

NRF2/Itaconate Axis Regulates Metabolism and Inflammatory Properties of T Cells in Children with JIA

Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Healthy Donors
2.2. Cell Isolation
2.3. Cell Culture
2.4. Flow Cytometry
| ACTB | For 5′ AGA TGG CCA CGG CTG CT 3′ |
| Rev 5′ AAC CGC TCA TTG CCA ATG G 3′ | |
| IL17a | For 5′ GAATCTCCACCGCAATGAGGA 3′ |
| Rev 5′ TGGTAGTCCACGTTCCCATCAG 3‘ | |
| NRF2 | For 5′ TCC AGT CAG AAA CCA GTG GAT 3′ |
| Rev 5′ AAT GTC TGC GCC AAA AGC TG 3′ | |
| NQO1 | For 5′ GGT TTG AGC GAG TGT TCA TAG G 3′ |
| Rev 5′ GCA GAG AGT ACA TGG AGA CAC 3′ | |
| HO-1 | For 5′ CAG TGC CAC CAA GTT CAA GC 3′ |
| Rev 5′ GTT GAG CAG GAA CGC AGT CTT 3′ | |
| TRX1 | For 5′ CCC TTT CTT TCA TTC CCT CTC TG 3′ |
| Rev 5′ ATT CAC CCA CCT TTT GTC CCT 3′ | |
| IFN-γ | For 5′ ACT AGG CAG CCA ACC TAA GCA AGA 3′ |
| Rev 5′ CAT CAG GGT CAC CTG ACA CAT TCA 3′ |
2.5. Transfection of Primary Cells
2.6. SDS Page and Western Blot
2.7. Mass Spectrometry
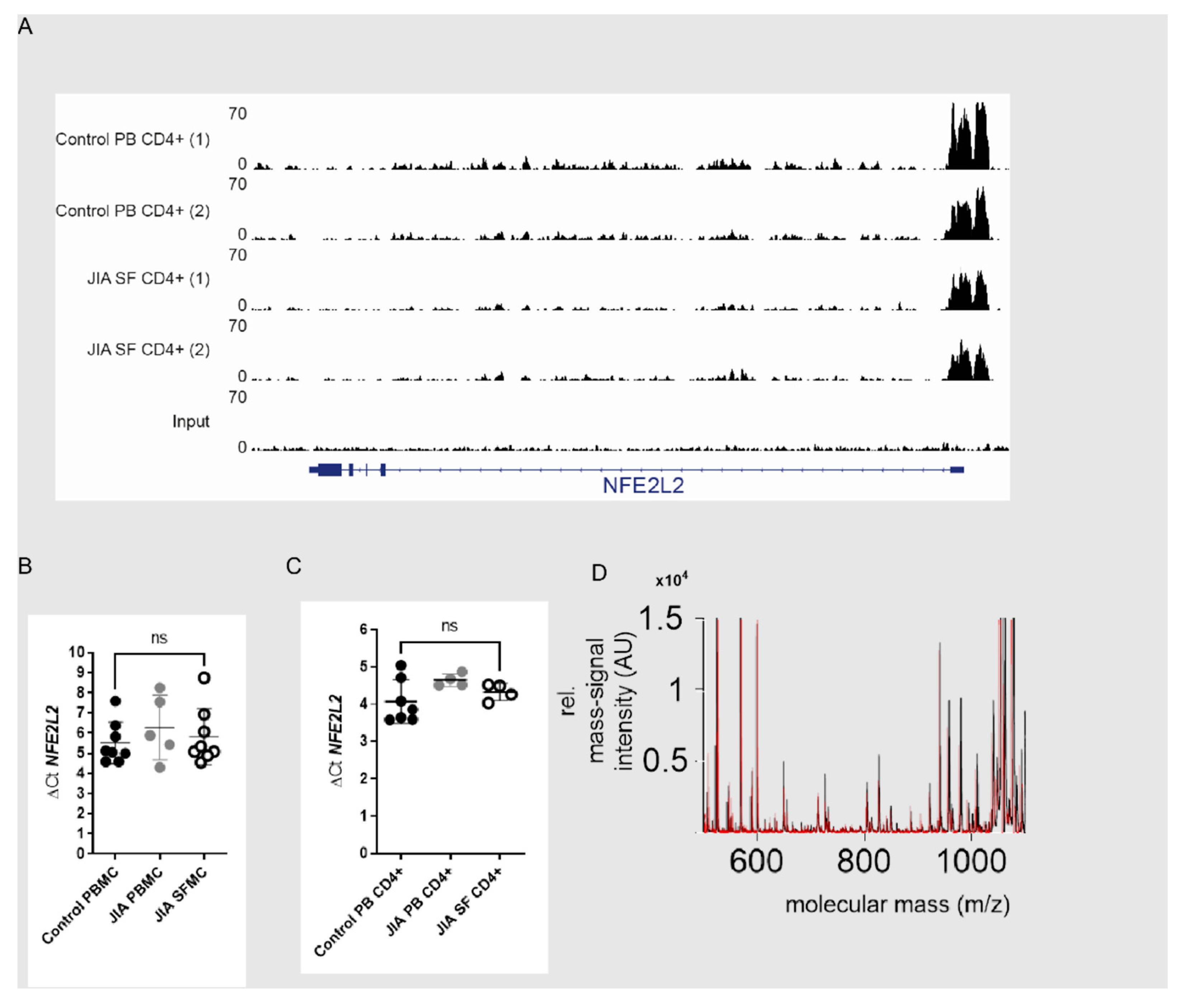
2.8. ChIP Sequencing
2.9. Statistics
3. Results
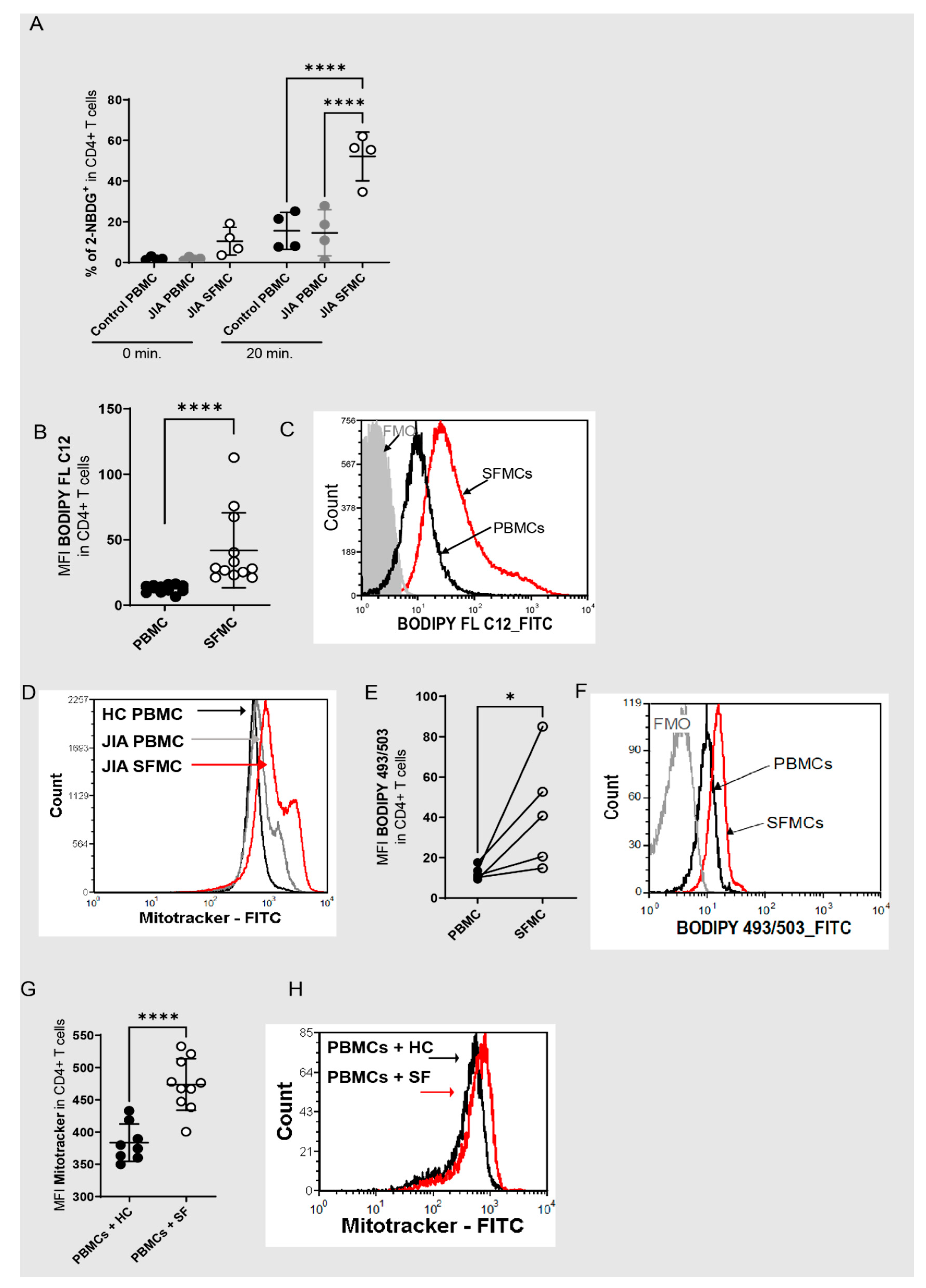
3.1. Redox Metabolism Is Dysregulated in JIA T Cells within Inflamed Joints
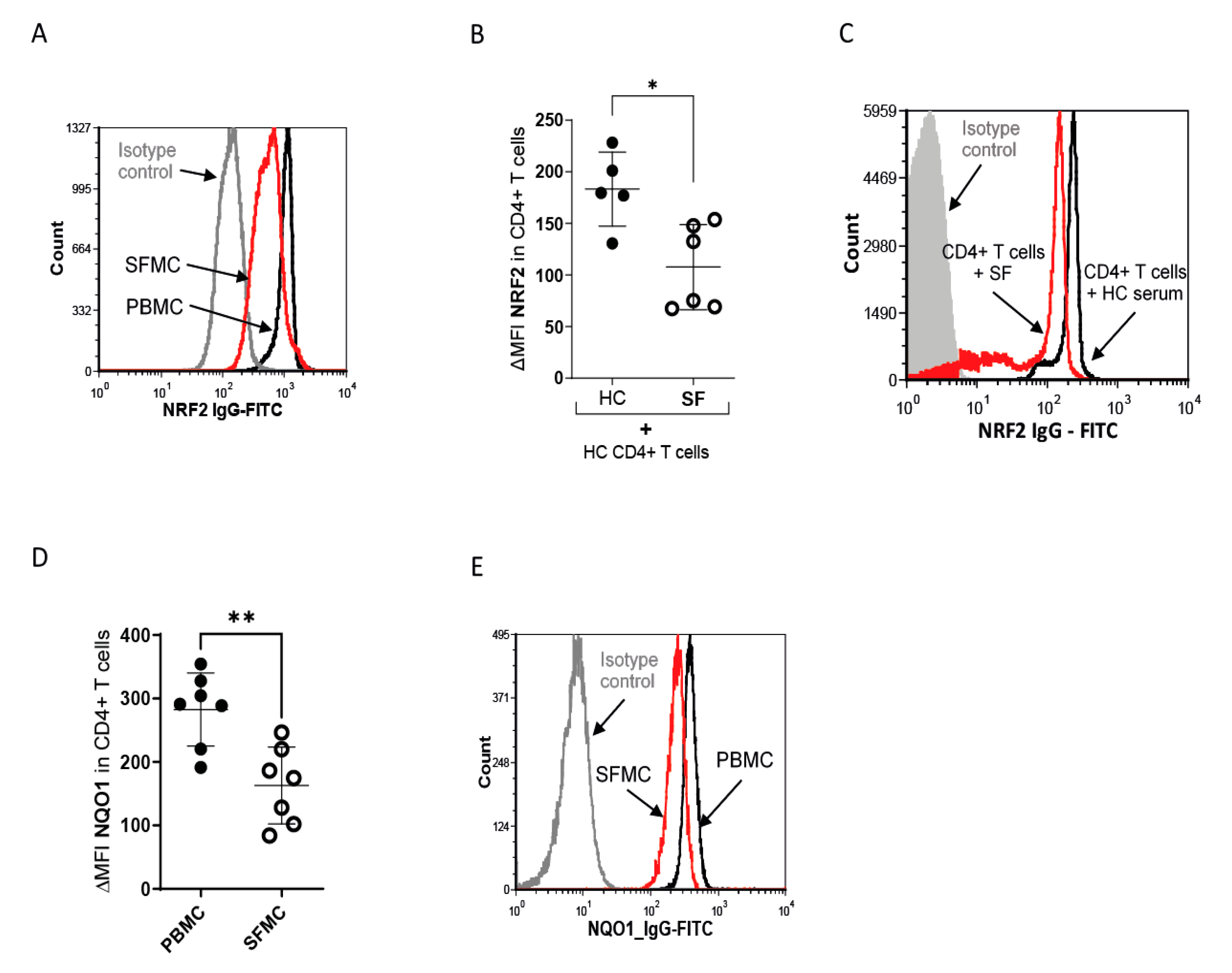
3.2. Nrf2 Signaling Is Reduced in SF T Cells
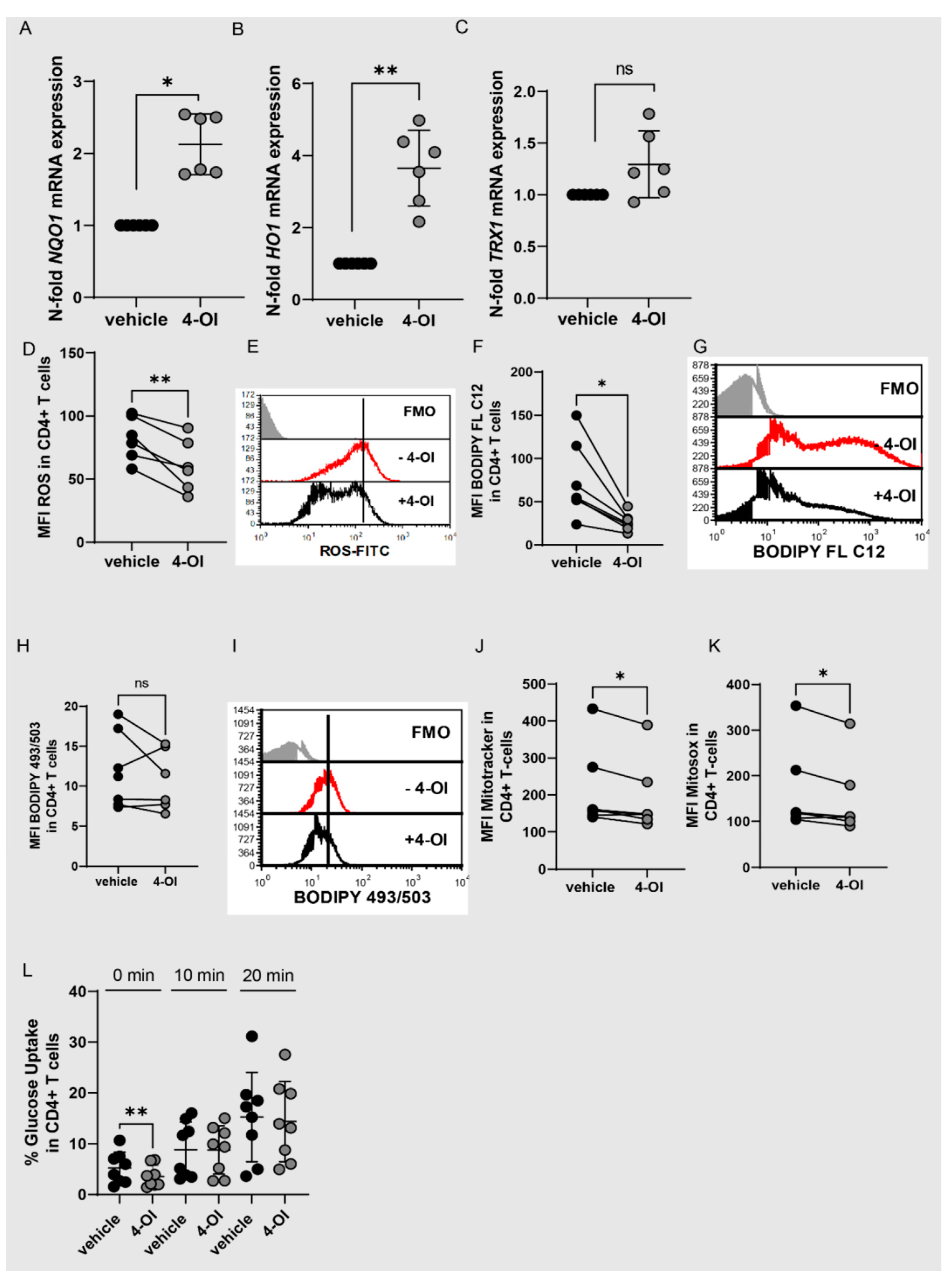
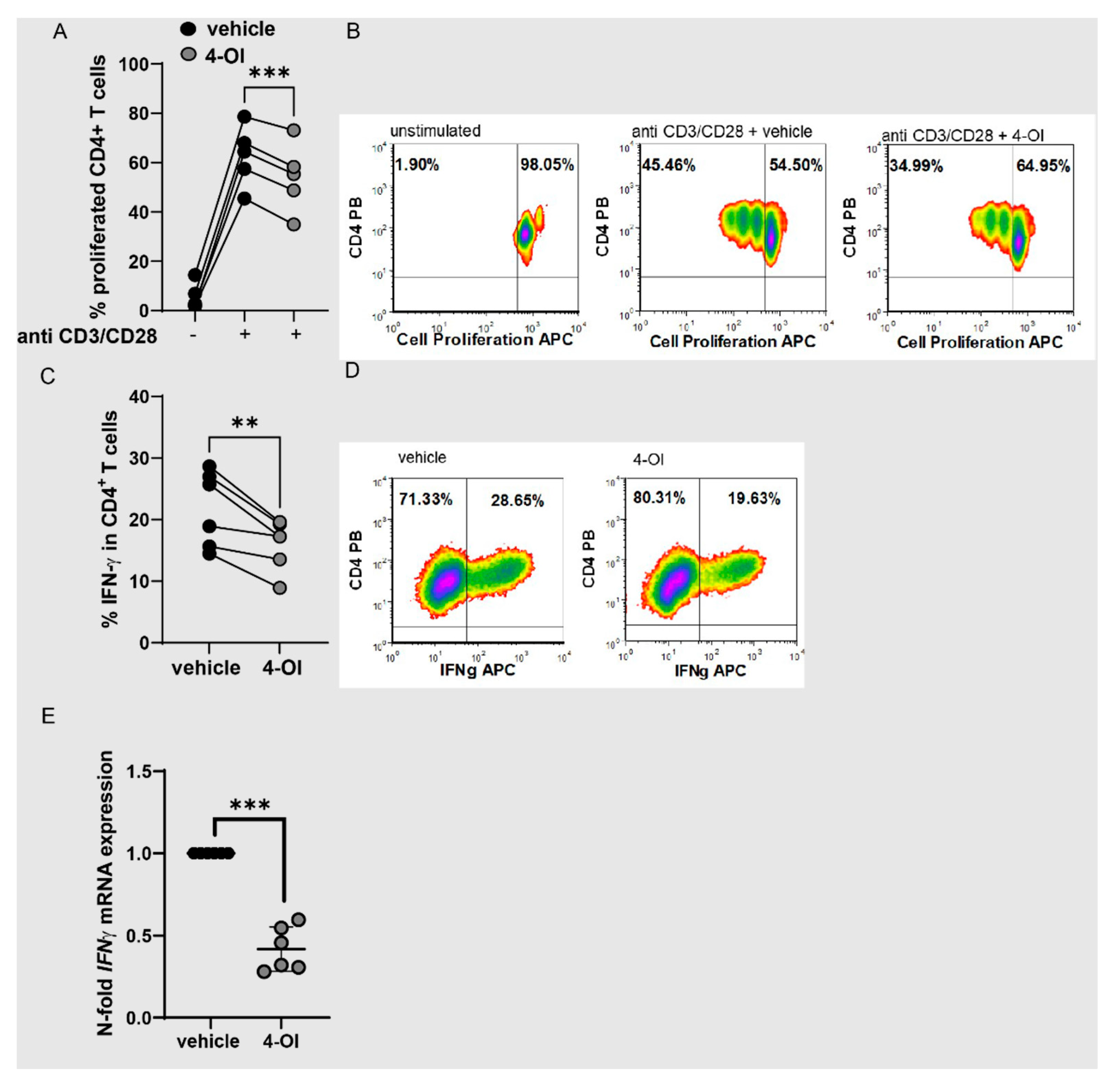
3.3. Alteration of Redox Metabolism in T Cells by 4-OI Upregulation of NRF2
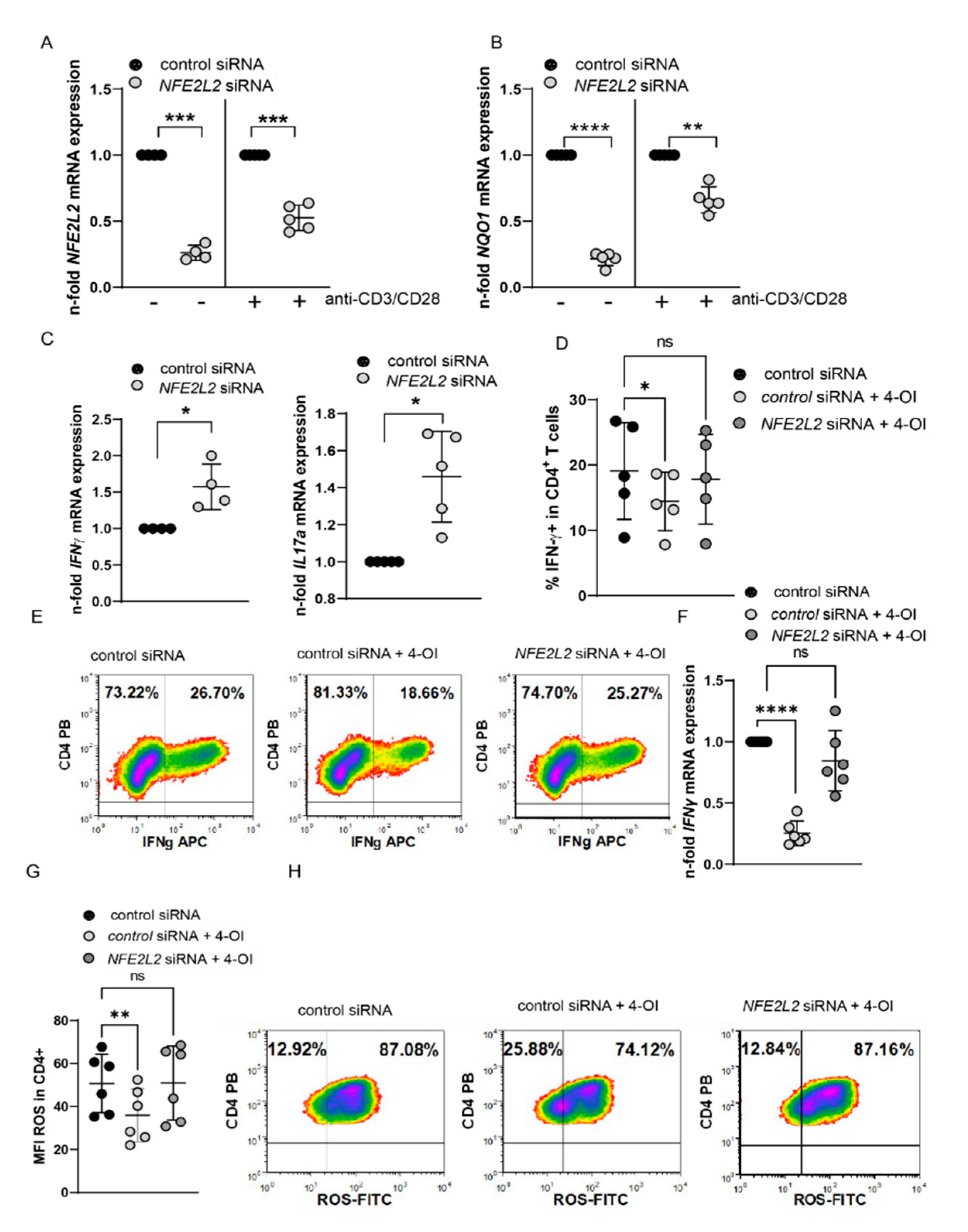
3.4. 4-OI Affects T Cell Function in a Nrf2-Dependent Manner
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurniawan, H.; Soriano-Baguet, L.; Brenner, D. Regulatory T cell metabolism at the intersection between autoimmune diseases and cancer. Eur. J. Immunol. 2020, 50, 1626–1642. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Sharabi, A.; Tsokos, G.C. T cell metabolism: New insights in systemic lupus erythematosus pathogenesis and therapy. Nat. Rev. Rheumatol. 2020, 16, 100–112. [Google Scholar] [CrossRef]
- Yang, Z.; Shen, Y.; Oishi, H.; Matteson, E.L.; Tian, L.; Goronzy, J.J.; Weyand, C.M. Restoring oxidant signaling suppresses proarthritogenic T cell effector functions in rheumatoid arthritis. Sci. Transl. Med. 2016, 8, 331ra38. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wen, Z.; Li, Y.; Matteson, E.L.; Hong, J.; Goronzy, J.J.; Weyand, C.M. Metabolic control of the scaffold protein TKS5 in tissue-invasive, proinflammatory T cells. Nat. Immunol. 2017, 18, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Weyand, C.M.; Goronzy, J.J. Immunometabolism in the development of rheumatoid arthritis. Immunol. Rev. 2020, 294, 177–187. [Google Scholar] [CrossRef]
- Zaslona, Z.; O’Neill, L.A.J. Cytokine-like Roles for Metabolites in Immunity. Mol. Cell 2020, 78, 814–823. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef]
- Ryan, D.G.; Murphy, M.P.; Frezza, C.; Prag, H.A.; Chouchani, E.T.; O’Neill, L.A.; Mills, E.L. Coupling Krebs cycle metabolites to signalling in immunity and cancer. Nat. Metab. 2019, 1, 16–33. [Google Scholar] [CrossRef]
- Bailis, W.; Shyer, J.A.; Zhao, J.; Canaveras, J.C.G.; Al Khazal, F.J.; Qu, R.; Steach, H.R.; Bielecki, P.; Khan, O.; Jackson, R.; et al. Distinct modes of mitochondrial metabolism uncouple T cell differentiation and function. Nature 2019, 571, 403–407. [Google Scholar] [CrossRef]
- Shyer, J.A.; Flavell, R.A.; Bailis, W. Metabolic signaling in T cells. Cell Res. 2020, 30, 649–659. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J.; Artyomov, M.N. Itaconate: The poster child of metabolic reprogramming in macrophage function. Nat. Rev. Immunol. 2019, 19, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Cordes, T.; Wallace, M.; Michelucci, A.; Divakaruni, A.S.; Sapcariu, S.C.; Sousa, C.; Koseki, H.; Cabrales, P.; Murphy, A.N.; Hiller, K.; et al. Immunoresponsive Gene 1 and Itaconate Inhibit Succinate Dehydrogenase to Modulate Intracellular Succinate Levels. J. Biol. Chem. 2016, 291, 14274–14284. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulou, V.; Sergushichev, A.; Bambouskova, M.; Nair, S.; Vincent, E.E.; Loginicheva, E.; Cervantes-Barragan, L.; Ma, X.; Huang, S.C.-C.; Griss, T.; et al. Itaconate Links Inhibition of Succinate Dehydrogenase with Macrophage Metabolic Remodeling and Regulation of Inflammation. Cell Metab. 2016, 24, 158–166. [Google Scholar] [CrossRef]
- Mills, E.L.; Ryan, D.G.; Prag, H.A.; Dikovskaya, D.; Menon, D.; Zaslona, Z.; Jedrychowski, M.P.; Costa, A.S.H.; Higgins, M.; Hams, E.; et al. Itaconate is an anti-inflammatory metabolite that activates Nrf2 via alkylation of KEAP1. Nature 2018, 556, 113–117. [Google Scholar] [CrossRef]
- Morzadec, C.; Macoch, M.; Sparfel, L.; Kerdine-Römer, S.; Fardel, O.; Vernhet, L. Nrf2 expression and activity in human T lymphocytes: Stimulation by T cell receptor activation and priming by inorganic arsenic and tert-butylhydroquinone. Free Radic. Biol. Med. 2014, 71, 133–145. [Google Scholar] [CrossRef]
- Klemm, P.; Rajendiran, A.; Fragoulis, A.; Wruck, C.; Schippers, A.; Wagner, N.; Bopp, T.; Tenbrock, K.; Ohl, K. Nrf2 expression driven by Foxp3 specific deletion of Keap1 results in loss of immune tolerance in mice. Eur. J. Immunol. 2020, 50, 515–524. [Google Scholar] [CrossRef]
- Tsai, J.J.; Velardi, E.; Shono, Y.; Argyropoulos, K.V.; Holland, A.M.; Smith, O.M.; Yim, N.L.; Rao, U.K.; Kreines, F.M.; Lieberman, S.R.; et al. Nrf2 regulates CD4(+) T cell-induced acute graft-versus-host disease in mice. Blood 2018, 132, 2763–2774. [Google Scholar] [CrossRef]
- Noel, S.; Martina, M.N.; Bandapalle, S.; Racusen, L.C.; Potteti, H.R.; Hamad, A.R.; Reddy, S.P.; Rabb, H. T Lymphocyte-Specific Activation of Nrf2 Protects from AKI. J. Am. Soc. Nephrol. 2015, 26, 2989–3000. [Google Scholar] [CrossRef]
- Karl, F.; Stoll, A.; Böttcher-Loschinski, R.; Böttcher, M.; Baur, R.; Jacobs, B.; Völkl, S.; Jitschin, R.; Rösler, W.; Mackensen, A.; et al. Impact of Nrf2 expression in reconstituting T-cells of allogeneic hematopoietic stem cell transplanted patients. Leukemia 2020. [Google Scholar] [CrossRef]
- Mougiakakos, D.; Johansson, C.C.; Kiessling, R. Naturally occurring regulatory T cells show reduced sensitivity toward oxidative stress-induced cell death. Blood 2009, 113, 3542–3545. [Google Scholar] [CrossRef]
- Maj, T.; Wang, W.; Crespo, J.; Zhang, H.; Wang, W.; Wei, S.; Zhao, L.; Vatan, L.; Shao, I.; Szeliga, W.; et al. Oxidative stress controls regulatory T cell apoptosis and suppressor activity and PD-L1-blockade resistance in tumor. Nat. Immunol. 2017, 18, 1332–1341. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, A.; Gambini, C.; Gerloni, V.; Parafioriti, A.; Sormani, M.P.; De Marco, G.; Rossi, F.; Martini, A.; Gattorno, M. Lymphoid neogenesis in juvenile idiopathic arthritis correlates with ANA positivity and plasma cells infiltration. Rheumatology 2007, 46, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Cosmi, L.; Cimaz, R.; Maggi, L.; Santarlasci, V.; Capone, M.; Borriello, F.; Frosali, F.; Querci, V.; Simonini, G.; Barra, G.; et al. Evidence of the transient nature of the Th17 phenotype of CD4+CD161+ T cells in the synovial fluid of patients with juvenile idiopathic arthritis. Arthritis Rheum. 2011, 63, 2504–2515. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.J.; Luyrink, L.; Grom, A.A.; Passo, M.H.; Emery, H.; Witte, D.; Glass, D.N. Immunohistological characteristics of T cell infiltrates in different forms of childhood onset chronic arthritis. J. Rheumatol. 1996, 23, 2116–2124. [Google Scholar]
- Buck, M.D.; O’Sullivan, D.; Pearce, E.L. T cell metabolism drives immunity. J. Exp. Med. 2015, 212, 1345–1360. [Google Scholar] [CrossRef]
- Ciurtin, C.; Cojocaru, V.M.; Miron, I.M.; Preda, F.; Milicescu, M.; Bojincă, M.; Costan, O.; Nicolescu, A.; Deleanu, C.; Kovàcs, E.; et al. Correlation between different components of synovial fluid and pathogenesis of rheumatic diseases. Rom. J. Intern. Med. 2006, 44, 171–181. [Google Scholar]
- Sido, B.; Seel, C.; Hochlehnert, A.; Breitkreutz, R.; Dröge, W. Low intestinal glutamine level and low glutaminase activity in Crohn’s disease: A rational for glutamine supplementation? Dig. Dis. Sci. 2006, 51, 2170–2179. [Google Scholar] [CrossRef]
- Bole, G.G. Synovial fluid lipids in normal individuals and patients with rheumatoid arthritis. Arthritis Rheum. 1962, 5, 589–601. [Google Scholar] [CrossRef]
- Leimer, E.M.; Pappan, K.L.; Nettles, D.L.; Bell, R.D.; Easley, M.E.; Olson, S.A.; Setton, L.A.; Adams, S.B. Lipid profile of human synovial fluid following intra-articular ankle fracture. J. Orthop. Res. 2017, 35, 657–666. [Google Scholar] [CrossRef]
- Kork, F.; Jankowski, J.; Goswami, A.; Weis, J.; Brook, G.; Yamoah, A.; Anink, J.; Aronica, E.; Fritz, S.; Huck, C.; et al. Golgin A4 in CSF and granulovacuolar degenerations of patients with Alzheimer disease. Neurology 2018, 91, e1799–e1808. [Google Scholar] [CrossRef] [PubMed]
- Peeters, J.G.; Vervoort, S.J.; Tan, S.C.; Mijnheer, G.; de Roock, S.; Vastert, S.J.; Nieuwenhuis, E.E.; van Wijk, F.; Prakken, B.J.; Creyghton, M.P.; et al. Inhibition of Super-Enhancer Activity in Autoinflammatory Site-Derived T Cells Reduces Disease-Associated Gene Expression. Cell Rep. 2015, 12, 1986–1996. [Google Scholar] [CrossRef] [PubMed]
- Black, A.P.; Bhayani, H.; Ryder, C.A.; Gardner-Medwin, J.M.; Southwood, T.R. T-cell activation without proliferation in juvenile idiopathic arthritis. Arthritis Res. 2002, 4, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Spreafico, R.; Rossetti, M.; Van Loosdregt, J.; Wallace, C.A.; Massa, M.; Manzoni, S.M.; Gattorno, M.; Martini, A.; Lovell, D.J.; Albani, S. A circulating reservoir of pathogenic-like CD4+ T cells shares a genetic and phenotypic signature with the inflamed synovial micro-environment. Ann. Rheum. Dis. 2016, 75, 459–465. [Google Scholar] [CrossRef]
- Ohl, K.; Nickel, H.; Moncrieffe, H.; Klemm, P.; Scheufen, A.; Föll, D.; Wixler, V.; Schippers, A.; Wagner, N.; Wedderburn, L.R.; et al. The transcription factor CREM drives an inflammatory phenotype of T cells in oligoarticular juvenile idiopathic arthritis. Pediatr. Rheumatol. Online J. 2018, 16, 39. [Google Scholar] [CrossRef] [PubMed]
- Omoyinmi, E.; Hamaoui, R.; Pesenacker, A.; Nistala, K.; Moncrieffe, H.; Ursu, S.; Wedderburn, L.; Woo, P. Th1 and Th17 cell subpopulations are enriched in the peripheral blood of patients with systemic juvenile idiopathic arthritis. Rheumatology 2012, 51, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Rajendiran, A.; Klemm, P.; Schippers, A.; Scheufen, A.; Schwarz, T.; Peitz, J.; Brandenburg, L.-O.; Wagner, N.; Consolaro, A.; Raggi, F.; et al. miR-23a contributes to T cellular redox metabolism in juvenile idiopathic oligoarthritis. Rheumatology 2022, 61, 2694–2703. [Google Scholar] [CrossRef]
- Lipinska, J.; Lipińska, S.; Stańczyk, J.; Sarniak, A.; Prymont, A.P.V.; Kasielski, M.; Smolewska, E. Reactive oxygen species and serum antioxidant defense in juvenile idiopathic arthritis. Clin. Rheumatol. 2015, 34, 451–456. [Google Scholar] [CrossRef]
- Kaur, G.; Sharma, A.; Bhatnagar, A. Role of oxidative stress in pathophysiology of rheumatoid arthritis: Insights into NRF2-KEAP1 signalling. Autoimmunity 2021, 54, 385–397. [Google Scholar] [CrossRef]
- Mutter, F.E.; Park, B.K.; Copple, I.M. Value of monitoring Nrf2 activity for the detection of chemical and oxidative stress. Biochem. Soc. Trans. 2015, 43, 657–662. [Google Scholar] [CrossRef]
- Hradilkova, K.; Maschmeyer, P.; Westendorf, K.; Schliemann, H.; Husak, O.; Von Stuckrad, A.S.L.; Kallinich, T.; Minden, K.; Durek, P.; Grün, J.R.; et al. Regulation of Fatty Acid Oxidation by Twist 1 in the Metabolic Adaptation of T Helper Lymphocytes to Chronic Inflammation. Arthritis Rheumatol. 2019, 71, 1756–1765. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Goodman, S.B.; Berry, G.J.; Goronzy, J.J.; Weyand, C.M. Metabolic Control of Autoimmunity and Tissue Inflammation in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 652771. [Google Scholar] [CrossRef] [PubMed]
- Kidani, Y.; Elsaesser, H.; Hock, M.B.; Vergnes, L.; Williams, K.J.; Argus, J.P.; Marbois, B.N.; Komisopoulou, E.; Wilson, E.B.; Osborne, T.F.; et al. Sterol regulatory element-binding proteins are essential for the metabolic programming of effector T cells and adaptive immunity. Nat. Immunol. 2013, 14, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Michelucci, A.; Cordes, T.; Ghelfi, J.; Pailot, A.; Reiling, N.; Goldmann, O.; Binz, T.; Wegner, A.; Tallam, A.; Rausell, A.; et al. Immune-responsive gene 1 protein links metabolism to immunity by catalyzing itaconic acid production. Proc. Natl. Acad. Sci. USA 2013, 110, 7820–7825. [Google Scholar] [CrossRef] [PubMed]
- Nastasi, C.; Willerlev-Olsen, A.; Dalhoff, K.; Ford, S.L.; Østergaard Gadsbøll, A.-S.; Buus, T.B.; Gluud, M.; Danielsen, M.; Litman, T.; Bonefeld, C.M.; et al. Inhibition of succinate dehydrogenase activity impairs human T cell activation and function. Sci. Rep. 2021, 11, 1458. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Yadav, J.; Makhija, S.; Mazumder, S.; Mitra, A.K.; Suryawanshi, A.; Sandey, M.; Mishra, A. Irg1/itaconate metabolic pathway is a crucial determinant of dendritic cells immune-priming function and contributes to resolute allergen-induced airway inflammation. Mucosal Immunol. 2022, 15, 301–313. [Google Scholar] [CrossRef]
- Roberts, L.M.; Leighton, I.; Schwarz, B.; Wehrly, T.D.; Evans, T.J.; Bosio, C.M. Itaconate indirectly influences expansion of effector T cells following vaccination with Francisella tularensis live vaccine strain. Cell Immunol. 2022, 373, 104485. [Google Scholar] [CrossRef]
- Tang, C.; Wang, X.; Xie, Y.; Cai, X.; Yu, N.; Hu, Y.; Zheng, Z. 4-Octyl Itaconate Activates Nrf2 Signaling to Inhibit Pro-Inflammatory Cytokine Production in Peripheral Blood Mononuclear Cells of Systemic Lupus Erythematosus Patients. Cell Physiol. Biochem. 2018, 51, 979–990. [Google Scholar] [CrossRef]
- Peace, C.G.; O’Neill, L.A. The role of itaconate in host defense and inflammation. J. Clin. Invest. 2022, 132, e148548. [Google Scholar] [CrossRef]
- Gold, R.; Giovanni, G.; Phillips, J.T.; Fox, R.J.; Zhang, A.; Marantz, J.L. Sustained Effect of Delayed-Release Dimethyl Fumarate in Newly Diagnosed Patients with Relapsing-Remitting Multiple Sclerosis: 6-Year Interim Results From an Extension of the DEFINE and CONFIRM Studies. Neurol. Ther. 2016, 5, 45–57. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gender | Age (Years) | Treatment |
|---|---|---|
| Female | 4 | none |
| Female | 15 | none |
| Male | 11 | none |
| Female | 7 | none |
| Male | 14 | none |
| Female | 3 | none |
| Female | 11 | none |
| Male | 4 | none |
| Male | 14 | none |
| Female | 8 | Methotrexate |
| Female | 13 | none |
| Male | 16 | Methotrexate & Decortin |
| Female | 10 | none |
| Male | 6 | none |
| Male | 12 | none |
| Female | 6 | Ibuprofen |
| Female | 12 | none |
| Female | 6 | none |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajendiran, A.; Subramanyam, S.H.; Klemm, P.; Jankowski, V.; van Loosdregt, J.; Vastert, B.; Vollbach, K.; Wagner, N.; Tenbrock, K.; Ohl, K. NRF2/Itaconate Axis Regulates Metabolism and Inflammatory Properties of T Cells in Children with JIA. Antioxidants 2022, 11, 2426. https://doi.org/10.3390/antiox11122426
Rajendiran A, Subramanyam SH, Klemm P, Jankowski V, van Loosdregt J, Vastert B, Vollbach K, Wagner N, Tenbrock K, Ohl K. NRF2/Itaconate Axis Regulates Metabolism and Inflammatory Properties of T Cells in Children with JIA. Antioxidants. 2022; 11(12):2426. https://doi.org/10.3390/antiox11122426
Chicago/Turabian StyleRajendiran, Anandhi, Sudheendra Hebbar Subramanyam, Patricia Klemm, Vera Jankowski, Jorg van Loosdregt, Bas Vastert, Kristina Vollbach, Norbert Wagner, Klaus Tenbrock, and Kim Ohl. 2022. "NRF2/Itaconate Axis Regulates Metabolism and Inflammatory Properties of T Cells in Children with JIA" Antioxidants 11, no. 12: 2426. https://doi.org/10.3390/antiox11122426
APA StyleRajendiran, A., Subramanyam, S. H., Klemm, P., Jankowski, V., van Loosdregt, J., Vastert, B., Vollbach, K., Wagner, N., Tenbrock, K., & Ohl, K. (2022). NRF2/Itaconate Axis Regulates Metabolism and Inflammatory Properties of T Cells in Children with JIA. Antioxidants, 11(12), 2426. https://doi.org/10.3390/antiox11122426