J.; Li, J.S.S.; Liu, Y.; Perrimon, N. Defining cell types and lineage in the Drosophila midgut using single cell transcriptomics. Curr. Opin. Insect Sci. 2021, 47, 12–17. [Google Scholar] [CrossRef]
- Boumard, B.; Bardin, A.J. An amuse-bouche of stem cell regulation: Underlying principles and mechanisms from adult Drosophila intestinal stem cells. Curr. Opin. Cell Biol. 2021, 73, 58–68. [Google Scholar] [CrossRef]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apidianakis, Y.; Rahme, L.G. Drosophila melanogaster as a model for human intestinal infection and pathology. Dis. Model Mech. 2011, 4, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, R.; Gunarta, I.K.; Boldbaatar, J.; Erdenebaatar, P.; Odongoo, R.; Yoshioka, K. Functional role of c-Jun NH2-terminal kinase-associated leucine zipper protein (JLP) in lysosome localization and autophagy. Drug Discov. Ther. 2020, 14, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liu, Z.; Chen, Y.; Jin, L.H. Identification of the protective effects of traditional medicinal plants against SDS-induced Drosophila gut damage. Exp. Ther. Med. 2016, 12, 2671–2680. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Chen, Y.; Zhang, H.; Jin, L.H. Crocus sativus L. protects against SDS-induced intestinal damage and extends lifespan in Drosophila melanogaster. Mol. Med. Rep. 2016, 14, 5601–5606. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, S.H.; Jin, L. Acanthopanax senticosus polysaccharide regulates the intestinal homeostasis disruption induced by toxic chemicals in Drosophila. Phytother. Res. 2020, 34, 193–200. [Google Scholar] [CrossRef]
- Zang, Y.R.; Wan, M.; Liu, M.; Ke, H.M.; Ma, S.C.; Liu, L.P.; Ni, J.Q.; Pastor-Pareja, J.C. Plasma membrane overgrowth causes fibrotic collagen accumulation and immune activation in Drosophila adipocytes. eLife 2015, 4, e07187. [Google Scholar] [CrossRef]
- Wei, T.; Ji, X.W.; Xue, J.S.; Gao, Y.; Zhu, X.M.; Xiao, G.R. Cyanidin-3-O-glucoside represses tumor growth and invasion in vivo by suppressing autophagy via inhibition of the JNK signaling pathways. Food Funct. 2021, 12, 387–396. [Google Scholar] [CrossRef]
- Xiao, G.; Fan, Q.; Wang, X.; Zhou, B. Huntington disease arises from a combinatory toxicity of polyglutamine and copper binding. Proc. Natl. Acad. Sci. USA 2013, 110, 14995–15000. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.Y.; Yang, Q.; Chen, X.; Tian, Y.Q.; Liu, Z.Y.; Wang, S.Y. Bioactive peptides derived from crimson snapper and in vivo anti-aging effects on fat diet-induced high fat Drosophila melanogaster. Food Funct. 2020, 11, 524–533. [Google Scholar] [CrossRef]
- Xiao, G.R.; Wan, Z.H.; Fan, Q.W.; Tang, X.N.; Zhou, B. The metal transporter ZIP13 supplies iron into the secretory pathway in Drosophila melanogaster. eLife 2014, 3, e03191. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Hong, J.; Xiao, G.; Feng, Y.; Peng, R.; Wang, M.; Qu, H. Descemet stripping endothelial keratoplasty in pediatric patients with congenital hereditary endothelial dystrophy. Am. J. Ophthalmol. 2019, 209, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Lee, C.; Heo, S.; Kim, B.; Hyun, C.K. DSS-induced colitis is associated with adipose tissue dysfunction and disrupted hepatic lipid metabolism leading to hepatosteatosis and dyslipidemia in mice. Sci. Rep. 2021, 11, 5283. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.H.; Penalva, C.; Kardorff, M.; Roca, M.; Pavlovic, B.; Thiel, A.; Teleman, A.A.; Edgar, B.A. Damage sensing by a Nox-Ask1-MKK3-p38 signaling pathway mediates regeneration in the adult Drosophila midgut. Nat. Commun. 2019, 10, 4365. [Google Scholar] [CrossRef] [Green Version]
- Dambroise, E.; Monnier, L.; Ruisheng, L.; Aguilaniu, H.; Joly, J.S.; Tricoire, H.; Rera, M. Two phases of aging separated by the Smurf transition as a public path to death. Sci. Rep. 2016, 6, 23523. [Google Scholar] [CrossRef] [Green Version]
- Kux, K.; Pitsouli, C. Tissue communication in regenerative inflammatory signaling: Lessons from the fly gut. Front. Cell Infect. Microbiol. 2014, 4, 49. [Google Scholar] [CrossRef] [Green Version]
- Loza-Coll, M.A.; Southall, T.D.; Sandall, S.L.; Brand, A.H.; Jones, D.L. Regulation of Drosophila intestinal stem cell maintenance and differentiation by the transcription factor Escargot. EMBO J. 2014, 33, 2983–2996. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Z.; Boquete, J.P.; Lemaitre, B. Cell-specific imd-nf-kappab responses enable simultaneous antibacterial immunity and intestinal epithelial cell shedding upon bacterial infection. Immunity 2018, 48, 897–910.e7. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Fang, R.; Wang, H.; Xu, D.X.; Yang, J.; Huang, X.; Cozzolino, D.; Fang, M.; Huang, Y. A review of environmental metabolism disrupting chemicals and effect biomarkers associating disease risks: Where exposomics meets metabolomics. Environ. Int. 2022, 158, 106941. [Google Scholar] [CrossRef]
- Morris, O.; Jasper, H. Reactive oxygen species in intestinal stem cell metabolism, fate and function. Free Radic. Biol. Med. 2021, 166, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, W.; Zheng, L.; Zhang, B.; Yang, X.; Zhang, Q.; Wang, N.; Wang, Y.; Yang, J.; Sha, J.; et al. Ameliorative effect of ursolic acid on ochratoxin A-induced renal cytotoxicity mediated by Lonp1/Aco2/Hsp75. Toxicon 2019, 168, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical relevance of biomarkers of oxidative stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeeva, J.S.; Sunitha, J.; Ananthalakshmi, R.; Rajkumari, S.; Ramesh, M.; Krishnan, R. Enzymatic antioxidants and its role in oral diseases. J. Pharm. Bioallied Sci. 2015, 7, S331–S333. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.M.; Liu, Z.G. JNK signaling pathway is a key modulator in cell death mediated by reactive oxygen and nitrogen species. Free Radic. Bio. Med. 2006, 40, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Kurata, S.; Yano, T. Immunoglobulin superfamily beat-Ib mediates intestinal regeneration induced by reactive oxygen species in Drosophila. Genes Cells 2020, 25, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Garlick, D.S.; Greiner, D.L.; Davis, R.J. The role of JNK in the development of hepatocellular carcinoma. Gene Dev. 2011, 25, 634–645. [Google Scholar] [CrossRef] [Green Version]
- Ahmed-de-Prado, S.; Diaz-Garcia, S.; Baonza, A. JNK and JAK/STAT signalling are required for inducing loss of cell fate specification during imaginal wing discs regeneration in Drosophila melanogaster. Dev. Biol. 2018, 441, 31–41. [Google Scholar] [CrossRef]
- Tian, A.G.; Shi, Q.; Jiang, A.; Li, S.X.; Wang, B.; Jiang, J. Injury-stimulated Hedgehog signaling promotes regenerative proliferation of Drosophila intestinal stem cells. J. Cell Biol. 2015, 208, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Zheng, F.; Goncalves, F.M.; Abiko, Y.; Li, H.; Kumagai, Y.; Aschner, M. Redox toxicology of environmental chemicals causing oxidative stress. Redox Biol. 2020, 34, 101475. [Google Scholar] [CrossRef] [PubMed]
- Wisidagama, D.R.; Thummel, C.S. Regulation of Drosophila intestinal stem cell proliferation by enterocyte mitochondrial pyruvate metabolism. G3 Genes Genom. Genet. 2019, 9, 3623–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.Q.; Tian, A.G.; Jiang, J. Intestinal stem cell response to injury: Lessons from Drosophila. Cell Mol. Life Sci. 2016, 73, 3337–3349. [Google Scholar] [CrossRef] [PubMed]
- Naszai, M.; Carroll, L.R.; Cordero, J.B. Intestinal stem cell proliferation and epithelial homeostasis in the adult Drosophila midgut. Insect Biochem. Mol. Biol. 2015, 67, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Herrera, S.C.; Bach, E.A. JAK/STAT signaling in stem cells and regeneration: From Drosophila to vertebrates. Development 2019, 146, dev167643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beira, J.V.; Torres, J.; Paro, R. Signalling crosstalk during early tumorigenesis in the absence of Polycomb silencing. PLoS Genet. 2018, 14, e1007187. [Google Scholar] [CrossRef]
- Shaw, R.L.; Kohlmaier, A.; Polesello, C.; Veelken, C.; Edgar, B.A.; Tapon, N. The Hippo pathway regulates intestinal stem cell proliferation during Drosophila adult midgut regeneration. Development 2010, 137, 4147–4158. [Google Scholar] [CrossRef] [Green Version]
- Santabarbara-Ruiz, P.; Lopez-Santillan, M.; Martinez-Rodriguez, I.; Binagui-Casas, A.; Perez, L.; Milan, M.; Corominas, M.; Serras, F. ROS-induced JNK and p38 signaling is required for unpaired cytokine activation during Drosophila regeneration. PLoS Genet. 2015, 11, e1005595. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, T.; Wu, L.; Ji, X.; Gao, Y.; Xiao, G. Ursolic Acid Protects Sodium Dodecyl Sulfate-Induced Drosophila Ulcerative Colitis Model by Inhibiting the JNK Signaling. Antioxidants 2022, 11, 426. https://doi.org/10.3390/antiox11020426
Wei T, Wu L, Ji X, Gao Y, Xiao G. Ursolic Acid Protects Sodium Dodecyl Sulfate-Induced Drosophila Ulcerative Colitis Model by Inhibiting the JNK Signaling. Antioxidants. 2022; 11(2):426. https://doi.org/10.3390/antiox11020426
Chicago/Turabian StyleWei, Tian, Lei Wu, Xiaowen Ji, Yan Gao, and Guiran Xiao. 2022. "Ursolic Acid Protects Sodium Dodecyl Sulfate-Induced Drosophila Ulcerative Colitis Model by Inhibiting the JNK Signaling" Antioxidants 11, no. 2: 426. https://doi.org/10.3390/antiox11020426
APA StyleWei, T., Wu, L., Ji, X., Gao, Y., & Xiao, G. (2022). Ursolic Acid Protects Sodium Dodecyl Sulfate-Induced Drosophila Ulcerative Colitis Model by Inhibiting the JNK Signaling. Antioxidants, 11(2), 426. https://doi.org/10.3390/antiox11020426