
Nitric Oxide Resistance in Leishmania (Viannia) braziliensis Involves Regulation of Glucose Consumption, Glutathione Metabolism and Abundance of Pentose Phosphate Pathway Enzymes

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Parasite Culture and Growth Curve
2.3. Inhibitory Activity of NO Donor on Promastigotes of L. braziliensis
2.4. Effect of Pro-Oxidant and Antioxidant Molecules on Intracellular Amastigotes
2.5. ROS and RNS Release by Infected Macrophages
2.6. Protein Extraction and Sample Preparation
2.7. LC-MS/MS Analysis
2.8. Data Analysis
2.9. Analysis of 2-NBDG Uptake
2.10. Analysis of Oxygen Uptake
2.11. Analysis of Enzymatic Activities
2.12. Statistical Analysis
3. Results
3.1. The NO-Resistant 2853 L. braziliensis Strain Is More Infective to Macrophages In Vitro
3.2. NO-Susceptible Parasites Trigger a More Intense ROS-Dependent Response in Peritoneal Macrophages
3.3. Difference in Protein Abundance Is Observed between NO-Resistant and NO-Susceptible L. braziliensis Strains and it Is Significantly Modulated in Response to NO
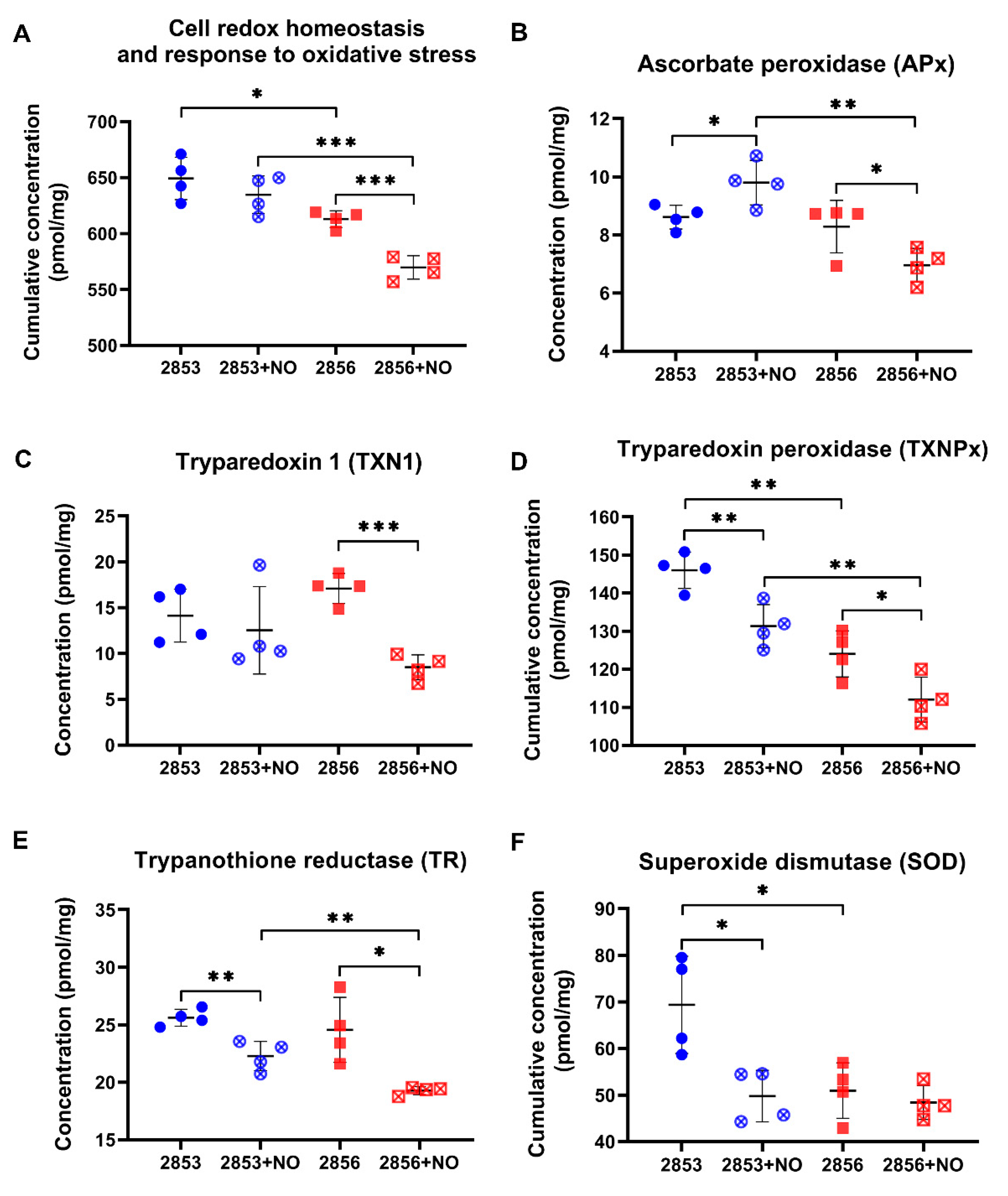
3.4. The Nitrosative Challenge Negatively Modulated Antioxidant Proteins of NO-Susceptible L. braziliensis Strain
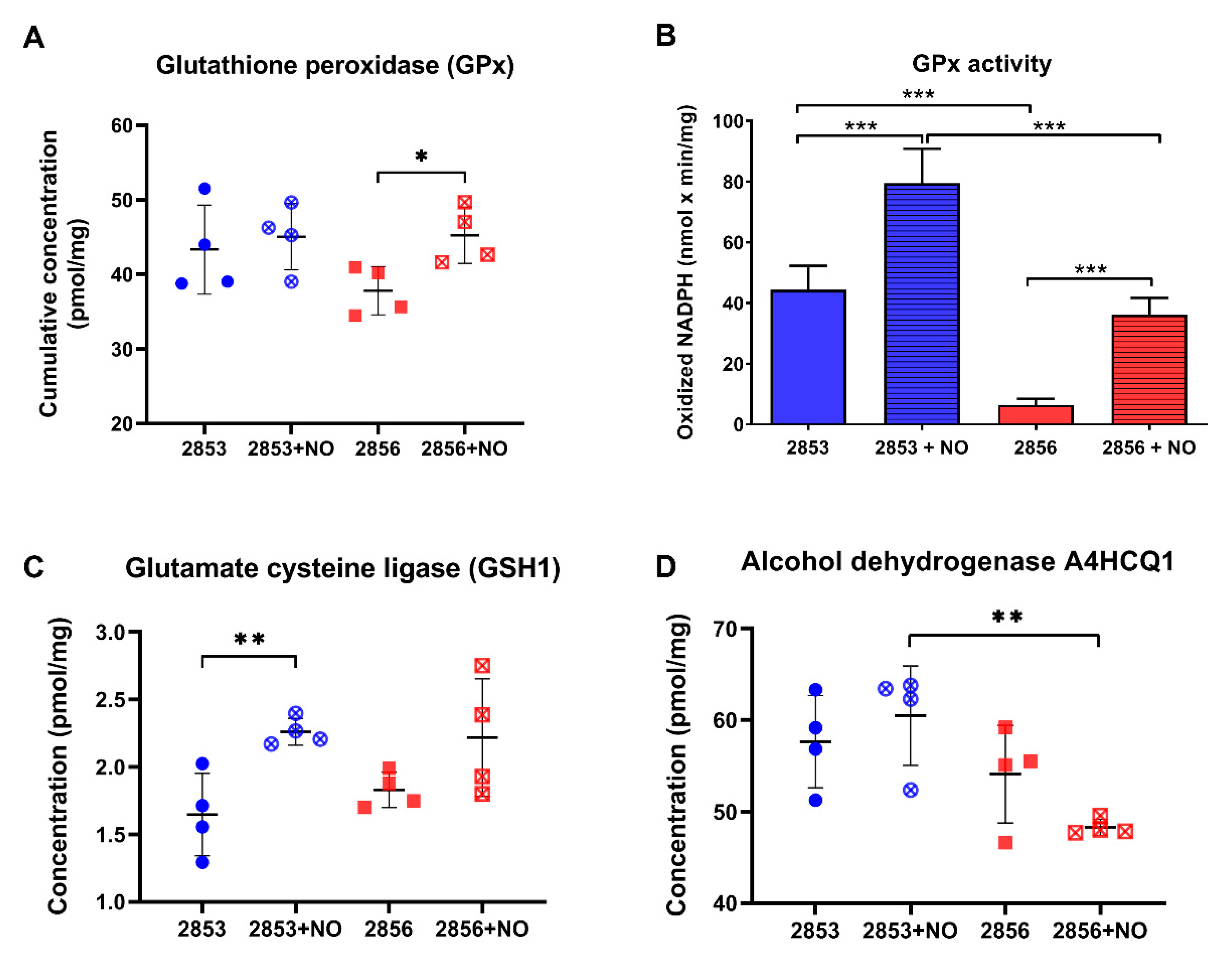
3.5. NO-Resistant L. braziliensis Strain Challenged with NO Increases the Abundance of Enzymes Involved in the Glutathione Pathway
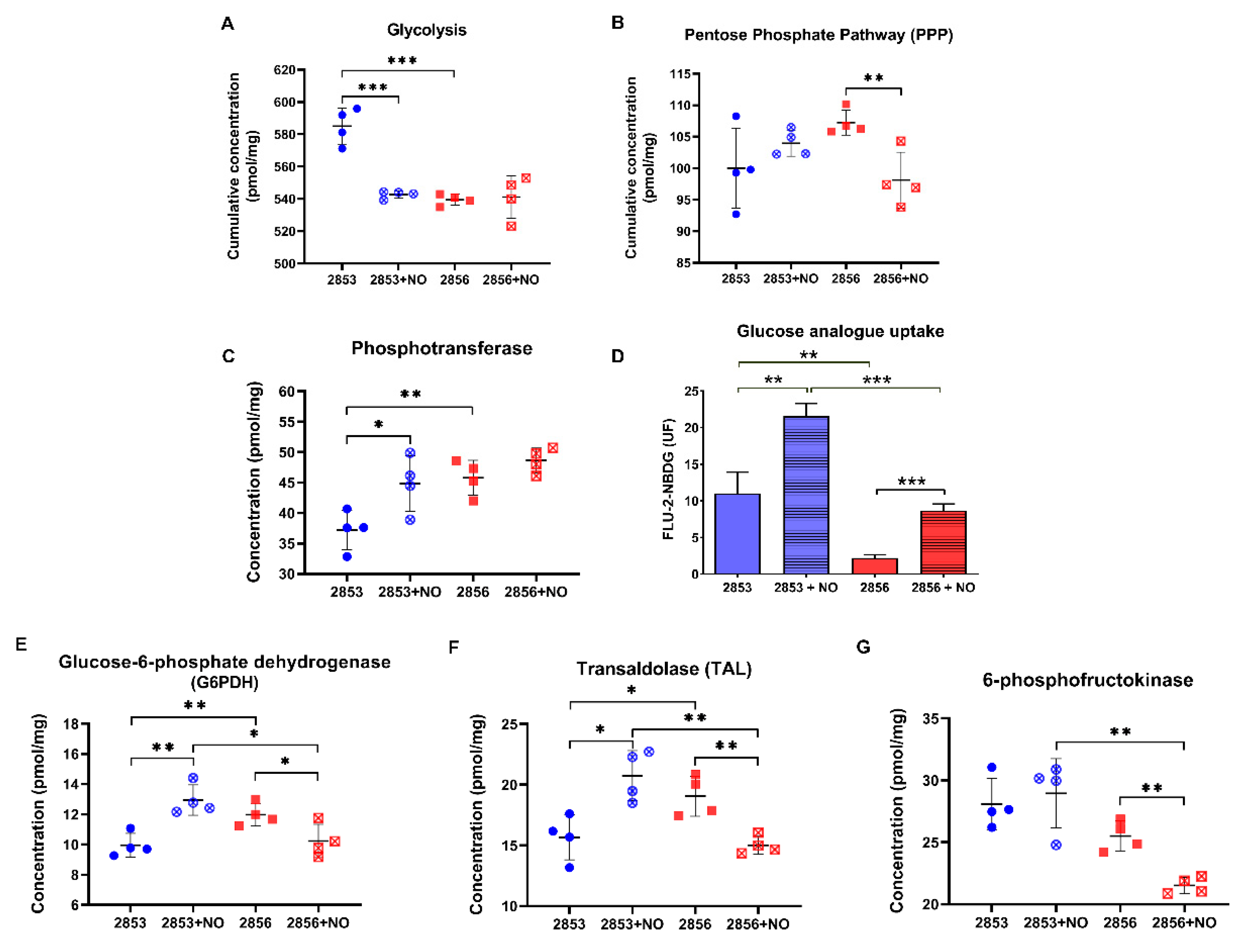
3.6. Uptake of Glucose Analog and G6PDH Protein Levels Increase in NO-Resistant L. braziliensis Strain after the Nitrosative Challenge
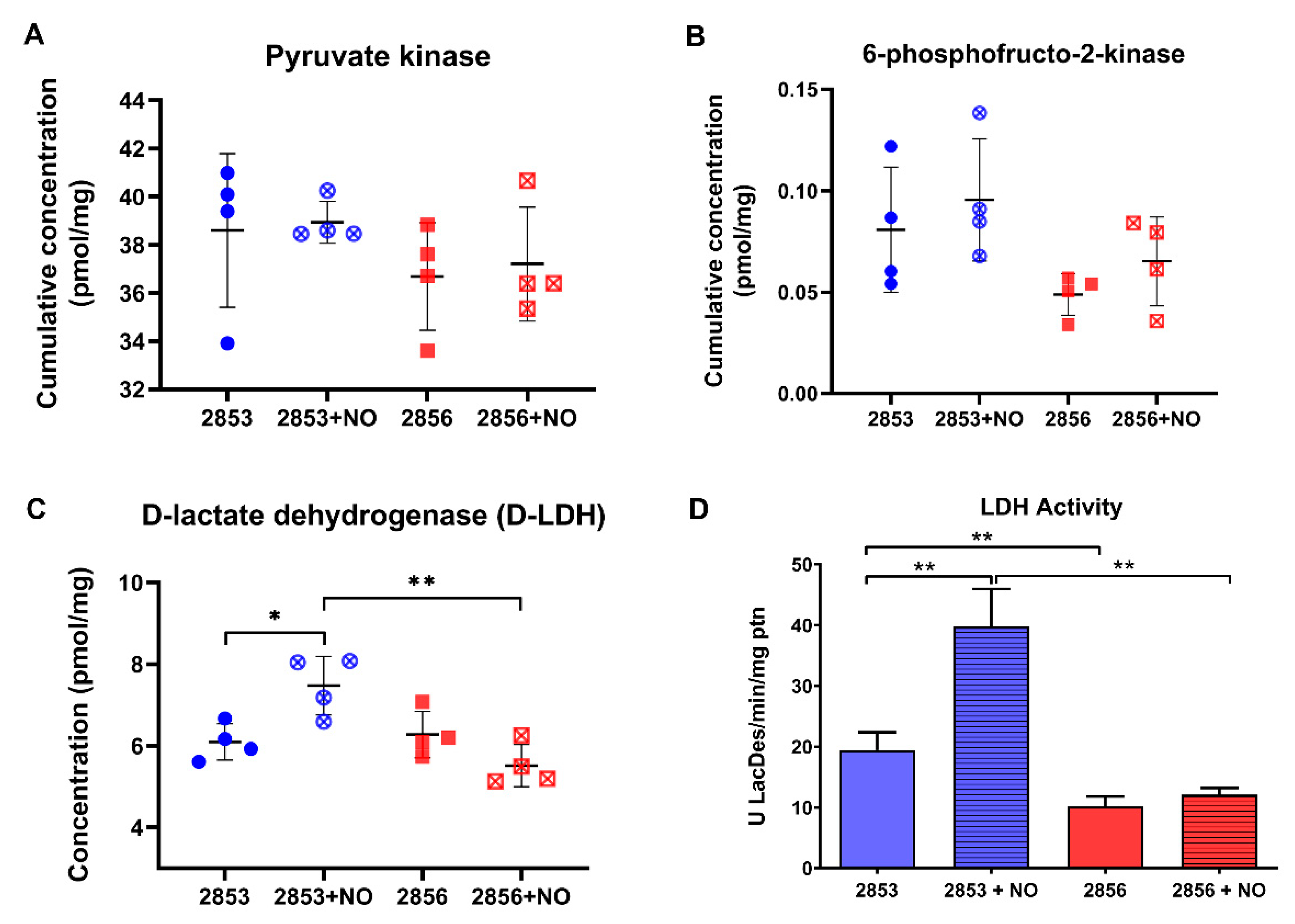
3.7. Nitrosative Challenge Increases D-Lactate Dehydrogenase Abundance in NO-Resistant L. braziliensis Strain
3.8. Nitrosative Challenge Impairs Mitochondrial O2 Consumption by L. braziliensis Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; de Boer, M. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Jirmanus, L.; Glesby, M.J.; Guimaraes, L.H.; Lago, E.; Rosa, M.E.; Machado, P.R.; Carvalho, E.M. Epidemiological and clinical changes in American tegumentary leishmaniasis in an area of Leishmania (viannia) braziliensis transmission over a 20-year period. Am. J. Trop. Med. Hyg. 2012, 86, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.D.; de Queiroz Sousa, A. Clinical spectrum of leishmaniasis. Clin. Infect. Dis. 1996, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Novais, F.O.; Scott, P. CD8+ T cells in cutaneous leishmaniasis: The good, the bad, and the ugly. Semin. Immunopathol. 2015, 37, 251–259. [Google Scholar] [CrossRef]
- Christensen, S.M.; Dillon, L.A.L.; Carvalho, L.P.; Passos, S.; Novais, F.O.; Hughitt, V.K.; Beiting, D.P.; Carvalho, E.M.; Scott, P.; El-Sayed, N.M.; et al. Meta-Transcriptome profiling of the human-Leishmania braziliensis cutaneous lesion. PLoS Negl. Trop. Dis. 2016, 10, e0004992. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.A.; Contreras, I.; Hallé, M.; Tremblay, M.L.; McMaster, R.W.; Olivier, M. Leishmania GP63 alters host signaling through cleavage-activated protein tyrosine phosphatases. Sci. Signal. 2009, 2, ra58. [Google Scholar] [CrossRef]
- Silverman, J.M.; Reiner, N.E. Leishmania exosomes deliver preemptive strikes to create an environment permissive for early infection. Front. Cell. Infect. Microbiol. 2011, 1, 26. [Google Scholar] [CrossRef]
- De Carvalho, R.V.H.; Lima-Junior, D.S.; da Silva, M.V.G.; Dilucca, M.; Rodrigues, T.S.; Horta, C.V.; Silva, A.L.N.; da Silva, P.F.; Frantz, F.G.; Lorenzon, L.B.; et al. Leishmania RNA virus exacerbates leishmaniasis by subverting innate immunity via TLR3-mediated NLRP3 inflammasome inhibition. Nat. Commun. 2019, 10, 5273. [Google Scholar] [CrossRef]
- Rodríguez-Vega, A.; Losada-Barragán, M.; Berbert, L.R.; Mesquita-Rodrigues, C.; Bombaça, A.C.S.; Menna-Barreto, R.; Aquino, P.; Carvalho, P.C.; Padrón, G.; de Jesus, J.B.; et al. Quantitative analysis of proteins secreted by Leishmania (Viannia) braziliensis strains associated to distinct clinical manifestations of American Tegumentary Leishmaniasis. J. Proteomics 2021, 232, 104077. [Google Scholar] [CrossRef]
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host-pathogen interface. Nat. Rev. Microbiol. 2011, 9, 604–615. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Sher, A. Evasion of innate immunity by parasitic protozoa. Nat. Immunol. 2002, 3, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y.; Wei, X.Q.; Proudfoot, L. Cytokines and nitric oxide as effector molecules against parasitic infections. Philos. Trans. R. Soc. B Biol. Sci. 1997, 352, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- Genestra, M.; de Souza, W.J.S.; Cysne-Finkelstein, L.; Leon, L.L. Comparative analysis of the nitric oxide production by Leishmania sp. Med. Microbiol. Immunol. 2003, 192, 217–223. [Google Scholar] [CrossRef]
- Novais, F.O.; Carvalho, L.P.; Graff, J.W.; Beiting, D.P.; Ruthel, G.; Roos, D.S.; Betts, M.R.; Goldschmidt, M.H.; Wilson, M.E.; de Oliveira, C.I.; et al. Cytotoxic T cells mediate pathology and metastasis in cutaneous leishmaniasis. PLoS Pathog. 2013, 9, e1003504. [Google Scholar] [CrossRef]
- Holzmuller, P.; Sereno, D.; Lemesre, J.L. Lower nitric oxide susceptibility of trivalent antimony-resistant amastigotes of Leishmania infantum. Antimicrob. Agents Chemother. 2005, 49, 4406–4409. [Google Scholar] [CrossRef]
- Giudice, A.; Camada, I.; Leopoldo, P.T.G.; Pereira, J.M.B.; Riley, L.W.; Wilson, M.E.; Ho, J.L.; de Jesus, A.R.; Carvalho, E.M.; Almeida, R.P. Resistance of Leishmania (Leishmania) amazonensis and Leishmania (Viannia) braziliensis to nitric oxide correlates with disease severity in Tegumentary Leishmaniasis. BMC Infect. Dis. 2007, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Santos, P.L.; Costa, R.V.; Braz, J.M.; Santos, L.F.V.C.; Batista, A.C.; Vasconcelos, C.R.O.; Rangel, M.R.; de Jesus, A.R.; de Moura, T.R.; Leopoldo, P.T.G.; et al. Leishmania chagasi naturally resistant to nitric oxide isolated from humans and dogs with visceral leishmaniasis in Brazil. Nitric Oxide Biol. Chem. 2012, 27, 67–71. [Google Scholar] [CrossRef][Green Version]
- Ávila, L.R.; Gomes, C.M.; Oliveira, P.G.; Gomes, R.S.; Vinaud, M.C.; Dorta, M.L.; Uliana, S.R.B.; Ribeiro-Dias, F.; Oliveira, M.A.P. Promastigote parasites cultured from the lesions of patients with mucosal leishmaniasis are more resistant to oxidative stress than promastigotes from a cutaneous lesion. Free Radic. Biol. Med. 2018, 129, 35–45. [Google Scholar] [CrossRef]
- Souza, A.S.; Giudice, A.; Pereira, J.M.B.; Guimarães, L.H.; de Jesus, A.R.; de Moura, T.R.; Wilson, M.E.; Carvalho, E.M.; Almeida, R.P. Resistance of Leishmania (Viannia) braziliensis to nitric oxide: Correlation with antimony therapy and TNF-α production. BMC Infect. Dis. 2010, 10, 209. [Google Scholar] [CrossRef]
- Costa, D.L.; Carregaro, V.; Lima-Júnior, D.S.; Silva, N.M.; Milanezi, C.M.; Cardoso, C.R.; Giudice, Â.; de Jesus, A.R.; Carvalho, E.M.; Almeida, R.P.; et al. BALB/c mice infected with antimony treatment refractory isolate of Leishmania braziliensis present severe lesions due to IL-4 production. PLoS Negl. Trop. Dis. 2011, 5, e965. [Google Scholar] [CrossRef] [PubMed]
- De Moura, T.R.; Santos, M.L.B.; Braz, J.M.; Santos, L.F.V.C.; Aragão, M.T.; de Oliveira, F.A.; Santos, P.L.; da Silva, Â.M.; de Jesus, A.R.; de Almeida, R.P. Cross-Resistance of Leishmania infantum isolates to nitric oxide from patients refractory to antimony treatment, and greater tolerance to antileishmanial responses by macrophages. Parasitol. Res. 2016, 115, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Alcolea, P.J.; Tuñón, G.I.L.; Alonso, A.; García-Tabares, F.; Ciordia, S.; Mena, M.C.; Campos, R.N.S.; Almeida, R.P.; Larraga, V. Differential protein abundance in promastigotes of nitric oxide-sensitive and resistant Leishmania chagasi strains. Proteome Clin. Appl. 2016, 10, 1132–1146. [Google Scholar] [CrossRef] [PubMed]
- Dias-Lopes, G.; Wiśniewski, J.R.; de Souza, N.P.; Vidal, V.E.; Padrón, G.; Britto, C.; Cuervo, P.; de Jesus, J.B. In-Depth quantitative proteomic analysis of trophozoites and pseudocysts of Trichomonas vaginalis. J. Proteome Res. 2018, 17, 3704–3718. [Google Scholar] [CrossRef]
- Pinho, N.; Wiśniewski, J.R.; Dias-Lopes, G.; Saboia-Vahia, L.; Bombaça, A.C.S.; Mesquita-Rodrigues, C.; Menna-Barreto, R.; Cupolillo, E.; de Jesus, J.B.; Padrón, G.; et al. In-Depth quantitative proteomics uncovers specie-specific metabolic programs in Leishmania (Viannia) species. PLoS Negl. Trop. Dis. 2020, 14, e0008509. [Google Scholar] [CrossRef]
- Gonçalves, R.L.S.; Barreto, R.F.S.M.; Polycarpo, C.R.; Gadelha, F.R.; Castro, S.L.; Oliveira, M.F. A comparative assessment of mitochondrial function in epimastigotes and bloodstream trypomastigotes of Trypanosoma cruzi. J. Bioenerg. Biomembr. 2011, 43, 651–661. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Wiśniewski, J.R. Filter-Aided Sample Preparation: The versatile and efficient method for proteomic analysis. Methods Enzymol. 2017, 585, 15–27. [Google Scholar] [CrossRef]
- Wiśniewski, J.R. Label-Free and standard-free absolute quantitative proteomics using the “total protein” and “proteomic ruler” approaches. Methods Enzymol. 2017, 585, 49–60. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Menna-Barreto, R.F.S.; Goncalves, R.L.S.; Costa, E.M.; Silva, R.S.F.; Pinto, A.V.; Oliveira, M.F.; de Castro, S.L. The effects on Trypanosoma cruzi of novel synthetic naphthoquinones are mediated by mitochondrial dysfunction. Free Radic. Biol. Med. 2009, 47, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Aebischer, T. Leishmania spp. proteome data sets: A comprehensive resource for vaccine development to target visceral leishmaniasis. Front. Immunol. 2014, 5, 260. [Google Scholar] [CrossRef] [PubMed]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New roles in redox signalling for an old antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [PubMed]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione is a crucial guardian of protein integrity in the brain upon nitric oxide imbalance. Commun. Integr. Biol. 2011, 4, 477–479. [Google Scholar] [CrossRef]
- Basu, S.; Keszler, A.; Azarova, N.A.; Nwanze, N.; Perlegas, A.; Shiva, S.; Broniowska, K.A.; Hogg, N.; Kim-Shapiro, D.B. A novel role for cytochrome c: Efficient catalysis of S-nitrosothiol formation. Free Radic. Biol. Med. 2010, 48, 255–263. [Google Scholar] [CrossRef]
- Broniowska, K.A.; Diers, A.R.; Hogg, N. S-Nitrosoglutathione. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3173–3181. [Google Scholar] [CrossRef]
- Baldelli, S.; Ciccarone, F.; Limongi, D.; Checconi, P.; Palamara, A.T.; Ciriolo, M.R. Glutathione and nitric oxide: Key team players in use and disuse of skeletal muscle. Nutrients 2019, 11, 2318. [Google Scholar] [CrossRef]
- Godoy, L.; Gonzàlez-Duarte, R.; Albalat, R. S-Nitrosogluthathione reductase activity of amphioxus ADH3: Insights into the nitric oxide metabolism. Int. J. Biol. Sci. 2006, 2, 117–124. [Google Scholar] [CrossRef]
- Barnett, S.D.; Buxton, I.L.O. The role of S-nitrosoglutathione reductase (GSNOR) in human disease and therapy. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 340–354. [Google Scholar] [CrossRef]
- Brown, G.C. Nitric oxide and mitochondrial respiration. Biochim. Biophys. Acta Bioenerg. 1999, 1411, 351–369. [Google Scholar] [CrossRef]
- McConville, M.J.; Saunders, E.C.; Kloehn, J.; Dagley, M.J. Leishmania carbon metabolism in the macrophage phagolysosome—Feast or Famine? F1000Research 2015, 4, 938. [Google Scholar] [CrossRef]
- Van Assche, T.; Deschacht, M.; da Luz, R.A.I.; Maes, L.; Cos, P. Leishmania-macrophage interactions: Insights into the redox biology. Free Radic. Biol. Med. 2011, 51, 337–351. [Google Scholar] [CrossRef]
- Volpedo, G.; Pacheco-Fernandez, T.; Holcomb, E.A.; Cipriano, N.; Cox, B.; Satoskar, A.R. Mechanisms of immunopathogenesis in cutaneous leishmaniasis and post kala-azar dermal leishmaniasis (PKDL). Front. Cell. Infect. Microbiol. 2021, 11, 685296. [Google Scholar] [CrossRef]
- Bogdan, C. Macrophages as host, effector and immunoregulatory cells in leishmaniasis: Impact of tissue micro-environment and metabolism. Cytokine X 2020, 2, 100041. [Google Scholar] [CrossRef] [PubMed]
- Colotti, G.; Ilari, A. Polyamine metabolism in Leishmania: From arginine to trypanothione. Amino Acids 2011, 40, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Morris, S.M. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Novais, F.O.; Nguyen, B.T.; Beiting, D.P.; Carvalho, L.P.; Glennie, N.D.; Passos, S.; Carvalho, E.M.; Scott, P. Human classical monocytes control the intracellular stage of Leishmania braziliensis by reactive oxygen species. J. Infect. Dis. 2014, 209, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.B.; Celes, F.S.; Paiva, C.N.; de Oliveira, C.I. The paradoxical leishmanicidal effects of superoxide dismutase (SOD) mimetic tempol in Leishmania braziliensis infection in vitro. Front. Cell. Infect. Microbiol. 2019, 9, 237. [Google Scholar] [CrossRef]
- Haidaris, C.G.; Bonventre, P.F. A role for oxygen-dependent mechanisms in killing of Leishmania donovani tissue forms by activated macrophages. J. Immunol. 1982, 129, 850–855. [Google Scholar]
- Gantt, K.R.; Goldman, T.L.; McCormick, M.L.; Miller, M.A.; Jeronimo, S.M.B.; Nascimento, E.T.; Britigan, B.E.; Wilson, M.E. Oxidative responses of human and murine macrophages during phagocytosis of Leishmania chagasi. J. Immunol. 2001, 167, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Linares, E.; Giorgio, S.; Augusto, O. Inhibition of in vivo leishmanicidal mechanisms by tempol: Nitric oxide down-regulation and oxidant scavenging. Free Radic. Biol. Med. 2008, 44, 1668–1676. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, B.; Meysman, P.; Erb, I.; Bittremieux, W.; Valkenborg, D.; Baggerman, G.; Mertens, I. Four layer multi-omics reveals molecular responses to aneuploidy in Leishmania. BioRxiv 2021. [Google Scholar] [CrossRef]
- Rogers, M.B.; Hilley, J.D.; Dickens, N.J.; Wilkes, J.; Bates, P.A.; Depledge, D.P.; Harris, D.; Her, Y.; Herzyk, P.; Imamura, H.; et al. Chromosome and gene copy number variation allow major structural change between species and strains of Leishmania. Genome Res. 2011, 21, 2129–2142. [Google Scholar] [CrossRef] [PubMed]
- Sterkers, Y.; Lachaud, L.; Crobu, L.; Bastien, P.; Pagès, M. FISH analysis reveals aneuploidy and continual generation of chromosomal mosaicism in Leishmania major. Cell. Microbiol. 2011, 13, 274–283. [Google Scholar] [CrossRef]
- Dumetz, F.; Imamura, H.; Sanders, M.; Seblova, V.; Myskova, J.; Pescher, P.; Vanaerschot, M.; Meehan, C.J.; Cuypers, B.; de Muylder, G.; et al. Modulation of aneuploidy in Leishmania donovani during adaptation to different in vitro and in vivo environments and its impact on gene expression. MBio 2017, 8, e00599-17. [Google Scholar] [CrossRef] [PubMed]
- Downing, T.; Imamura, H.; Decuypere, S.; Clark, T.G.; Coombs, G.H.; Cotton, J.A.; Hilley, J.D.; de Doncker, S.; Maes, I.; Mottram, J.C.; et al. Whole genome sequencing of multiple Leishmania donovani clinical isolates provides insights into population structure and mechanisms of drug resistance. Genome Res. 2011, 21, 2143–2156. [Google Scholar] [CrossRef]
- Imamura, H.; Downing, T.; van den Broeck, F.; Sanders, M.J.; Rijal, S.; Sundar, S.; Mannaert, A.; Vanaerschot, M.; Berg, M.; de Muylder, G.; et al. Evolutionary genomics of epidemic visceral leishmaniasis in the Indian subcontinent. Elife 2016, 5, e12613. [Google Scholar] [CrossRef]
- Bussotti, G.; Gouzelou, E.; Boité, M.C.; Kherachi, I.; Harrat, Z.; Eddaikra, N.; Mottram, J.C.; Antoniou, M.; Christodoulou, V.; Bali, A.; et al. Leishmania genome dynamics during environmental adaptation reveal strain-specific differences in gene copy number variation, karyotype instability, and telomeric amplification. MBio 2018, 9, e01399-18. [Google Scholar] [CrossRef]
- Patino, L.H.; Imamura, H.; Cruz-Saavedra, L.; Pavia, P.; Muskus, C.; Méndez, C.; Dujardin, J.C.; Ramírez, J.D. Major changes in chromosomal somy, gene expression and gene dosage driven by SbIII in Leishmania braziliensis and Leishmania panamensis. Sci. Rep. 2019, 9, 9485. [Google Scholar] [CrossRef]
- Zhu, J.; Tsai, H.J.; Gordon, M.R.; Li, R. Cellular stress associated with aneuploidy. Dev. Cell. 2018, 44, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Sardar, A.H.; Kumar, S.; Kumar, A.; Purkait, B.; Das, S.; Sen, A.; Kumar, M.; Sinha, K.K.; Singh, D.; Equbal, A.; et al. Proteome changes associated with Leishmania donovani promastigote adaptation to oxidative and nitrosative stresses. J. Proteomics 2013, 81, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Laranjeira-Silva, M.F.; Maeda, F.Y.; Hauzel, J.; Andrews, N.W.; Mittra, B. Ascorbate-Dependent peroxidase (APX) from Leishmania amazonensis is a reactive oxygen species-induced essential enzyme that regulates virulence. Infect. Immun. 2019, 87, e00193-19. [Google Scholar] [CrossRef] [PubMed]
- De Moreira, D.S.; Xavier, M.V.; Murta, S.M.F. Ascorbate peroxidase overexpression protects Leishmania braziliensis against trivalent antimony effects. Mem. Inst. Oswaldo Cruz. 2018, 113, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ariyanayagam, M.R.; Fairlamb, A.H. Ovothiol and trypanothione as antioxidants in trypanosomatids. Mol. Biochem. Parasitol. 2001, 115, 189–198. [Google Scholar] [CrossRef]
- Krauth-Siegel, L.R.; Comini, M.A.; Schlecker, T. The trypanothione system. Subcell. Biochem. 2007, 44, 231–251. [Google Scholar] [CrossRef] [PubMed]
- Romão, P.R.T.; Tovar, J.; Fonseca, S.G.; Moraes, R.H.; Cruz, A.K.; Hothersall, J.S.; Noronha-Dutra, A.A.; Ferreira, S.H.; Cunha, F.Q. Glutathione and the redox control system trypanothione/trypanothione reductase are involved in the protection of Leishmania spp. against nitrosothiol-induced cytotoxicity. Braz. J. Med. Biol. Res. 2006, 39, 355–363. [Google Scholar] [CrossRef]
- Halliwell, B. Vitamin C: Poison, prophylactic or panacea? Trends Biochem. Sci. 1999, 24, 255–259. [Google Scholar] [CrossRef]
- Krauth-Siegel, R.L.; Lüdemann, H. Reduction of dehydroascorbate by trypanothione. Mol. Biochem. Parasitol. 1996, 80, 203–208. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Sardar, A.H.; Mandal, A.; Saini, S.; Abhishek, K.; Kumar, A.; Purkait, B.; Singh, R.; Das, S.; Mukhopadhyay, R.; et al. Metabolic reconfiguration of the central glucose metabolism: A crucial strategy of Leishmania donovani for its survival during oxidative stress. FASEB J. 2015, 29, 2081–2098. [Google Scholar] [CrossRef]
- Holzmuller, P.; Hide, M.; Sereno, D.; Lemesre, J.L. Leishmania infantum amastigotes resistant to nitric oxide cytotoxicity: Impact on in vitro parasite developmental cycle and metabolic enzyme activities. Infect. Genet. Evol. 2006, 6, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Darling, T.N.; Blum, J.J. D-Lactate production by Leishmania braziliensis through the glyoxalase pathway. Mol. Biochem. Parasitol. 1988, 28, 121–127. [Google Scholar] [CrossRef]
- Wyllie, S.; Fairlamb, A.H. Methylglyoxal metabolism in trypanosomes and Leishmania, Semin. Cell Dev. Biol. 2011, 22, 271–277. [Google Scholar] [CrossRef]
- Silva, M.S.; Ferreira, A.E.N.; Gomes, R.; Tomás, A.M.; Freire, A.P.; Cordeiro, C. Glyoxalase Enzymes in Trypanosomatids. In Trypanosomatid Diseases; Selzer, P.M., Jäger, T., Koch, O., Flohé, L., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2013. [Google Scholar] [CrossRef]
- Rastrojo, A.; García-Hernández, R.; Vargas, P.; Camacho, E.; Corvo, L.; Imamura, H.; Dujardin, J.C.; Castanys, S.; Aguado, B.; Gamarro, F.; et al. Genomic and transcriptomic alterations in Leishmania donovani lines experimentally resistant to antileishmanial drugs. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 246–264. [Google Scholar] [CrossRef] [PubMed]
- Saunders, E.C.; Naderer, T.; Chambers, J.; Landfear, S.M.; McConville, M.J. Leishmania mexicana can utilize amino acids as major carbon sources in macrophages but not in animal models. Mol. Microbiol. 2018, 108, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, T.; Hammami, A.; Stäger, S. Hypoxia inducible factor 1α: A critical factor for the immune response to pathogens and Leishmania. Cell. Immunol. 2016, 309, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Degrossoli, A.; Arrais-Silva, W.W.; Colhone, M.C.; Gadelha, F.R.; Joazeiro, P.P.; Giorgio, S. The influence of low oxygen on macrophage response to Leishmania infection. Scand. J. Immunol. 2011, 74, 165–175. [Google Scholar] [CrossRef]
- Subramanian, A.; Jhawar, J.; Sarkar, R.R. Dissecting Leishmania infantum energy metabolism—A systems perspective. PLoS ONE 2015, 10, e0137976. [Google Scholar] [CrossRef]
- Tomás, A.M.; Castro, H. Redox metabolism in mitochondria of trypanosomatids. Antioxid. Redox Signal. 2013, 19, 696–707. [Google Scholar] [CrossRef]
- Chen, M.; Bennedsen, M.; Zhai, L.; Kharazmi, A. Purification and enzymatic activity of an NADH-fumarate reductase and other mitochondrial activities of Leishmania parasites. Apmis 2001, 109, 801–808. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinho, N.; Bombaça, A.C.; Wiśniewski, J.R.; Dias-Lopes, G.; Saboia-Vahia, L.; Cupolillo, E.; de Jesus, J.B.; de Almeida, R.P.; Padrón, G.; Menna-Barreto, R.; et al. Nitric Oxide Resistance in Leishmania (Viannia) braziliensis Involves Regulation of Glucose Consumption, Glutathione Metabolism and Abundance of Pentose Phosphate Pathway Enzymes. Antioxidants 2022, 11, 277. https://doi.org/10.3390/antiox11020277
Pinho N, Bombaça AC, Wiśniewski JR, Dias-Lopes G, Saboia-Vahia L, Cupolillo E, de Jesus JB, de Almeida RP, Padrón G, Menna-Barreto R, et al. Nitric Oxide Resistance in Leishmania (Viannia) braziliensis Involves Regulation of Glucose Consumption, Glutathione Metabolism and Abundance of Pentose Phosphate Pathway Enzymes. Antioxidants. 2022; 11(2):277. https://doi.org/10.3390/antiox11020277
Chicago/Turabian StylePinho, Nathalia, Ana Cristina Bombaça, Jacek R. Wiśniewski, Geovane Dias-Lopes, Leonardo Saboia-Vahia, Elisa Cupolillo, José Batista de Jesus, Roque P. de Almeida, Gabriel Padrón, Rubem Menna-Barreto, and et al. 2022. "Nitric Oxide Resistance in Leishmania (Viannia) braziliensis Involves Regulation of Glucose Consumption, Glutathione Metabolism and Abundance of Pentose Phosphate Pathway Enzymes" Antioxidants 11, no. 2: 277. https://doi.org/10.3390/antiox11020277
APA StylePinho, N., Bombaça, A. C., Wiśniewski, J. R., Dias-Lopes, G., Saboia-Vahia, L., Cupolillo, E., de Jesus, J. B., de Almeida, R. P., Padrón, G., Menna-Barreto, R., & Cuervo, P. (2022). Nitric Oxide Resistance in Leishmania (Viannia) braziliensis Involves Regulation of Glucose Consumption, Glutathione Metabolism and Abundance of Pentose Phosphate Pathway Enzymes. Antioxidants, 11(2), 277. https://doi.org/10.3390/antiox11020277