
Interaction between the Effects of Sustained Swimming Activity and Dietary Macronutrient Proportions on the Redox Status of Gilthead Sea Bream Juveniles (Sparus aurata L.)

 , ,
, ,  ,
,  , ,
, ,
Abstract
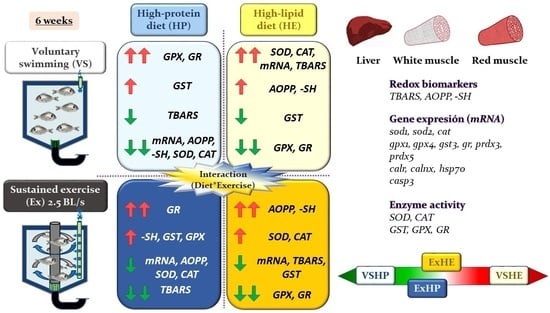
:
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Conditions
2.2. Sampling
2.3. Biochemical Markers and Enzyme Activity
2.4. RNA Extraction, cDNA Synthesis and Real-Time Quantitative PCR (qPCR)
2.5. Statistics
3. Results
3.1. Redox Metabolites
3.2. Relative Gene Expression
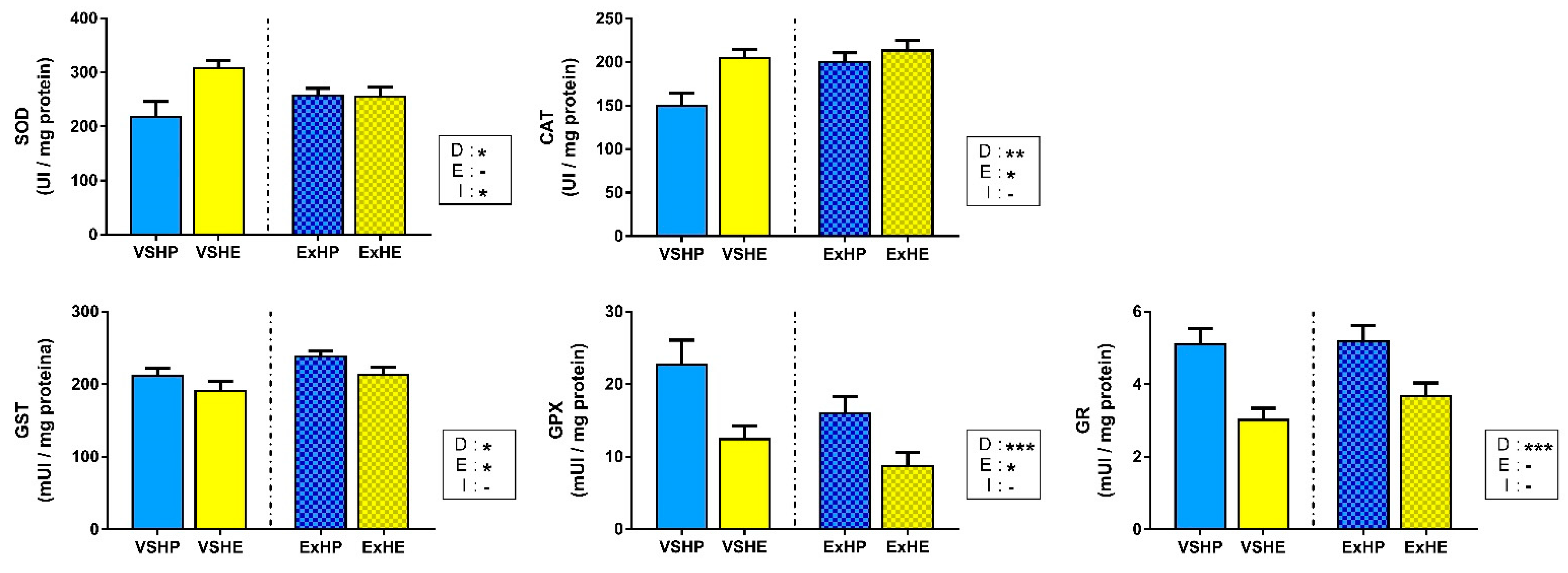
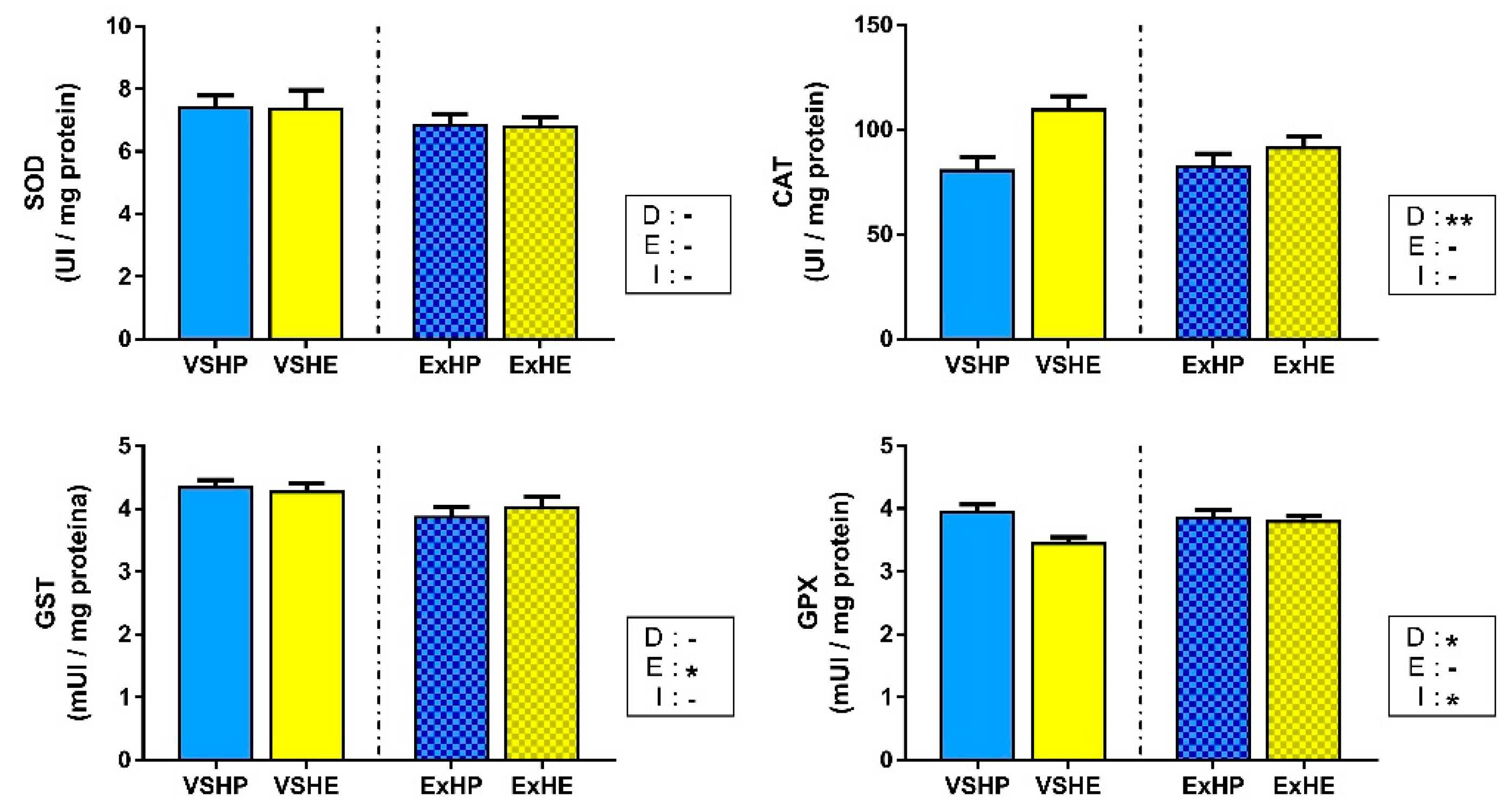
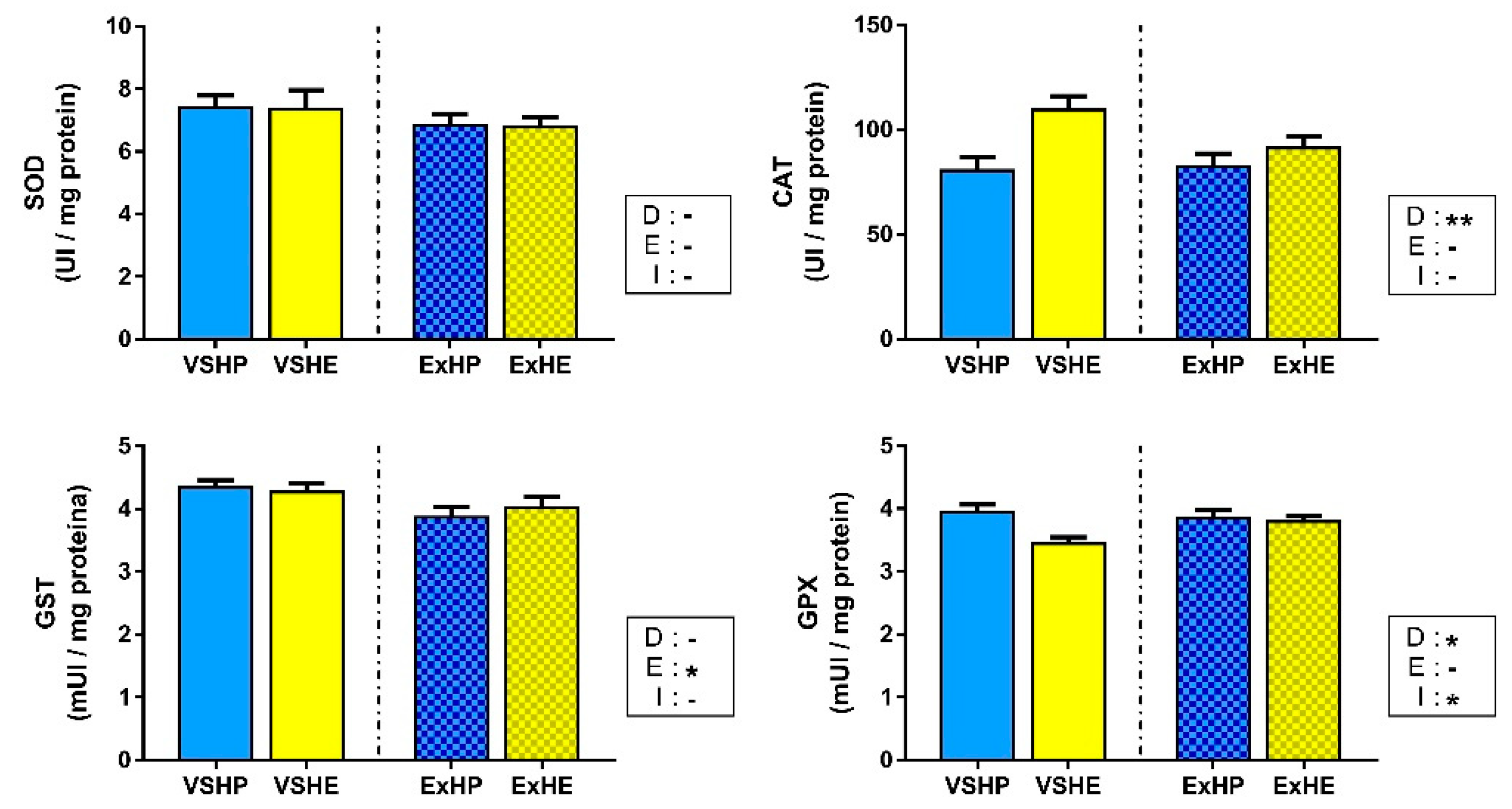
3.3. Enzyme Activity
4. Discussion
4.1. Effect of Diet on Oxidative Stress Status
4.2. Effect of Exercise on the Oxidative Stress Status
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Alvarez, M.J.; Lopez-Bote, C.J.; Diez, A.; Corraze, G.; Arzel, J.; Dias, J.; Kaushik, S.J.; Bautista, J.M. Dietary fish oil and digestible protein modify susceptibility to lipid peroxidation in the muscle of rainbow trout (Oncorhynchus mykiss) and sea bass (Dicentrarchus labrax). Br. J. Nutr. 1998, 80, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Rueda-Jasso, R.; Conceição, L.E.C.; Dias, J.; de Coen, W.; Gomes, E.; Rees, J.F.; Soares, F.; Dinis, M.T.; Sorgeloos, P. Effect of dietary non-protein energy levels on condition and oxidative status of Senegalese sole (Solea senegalensis) juveniles. Aquaculture 2004, 231, 417–433. [Google Scholar] [CrossRef]
- Pérez-Jiménez, A.; Hidalgo, M.C.; Morales, A.E.; Arizcun, M.; Abellán, E.; Cardenete, G. Antioxidant enzymatic defenses and oxidative damage in Dentex dentex fed on different dietary macronutrient levels. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 150, 537–545. [Google Scholar] [CrossRef]
- Li, X.; Jiang, Y.; Liu, W.; Ge, X. Protein-sparing effect of dietary lipid in practical diets for blunt snout bream (Megalobrama amblycephala) fingerlings: Effects on digestive and metabolic responses. Fish Physiol. Biochem. 2012, 38, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Karaman, Y.K.; Novgorodtseva, T.P.; Yan’Kova, V.I. Effects on Alimentary High-Fat Diet on Thiol Disulfide Homeostasis in Rats. Bull. Exp. Biol. Med. 2013, 155, 752–756. [Google Scholar] [CrossRef]
- Sánchez-Nuño, S.; Carbonell, T.; Valls, A.I. Redox Balance Affects Fish Welfare. In Redox; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Magalhães, R.; Guerreiro, I.; Santos, R.A.; Coutinho, F.; Couto, A.; Serra, C.R.; Olsen, R.E.; Peres, H.; Oliva-Teles, A. Oxidative status and intestinal health of gilthead sea bream (Sparus aurata) juveniles fed diets with different ARA/EPA/DHA ratios. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Burneiko, R.C.M.; Diniz, Y.S.; Galhardi, C.M.; Rodrigues, H.G.; Ebaid, G.M.X.; Faine, L.A.; Padovani, C.R.; Cicogna, A.C.; Novelli, E.L.B. Interaction of hypercaloric diet and physical exercise on lipid profile, oxidative stress and antioxidant defenses. Food Chem. Toxicol. 2006, 44, 1167–1172. [Google Scholar] [CrossRef]
- Rindler, P.M.; Plafker, S.M.; Szweda, L.I.; Kinter, M. High Dietary Fat Selectively Increases Catalase Expression within Cardiac Mitochondria. J. Biol. Chem. 2013, 288, 1979–1990. [Google Scholar] [CrossRef] [Green Version]
- Lasker, S.; Rahman, M.M.; Parvez, F.; Zamila, M.; Miah, P.; Nahar, K.; Kabir, F.; Sharmin, S.B.; Subhan, N.; Ahsan, G.U.; et al. High-fat diet-induced metabolic syndrome and oxidative stress in obese rats are ameliorated by yogurt supplementation. Sci. Rep. 2019, 9, 20026. [Google Scholar] [CrossRef]
- Sánchez, J.A.; Lopez-Olmeda, J.F.; Blanco-Vives, B.; Vázquez, F.J.S. Effects of feeding schedule on locomotor activity rhythms and stress response in sea bream. Physiol. Behav. 2009, 98, 125–129. [Google Scholar] [CrossRef]
- Zhang, Y.; Timmerhaus, G.; Anttila, K.; Mauduit, F.; Jørgensen, S.M.; Kristensen, T.; Claireaux, G.; Takle, H.; Farrell, A.P. Domestication compromises athleticism and respiratory plasticity in response to aerobic exercise training in Atlantic salmon (Salmo salar). Aquaculture 2016, 463, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Martin-Perez, M.; Fernandez-Borras, J.; Ibarz, A.; Millan-Cubillo, A.; Felip, O.; de Oliveira, E.; Blasco, J. New Insights into Fish Swimming: A Proteomic and Isotopic Approach in Gilthead Sea Bream. J. Proteome Res. 2012, 11, 3533–3547. [Google Scholar] [CrossRef] [PubMed]
- Blasco, J.; Moya, A.; Millán-Cubillo, A.; Vélez, E.J.; Capilla, E.; Pérez-Sánchez, J.; Gutierrez, J.; Fernandez-Borras, J. Growth-promoting effects of sustained swimming in fingerlings of gilthead sea bream (Sparus aurata L.). J. Comp. Physiol. B 2015, 185, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez, E.J.; Azizi, S.; Lutfi, E.; Capilla, E.; Moya, A.; Navarro, I.; Fernández-Borràs, J.; Blasco, J.; Gutiérrez, J. Moderate and sustained exercise modulates muscle proteolytic and myogenic markers in gilthead sea bream (Sparus aurata). Am. J. Physiol. Integr. Comp. Physiol. 2017, 312, R643–R653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moya, A.; Torrella, J.R.; Fernández-Borràs, J.; Rizo-Roca, D.; Millán-Cubillo, A.; Vélez, E.J.; Arcas, A.; Gutiérrez, J.; Blasco, J. Sustained swimming enhances white muscle capillarisation and growth by hyperplasia in gilthead sea bream (Sparus aurata) fingerlings. Aquaculture 2019, 501, 397–403. [Google Scholar] [CrossRef]
- Perelló-Amorós, M.; Fernández-Borràs, J.; Sánchez-Moya, A.; Vélez, E.J.; García-Pérez, I.; Gutiérrez, J.; Blasco, J. Mitochondrial Adaptation to Diet and Swimming Activity in Gilthead Seabream: Improved Nutritional Efficiency. Front. Physiol. 2021, 12, 875. [Google Scholar] [CrossRef]
- Perelló-Amorós, M.; García-Pérez, I.; Sánchez-Moya, A.; Innamorati, A.; Vélez, E.J.; Achaerandio, I.; Pujolà, M.; Calduch-Giner, J.; Pérez-Sánchez, J.; Fernández-Borràs, J.; et al. Diet and Exercise Modulate GH-IGFs Axis, Proteolytic Markers and Myogenic Regulatory Factors in Juveniles of Gilthead Sea Bream (Sparus aurata). Animals 2021, 11, 2182. [Google Scholar] [CrossRef] [PubMed]
- Felip, O.; Blasco, J.; Ibarz, A.; Martin-Perez, M.; Fernández-Borràs, J. Beneficial effects of sustained activity on the use of dietary protein and carbohydrate traced with stable isotopes 15N and 13C in gilthead sea bream (Sparus aurata). J. Comp. Physiol. B 2013, 183, 223–234. [Google Scholar] [CrossRef]
- Magnoni, L.J.; Felip, O.; Blasco, J.; Planas, J.V. Metabolic Fuel Utilization During Swimming: Optimizing Nutritional Requirements for Enhanced Performance. In Swimming Physiology of Fish; Springer: Berlin/Heidelberg, Germany, 2013; pp. 203–235. [Google Scholar]
- Castro, V.; Grisdale-Helland, B.; Helland, S.J.; Kristensen, T.; Jørgensen, S.M.; Helgerud, J.; Claireaux, G.; Farrell, A.P.; Krasnov, A.; Takle, H. Aerobic training stimulates growth and promotes disease resistance in Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 160, 278–290. [Google Scholar] [CrossRef]
- McKenzie, D.J.; Palstra, A.P.; Planas, J.; MacKenzie, S.; Bégout, M.; Thorarensen, H.; Vandeputte, M.; Mes, D.; Rey, S.; de Boeck, G.; et al. Aerobic swimming in intensive finfish aquaculture: Applications for production, mitigation and selection. Rev. Aquac. 2021, 13, 138–155. [Google Scholar] [CrossRef]
- Skov, P.V.; Lund, I.; Pargana, A.M. No evidence for a bioenergetic advantage from forced swimming in rainbow trout under a restrictive feeding regime. Front. Physiol. 2015, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palstra, A.P.; Mendez, S.; Dirks, R.P.; Schaaf, M.J.M. Cortisol Acting Through the Glucocorticoid Receptor Is Not Involved in Exercise-Enhanced Growth, But Does Affect the White Skeletal Muscle Transcriptome in Zebrafish (Danio rerio). Front. Physiol. 2019, 9, 1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pengam, M.; Moisan, C.; Simon, B.; Guernec, A.; Inizan, M.; Amérand, A. Training protocols differently affect AMPK–PGC-1α signaling pathway and redox state in trout muscle. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 243, 110673. [Google Scholar] [CrossRef] [PubMed]
- Pengam, M.; Amérand, A.; Simon, B.; Guernec, A.; Inizan, M.; Moisan, C. How do exercise training variables stimulate processes related to mitochondrial biogenesis in slow and fast trout muscle fibres? Exp. Physiol. 2021, 106, 938–957. [Google Scholar] [CrossRef]
- Weber, J.-M.; Choi, K.; Gonzalez, A.; Omlin, T. Metabolic fuel kinetics in fish: Swimming, hypoxia and muscle membranes. J. Exp. Biol. 2016, 219, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Palstra, A.P.; Roque, A.; Kruijt, L.; Jéhannet, P.; Pérez-Sánchez, J.; Dirks, R.P. Physiological Effects of Water Flow Induced Swimming Exercise in Seabream Sparus aurata. Front. Physiol. 2020, 11, 610049. [Google Scholar] [CrossRef]
- Altringham, J.D.; Ellerby, D.J. Fish swimming: Patterns in muscle function. J. Exp. Biol. 1999, 202, 3397–3403. [Google Scholar] [CrossRef]
- Teulier, L.; Thoral, E.; Queiros, Q.; McKenzie, D.J.; Roussel, D.; Dutto, G.; Gasset, E.; Bourjea, J.; Saraux, C. Muscle bioenergetics of two emblematic Mediterranean fish species: Sardina pilchardus and Sparus aurata. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 235, 174–179. [Google Scholar] [CrossRef]
- McClelland, G.B.; Scott, G.R. Muscle plasticity. In The Physiology of Fishes; Evans, D.H., Claiborne, J.B., Currie, S., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–31. [Google Scholar]
- Loschen, G.; Azzi, A.; Richter, C.; Flohé, L. Superoxide radicals as precursors of mitochondrial hydrogen peroxide. FEBS Lett. 1974, 42, 68–72. [Google Scholar] [CrossRef] [Green Version]
- St-Pierre, J.; Buckingham, J.A.; Roebuck, S.J.; Brand, M.D. Topology of Superoxide Production from Different Sites in the Mitochondrial Electron Transport Chain. J. Biol. Chem. 2002, 277, 44784–44790. [Google Scholar] [CrossRef] [Green Version]
- Powers, S.K.; Bomkamp, M.; Ozdemir, M.; Hyatt, H. Mechanisms of exercise-induced preconditioning in skeletal muscles. Redox Biol. 2020, 35, 101462. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Deminice, R.; Ozdemir, M.; Yoshihara, T.; Bomkamp, M.P.; Hyatt, H. Exercise-induced oxidative stress: Friend or foe? J. Sport Health Sci. 2020, 9, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Sánchez-Nuño, S.; Sanahuja, I.; Fernández-Alacid, L.; Ordóñez-Grande, B.; Carbonell, T.; Ibarz, A. Oxidative attack during temperature fluctuation challenge compromises liver protein homeostasis of a temperate fish model. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 236, 110311. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Gausson, R.; Nguyen, A.-T.; Touam, M.; Drüeke, T.; Santangelo, F.; Descamps-Latscha, B. AOPP-induced activation of human neutrophil and monocyte oxidative metabolism: A potential target for N-acetylcysteine treatment in dialysis patients. Kidney Int. 2003, 64, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar] [CrossRef]
- Mannervik, B. The Isoenzymes of Glutathione Transferase. Adv. Enzymol. Relat. Areas Mol. Biol. 1985, 57, 357–417. [Google Scholar] [CrossRef]
- Bell, J.G.; Cowey, C.B.; Adron, J.W.; Shanks, A.M. Some effects of vitamin E and selenium deprivation on tissue enzyme levels and indices of tissue peroxidation in rainbow trout (Salmo gairdneri). Br. J. Nutr. 1985, 53, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagone, A.L.; Greenwald, J.; Kraut, E.H.; Bianchine, J.; Singh, D. Glucose: A role as a free radical scavenger in biological systems. J. Lab. Clin. Med. 1983, 101, 97–104. [Google Scholar] [PubMed]
- Fang, Y.-Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Liemburg-Apers, D.C.; Willems, P.H.G.M.; Koopman, W.J.H.; Grefte, S. Interactions between mitochondrial reactive oxygen species and cellular glucose metabolism. Arch. Toxicol. 2015, 89, 1209–1226. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Hong, C. Evaluation of malonaldehyde as a marker of oxidative rancidity in meat products. J. Food Biochem. 1991, 15, 97–105. [Google Scholar] [CrossRef]
- Zacarias, A.C.; Barbosa, M.A.; Guerra-Sá, R.; de Castro, U.G.M.; Bezerra, F.S.; de Lima, W.G.; Cardoso, L.M.; dos Santos, R.A.S.; Campagnole-Santos, M.J.; Alzamora, A.C. Swimming training induces liver adaptations to oxidative stress and insulin sensitivity in rats submitted to high-fat diet. Redox Rep. 2017, 22, 515–523. [Google Scholar] [CrossRef]
- Camiletti-Móiron, D.; Arianna Aparicio, V.; Nebot, E.; Medina, G.; Martínez, R.; Kapravelou, G.; Andrade, A.; Porres, J.M.; López-Jurado, M.; Aranda, P. High-protein diet induces oxidative stress in rat brain: Protective action of high-intensity exercise against lipid peroxidation. Nutr. Hosp. 2015, 31, 866–874. [Google Scholar] [CrossRef]
- Weisiger, R.A.; Fridovich, I. Mitochondrial Superoxide Dismutase. J. Biol. Chem. 1973, 248, 4793–4796. [Google Scholar] [CrossRef]
- Tyler, D.D. Polarographic assay and intracellular distribution of superoxide dismutase in rat liver. Biochem. J. 1975, 147, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Tsuboyama-Kasaoka, N.; Nakatani, T.; Ishii, M.; Tsutsumi, S.; Aburatani, H.; Ezaki, O. Fish oil feeding alters liver gene expressions to defend against PPARα activation and ROS production. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G338–G348. [Google Scholar] [CrossRef] [Green Version]
- Salvi, M.; Battaglia, V.; Brunati, A.M.; La Rocca, N.; Tibaldi, E.; Pietrangeli, P.; Marcocci, L.; Mondovi, B.; Rossi, C.A.; Toninello, A. Catalase Takes Part in Rat Liver Mitochondria Oxidative Stress Defense. J. Biol. Chem. 2007, 282, 24407–24415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarukamjorn, K.; Jearapong, N.; Pimson, C.; Chatuphonprasert, W. A High-Fat, High-Fructose Diet Induces Antioxidant Imbalance and Increases the Risk and Progression of Nonalcoholic Fatty Liver Disease in Mice. Scientifica 2016, 2016, 9060649. [Google Scholar] [CrossRef] [Green Version]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen Peroxide Sensing and Signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Kim, M.H.; Park, S.-J.; Kim, J.-H.; Seong, J.B.; Kim, K.-M.; Woo, H.A.; Lee, D.-S. Peroxiredoxin 5 regulates adipogenesis-attenuating oxidative stress in obese mouse models induced by a high-fat diet. Free Radic. Biol. Med. 2018, 123, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Grim, J.M.; Hyndman, K.A.; Kriska, T.; Girotti, A.W.; Crockett, E.L. Relationship between oxidizable fatty acid content and level of antioxidant glutathione peroxidases in marine fish. J. Exp. Biol. 2011, 214, 3751–3759. [Google Scholar] [CrossRef] [Green Version]
- Kapravelou, G.; Martínez, R.; Andrade, A.M.; Nebot, E.; Camiletti-Moirón, D.; Aparicio, V.A.; Lopez-Jurado, M.; Aranda, P.; Arrebola, F.; Fernandez-Segura, E.; et al. Aerobic interval exercise improves parameters of nonalcoholic fatty liver disease (NAFLD) and other alterations of metabolic syndrome in obese Zucker rats. Appl. Physiol. Nutr. Metab. 2015, 40, 1242–1252. [Google Scholar] [CrossRef] [Green Version]
- Wadley, A.J.; Aldred, S.; Coles, S.J. An unexplored role for Peroxiredoxin in exercise-induced redox signalling? Redox Biol. 2016, 8, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Cabrera, M.C.; Arc-Chagnaud, C.; Salvador-Pascual, A.; Brioche, T.; Chopard, A.; Olaso-Gonzalez, G.; Viña, J. Redox modulation of muscle mass and function. Redox Biol. 2020, 35, 101531. [Google Scholar] [CrossRef]
- Ihara, Y.; Kageyama, K.; Kondo, T. Overexpression of calreticulin sensitizes SERCA2a to oxidative stress. Biochem. Biophys. Res. Commun. 2005, 329, 1343–1349. [Google Scholar] [CrossRef]
- Gomez-Cabrera, M.C.; Carretero, A.; Millan-Domingo, F.; Garcia-Dominguez, E.; Correas, A.G.; Olaso-Gonzalez, G.; Viña, J. Redox-related biomarkers in physical exercise. Redox Biol. 2021, 42, 101956. [Google Scholar] [CrossRef]
- Zembron-Lacny, A.; Slowinska-Lisowska, M.; Ziemba, A. Integration of the Thiol Redox Status with Cytokine Response to Physical Training in Professional Basketball Players. Physiol. Res. 2010, 59, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, C.V.; Balakrishnan, S.D. Effect of training on lipid peroxidation, thiol status and antioxidant enzymes in tissues of rats. Indian J. Physiol. Pharmacol. 1998, 42, 64–70. [Google Scholar]
- Brooks, S.V.; Vasilaki, A.; Larkin, L.M.; McArdle, A.; Jackson, M.J. Repeated bouts of aerobic exercise lead to reductions in skeletal muscle free radical generation and nuclear factor κB activation. J. Physiol. 2008, 586, 3979–3990. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K.; Packer, L. Thiol homeostasis and supplements in physical exercise. Am. J. Clin. Nutr. 2000, 72, 653S–669S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elokda, A.S.; Nielsen, D.H. Effects of exercise training on the glutathione antioxidant system. Eur. J. Cardiovasc. Prev. Rehabil. 2007, 14, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Venditti, P.; di Meo, S. Effect of Training on Antioxidant Capacity, Tissue Damage, and Endurance of Adult Male Rats. Int. J. Sports Med. 1997, 18, 497–502. [Google Scholar] [CrossRef]
- Ibarz, A.; Felip, O.; Fernández-Borràs, J.; Martín-Pérez, M.; Blasco, J.; Torrella, J.R. Sustained swimming improves muscle growth and cellularity in gilthead sea bream. J. Comp. Physiol. B 2011, 181, 209–217. [Google Scholar] [CrossRef]
- Leaver, M.J.; George, S.G. A piscine glutathione S-transferase which efficiently conjugates the end-products of lipid peroxidation. Mar. Environ. Res. 1998, 46, 71–74. [Google Scholar] [CrossRef]
- Atalay, M.; Laaksonen, D.E.; Khanna, S.; Kaliste-Korhonen, E.; Hänninen, O.; Sen, C.K. Vitamin E regulates changes in tissue antioxidants induced by fish oil and acute exercise. Med. Sci. Sports Exerc. 2000, 32, 601–607. [Google Scholar] [CrossRef]
- Avula, C.P.R.; Fernandes, G. Modulation of antioxidant enzymes and apoptosis in mice by dietary lipids and treadmill exercise. J. Clin. Immunol. 1999, 19, 35–44. [Google Scholar] [CrossRef]
- Sen, C.K. Glutathione homeostasis in response to exercise training and nutritional supplements. Mol. Cell. Biochem. 1999, 196, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Lambertucci, R.H.; Levada-Pires, A.C.; Rossoni, L.V.; Curi, R.; Pithon-Curi, T.C. Effects of aerobic exercise training on antioxidant enzyme activities and mRNA levels in soleus muscle from young and aged rats. Mech. Ageing Dev. 2007, 128, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Ellgaard, L.; Frickel, E.-M. Calnexin, Calreticulin, and ERp57: Teammates in Glycoprotein Folding. Cell Biochem. Biophys. 2003, 35, 223–247. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HP DIET | HE DIET | |
|---|---|---|
| Digestible energy (MJ/kg) | 18 | 19.9 |
| Protein (% dry mass) | 54 | 50 |
| Lipids (% dry mass) | 15 | 20 |
| DHA (% dry mass) | 1 | 1.4 |
| EPA (% dry mass) | 2.5 | 3 |
| ARA (% dry mass) | 0.2 | 0.4 |
| DHA/EPA/ARA | 5/12.5/1 | 3.5/7.5/1 |
| Voluntary Swimming | Exercise | ANOVA | ||||||
|---|---|---|---|---|---|---|---|---|
| High protein (VSHP) | High lipid (VSHE) | High protein (ExHP) | High lipid (ExHE) | D | E | I | ||
| TBARS | 0.65 ± 0.06 | 0.99 ± 0.09 | 0.68 ± 0.06 | 0.9 ± 0.1 | *** | - | - | |
| Liver | AOPP | 3.32 ± 0.43 | 5.03 ± 0.4 | 4.64 ± 0.4 | 5.67 ± 0.31 | *** | * | - |
| -SH | 262.5 ± 19.1 | 368.7 ± 17.2 | 336.8 ± 18.1 | 385.4 ± 20.8 | *** | * | - | |
| TBARS | 0.43 ± 0.05 | 0.57 ± 0.06 | 0.37 ± 0.04 | 0.43 ± 0.04 | * | * | - | |
| White muscle | AOPP | 6.04 ± 1.19 | 7.9 ± 2.01 | 4.58 ± 0.63 | 8.18 ± 0.85 | * | - | - |
| -SH | 145.8 ± 3.7 | 137.8 ± 5.3 | 138.9 ± 5 | 132.7 ± 4.3 | - | - | - | |
| Red muscle | TBARS | 0.99 ± 0.35 | 2.37 ± 0.72 | 0.81 ± 0.09 | 3.26 ± 0.82 | ** | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Moya, A.; Perelló-Amorós, M.; Vélez, E.J.; Viñuales, J.; García-Pérez, I.; Blasco, J.; Gutiérrez, J.; Fernández-Borràs, J. Interaction between the Effects of Sustained Swimming Activity and Dietary Macronutrient Proportions on the Redox Status of Gilthead Sea Bream Juveniles (Sparus aurata L.). Antioxidants 2022, 11, 319. https://doi.org/10.3390/antiox11020319
Sánchez-Moya A, Perelló-Amorós M, Vélez EJ, Viñuales J, García-Pérez I, Blasco J, Gutiérrez J, Fernández-Borràs J. Interaction between the Effects of Sustained Swimming Activity and Dietary Macronutrient Proportions on the Redox Status of Gilthead Sea Bream Juveniles (Sparus aurata L.). Antioxidants. 2022; 11(2):319. https://doi.org/10.3390/antiox11020319
Chicago/Turabian StyleSánchez-Moya, Albert, Miquel Perelló-Amorós, Emilio J. Vélez, Julia Viñuales, Isabel García-Pérez, Josefina Blasco, Joaquim Gutiérrez, and Jaume Fernández-Borràs. 2022. "Interaction between the Effects of Sustained Swimming Activity and Dietary Macronutrient Proportions on the Redox Status of Gilthead Sea Bream Juveniles (Sparus aurata L.)" Antioxidants 11, no. 2: 319. https://doi.org/10.3390/antiox11020319
APA StyleSánchez-Moya, A., Perelló-Amorós, M., Vélez, E. J., Viñuales, J., García-Pérez, I., Blasco, J., Gutiérrez, J., & Fernández-Borràs, J. (2022). Interaction between the Effects of Sustained Swimming Activity and Dietary Macronutrient Proportions on the Redox Status of Gilthead Sea Bream Juveniles (Sparus aurata L.). Antioxidants, 11(2), 319. https://doi.org/10.3390/antiox11020319