
Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Literature Search Methodology
3. Bioactive Compounds of Citrus Fruits
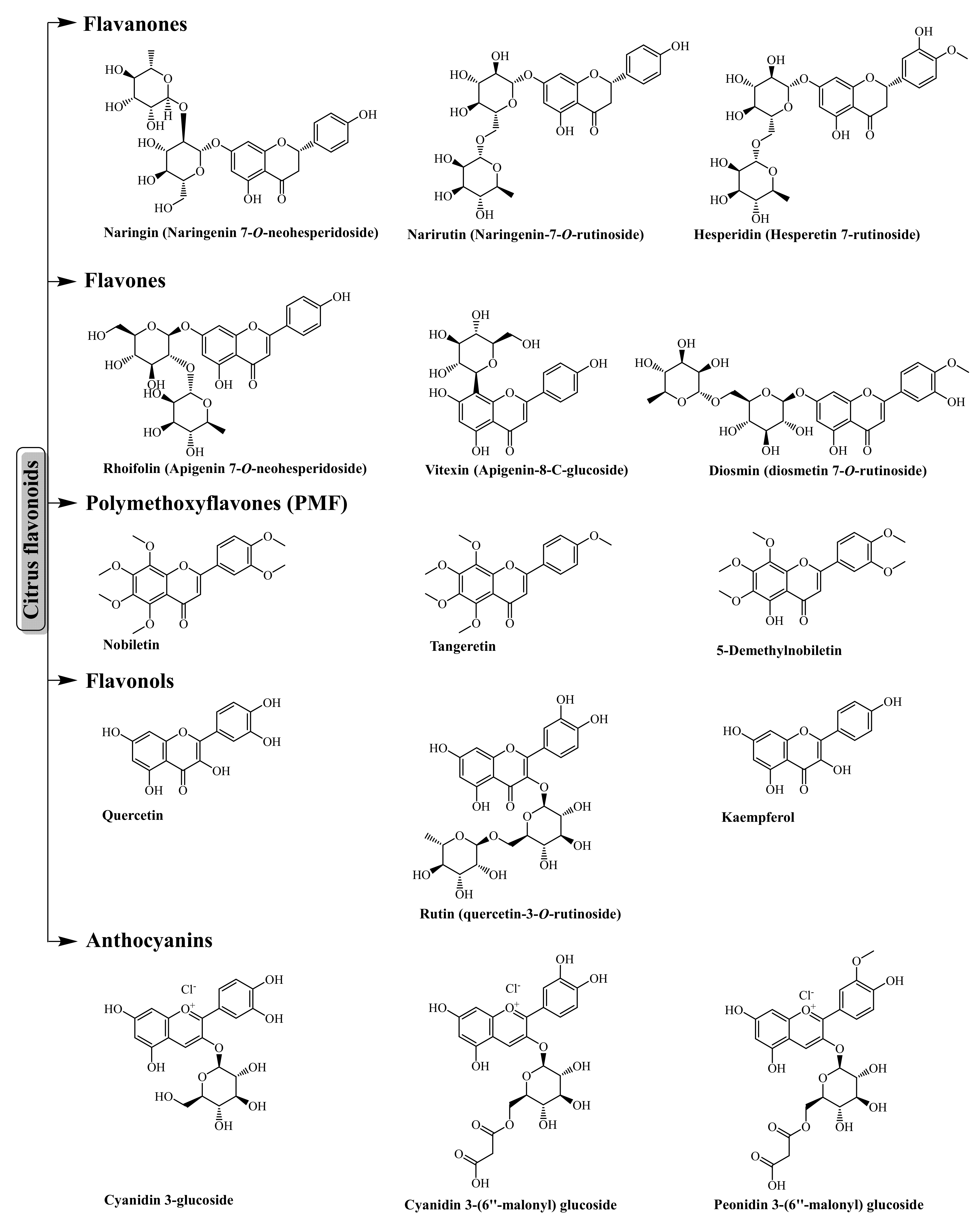
3.1. Flavonoids
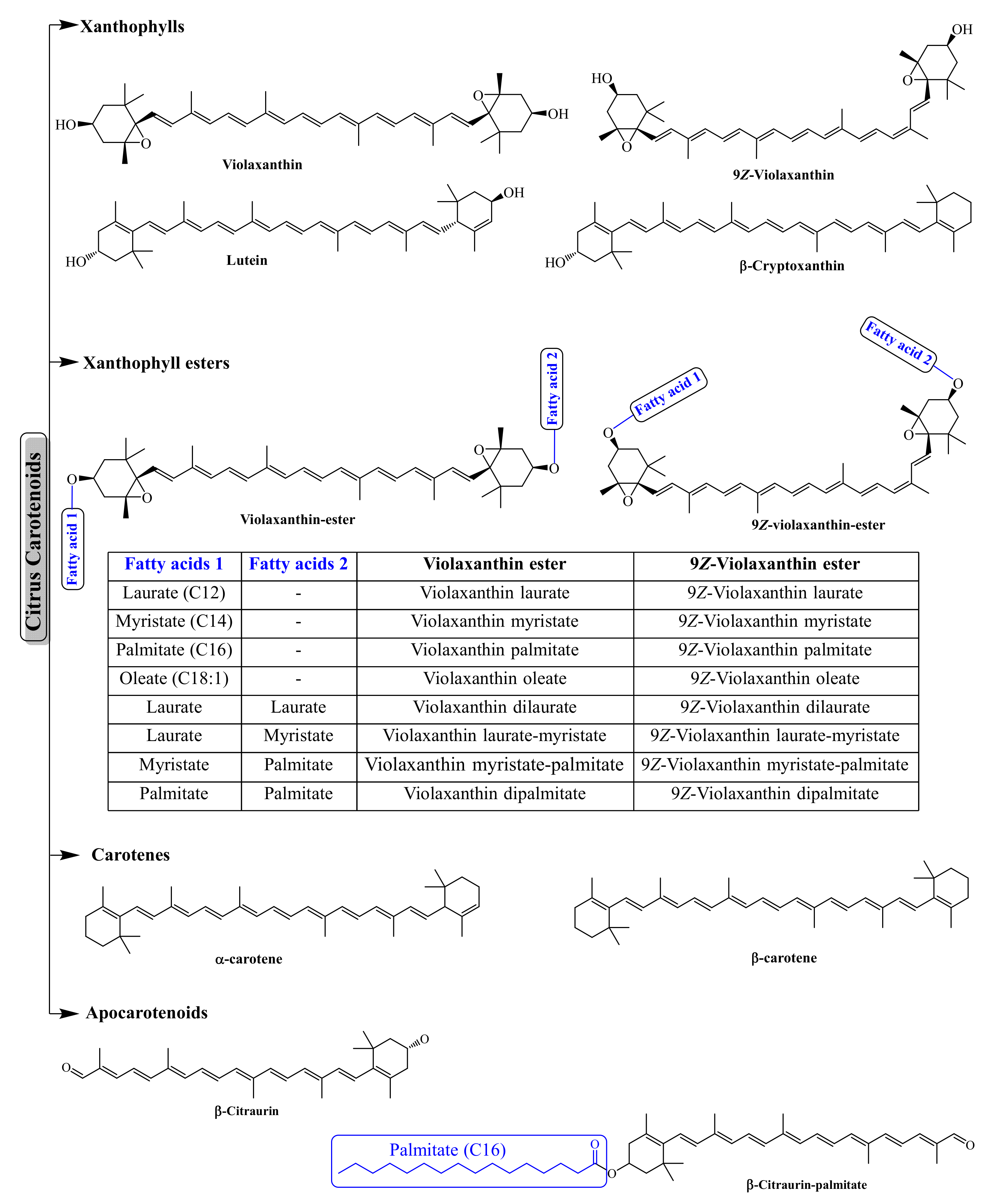
3.2. Carotenoids and Apocarotenoids
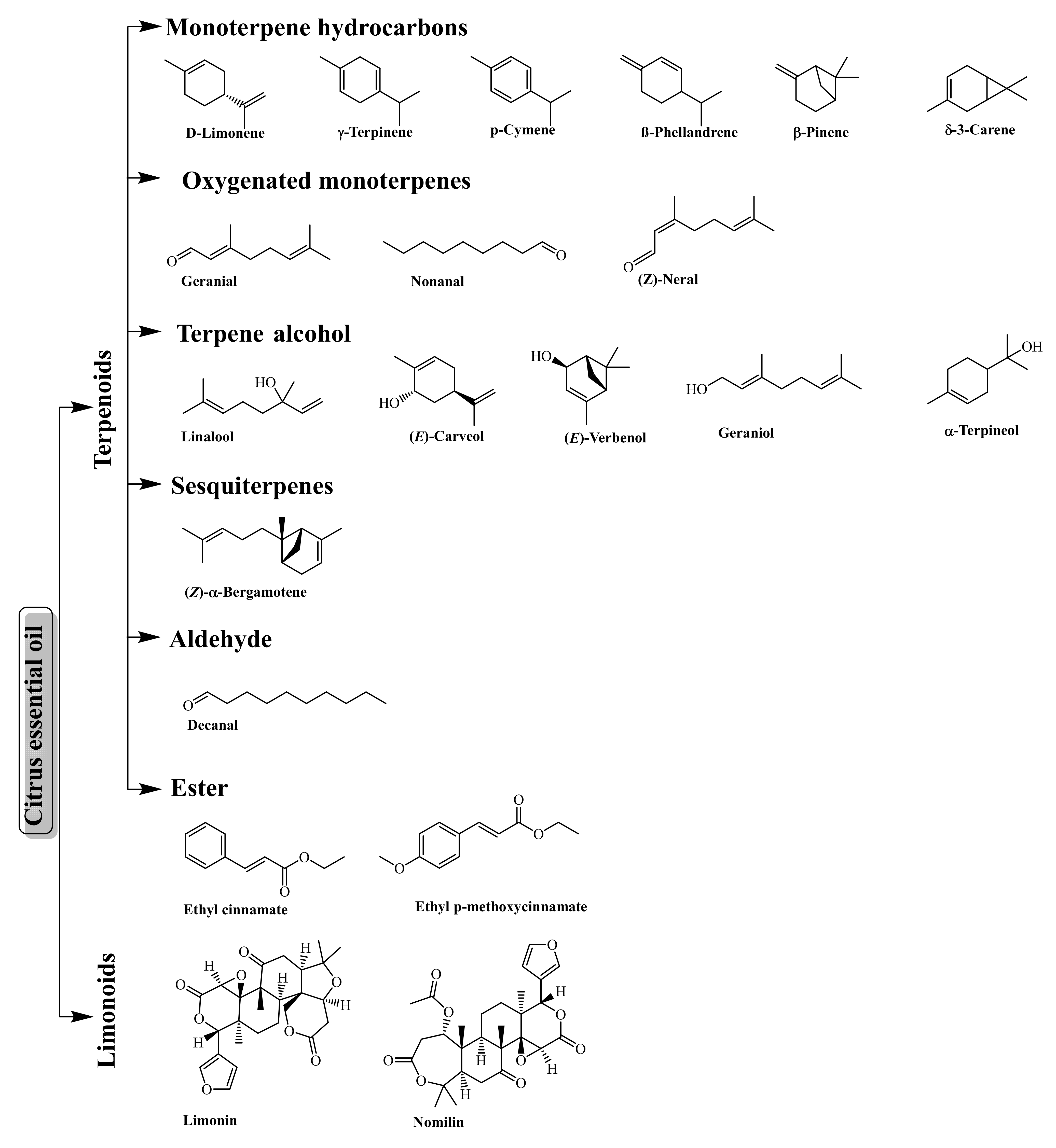
3.3. Essential Oil (Terpenes and Limonoids)
4. Bioactive Compounds of Citrus Fruit Byproducts
5. Health Benefits of Citrus Fruit Bioactive Compounds
5.1. In Vitro Studies
5.1.1. Flavonoids
5.1.2. Carotenoids
5.1.3. Essential Oil (Terpenes and Limonoids)
5.1.4. Other Bioactives

{kind=link}
{kind=link}
{kind=link}
| Compounds | Experimental System | Disease Target | Mechanism of Action | Reference |
|---|---|---|---|---|
| Flavanone-rich mandarin juice extract (0.001–1 mg/mL) | 6-hydroxydopamine (6-OHDA)-stimulated SH-SY5Y human neuroblastoma cells | Parkinson’s disease (PD) | ↓ROS and NO, restored SOD and CAT activity, ↓caspase 3 activity, ↑Bcl-2 mRNA, ↓p53 and Bax mRNA, restored mitochondrial membrane potential, ↓oxidative DNA damage, balanced α-synuclein, LRRK2, parkin, PINK1, and DJ-1 mRNA levels | [88] |
| Flavanones (10 µM) | Caco-2 cells stimulated with IL-1 | Bowel diseases | ↓IL-6, IL-8, and NO release | [83] |
| Hesperetin and gardenin A (5–10 µM) | PMA/ ionomycin-induced EL-4 murine T-lymphoma cell cells | Asthma | ↓ROS and IL-5 production, ↓NFAT activity and IL-5 secretion, ↑HO-1 through ↑Nrf2, PPARγ, PI3K/AKT, or ERK/JNK signaling | [84] |
| Limonene (1–100 mg/mL) | Aβ1–42 triggered toxicity in primary cortical neurons | Alzheimer’s disease (AD) | ↓AchE, ROS production, voltage-gated K+ channel KV3.4 hyperfunction, and phosphorylated ERK | [87] |
| Limonin, nomilin, and limonexic acid (20–60 µM) | Human pancreatic Panc-28 cells | Cancer (pancreatic) | ↓Cell proliferation (IC50 values < 50 μm after 72 h), ↑cleavage of caspase-3, mitochondrial membrane potential, ↑Bax/Bcl2 expression, and p21, ↓COX-2, NF-κβ, and IL-6 | [90] |
| Limonoids (Fortunellon and nomilin; 30 µM) | HeLa cells | Cancer (Cervical) | ↑Adriamycin-dependent cell death | [89] |
| Naringenin (62.5–2000 µM) | Human A549 lung epithelial cells and primary human monocyte-derived dendritic cells | Zika virus infection | ↓Replication or assembly of viral particles | [100] |
| Naringin- and hesperidin-rich junos peel extract (0.5 mg/mL) | Human lung basal epithelial NCI-H460 cells exposed to H2O2 | Oxidative stress-induced diseases | ↓p53, cytochrome c, and Bax proteins | [80] |
| Pectin oligosaccharides (5 mg/mL) | LPS-stimulated human macrophages | Atherosclerosis | ↑Immune responses, ↓TNF-α, IL-6, IL-10, and NF-κβ mRNA, ↑cholesterol efflux via LXRα and ABCA1, and ABCG1 pathway, ↓cholesterol synthesis via ↓HMGCR | [85] |
| Phase-II flavanone metabolites (2–100 µM) | Pancreatic β-cell MIN6 cells exposed to cholesterol | Oxidative stress-induced diseases | ↓Oxidative biomarkers (superoxide anion, H2O2, and MDA), ↓SOD and GPx, ↑insulin secretion, ↓apoptosis | [81] |
| PMF nobiletin (10–50 µM) | LPS-stimulated RAW264.7 cells | Inflammatory diseases | ↓Release of NO, ↓expression of iNOS and COX-2, ↑autophagy, activation of the IL-6/STAT3/FOXO3a signal pathway | [32] |
| β-cryptoxanthin from mandarin oranges; IC50—4.5µM (24 h treatment) | HeLa cells | Cancer (cervical) | ↓Bcl-2 mRNA, ↑Bax, caspase-3, -7, and -9 mRNA, nuclear condensation and disruption of the integrity of the mitochondrial membrane, activation of caspase-3 proteins, nuclei DNA damage, and apoptosis | [101] |
5.2. In Vivo Studies
5.2.1. Flavonoids
5.2.2. Carotenoids
5.2.3. Essential Oil (Terpenes and Limonoids)
| Bioactive and Doses | Experimental System | Disease Target | Mechanism of Action | Reference |
|---|---|---|---|---|
| Auraptene (50 mg/kg), naringin (50 mg/kg) for 14 days | STZ-induced hyperglycemia in C57BL/6 mice | Alzheimer’s disease (AD) | ↓Tau hyperphosphorylation, astroglial activation, and ↑neurogenesis in the hippocampus | [108] |
| Citrus concentrate containing 0.086 mg β-cryptoxanthin, 5.69 mg hesperidin, and 7.5 mg pectin for 8 weeks | Wistar male rats fed with high-fructose diet | Metabolic syndrome (type 2 diabetes) | ↓Plasma glucose, glycemia, insulinemia, and LDL-C, VLDL-C, and TG levels, ↑liver retinyl palmitate, and plasma β-cryptoxanthin | [103] |
| Coumarin (auraptene, 7.5–30 mg/kg for three days a week for total of 8 weeks) | TAA-induced hepatic fibrosis in male C57BL/6 mice | Hepatic fibrosis (cirrhosis and liver cancer) | ↓Bile acids in liver by increasing their efflux, ↓activation of HSCs by suppressing the expression of TGF-β1 and -SMAα and ↓expression of NF-κB, TNF-α, and IL-1β | [116] |
| Coumarin auraptene (5–20 mg/kg) | 17α-Ethinylestradiol (synthetic estrogen) induced cholestasis in male C57BL/6 mice | Estrogen-induced cholestasis | ↑Bile acid transporters (Bsep and Mrp2) mRNA and proteins, ↑Shp and Fgf15, FXR, ↑bile acid metabolism, ↑SULT2A1, ↓Cyp7a1 and Cyp8b1 mRNA, ↓hepatic inflammation (↓TNF-α, IL-1β, and IL-6) | [117] |
| Essential oil (0.75% of the diet for 42 weeks) | Male SD rats fed with HFD | Metabolic syndrome (hyperlipidemia) | ↓TC, LDL-C, hepatic TC, TG, and hepatic lipid droplet accumulation, ↓liver FFAs, TG, and CE | [102] |
| Essential oil (limonene; daily inhalation for 1.5 and 24 h, for five days) | CUMS male Kunming mice mouse model | Depression | ↑Curiosity, body weight, sucrose preference, 5-HT, DA, NE, BNDF, TrkB, GR, ↓CRF, CORT | [106] |
| Flavanone aglycones rich ougan (cv. Suavissima) juice (20 mL/kg for 10 weeks) | HFD-fed C57BL/6J mice | Metabolic syndrome (obesity) | ↓Weight gain, ↓fat accumulation, ↓liver steatosis, ↑glucose homeostasis and insulin sensitivity, ↑BAT activity, and ↑WAT browning, ↑diversity of gut microbiota | [109] |
| Flavanones (eriocitrin and eriodictyol), 25 and 50 mg/kg | BALB/c mice with LPS-induced periodontal disease | Periodontitis | ↓Gingival IL-1β and TNF-α, ↑IL-10, ↓MPO and EPO activity, SOD, ↑CAT and GPx activities, ↓MDA | [118] |
| Flavanones (naringin, naringenin, hesperidin, and hesperetin; 100 mg/kg/day for 16 weeks) | ApoE−/− mice | Atherosclerosis | ↑Bile acid synthesis (naringin), ↑cholesterol reverse transport (hesperidin), ↓cholesterol synthesis (naringenin and hesperetin) | [33] |
| Flavonoid-rich bitter/sour orange fruit peel extract (125–500 mg/kg for 3 days) | TNBS-induced IBD in male Sprague/Dawley (SD) rats | IBD | ↓Weight loss and diarrhea, colitis inflammatory cell infiltration, and proinflammatory cytokines (TNF-α, iNOS, COX-2), ↓serum and colon NO and MPO activity | [119] |
| Hesperetin (40 mg/kg for 45 days) | STZ-induced diabetes in male albino Wistar strain rats | Diabetes | ↓Plasma glucose, ↑plasma insulin and glycogen, ↑antioxidant system (↑SOD, CAT, GPx), ↑insulin secretion by renovating pancreatic β-cells, ↓dyslipidemia (hepatic cholesterol, FFAs, TG, and PLs), ↓serum levels of ALT, AST, and ALP, ↓renal damage (serum urea, creatinine, and uric acid) | [104] |
| Hesperetin (50 mg/kg daily for five weeks) | LPS-induced neuroinflammation C57BL/6 N mice | Alzheimer’s disease (AD) and Parkinson’s disease (PD) | ↓Inflammatory mediators (phosphorylated-NF-κβ, TNF-α, and IL-1), ROS/lipid peroxidation, ↑antioxidant protein (Nrf2 and HO-1), ↓phosphorylated-JNK, Bax, and caspase-3 protein, ↑Bcl-2, ↑synaptic integrity, cognition, and memory processes, ↑ phosphorylated-CREB, PSD-95, and Syntaxin | [107] |
| Hesperetin (50 mg/kg/day for 46 days) | STZ-induced diabetes in male Wistar rats | Diabetes-associated testicular injury | ↓Body weight loss, ↓ serum glucose, ↓MDA, ROS, protein carbonyl, DNA fragmentation, and caspase 3 activity, ↑testicular antioxidant system (↑GSH, MMP, FRAP, SOD, CAT, GPx) | [120] |
| Hesperidin (100 mg/kg for eight weeks) | Male SD rats fed an obesogenic cafeteria diet | Metabolic syndrome (obesity) | ↓TC, LDL-C, FFAs, MCP-1 | [121] |
| Limonene-rich essential oil (0.0765 mL/kg for 7 days) | SD rats with estradiol benzoate and oxytocin-induced uterine contraction | Primary dysmenorrhea (PD) | ↑Antioxidant status markers (SOD,T-AOC, CAT, and GSH), ↓MDA and iNOS, and PGF2α/PGE2 | [105] |
| Naringin- and hesperidin-rich C. junos peel extract (200 mg/kg/day, 10 days | Acrolein-induced pulmonary apoptosis in male C57BL/6J mice | Pulmonary edema and COPD | ↓Cleaved caspase 3, cleaved PARP, Bax and PUMA, p53, Prx-SO3 | [80] |
| Pectin oligosaccharides (0.15–0.9 g/kg for 30 days) | Male C57BL/6 mice fed with HFD | Metabolic syndrome | ↓Serum TC, LDL-C, ↓Firmicutes ↑Bacteroidetes ↑SCFAs (acetate, propionate, and butyrate) | [122] |
| PMF (nobilitin, tangeritin)-rich extract (30–120 mg/kg) | C57BL/6J male mice fed with HFD | Metabolic syndrome | ↓Firmicutes-to-Bacteroidetes ratio, ↓serum BCAA, ↓mTORC1 and P70S6K activation, ↓SREBPs | [35] |
| PMF (nobilitin, tangeritin, and 5-OH nobiletin)-rich aged chenpi peel extract (0.25 and 0.5% of diet weight for 11 weeks) | Male C57BL/6J mice fed with HFD | Metabolic syndrome (obesity) | ↑Fecal SCFAs (acetic acid and propionic acid), ↑healthy gut microbiota | [34] |
| PMF 5-Demethylnobiletin (12 mg per kg) | Azoxymethane/DSS-driven colorectal carcinogenesis in male CD-1 mice | Cancer (colorectal) | ↓Cell proliferation, ↑apoptosis, and ↓mRNA and protein levels of IL-1β, IL-6, and TNF-α in the colon | [36] |
| PMF- and HOPMF-rich extract (0.5% of HFD for 16 weeks) | Male C57BL/6 mice fed with HFD | Metabolic syndrome (obesity) | ↓Adipocyte size, adipose tissue weight, and alleviated the total body weight, levels of lipid droplets, and perilipin 1 protein and SREBP-1 expression, ↑gut microbiota Prevotella, ↓rc4–4 bacteria | [111] |
5.3. Human Studies
6. Toxicity and Safety Profile of Citrus Fruit Bioactive Compounds
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lapuente, M.; Estruch, R.; Shahbaz, M.; Casas, R. Relation of Fruits and Vegetables with Major Cardiometabolic Risk Factors, Markers of Oxidation, and Inflammation. Nutrients 2019, 11, 2381. [Google Scholar] [CrossRef] [Green Version]
- Medina-Remón, A.; Kirwan, R.; Lamuela-Raventós, R.M.; Estruch, R. Dietary patterns and the risk of obesity, type 2 diabetes mellitus, cardiovascular diseases, asthma, and neurodegenerative diseases. Crit. Rev. Food Sci. Nutr. 2018, 58, 262–296. [Google Scholar] [CrossRef]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhou, Z. Antioxidant activity of Citrus fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef]
- Wu, G.A.; Terol, J.; Ibanez, V.; Lopez-Garcia, A.; Perez-Roman, E.; Borreda, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zhao, C.; Shi, H.; Liao, Y.; Xu, F.; Du, H.; Xiao, H.; Zheng, J. Nutrients and bioactives in citrus fruits: Different citrus varieties, fruit parts, and growth stages. Crit. Rev. Food Sci. Nutr. 2021, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, S.Y.; Yang, W.J.; Huang, Q.R.; Ho, C.T. The biological fate and bioefficacy of citrus flavonoids: Bioavailability, biotransformation, and delivery systems. Food Funct. 2021, 12, 3307–3323. [Google Scholar] [CrossRef]
- United States Department of Agriculture Foreign Agricultural Service. Citrus: World Markets and Trade. 2021. Available online: https://www.fas.usda.gov/data/citrus-world-markets-and-trade (accessed on 23 December 2021).
- Cebadera-Miranda, L.; Dominguez, L.; Dias, M.I.; Barros, L.; Ferreira, I.; Igual, M.; Martinez-Navarrete, N.; Fernandez-Ruiz, V.; Morales, P.; Camara, M. Sanguinello and Tarocco (Citrus sinensis L. Osbeck): Bioactive compounds and colour appearance of blood oranges. Food Chem. 2019, 270, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Lado, J.; Gambetta, G.; Zacarias, L. Key determinants of citrus fruit quality: Metabolites and main changes during maturation. Sci. Hortic. 2018, 233, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Bora, H.; Kamle, M.; Mahato, D.K.; Tiwari, P.; Kumar, P. Citrus Essential Oils (CEOs) and Their Applications in Food: An Overview. Plants 2020, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Mas, M.C.; Rambla, J.L.; Lopez-Gresa, M.P.; Blazquez, M.A.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef]
- Stevens, Y.; Van Rymenant, E.; Grootaert, C.; Van Camp, J.; Possemiers, S.; Masclee, A.; Jonkers, D. The Intestinal Fate of Citrus Flavanones and Their Effects on Gastrointestinal Health. Nutrients 2019, 11, 1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreca, D.; Mandalari, G.; Calderaro, A.; Smeriglio, A.; Trombetta, D.; Felice, M.R.; Gattuso, G. Citrus Flavones: An Update on Sources, Biological Functions, and Health Promoting Properties. Plants 2020, 9, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addi, M.; Elbouzidi, A.; Abid, M.; Tungmunnithum, D.; Elamrani, A.; Hano, C. An Overview of Bioactive Flavonoids from Citrus Fruits. Appl. Sci. 2022, 12, 29. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.J.; Chen, J.B.; Cao, J.P.; Li, X.; Sun, C.D. Citrus flavonoids and their antioxidant evaluation. Crit. Rev. Food Sci. Nutr. 2021, 1–22. [Google Scholar] [CrossRef]
- Pontifex, M.G.; Malik, M.; Connell, E.; Muller, M.; Vauzour, D. Citrus Polyphenols in Brain Health and Disease: Current Perspectives. Front. Neurosci. 2021, 15, 640648. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Y.; Wang, F.; Lian, Y.H.; Xiao, H.; Zheng, J.K. Biosynthesis of citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef]
- Musumeci, L.; Maugeri, A.; Cirmi, S.; Lombardo, G.E.; Russo, C.; Gangemi, S.; Calapai, G.; Navarra, M. Citrus fruits and their flavonoids in inflammatory bowel disease: An overview. Nat. Prod. Res. 2020, 34, 122–136. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Vasconcelos, A.B.S.; Wu, D.T.; Li, H.B.; Antony, P.J.; Li, H.; Geng, F.; Gurgel, R.Q.; Narain, N.; Gan, R.Y. Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies. Nutrients 2020, 12, 2907. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Bautista, R.J.H.; Sandhu, M.A.; Hussein, O.E. Beneficial Effects of Citrus Flavonoids on Cardiovascular and Metabolic Health. Oxid. Med. Cell. Longev. 2019, 2019, 5484138. [Google Scholar] [CrossRef] [Green Version]
- Bellavite, P.; Donzelli, A. Hesperidin and SARS-CoV-2: New Light on the Healthy Function of Citrus Fruits. Antioxidants 2020, 9, 742. [Google Scholar] [CrossRef]
- Den Hartogh, D.J.; Tsiani, E. Antidiabetic Properties of Naringenin: A Citrus Fruit Polyphenol. Biomolecules 2019, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidun, N.H.; Thent, Z.C.; Abd Latiff, A. Combating oxidative stress disorders with citrus flavonoid: Naringenin. Life Sci. 2018, 208, 111–122. [Google Scholar] [CrossRef]
- Nakajima, A.; Ohizumi, Y. Potential Benefits of Nobiletin, A Citrus Flavonoid, against Alzheimer’s Disease and Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, J.X.H.; Tan, L.T.H.; Goh, J.K.; Chan, K.G.; Pusparajah, P.; Lee, L.H.; Goh, B.H. Nobiletin and Derivatives: Functional Compounds from Citrus Fruit Peel for Colon Cancer Chemoprevention. Cancers 2019, 11, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimek-Szczykutowicz, M.; Szopa, A.; Ekiert, H. Citrus limon (Lemon) Phenomenon—A Review of the Chemistry, Pharmacological Properties, Applications in the Modern Pharmaceutical, Food, and Cosmetics Industries, and Biotechnological Studies. Plants 2020, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Gao, W.; Zeng, S.L.; Li, P.; Liu, E.H. Chemical structures, bioactivities and molecular mechanisms of citrus polymethoxyflavones. J. Funct. Foods 2018, 40, 498–509. [Google Scholar] [CrossRef]
- Peng, Z.X.; Zhang, H.P.; Li, W.Y.; Yuan, Z.Y.; Xie, Z.Z.; Zhang, H.Y.; Cheng, Y.J.; Chen, J.J.; Xu, J. Comparative profiling and natural variation of polymethoxylated flavones in various citrus germplasms. Food Chem. 2021, 354, 129499. [Google Scholar] [CrossRef]
- Zhang, H.J.; Tian, G.F.; Zhao, C.Y.; Han, Y.H.; DiMarco-Crook, C.; Lu, C.; Bao, Y.M.; Li, C.X.; Xiao, H.; Zheng, J.K. Characterization of polymethoxyflavone demethylation during drying processes of citrus peels. Food Funct. 2019, 10, 5707–5717. [Google Scholar] [CrossRef]
- Smeriglio, A.; Cornara, L.; Denaro, M.; Barreca, D.; Burlando, B.; Xiao, J.B.; Trombetta, D. Antioxidant and cytoprotective activities of an ancient Mediterranean citrus (Citrus lumia Risso) albedo extract: Microscopic observations and polyphenol characterization. Food Chem. 2019, 279, 347–355. [Google Scholar] [CrossRef]
- Mazzotti, F.; Bartella, L.; Talarico, I.R.; Napoli, A.; Di Donna, L. High-throughput determination of flavanone-O-glycosides in citrus beverages by paper spray tandem mass spectrometry. Food Chem. 2021, 360, 130060. [Google Scholar] [CrossRef]
- Rong, X.; Xu, J.; Jiang, Y.; Li, F.; Chen, Y.L.; Dou, Q.P.; Li, D.P. Citrus peel flavonoid nobiletin alleviates lipopolysaccharide-induced inflammation by activating IL-6/STAT3/FOXO3a-mediated autophagy. Food Funct. 2021, 12, 1305–1317. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhao, C.Y.; Yang, M.K.; Zhang, L.; Wei, R.J.; Meng, K.; Bao, Y.M.; Zhang, L.N.; Zheng, J.K. Four Citrus Flavanones Exert Atherosclerosis Alleviation Effects in ApoE(−/−) Mice via Different Metabolic and Signaling Pathways. J. Agric. Food Chem. 2021, 69, 5226–5237. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, J.Y.; Zhang, X.; Zhao, D.G.; Ma, Y.Y.; Li, D.L.; Ho, C.T.; Huang, Q.R. Aged citrus peel (chenpi) extract causes dynamic alteration of colonic microbiota in high-fat diet induced obese mice. Food Funct. 2020, 11, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.L.; Li, S.Z.; Xiao, P.T.; Cai, Y.Y.; Chu, C.; Chen, B.Z.; Li, P.; Li, J.; Liu, E.H. Citrus polymethoxyflavones attenuate metabolic syndrome by regulating gut microbiome and amino acid metabolism. Sci. Adv. 2020, 6, eaax6208. [Google Scholar] [CrossRef] [Green Version]
- Song, M.Y.; Lan, Y.Q.; Wu, X.; Han, Y.H.; Wang, M.Q.; Zheng, J.K.; Li, Z.Z.; Li, F.; Zhou, J.Z.; Xiao, J.; et al. The chemopreventive effect of 5-demethylnobiletin, a unique citrus flavonoid, on colitis-driven colorectal carcinogenesis in mice is associated with its colonic metabolites. Food Funct. 2020, 11, 4940–4952. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Pan, M.-H.; Lo, C.-Y.; Tan, D.; Wang, Y.; Shahidi, F.; Ho, C.-T. Chemistry and health effects of polymethoxyflavones and hydroxylated polymethoxyflavones. J. Funct. Foods 2009, 1, 2–12. [Google Scholar] [CrossRef]
- Deng, M.; Jia, X.C.; Dong, L.H.; Liu, L.; Huang, F.; Chi, J.W.; Ma, Q.; Zhao, D.; Zhang, M.W.; Zhang, R.F. Structural elucidation of flavonoids from Shatianyu (Citrus grandis L. Osbeck) pulp and screening of key antioxidant components. Food Chem. 2022, 366, 130605. [Google Scholar] [CrossRef]
- Chen, J.; Yuan, Z.; Zhang, H.; Li, W.; Shi, M.; Peng, Z.; Li, M.; Tian, J.; Deng, X.; Cheng, Y.; et al. Cit1,2RhaT and two novel CitdGlcTs participate in flavor-related flavonoid metabolism during citrus fruit development. J. Exp. Bot. 2019, 70, 2759–2771. [Google Scholar] [CrossRef]
- Ledesma-Escobar, C.A.; Priego-Capote, F.; Olvera, V.J.R.; de Castro, M.D.L. Targeted Analysis of the Concentration Changes of Phenolic Compounds in Persian Lime (Citrus latifolia) during Fruit Growth. J. Agric. Food Chem. 2018, 66, 1813–1820. [Google Scholar] [CrossRef]
- Multari, S.; Licciardello, C.; Caruso, M.; Martens, S. Monitoring the changes in phenolic compounds and carotenoids occurring during fruit development in the tissues of four citrus fruits. Food Res. Int. 2020, 134, 109228. [Google Scholar] [CrossRef]
- Saini, R.K.; Nile, S.H.; Park, S.W. Carotenoids from fruits and vegetables: Chemistry, analysis, occurrence, bioavailability and biological activities. Food Res. Int. 2015, 76, 735–750. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Etzbach, L.; Stolle, R.; Anheuser, K.; Herdegen, V.; Schieber, A.; Weber, F. Impact of Different Pasteurization Techniques and Subsequent Ultrasonication on the In Vitro Bioaccessibility of Carotenoids in Valencia Orange (Citrus sinensis (L.) Osbeck) Juice. Antioxidants 2020, 9, 534. [Google Scholar] [CrossRef]
- Lux, P.E.; Carle, R.; Zacarias, L.; Rodrigo, M.J.; Schweiggert, R.M.; Steingass, C.B. Genuine Carotenoid Profiles in Sweet Orange Citrus sinensis (L.) Osbeck cv. Navel Peel and Pulp at Different Maturity Stages. J. Agric. Food Chem. 2019, 67, 13164–13175. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.J.; Mi, J.N.; Deng, X.X.; Al-Babili, S. LC-MS-Based Profiling Provides New Insights into Apocarotenoid Biosynthesis and Modifications in Citrus Fruits. J. Agric. Food Chem. 2021, 69, 1842–1851. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.T.; Wang, S.S.; Wang, R.Q.; Xu, C.J. Accumulation of red apocarotenoid beta-citraurin in peel of a spontaneous mutant of huyou (Citrus changshanensis) and the effects of storage temperature and ethylene application. Food Chem. 2020, 309, 125705. [Google Scholar] [CrossRef]
- Dhakane-Lad, J.; Kar, A. Supercritical CO2 extraction of lycopene from pink grapefruit (Citrus paradise Macfad) and its degradation studies during storage. Food Chem. 2021, 361, 130113. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lin, J.R.; Xu, L.L.; Peng, Q.Q.; Huang, H.Y.; Tong, L.J.; Lu, Q.; Wang, C.C.; Yang, L. On higher nutritional and medical properties of a carotenoid-rich mutant pomelo (Citrus maxima (L.) Osbeck). Ind. Crop. Prod. 2019, 127, 142–147. [Google Scholar] [CrossRef]
- Petry, F.C.; de Nadai, F.B.; Cristofani-Yaly, M.; Latado, R.R.; Mercadante, A.Z. Carotenoid biosynthesis and quality characteristics of new hybrids between tangor (Citrus reticulata × C. sinensis) cv. ‘Murcott’ and sweet orange (C. sinensis) cv. ‘Pera’. Food Res. Int. 2019, 122, 461–470. [Google Scholar] [CrossRef]
- Raspo, M.A.; Vignola, M.B.; Andreatta, A.E.; Juliani, H.R. Antioxidant and antimicrobial activities of citrus essential oils from Argentina and the United States. Food Biosci. 2020, 36, 100651. [Google Scholar] [CrossRef]
- Gao, Z.P.; Zhong, W.M.; Chen, K.Y.; Tang, P.Y.; Guo, J.J. Chemical composition and anti-biofilm activity of essential oil from Citrus medica L. var. sarcodactylis Swingle against Listeria monocytogenes. Ind Crop. Prod. 2020, 144, 112036. [Google Scholar] [CrossRef]
- Mahato, N.; Sharma, K.; Koteswararao, R.; Sinha, M.; Baral, E.; Cho, M.H. Citrus essential oils: Extraction, authentication and application in food preservation. Crit. Rev. Food Sci. Nutr. 2019, 59, 611–625. [Google Scholar] [CrossRef]
- Li, Z.H.; Cai, M.; Liu, Y.S.; Sun, P.L.; Luo, S.L. Antibacterial Activity and Mechanisms of Essential Oil from Citrus medica L. var. sarcodactylis. Molecules 2019, 24, 1577. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, C.M.S.; Ikeda, N.Y.; Miano, A.C.; Saldana, E.; Moreno, A.M.; Stashenko, E.; Contreras-Castillo, C.J.; Da Gloria, E.M. Unraveling the selective antibacterial activity and chemical composition of citrus essential oils. Sci. Rep. 2019, 9, 17719. [Google Scholar] [CrossRef] [Green Version]
- Denkova-Kostova, R.; Teneva, D.; Tomova, T.; Goranov, B.; Denkova, Z.; Shopska, V.; Slavchev, A.; Hristova-Ivanova, Y. Chemical composition, antioxidant and antimicrobial activity of essential oils from tangerine (Citrus reticulata L.), grapefruit (Citrus paradisi L.), lemon (Citrus lemon L.) and cinnamon (Cinnamomum zeylanicum Blume). Z. Nat. Sect. C J. Biosci. 2021, 76, 175–185. [Google Scholar] [CrossRef]
- Paw, M.; Begum, T.; Gogoi, R.; Pandey, S.K.; Lal, M. Chemical Composition of Citrus limon L. Burmf Peel Essential Oil from North East India. J. Essent. Oil Bear. Plants 2020, 23, 337–344. [Google Scholar] [CrossRef]
- Zoccali, M.; Arigo, A.; Russo, M.; Salafia, F.; Dugo, P.; Mondello, L. Characterization of Limonoids in Citrus Essential Oils by Means of Supercritical Fluid Chromatography Tandem Mass Spectrometry. Food Anal. Methods 2018, 11, 3257–3266. [Google Scholar] [CrossRef]
- Rossi, R.C.; da Rosa, S.R.; Weimer, P.; Moura, J.G.L.; de Oliveira, V.R.; de Castilhos, J. Assessment of compounds and cytotoxicity of Citrus deliciosa Tenore essential oils: From an underexploited by-product to a rich source of high-value bioactive compounds. Food Biosci. 2020, 38, 100779. [Google Scholar] [CrossRef]
- Yu, M.; Xia, Y.; Xie, W.; Li, Y.; Yu, X.; Zheng, J.; Zhang, Y. Enzymatic extraction of pectic oligosaccharides from finger citron (Citrus medica L. var. sarcodactylis Swingle) pomace with antioxidant potential. Food Funct. 2021, 12, 9855–9865. [Google Scholar] [CrossRef] [PubMed]
- Zema, D.A.; Calabro, P.S.; Folino, A.; Tamburino, V.; Zappia, G.; Zimbone, S.M. Valorisation of citrus processing waste: A review. Waste Manag. 2018, 80, 252–273. [Google Scholar] [CrossRef] [PubMed]
- Mahato, N.; Sharma, K.; Sinha, M.; Cho, M.H. Citrus waste derived nutra-/pharmaceuticals for health benefits: Current trends and future perspectives. J. Funct. Foods 2018, 40, 307–316. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Yadav, M.P. Insights into the chemical composition and bioactivities of citrus peel essential oils. Food Res. Int. 2021, 143, 110231. [Google Scholar] [CrossRef] [PubMed]
- Saini, A.; Panesar, P.S.; Bera, M.B. Valuation of Citrus reticulata (kinnow) peel for the extraction of lutein using ultrasonication technique. Biomass Convers. Biorefinery 2021, 11, 2157–2165. [Google Scholar] [CrossRef]
- Costanzo, G.; Iesce, M.R.; Naviglio, D.; Ciaravolo, M.; Vitale, E.; Arena, C. Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components. Antioxidants 2020, 9, 517. [Google Scholar] [CrossRef] [PubMed]
- Murador, D.C.; Salafia, F.; Zoccali, M.; Martins, P.L.G.; Ferreira, A.G.; Dugo, P.; Mondello, L.; De Rosso, V.V.; Giuffrida, D. Green Extraction Approaches for Carotenoids and Esters: Characterization of Native Composition from Orange Peel. Antioxidants 2019, 8, 613. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, D.; Picone, P.; Giardina, C.; Scordino, M.; Mudò, G.; Pagliaro, M.; Scurria, A.; Meneguzzo, F.; Ilharco, L.M.; Fidalgo, A.; et al. New Neuroprotective Effect of Lemon IntegroPectin on Neuronal Cellular Model. Antioxidants 2021, 10, 669. [Google Scholar] [CrossRef] [PubMed]
- Colodel, C.; Vriesmann, L.C.; Teofilo, R.F.; Petkowicz, C.L.D. Extraction of pectin from ponkan (Citrus reticulata Blanco cv. Ponkan) peel: Optimization and structural characterization. Int. J. Biol. Macromol. 2018, 117, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Long, X.Y.; Zeng, X.G.; Yan, H.T.; Xu, M.J.; Zeng, Q.T.; Xu, C.; Xu, Q.M.; Liang, Y.; Zhang, J. Flavonoids composition and antioxidant potential assessment of extracts from Gannanzao Navel Orange (Citrus sinensis Osbeck Cv. Gannanzao) peel. Nat. Prod. Res. 2021, 35, 702–706. [Google Scholar] [CrossRef]
- El-Kersh, D.M.; Ezzat, S.M.; Salama, M.M.; Mahrous, E.A.; Attia, Y.M.; Ahmed, M.S.; Elmazar, M.M. Anti-estrogenic and anti-aromatase activities of citrus peels major compounds in breast cancer. Sci. Rep. 2021, 11, 7121. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pan, H.L.; Hao, S.X.; Pan, D.M.; Wang, G.J.; Yu, W.Q. Evaluation of phenolic composition and antioxidant properties of different varieties of Chinese citrus. Food Chem. 2021, 364, 130413. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Li, X.; Zhao, P.; Zhang, X.Q.; Qiao, O.; Huang, L.Q.; Guo, L.P.; Gao, W.Y. A review of chemical constituents and health-promoting effects of citrus peels. Food Chem. 2021, 365, 130585. [Google Scholar] [CrossRef]
- Gogoi, M.; Boruah, J.L.H.; Bora, P.K.; Das, D.J.; Famhawite, V.; Biswas, A.; Puro, N.; Kalita, J.; Haldar, S.; Baishya, R. Citrus macroptera induces apoptosis via death receptor and mitochondrial mediated pathway as prooxidant in human non-small cell lung cancer cells. Food Biosci. 2021, 43, 101293. [Google Scholar] [CrossRef]
- Abdelghffar, E.A.; El-Nashar, H.A.S.; Al-Mohammadi, A.G.A.; Eldahshan, O.A. Orange fruit (Citrus sinensis) peel extract attenuates chemotherapy-induced toxicity in male rats. Food Funct. 2021, 12, 9443–9455. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Res. Int. 2020, 132, 109114. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Park, H.; Jang, B.K.; Ju, Y.B.; Shin, M.H.; Oh, E.J.; Lee, E.J.; Kim, S.R. Recent advances in the biological valorization of citrus peel waste into fuels and chemicals. Bioresour. Technol. 2021, 323, 124603. [Google Scholar] [CrossRef]
- Zayed, A.; Badawy, M.T.; Farag, M.A. Valorization and extraction optimization of Citrus seeds for food and functional food applications. Food Chem. 2021, 355, 129609. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Kim, H.J.; Ko, M.J.; Chung, M.S. Recovery of hesperidin and narirutin from waste Citrus unshiu peel using subcritical water extraction aided by pulsed electric field treatment. Food Sci. Biotechnol. 2021, 30, 217–226. [Google Scholar] [CrossRef]
- Kim, D.-S.; Lim, S.-B. Semi-Continuous Subcritical Water Extraction of Flavonoids from Citrus unshiu Peel: Their Antioxidant and Enzyme Inhibitory Activities. Antioxidants 2020, 9, 360. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Jo, E.H.; Moon, J.E.; Cha, H.; Chang, M.H.; Cho, H.T.; Lee, M.K.; Jung, W.S.; Lee, J.H.; Heo, W.; et al. In Vitro and In Vivo Inhibitory Effect of Citrus Junos Tanaka Peel Extract against Oxidative Stress-Induced Apoptotic Death of Lung Cells. Antioxidants 2020, 9, 1231. [Google Scholar] [CrossRef]
- Anacleto, S.L.; Milenkovic, D.; Kroon, P.A.; Needs, P.W.; Lajolo, F.M.; Hassimotto, N.M.A. Citrus flavanone metabolites protect pancreatic-beta cells under oxidative stress induced by cholesterol. Food Funct. 2020, 11, 8612–8624. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Y.; Shen, S.Y.; Zhi, Z.J.; Cheng, H.; Chen, S.G.; Ye, X.Q. Antioxidant and pancreatic lipase inhibitory effects of flavonoids from different citrus peel extracts: An in vitro study. Food Chem. 2020, 326, 126785. [Google Scholar] [CrossRef]
- Denaro, M.; Smeriglio, A.; Trombetta, D. Antioxidant and Anti-Inflammatory Activity of Citrus Flavanones Mix and Its Stability after In Vitro Simulated Digestion. Antioxidants 2021, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.L.; Chen, S.Y.; Ho, C.Y.; Yen, G.C. Citrus flavonoids suppress IL-5 and ROS through distinct pathways in PMA/ionomycin-induced EL-4 cells. Food Funct. 2020, 11, 824–833. [Google Scholar] [CrossRef]
- Hu, H.J.; Zhang, S.S.; Pan, S.Y. Characterization of Citrus Pectin Oligosaccharides and Their Microbial Metabolites as Modulators of Immunometabolism on Macrophages. J. Agric. Food Chem. 2021, 69, 8403–8414. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Alloisio, S.; Raimondo, F.M.; Denaro, M.; Xiao, J.B.; Cornara, L.; Trombetta, D. Essential oil of Citrus lumia Risso: Phytochemical profile, antioxidant properties and activity on the central nervous system. Food Chem. Toxicol. 2018, 119, 407–416. [Google Scholar] [CrossRef]
- Piccialli, I.; Tedeschi, V.; Caputo, L.; Amato, G.; De Martino, L.; De Feo, V.; Secondo, A.; Pannaccione, A. The Antioxidant Activity of Limonene Counteracts Neurotoxicity Triggered byAβ1-42 Oligomers in Primary Cortical Neurons. Antioxidants 2021, 10, 937. [Google Scholar] [CrossRef]
- Cirmi, S.; Maugeri, A.; Lombardo, G.E.; Russo, C.; Musumeci, L.; Gangemi, S.; Calapai, G.; Barreca, D.; Navarra, M. A Flavonoid-Rich Extract of Mandarin Juice Counteracts 6-OHDA-Induced Oxidative Stress in SH-SY5Y Cells and Modulates Parkinson-Related Genes. Antioxidants 2021, 10, 539. [Google Scholar] [CrossRef]
- Kitagawa, T.; Matsumoto, T.; Imahori, D.; Kobayashi, M.; Okayama, M.; Ohta, T.; Yoshida, T.; Watanabe, T. Limonoids isolated from the Fortunella crassifolia and the Citrus junos with their cell death-inducing activity on Adriamycin-treated cancer cell. J. Nat. Med. 2021, 75, 998–1004. [Google Scholar] [CrossRef]
- Murthy, K.N.C.; Jayaprakasha, G.K.; Safe, S.; Patil, B.S. Citrus limonoids induce apoptosis and inhibit the proliferation of pancreatic cancer cells. Food Funct. 2021, 12, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Wei, X.; Zhang, F.; Hao, J.; Huang, F.; Zhang, C.; Liang, W. A dual character of flavonoids in influenza A virus replication and spread through modulating cell-autonomous immunity by MAPK signaling pathways. Sci. Rep. 2014, 4, 7237. [Google Scholar] [CrossRef]
- Lin, C.W.; Tsai, F.J.; Tsai, C.H.; Lai, C.C.; Wan, L.; Ho, T.Y.; Hsieh, C.C.; Chao, P.D. Anti-SARS coronavirus 3C-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef]
- De Clercq, E. Potential antivirals and antiviral strategies against SARS coronavirus infections. Expert Rev. Anti-Infect. Ther. 2006, 4, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Tutunchi, H.; Naeini, F.; Ostadrahimi, A.; Hosseinzadeh-Attar, M.J. Naringenin, a flavanone with antiviral and anti-inflammatory effects: A promising treatment strategy against COVID-19. Phytother. Res. 2020, 34, 3137–3147. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Silva, A.M.; Nunes, F.M. Citrus reticulata Blanco peels as a source of antioxidant and anti-proliferative phenolic compounds. Ind. Crop. Prod. 2018, 111, 141–148. [Google Scholar] [CrossRef]
- Saini, R.K.; Rengasamy, K.R.R.; Mahomoodally, F.M.; Keum, Y.S. Protective effects of lycopene in cancer, cardiovascular, and neurodegenerative diseases: An update on epidemiological and mechanistic perspectives. Pharm. Res. 2020, 155, 104730. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.S.; Daglia, M.; Rengasamy, K.R. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharm. Res. 2020, 157, 104830. [Google Scholar] [CrossRef]
- Shin, J.; Song, M.H.; Oh, J.W.; Keum, Y.S.; Saini, R.K. Pro-Oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants 2020, 9, 532. [Google Scholar] [CrossRef]
- Tian, D.M.; Wang, F.F.; Duan, M.L.; Cao, L.Y.; Zhang, Y.W.; Yao, X.S.; Tang, J.S. Coumarin Analogues from the Citrus grandis (L.) Osbeck and Their Hepatoprotective Activity. J. Agric. Food Chem. 2019, 67, 1937–1947. [Google Scholar] [CrossRef]
- Cataneo, A.H.D.; Kuczera, D.; Koishi, A.C.; Zanluca, C.; Silveira, G.F.; de Arruda, T.B.; Suzukawa, A.A.; Bortot, L.O.; Dias-Baruffi, M.; Verri, W.A.; et al. The citrus flavonoid naringenin impairs the in vitro infection of human cells by Zika virus. Sci. Rep. 2019, 9, 16348. [Google Scholar] [CrossRef]
- Gansukh, E.; Nile, A.; Sivanesan, I.; Rengasamy, K.R.R.; Kim, D.H.; Keum, Y.S.; Saini, R.K. Chemopreventive Effect of beta-Cryptoxanthin on Human Cervical Carcinoma (HeLa) Cells Is Modulated through Oxidative Stress-Induced Apoptosis. Antioxidants 2019, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.L.; Zhu, X.A.; Liu, G.; Kan, Q.X.; Chen, T.; Chen, Y.J.; Cao, Y. Dietary citrus peel essential oil ameliorates hypercholesterolemia and hepatic steatosis by modulating lipid and cholesterol homeostasis. Food Funct. 2020, 11, 7217–7230. [Google Scholar] [CrossRef] [PubMed]
- Dhuique-Mayer, C.; Gence, L.; Portet, K.; Tousch, D.; Poucheret, P. Preventive action of retinoids in metabolic syndrome/type 2 diabetic rats fed with citrus functional food enriched in beta-cryptoxanthin. Food Funct. 2020, 11, 9263–9271. [Google Scholar] [CrossRef]
- Revathy, J.; Srinivasan, S.; Abdullah, S.H.S.; Muruganathan, U. Antihyperglycemic effect of hesperetin, a citrus flavonoid, extenuates hyperglycemia and exploring the potential role in antioxidant and antihyperlipidemic in streptozotocin-induced diabetic rats. Biomed. Pharmacother. 2018, 97, 98–106. [Google Scholar] [CrossRef]
- Bi, W.Y.; Zhou, J.X.; Zhao, L.; Wang, C.T.; Wu, W.; Zhang, L.B.; Ji, B.P.; Zhang, N.H.; Zhou, F. Preventive effect of different citrus essential oils on primary dysmenorrhea: In vivo and in vitro study. Food Biosci. 2021, 42, 101135. [Google Scholar] [CrossRef]
- Zhang, L.L.; Yang, Z.Y.; Fan, G.; Ren, J.N.; Yin, K.J.; Pan, S.Y. Antidepressant-like Effect of Citrus sinensis (L.) Osbeck Essential Oil and Its Main Component Limonene on Mice. J. Agric. Food Chem. 2019, 67, 13817–13828. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Ikram, M.; Ullah, R.; Rehman, S.U.; Kim, M.O. Hesperetin, a Citrus Flavonoid, Attenuates LPS-Induced Neuroinflammation, Apoptosis and Memory Impairments by Modulating TLR4/NF-kappa B Signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, S.; Nakashima, T.; Nakamura, K.; Shinoka, W.; Kotani, M.; Sawamoto, A.; Nakajima, M.; Furukawa, Y. Inhibitory Effects of Auraptene and Naringin on Astroglial Activation, Tau Hyperphosphorylation, and Suppression of Neurogenesis in the Hippocampus of Streptozotocin-Induced Hyperglycemic Mice. Antioxidants 2018, 7, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Cao, X.D.; Fang, X.G.; Guo, A.L.; Li, E.H. Inhibitory effects of fermented Ougan (Citrus reticulata cv. Suavissima) juice on high-fat diet-induced obesity associated with white adipose tissue browning and gut microbiota modulation in mice. Food Funct. 2021, 12, 9300–9314. [Google Scholar] [CrossRef]
- Lazar, V.; Ditu, L.-M.; Pircalabioru, G.G.; Picu, A.; Petcu, L.; Cucu, N.; Chifiriuc, M.C. Gut Microbiota, Host Organism, and Diet Trialogue in Diabetes and Obesity. Front. Nutr. 2019, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Tung, Y.C.; Chang, W.T.; Li, S.M.; Wu, J.C.; Badmeav, V.; Ho, C.T.; Pan, M.H. Citrus peel extracts attenuated obesity and modulated gut microbiota in mice with high-fat diet-induced obesity. Food Funct. 2018, 9, 3363–3373. [Google Scholar] [CrossRef]
- Miler, M.; Zivanovic, J.; Ajdzanovic, V.; Milenkovic, D.; Jaric, I.; Sosic-Jurjevic, B.; Milosevic, V. Citrus Flavanones Upregulate Thyrotroph Sirt1 and Differently Affect Thyroid Nrf2 Expressions in Old-Aged Wistar Rats. J. Agric. Food Chem. 2020, 68, 8242–8254. [Google Scholar] [CrossRef]
- Kawabata, A.; Hung, T.V.; Nagata, Y.; Fukuda, N.; Suzuki, T. Citrus kawachiensis Peel Powder Reduces Intestinal Barrier Defects and Inflammation in Colitic Mice. J. Agric. Food Chem. 2018, 66, 10991–10999. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Nie, M.M.; Liu, C.Q.; Jiang, N.; Liu, C.J.; Li, D.J. Citrus Flavanones Enhance beta-Carotene Uptake in Vitro Experiment Using Caco-2 Cell: Structure-Activity Relationship and Molecular Mechanisms. J. Agric. Food Chem. 2019, 67, 4280–4288. [Google Scholar] [CrossRef]
- Castro, M.A.; Rodenak-Kladniew, B.; Massone, A.; Polo, M.; de Bravo, M.G.; Crespo, R. Citrus reticulata peel oil inhibits non-small cell lung cancer cell proliferation in culture and implanted in nude mice. Food Funct. 2018, 9, 2290–2299. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.G.; Wang, C.Y.; Ning, C.Q.; Liu, K.X.; Wang, X.Y.; Liu, Z.H.; Sun, H.J.; Ma, X.D.; Sun, P.Y.; Meng, Q. Hepatoprotection of auraptene from peels of citrus fruits against thioacetamide-induced hepatic fibrosis in mice by activating farnesoid X receptor. Food Funct. 2018, 9, 2684–2694. [Google Scholar] [CrossRef]
- Wang, J.Q.; Fu, T.; Dong, R.C.; Wang, C.Y.; Liu, K.X.; Sun, H.J.; Huo, X.K.; Ma, X.D.; Yang, X.B.; Meng, Q. Hepatoprotection of auraptene from the peels of citrus fruits against 17 alpha-ethinylestradiol-induced cholestasis in mice by activating farnesoid X receptor. Food Funct. 2019, 10, 3839–3850. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.D.; Ramadan, D.; Goncalves, V.D.; Maquera-Huacho, P.M.; Assis, R.P.; Lima, T.F.O.; Brunetti, I.L.; Spolidorio, D.M.P.; Cesar, T.; Manthey, J.A.; et al. Impact of citrus flavonoid supplementation on inflammation in lipopolysaccharide-induced periodontal disease in mice. Food Funct. 2021, 12, 5007–5017. [Google Scholar] [CrossRef]
- He, W.; Li, Y.M.; Liu, M.Y.; Yu, H.Y.; Chen, Q.; Chen, Y.; Ruan, J.Y.; Ding, Z.J.; Zhang, Y.; Wang, T. Citrus aurantium L. and Its Flavonoids Regulate TNBS-Induced Inflammatory Bowel Disease through Anti-Inflammation and Suppressing Isolated Jejunum Contraction. Int. J. Mol. Sci. 2018, 19, 3057. [Google Scholar] [CrossRef]
- Samie, A.; Sedaghat, R.; Baluchnejadmojarad, T.; Roghani, M. Hesperetin, a citrus flavonoid, attenuates testicular damage in diabetic rats via inhibition of oxidative stress, inflammation, and apoptosis. Life Sci. 2018, 210, 132–139. [Google Scholar] [CrossRef]
- Guirro, M.; Gual-Grau, A.; Gibert-Ramos, A.; Alcaide-Hidalgo, J.M.; Canela, N.; Arola, L.; Mayneris-Perxachs, J. Metabolomics Elucidates Dose-Dependent Molecular Beneficial Effects of Hesperidin Supplementation in Rats Fed an Obesogenic Diet. Antioxidants 2020, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.J.; Zhang, S.S.; Liu, F.X.; Zhang, P.P.; Muhammad, Z.; Pan, S.Y. Role of the Gut Microbiota and Their Metabolites in Modulating the Cholesterol-Lowering Effects of Citrus Pectin Oligosaccharides in C57BL/6 Mice. J. Agric. Food Chem. 2019, 67, 11922–11930. [Google Scholar] [CrossRef] [PubMed]
- Cirmi, S.; Navarra, M.; Woodside, J.V.; Cantwell, M.M. Citrus fruits intake and oral cancer risk: A systematic review and meta-analysis. Pharmacol. Res. 2018, 133, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victoria-Montesinos, D.; Abellán Ruiz, M.S.; Luque Rubia, A.J.; Guillén Martínez, D.; Pérez-Piñero, S.; Sánchez Macarro, M.; García-Muñoz, A.M.; Cánovas García, F.; Castillo Sánchez, J.; López-Román, F.J. Effectiveness of Consumption of a Combination of Citrus Fruit Flavonoids and Olive Leaf Polyphenols to Reduce Oxidation of Low-Density Lipoprotein in Treatment-Naïve Cardiovascular Risk Subjects: A Randomized Double-Blind Controlled Study. Antioxidants 2021, 10, 589. [Google Scholar] [CrossRef]
- Sánchez Macarro, M.; Martínez Rodríguez, J.P.; Bernal Morell, E.; Pérez-Piñero, S.; Victoria-Montesinos, D.; García-Muñoz, A.M.; Cánovas García, F.; Castillo Sánchez, J.; López-Román, F.J. Effect of a Combination of Citrus Flavones and Flavanones and Olive Polyphenols for the Reduction of Cardiovascular Disease Risk: An Exploratory Randomized, Double-Blind, Placebo-Controlled Study in Healthy Subjects. Nutrients 2020, 12, 1475. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, R.; Williamson, G. Effect of citrus fruit and juice consumption on risk of developing type 2 diabetes: Evidence on polyphenols from epidemiological and intervention studies. Trends Food Sci. Technol. 2021, 115, 133–146. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.Á.; Giménez-Bastida, J.A.; González-Sarrías, A.; Espín, J.C. New Insights into the Metabolism of the Flavanones Eriocitrin and Hesperidin: A Comparative Human Pharmacokinetic Study. Antioxidants 2021, 10, 435. [Google Scholar] [CrossRef]
- Brasili, E.; Hassimotto, N.M.A.; Del Chierico, F.; Marini, F.; Quagliariello, A.; Sciubba, F.; Miccheli, A.; Putignani, L.; Lajolo, F. Daily Consumption of Orange Juice from Citrus sinensis L. Osbeck cv. Cara Cara and cv. Bahia Differently Affects Gut Microbiota Profiling as Unveiled by an Integrated Meta-Omits Approach. J. Agric. Food Chem. 2019, 67, 1381–1391. [Google Scholar] [CrossRef]
- Garcia-Flores, L.A.; Medina, S.; Gomez, C.; Wheelock, C.E.; Cejuela, R.; Martinez-Sanz, J.M.; Oger, C.; Galano, J.M.; Durand, T.; Hernandez-Saez, A.; et al. Aronia-citrus juice (polyphenol-rich juice) intake and elite triathlon training: A lipidomic approach using representative oxylipins in urine. Food Funct. 2018, 9, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kandhare, A.D.; Mukherjee, A.A.; Bodhankar, S.L. Acute and sub-chronic oral toxicity studies of hesperidin isolated from orange peel extract in Sprague Dawley rats. Regul. Toxicol. Pharmacol. 2019, 105, 77–85. [Google Scholar] [CrossRef]
- Nakajima, A.; Nemoto, K.; Ohizumi, Y. An evaluation of the genotoxicity and subchronic toxicity of the peel extract of Ponkan cultivar ‘Ohta ponkan’ (Citrus reticulata Blanco) that is rich in nobiletin and tangeretin with anti-dementia activity. Regul. Toxicol. Pharmacol. 2020, 114, 104670. [Google Scholar] [CrossRef]
- Li, P.; Wu, H.; Wang, Y.; Peng, W.; Su, W. Toxicological evaluation of naringin: Acute, subchronic, and chronic toxicity in Beagle dogs. Regul. Toxicol. Pharmacol. 2020, 111, 104580. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Eisenbrand, G.; Fukushima, S.; Gooderham, N.J.; Guengerich, F.P.; Hecht, S.S.; Rietjens, I.M.C.M.; Bastaki, M.; Davidsen, J.M.; Harman, C.L.; et al. FEMA GRAS assessment of natural flavor complexes: Citrus-derived flavoring ingredients. Food Chem. Toxicol. 2019, 124, 192–218. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, C.; Badgujar, P.C.; Gundev, P.; Upadhyay, A. Review of toxicological assessment of d-limonene, a food and cosmetics additive. Food Chem. Toxicol. 2018, 120, 668–680. [Google Scholar] [CrossRef] [PubMed]
| Botanical Name | Common Name |
|---|---|
| C. aurantifolia (Christm.) Swingle or C. × lumia Risso. & Poit. | Lime, key lime, lumy, ancient Mediterranean citrus |
| C. × aurantium L. | Bitter/sour orange |
| C. × clementina | Clementine |
| C. × deliciosa Tanore | Montenegrin mandarin |
| C. japonica Thunb. | Kumquat |
| C. junos Siebold ex Tanaka | Junos, yuzu |
| C. × latifolia (Yu.Tanaka) Tanaka | Persian lime |
| C. limon (L.) Osbeck) | Lemon |
| C. × limonia Osbeck | Rangpur lime |
| C. maxima (J. Burman) Merr. or C. grandis (L.) Osbeck | Pomelo, pummelo |
| C. medica L. | Citron, finger citron |
| C. poonensis Hort. ex Tanaka | Ponkan |
| C. × paradisi Macfad. or C. × paradise Macfad | Grapefruit, pink/white grapefruit |
| C. reticulata × C. paradisi | Tangelo |
| C. reticulata Blanco | Mandarin, tangerine, Phlegraean mandarin, ougan |
| C. reticulata × C. sinensis | Tangor |
| C. × sinensis (L.) Osbeck | Orange, Valencia orange, blood orange, sweet orange |
| C. unshiu Marc. | Satsuma mandarin, Mandarin orange |
| Compounds | Review Highlights | Reference |
|---|---|---|
| Essential oil | Extraction, purification, detection methods, composition, and applications of citrus essential oil | [10] |
| Composition of volatile compounds from peel, leaves, and flowers of different citrus species | [11] | |
| Flavanones (hesperidin and naringin) | The intestinal fate, bioavailability, intestinal metabolism, and interaction with the gut microbiota | [12] |
| Flavones | Sources, antioxidant, anti-inflammatory, antimicrobial, anticancer properties | [13] |
| Flavonoids | Chemistry, biosynthesis, composition, extraction techniques, health benefits, and industrial applications | [14] |
| Composition, antioxidant evaluation, and regulation of Nrf2-Keap1 pathway by citrus flavonoids | [15] | |
| Role of citrus flavonoids in brain health: evidence from preclinical and human studies | [16] | |
| Biosynthesis, location, and distribution of flavonoids in citrus plants, factors affecting biosynthesis, and health-promoting properties | [17] | |
| In vitro, in vivo, and human studies of citrus flavonoids in minimizing the incidence of inflammatory bowel disease | [18] | |
| Antidiabetic potential of 19 citrus flavonoids, including diosmin, hesperidin, hesperetin, naringin, naringenin, nobiletin, neohesperidin, quercetin, rutin, and tangeretin | [19] | |
| Therapeutic potential in diabetes and diabetic cardiomyopathy, endothelial dysfunction, atherosclerosis, and platelet function | [20] | |
| Chemistry, metabolism, bioavailability, biotransformation and delivery systems, and health benefits | [6] | |
| Hesperidin and vitamin C | Antiviral properties against acute respiratory syndrome coronavirus 2 (SARS-CoV-2) | [21] |
| Naringenin | Antidiabetic properties; in vitro, in vivo, and human studies | [22] |
| Combating oxidative stress disorders: cardiovascular disease, diabetes mellitus, neurodegenerative disease, pulmonary disease, cancer, and nephropathy | [23] | |
| Nobiletin | Beneficial effects against Alzheimer’s disease (AD) and Parkinson’s disease (PD) | [24] |
| Nobiletin, 5-demethylnobiletin, and derivatives | Beneficial effects against colon cancer, pharmacokinetics, and bioavailability | [25] |
| Nutrients and bioactive | Description of the genus Citrus, the composition of nutrients and bioactive components, and biological activities of lemon extract and essential oil | [26] |
| Nutrients (proteins, lipids, vitamins, minerals, fiber) and bioactive (flavonoids, essential oil, limonoids, carotenoids, synephrine) content, their structural characteristics, and health benefits | [5] | |
| Polymethoxyflavones (PMF) | Biological properties against metabolic disorder, atherosclerosis, inflammation, neuroinflammation, cancer, and oxidation | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saini, R.K.; Ranjit, A.; Sharma, K.; Prasad, P.; Shang, X.; Gowda, K.G.M.; Keum, Y.-S. Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes. Antioxidants 2022, 11, 239. https://doi.org/10.3390/antiox11020239
Saini RK, Ranjit A, Sharma K, Prasad P, Shang X, Gowda KGM, Keum Y-S. Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes. Antioxidants. 2022; 11(2):239. https://doi.org/10.3390/antiox11020239
Chicago/Turabian StyleSaini, Ramesh Kumar, Arina Ranjit, Kavita Sharma, Parchuri Prasad, Xiaomin Shang, Karekal Girinur Mallikarjuna Gowda, and Young-Soo Keum. 2022. "Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes" Antioxidants 11, no. 2: 239. https://doi.org/10.3390/antiox11020239
APA StyleSaini, R. K., Ranjit, A., Sharma, K., Prasad, P., Shang, X., Gowda, K. G. M., & Keum, Y.-S. (2022). Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes. Antioxidants, 11(2), 239. https://doi.org/10.3390/antiox11020239