Abstract
Hydroxycinnamoyl-quinic acids (HCQAs) are polyphenol esters formed of hydroxycinnamic acids and (-)-quinic acid. They are naturally synthesized by plants and some micro-organisms. The ester of caffeic acid and quinic acid, the chlorogenic acid, is an intermediate of lignin biosynthesis. HCQAs are biologically active dietary compounds exhibiting several important therapeutic properties, including antioxidant, antimicrobial, anti-inflammatory, neuroprotective, and other activities. They can also be used in the synthesis of nanoparticles or drugs. However, extraction of these compounds from biomass is a complex process and their synthesis requires costly precursors, limiting the industrial production and availability of a wider variety of HCQAs. The recently emerged production through the bioconversion is still in an early stage of development. In this paper, we discuss existing and potential future strategies for production of HCQAs.
1. Introduction
HCQAs (Table 1) are mostly produced in plants by ester formation of a hydroxycinnamic acid (primarily p-coumaric, caffeic, ferulic and sinapic acids) with a (-)-quinic acid from the phenylpropanoid pathway, and they are linked with lignin synthesis [1]. They belong to a large and diverse group of phenolic compounds, often termed as chlorogenic acids [2]. The extended list of chlorogenic acids, which contains approximately 400 compounds [2], also encompass several derivatives and isomers of quinic acid, including shikimic acid, its epimers, 4-deoxy-, muco-, methyl- and butyl-quinic acids esterified with hydroxycinnamic and hydroxybenzoic acids or some of their derivatives. The nomenclature and trivial names of compounds of chlorogenic acids are rather complicated and they are explained in [3].

Table 1.
The principal chemical structure of HCQAs.
Here, we primarily focus on the HCQAs (Table 1). Amongst these, the most abundant and important compound is 5-caffeoylquinic acid (5-CQA), often referred to as chlorogenic acid. In 2018, the market size of 5-CQA was 130 million and it is expected to reach 150 million US$ by 2025 [4].
Over the last decade, a wide interest in HCQAs has been reflected by an exponentially growing number of scientific publications (Figure 1). However, the main focus has been on HCQAs extraction from plants and to some extent on their chemical synthesis, whereas studies on microbial production of these acids have been limited.
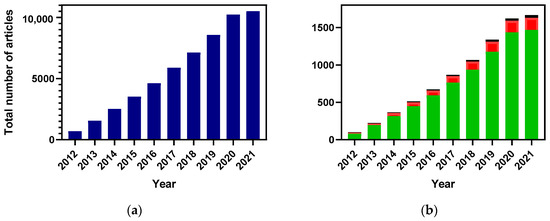
Figure 1.
The number of publications dedicated to HCQAs research over the last 10 years (data based on information retrieved from scopus.com on 22 February 2022). Number of publications is represented as following: total ((a) blue bar), HCQAs extraction from plants ((b) green bar), HCQAs chemical synthesis ((b) red bar), and HCQAs production in micro-organisms ((b) black bar).
HCQAs are used in pharmaceuticals, cosmetics, foods due to their therapeutic properties, such as antioxidant [5], anticancer [6], antimicrobial [7,8], antiobesity [9], hepatoprotective [10], antiviral and anti-inflammatory [11], antihypertensive [12] and neuroprotective [13]. The hydroxycinnamoyl moiety in these compounds determines the antimicrobial activity [14,15]. The strength of this property increases with the number of these moieties in the molecule. HCQAs exhibit antimicrobial activity against many bacteria, including Enterococcus faecium, Escherichia coli and others [16]. However, some cider yeasts (Lactobacillus collinoides, Lactobacillus paracollinoides) [17,18], fungi (Aspergillus niger C23308, Fusarium graminearum [19,20], Fusarium culmorum and Fusarium graminearum sensu stricto [21]) and lactic acid producing bacteria (Lactobacillus johnsonii NCC 533) [17,22] are resistant to 5-CQA due to their ability to catabolize this compound.
5-CQA has been found to be useful for diverse applications. Recently, it has been shown to have potential to counteract SARS-CoV-2 by reducing viral attachment to the host cell-surface heat shock protein A5 (HSPA5) [23]. 5-CQA stimulates short-chain fatty acid production in bacteria. The fermentation products of this acid stimulate the proliferation of Bifidobacterium spp. causing the decreased ratio between Firmicutes and Bacteroidetes [24]. The final compounds formed from 5-CQA and other HCQAs are hippuric acid and 3-hydroxyhippuric acid, which are used as non-specific biomarkers for polyphenol uptake or metabolism [25,26,27].
Moreover, 5-CQA has found applications in the synthesis of metals’ nanoparticles as a reducing and stabilizing agent [28,29]; in the production modification of carbon ceramic electrode for NADH detection [30]; in red food dye preparation from coupling of tryptophan and 5-CQA [31]; and the preservation of food by preparing the edible coating with chitosan [32]. This compound is also referred to as the major compound from instant coffee extract responsible for graphene green production from graphite and functionalization [33]. 5-CQA is considered as the precursor of caffeic and quinic acids, which may be obtained via hydrolysis reactions during extraction from plant material [34].
Importantly, the derivatives of HCQAs are also widely researched for their potential use in the production of drugs or the use in biopharmaceuticals [35,36]. Moreover, the extract of green coffee beans containing a large amount of 5-CQA has been certified by COSMOS to obtain a label of a natural raw material for cosmetics [37]. It is also produced by Naturex under the trade name Svetol® as a supplement for body weight loss [38].
This paper aims to discuss and assess the recent advances in chemical synthesis of HCQAs, their extraction from plant material and agricultural waste as well as emerging bioproduction of these compounds using natural or modified micro-organisms. The HCQAs production strategies and relevant research developments are summarized.
2. HCQAs Extraction from Plants and Agricultural Waste
2.1. HCQAs Biosynthesis in Plants
The phenylpropanoid pathway forms a platform for HCQAs biosynthesis in plants with the main biochemical reactions presented in Figure 2. HCQAs can be synthesized from p-coumaric acid or p-coumaroyl-CoA by four different routes. The first route (blue arrows in Figure 2) requires the direct conversion of p-coumaric acid into other hydroxycinnamic acids (caffeic, ferulic, sinapic acids) via hydroxylation or methylation reactions mediated by p-coumarate 3-hydroxylase (C3H), caffeic/5-hydroxyferulic acid O-methyltransferase (COMT) or ferulic acid 5-hydroxylase (F5H) [39]. Then, hydroxycinnamic acids CoA esters are formed with mediation of 4-coumaroyl-CoA ligase (4CL) [40] and HCQAs are produced by transesterification reaction with quinic acid catalyzed by HCT/HQT [41]. The second route (pink arrows in Figure 3) is the conversion of hydroxycinnamates into glucosides and transesterification with quinic acid [41]. The third and fourth routes starts from p-coumaroyl-CoA, which is converted into p-coumaroyl-quinic (quinate shunt, red arrows in Figure 2), or p-coumaroyl-shikimic (shikimate shunt, green arrows in Figure 2) acids with additional conversion of obtained shikimate into ester of CoA and following esterification with quinic acid, respectively [39,41,42]. 3-CQA, 4-CQA are produced from 5-CQA, and 3,4-diCQA or 4,5-diCQAs are synthesized from 1,5-diCQA with mediation of isomerase or hydroxycinnamoyltransferase, respectively [43,44]. The dual activity of HQT enzyme results in 5-HCQAs production in cytosol and in the production of 3,4-diCQA, 3,5-diCQA or 4,5-diCQAs from two molecules of 5-CQA (acting as acyl-donor and acyl-acceptor) in the vacuoles [45]. In addition, 3,5-diCQA can be obtained from 5-CQA and caffeoyl-CoA with mediation of HCT and ICS enzymes [46]. Cis isomerization may occur with mediation of the non-specified/non-identified enzyme [47] or due to the exposure to ultraviolet irradiation [48]. To date, the major synthesis routes in plants are shikimic or quinic acids shunts, which results in the production of p-CoQAs, CQAs or FQAs [39,44,49,50].
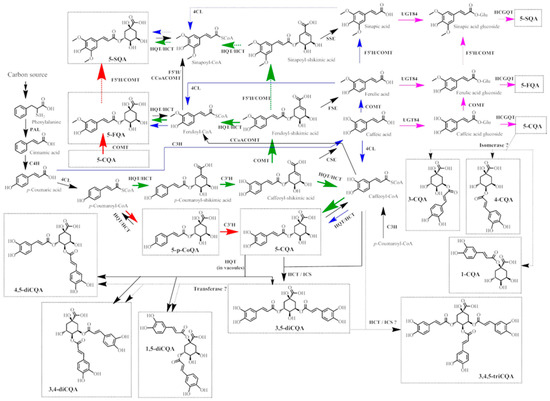
Figure 2.
Pathways for the biosynthesis of HCQAs in plants. The four main routes of phenylpropanoid metabolism are highlighted in different colors: green, shikimate shunt; red, quinate shunt; blue and pink, direct conversion and cinnamoyl glucosides pathway. Dashed arrows show the suggested enzymatic reactions. Abbreviations: PAL, L-phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, p-coumaroyl-CoA ligase; HCT/HQT, 4-hydroxycinnamoyl CoA - shikimate/quinate hydroxycinnamoyl transferase; C3’H, p-coumaroyl shikimate/quinate 3’-hydroxylase, CSE - caffeoyl shikimate esterase, ICS isochlorogenate synthase, HCT - hydroxycinnamoyl-CoA shikimate/quinate hydroxycinnamoyltransferase; HQT, hydroxycinnamoyl-CoA quinate hydroxycinnamoyl transferase; CSE, caffeoyl shikimate esterase; FSE, feruoyl shikimate esterase; SSE, sinapoyl shikimate esterase; COMT, caffeic/5-hydroxyferulic acid O-methyltransferase; C3H, p-coumarate 3-hydroxylase (ascorbate peroxidase); CCoAOMT, caffeoyl-CoA 3-O-methyltransferase; UGT84, UDP-glucoside transferase; HCGQT, hydroxycinnamoyl D-glucose:quinate hydroxycinnamoyl transferase; F5’H—ferulic acid 5-hydroxylase.

Figure 3.
Some natural sources of HCQAs: tea tree (Camellia sinensis) (a), rosemary (Rosmarinus officinalis) (b), mountain arnica (Arnica montana) (c), coffee beans (Coffea sp.) (d).
2.1.1. Plant Sources of HCQAs
All types of HCQAs (from Table 2) may be found in plants (Figure 3) and they are mainly localized in fruit skin, seeds, kernels, leaves, or husks. The richest sources of HCQAs (mainly mono- and dicaffeoylquinic acids) are Yerba mate, white, green teas and coffee (Table 2). Although coffee is considered a major 5-CQA source, depending on the coffee type it may contain a lower total amount of 5-CQA but a larger variety of other HCQAs (including sinapoyl- derivatives such as 3-SQA, 4-SQA, 5-SQA, 3-S-5-CQA, 3-S-4-CQA, 4-S-3-CQA, 3-S-5-FQA, 3-F-4-SQA, 4-S-5-FQA) than teas [51,52,53]. The plants from Asteraceae, Cichorium, Phaseoullus, Brassica, Solanaceae and Lamiaceae families possess high amounts of different HCQAs [54,55,56]. Many fruits and berries are rich in CQAs with the highest total concentrations of 200–570 mg/kg wet biomass determined in cherry, quince, mulberry, bilberry, and sweet granadilla [57]. Food, crop and agro-industrial waste can be used as an alternative source for the bio-refinery of HCQAs [58,59]. Coffee by-products are some of the richest waste sources containing up to 10–23% of HCQAs [51,60]. Principally, all plants containing high levels of alkaloids possess significant amounts of HCQAs [61,62].
Table 2.
Plant sources with the most abundant HCQAs available.
2.1.2. Marine Sources of HCQAs
HCQAs are present in marine sources, mainly microalgae (Spongiochlori sp., Euglena cantabrica, Anabaena doliolum, Porphyra tenera, Undaria pinnatifia) and cyanobacteria (Nostoc sp.) [64,65,66,67]. The HCQAs synthesis pathway in these organisms is similar to higher plants [68]. Although microalgae and cyanobacteria contain high levels of extractable phenolic compounds (phenolic acids) and quinic acid, the abundance of various HCQAs is limited. To date, the data on abundance of 5-CQA is only available for some species of marine plants and cyanobacteria. Its concentration reaches up to 78 µg/g DW for microalgae and 9.55 µg/g DW for cyanobacteria (Table 3). The higher amount of 5-CQA is determined in algae due to the adaptation to abiotic and biotic stress occurring in the evolutionary advanced micro-organisms [65].
Table 3.
Marine sources of 5-CQA.
2.2. HCQAs Extraction
HCQAs are usually extracted by conventional, solid–liquid extraction (SLE) and nonconventional or intensified techniques, such as ultrasound assisted (UAE), microwave assisted (MAE), pressurized liquid (PLE), supercritical fluid (SFE), enzymatic extraction (EAE) [69,70] or infrared assisted extraction (IAE) [71] (Table 4). Only in EAE, the release and extraction is performed for HCQAs trapped within the plant cell walls using enzymes (cellulases, glucosidases, proteases, dextranases, xylanases and ligninolytic enzymes that do not hydrolase the HCQAs into their constituents) in aqueous buffers or ionic liquids [72,73,74,75]. The non-conventional or intensified extraction methods are considered to be more efficient. In extraction process, parameters (such as temperature, time, pH of solvents, particle size, solvent type and concentration, its volume and other specific parameters of the process (e.g., microwave power, ultrasonic frequency, enzyme concentration)) can be optimized [69,76]. The final yield of HCQAs depends on the raw material as well as the extraction method (Table 4). The highest yields (up to 4% of raw material) of HCQAs are determined for the coffee by-products, honeysuckle (Lonicera japonicae) and its flowers or by-products, sunflower seed kernels [60,77,78,79,80,81,82]. All the extraction methods are potentially suitable for the HCQAs except for SFE, which is considered as the green extraction technique, but the recovery yields do not exceed~52%, due to reduced solubility in the nonpolar supercritical fluid [80,83]. The MAE and IRE are usually very efficient and require a short time for the extraction process, but the large size microwave extractors in industry remain too expensive. Therefore, SLE is considered the most relevant method for industrial application due to its simplicity, reproducibility, low cost and possibility to use environment friendly solvents [84]. Intensified methods are mostly applied for the recovery of HCQAs from waste [71,85,86,87,88,89,90,91,92,93] (Table 4).
Table 4.
Yields HCQAs extracted from plants and wastes.
Many intensified extraction methods have been adopted for waste valorization. Furthermore, combined processes, such as microwave-assisted simultaneous distillation and dual extraction (MSDDE) [90], multi-frequency multimode modulated vibration (acoustic probe) technique (MMM) [91] or simultaneous SLE extraction and solid-state fermentation (SLE-SSF) [92] have been developed for 5-CQA extraction (Table 4).These three methods are more effective than SLE or other non-conventional extraction methods, however, MSDDE and MMM are too expensive for industrial application, and only SLE-SSF can be applied for large scale extraction and production of 5-CQA with high enrichment of raw material (for example, up to 400% for coffee pulp) [92].
For the separation, concentration and purification of HCQAs, mesoporous resins (recovery 65.03%, purity of CQAs 89.27%) [94], a high-speed countercurrent or high-performance liquid chromatography (recovery up to 99.56%) [82,89,95], fractional extraction in centrifuges (yield 35%, purity 99.5%) [96], isocratic system with three zones of simulating moving beds (0.04–0.2 g/L extract, purity 99.27%) [77], ultrafiltration membranes (rejection 92%) [97], molecularly imprinted polymer (recovery 60.08–72.59%, extraction yield 12.57 mg/g raw material) [79], imprinted magnetic nanomaterials or membranes (recovery 86–102%) [98,99], ethanol/salt aqueous two-phase system (95.76% maximum extraction) [100] are used. The chromatography-based purification is the most popular method, which is often used in industrial applications, due to its simplicity, effectiveness and reasonable cost. The fractional extraction in centrifuge or three-zone simulated moving bed method are based on continuous and selective processes, which allows for separating and concentrating 5-CQA with high purity (97.25–99.27%) [77,96]. However, only the fractional extraction in centrifuge is a cost-effective method and can be applied in industrial scale production [96]. The other separation and purification methods have been tested only in lab-scale production and may be limited due to some disadvantages, such as high cost, low effectiveness, low purity or selectivity, long analysis time, usage of toxic (organic) solvents, or membrane fouling [79,94,98,100,101,102,103].
The extraction and identification of HCQAs suffer from some limitations. Firstly, the stability of HCQAs is reduced by the isomerization or destruction reactions under extraction conditions, especially if these conditions are harsh [2,93]. Secondly, transesterification reactions can occur due to the activity of chlorogenate-dependent caffeoyltransferase, when the extraction of fresh plant material is performed in alcohol or alcohol-water mixtures [104]. The transformed products may be mistakenly identified as the new bioactive ingredients of the plant sources [105]. Another problem is the identification of the compounds in the extracts due to the limited number of commercially available standards and similar properties or spectra of HCQAs as summarized in [2,106].
Extraction and purification processes can be more environment-friendly if ionic liquids or eco-friendly natural deep eutectic solvents are used instead of organic solvents. They have a higher affinity to HCQAs than water [107], and it is easy to purify these solvents by distillation and to re-use them for extraction processes [108,109]. Furthermore, the use of ionic liquids can result in a shorter extraction time [75,110]. Generally, the recycling and re-use of the extraction solvents, application of high solid–to–liquid ratio, and reduced processing time helps to lower the total cost of the extraction [100,111].
The extraction of HCQAs results in large amounts of the residual plant material. Aiming for the sustainable circular economy and resource conservation, these residues can be utilized for combustion [112], as a biofuel (e.g., biodiesel, bioethanol, biochar and liquid pyrolysis product production) [113,114,115] or other applications [60,113,116,117,118,119,120,121,122,123] (Figure 4), which allows for reducing the solid waste amount and/or covering up to 100% of phytoremediation cost [124,125].
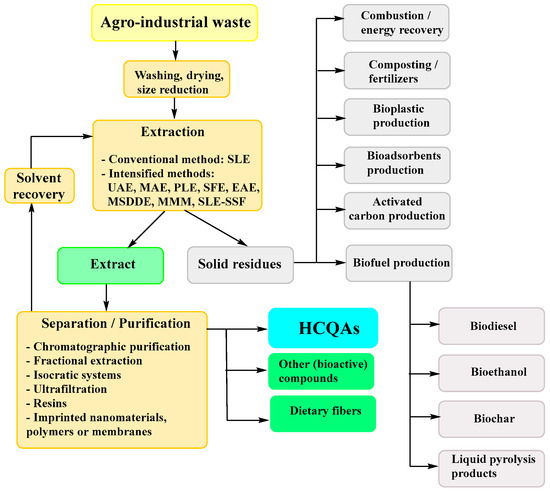
Figure 4.
General agro-industrial waste treatment possibilities. Abbreviations: EAE—enzyme assisted extraction; MAE—microwave assisted extraction; MMM—multi-frequency multimode modulated vibration (acoustic probe) technique; MSDDE—microwave-assisted simultaneous distillation and dual extraction; PLE—pressurized liquid extraction; SFE—supercritical fluid extraction; SLE—solid-liquid extraction techniques; SLE-SSF—simultaneous SLE extraction and solid-state fermentation; UAE—ultrasound assisted extraction.
3. Chemical Synthesis of HCQAs
The major HCQAs containing hydroxycinnamoyl moieties can be chemically synthesized performing either esterification or condensation reaction between quinic and hydroxycinnamic acids using pyridine or DMAP as homogenous catalyst in organic solvents (dichlormethane or DMF). The reactions with the best total yields are presented in Figure 5.
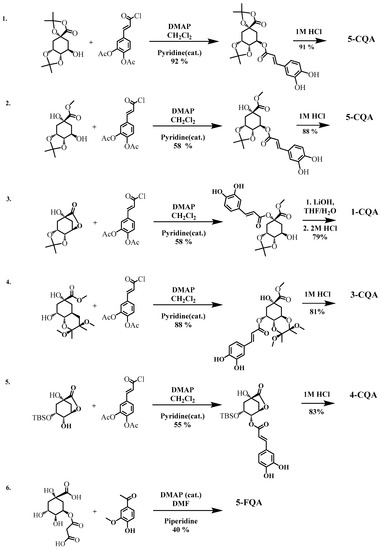
Figure 5.
Chemical synthesis of 1-CQA, 3-CQA, 3-CQA and 5-CQA. Selected reactions with highest yields reported by [127] (1); [130] (2); [126] (3–5); and [129] (6). All reactions performed at room temperature.
The highest total yields have been obtained with Sefkow synthesis method for 1-CQA, 3-CQA, 4-CQA or 5-CQA isomers (Table 5) using esterification reaction between caffeoylchloride derivative and quinic acid derivatives with unprotected OH groups at 1, 3, 4 and/or 5 positions followed by 1–2 steps of hydrolysis reaction of protecting groups [126,127]. The non-protected quinic acid can also be used in the esterification with suitably protected hydroxycinnamoyl derivative, which results in the formation of all mono- substituted HCQAs with moieties at 1-, 3-, 4- and 5- positions [128]. The synthesis of 5-FQA can be obtained from quinic acid ester with malonate and vanillin without any protected hydroxyl group via Knoevenagel condensation reaction when the (E)-double bond is formed (the total yield of 19%) [129]. Independently of the synthesis method, the final crystalline HCQAs can be purified by recrystallization or by a complex procedure (extraction, concentration and chromatographic purification followed by recrystallization) for the improved purity of the final compound [126,127,129]. All the above methods are commonly used in manufacturing, and they are considered cost-effective as the solvents can be recovered by distillation and reused.
Table 5.
The highest yields of other HCQAs obtained by synthetic chemistry methods.
There are positive and negative aspects of condensation and esterification methods. Higher total yields can be achieved with the esterification method compared to the condensation method. Irrespective of method, the synthesis of di- or tri-HCQAs is more complex than that of mono-HCQAs. Esterification reaction requires the protection/deprotection of active hydroxyl groups because the HCQAs molecules are sensitive to basic and strong acidic or hydrogenative conditions due to the double bond reduction, transesterification or isomerization reactions as well as possible cleavage reactions [129,131,132]. Additionally, esterification may require low or ultralow temperature, solvent system CH2Cl2/pyridine/DMAP ratio optimization, and long acidic hydrolysis time for the deprotection reaction [126,127,131,132,136], which could cause the decreased yields of HCQAs due to the side reactions. In contrast, the condensation reaction by Knoevenagel involves benzyl aldehydes use of which does not require any protection of reactive hydroxyl groups [137]. 1-HCQAs and di- or tri- substituted HCQAs are more difficult to obtain by both condensation and esterification methods due to increased steric hindrance.
The prices of the major chemical substances for both synthesis methods are presented in Table 6. The Knoevenagel condensation reaction requires benzyl aldehydes, which are currently commercially produced from naphtha. Hydroxycinnamic acids for the esterification reaction are expensive as they are produced mainly by chemical synthesis. Even biotechnological production of vanillin, syringaldehyde or extraction of hydroxycinnamic acids from plant biomass could not reduce the high synthesis costs of HCQAs.
Table 6.
The average prices for the chemical substances and catalysts required for the HCQAs synthesis (Prices are listed from [138].
4. Biosynthesis of HCQAs in Non-Modified and Modified Micro-Organisms
4.1. Non-Modified Micro-Organisms
Non-modified micro-organisms, such as bacteria and fungi are able to produce 5-CQA from organic carbon (Table 7). The detectable amounts of 5-CQA have been observed with bacteria such as Brevibacillus borstelensis, Bacillus amyloliquefaciens, Bacillus badius, Sphingomonas yabuuchiae, Enterobacter tabaci, Paenibacillus phoenicis and fungi including Colletotrichum acutatum, Lodderomyces elongisporus, Sphingomonas yabuuchiae, Enterobacter tabaci, Paenibacillus phoenicis [139], Sordariomycetes sp. [140], Penicillium flavigenum, Screlotium rolfsii [141,142]. L. elongisporus sp. S216 and P. flavigenum (CML2965) exhibit the 5-CQA titers that could be of interest for industrial scale production [139,141] (Table 7). The pathway of 5-CQA biosynthesis in micro-organisms is considered to be similar to plants [139]. Interestingly, an ortho-adipate pathway enabling production of 1-CQA and 3,4,5-triCQA may be functional in Streptomyces albogriseolus KF977548, which has been isolated from decaying wood [143].
Table 7.
Comparison of HCQAs production cases in non-modified micro-organisms.
Micro-organisms producing 5-CQA can be used for the valorization of agricultural waste. For example, B. amyloliquefaciens B17 has been applied for the successful fermentation of mango peels in liquid state fermentation at 37 °C [145]. Further research is required to screen and develop micro-organisms for the valorization of different types of waste.
4.2. Modified Micro-Organisms
The HCQAs biosynthesis pathways have been developed in E. coli, Saccharomyces cerevisiae, and Pichia pastoris. For the production of 5-CQA from caffeic acid, the E. coli has been engineered by introducing hydroxycinnamoyl-CoA quinate transferase gene HQT from Nicotiana tabacum and 4-coumarate CoA:ligase gene 4CL, which mediates the formation of coenzyme A thioester with hydroxycinnamic acids and deletion of aroD gene, which is required for the conversion of 3-dehydroquinate into 3-dehydroshikimate [145]. This recombinant strain B-101 was able to accumulate 3-dehydroquinate and caffeoyl-3-dehydroquinate, but it was not able to produce 5-CQA in higher concentrations than 16 mg/L. When gene ydiB encoding shikimate/quinate dehydrogenase was overexpressed followed by the vector and cell concentration optimization, then the production up to 450 mg/L in 24 h was achieved from quinate and additionally supplied caffeic acid [145].
Recently, the synthesis of 5-CQA and p-coumaroyl shikimate by expressing shikimate gene modules in E. coli has been demonstrated. The overexpression of five genes of the shikimate pathway (ppsA, tktA, aroGf, aroB, ydiB) and heterologous genes TAL, HpaBC, 4CL, HST, and HQT resulted in the biosynthesis of 109.7 mg/L 5-CQA from glucose [146] (Figure 6).
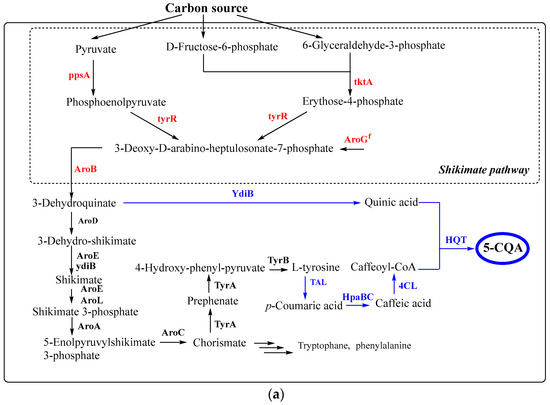
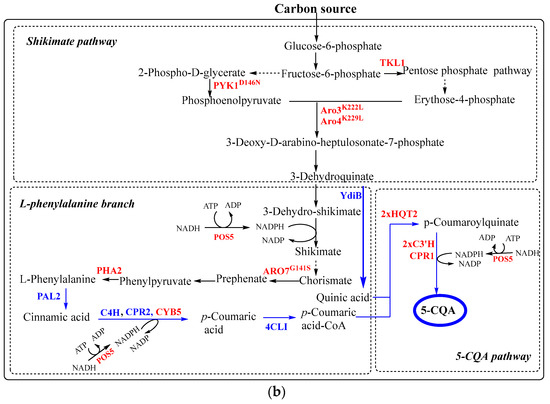
Figure 6.
The synthesis of chlorogenic acid in E. coli (a) and S. cerevisiae (b) from carbon source according to [146; 151]. Black and blue arrows correspond to native and non-native pathways, respectively. Dashed arrows represent the complex processes. The blue and red names of genes correspond to inserted and overexpressed genes, respectively. Abbreviations: AroH - phospho-2-dehydro-3-deoxyheptonate aldolase; TyrR—transcriptional regulatory protein; AroF - phospho-2-dehydro-3-deoxyheptonate aldolase; AroG—phospho-2-dehydro-3-deoxyheptonate aldolase; AroD—5-dehydroquinate dehydratase; AroB—dehydroquinate synthase; PAL2—phenylalanine ammonia lyase from Arabidopsis thaliana; C3′H—cytochrome P450 98A3 from A. thaliana; CPR1 and AtCPR2—P450 reductases from A. thaliana; YdiB—quinate/shikimate dehydrogenase from E. coli; AtC4H, cinnamate-4-hydroxylase from A. thaliana; 4CL—4-coumarateCoA:ligase from Oryza sativa; 4CL1, 4-coumarate:CoA ligase 1 from A. thaliana; HQT—hydroxycinnamoyl-CoA quinate transferase from Nicotiana tabacum; HQT2—hydroxycinnamoyl-CoA quinate transferase 2 from Cynara scolymus; ARO3K222L—l-phenylalanine feedback-insensitive DAHP synthase; ARO4K229L—l-tyrosine feedback-insensitive DAHP synthase; ARO7G141S—l-tyrosine feedback-insensitive chorismate mutase; PYK1D146N—pyruvate kinase 1 mutant with reduced catalytic activity.
The polyculture technique has been applied for 5-CQA production using engineered E. coli. In the two culture technique, E. coli B-102 was inoculated in a filtrated culture mixture of modified E. coli strain B-TP-CA2, which produced caffeic acid from glucose [147]. After 45 h the highest concentration (78 mg/L) of 5-CQA was obtained. Similarly, de novo 5-CQA production using the polyculture technique with three recombinant E. coli strains containing biosynthetic modules of caffeic acid, quinic acid and 5-CQA, reached 250 µM (or ~88 mg/L) concentration after 18 h of incubation [148]. In both cases, a rather low yield of targeted compound could be acceptable due to the usage of a low-cost primary compound (glucose).
Yeast has also been engineered for the synthesis of HCQAs. The expression of tobacco 4CL and globe artichoke HCT genes in yeast Saccharomyces cerevisiae resulted in the formation of N-(E)-p-coumaroyl-3-hydroxyanthranilic acid as a primary product, which is similar to avenanthramides [149]. Subsequently, a successful biosynthesis of 5-CQA and 5-pCoQA in yeast was achieved by expressing a BAHD enzyme NtHQT from tobacco Nicotiana tabacum and 4CL5 [150]. Recently, it has been shown that GDSL lipase-like ICS enzyme from Ipomoea batatas can be used for the efficient conversion of 5-CQA into 3,5-diCQA in P. pastoris [46].
Further improvements have been achieved by introducing into S. cerevisiae the de novo 5-CQA synthesis pathway, including PAL2, C4H, 4CL1, C3′H, CPR1, CPR2, HQT2, YdiB, CYB5 and implementing the following modifications (Figure 6): (1) unlocking the shikimate pathway and optimizing carbon distribution by overexpressing the L-phenylalanine feedback-insensitive DAHP synthase (ARO3K222L), L-tyrosine feedback-insensitive DAHP synthase (ARO4K229L), pyruvate kinase 1 mutant with reduced catalytic activity (PYK1D146N) and transketolase (TKL1); (2) optimizing the L-phenylalanine branch and pathway balancing by overexpressing the l-tyrosine feedback-insensitive chorismate mutase (ARO7G141S), endogenous prephenate dehydratase (PHA2), NADH kinase (POS5), and cytochrome b5 (Cyb5); (3) increasing the copy number of 5-CQA pathway genes encoding hydroxycinnamoyl-CoA quinate transferase 2 (HQT2) and cytochrome P450 98A3 (C3′H) [151]. The engineered S. cerevisiae strain produced 5-CQA to 234.8 ± 11.1 mg/L in shake flask culture and 806.8 ± 1.7 mg/L in fed-batch fermentation [151].
5. Conclusions
HCQAs are becoming very important compounds in cosmetics, medicine and food supplements production due to their outstanding properties. Therefore, the need of HCQAs, especially 5-CQA, is constantly increasing. The main strategies to obtain these compounds are based on their extraction from plant biomass and chemical synthesis. Recently, biosynthesis using modified or non-modified micro-organisms has attracted significant research efforts.
Several conventional and intensified methods have been developed for extraction of HCQAs from plants or marine biomass. Amongst the most promising are solid–liquid extraction techniques (SLE), pressurized liquid (PLE), supercritical fluid (SFE), enzymatic extraction (EAE), even if SLE remains dominant. The successful application of other intensified methods, such as microwave-assisted simultaneous distillation and dual extraction (MSDDE), multi-frequency multimode modulated vibration (acoustic probe) technique (MMM) or simultaneous SLE extraction and solid-state fermentation (SLE-SSF), was demonstrated with some evident success for the 5-CQA. Their suitability for other HCQAs remains to be explored. Despite that, intensified methods can result in better yields and can require less time; however, they are expensive, which limits their industrial application. 5-CQA extraction has already reached the stage of industrialization. 5-CQA is mainly produced (>75%) from honeysuckle, eucommia and green coffee bean. For the less abundant HCQAs, such as SQAs (which is present at extremely low yields in plants), further improvements of the extraction methodology are required.
Chemical synthesis enables achievement of moderate yields of mono-, di- or tri- HCQAs. It requires large quantities of hydroxycinnamic acid and is based on use of condensation or esterification methods. Esterification enables achievement of a better yield, but the high costs of reagents limits use of this method. Overall, the chemical synthesis of HCQAs is expensive and environmentally unfriendly because the primary compounds of the synthesis are obtained mainly from naphtha and its refinery products. Moreover, the use of halogenized organic compounds generates toxic waste. Despite this, this method could be preferable for the production of naturally scarce SQAs or to synthesize HCQAs that are not obtainable by other methods.
Biotechnological production of HCQAs is considered a most promising approach that enables consolidation of green chemistry and circular economy objectives. Despite its early stage, several non-modified micro-organisms have already been shown to produce the CQAs to yields that are comparable to those obtained by extraction from plant biomass. Significantly, the engineered bacteria Escherichia coli or yeast Saccharomyces cerevisiae have been developed to synthesize a greater variety of HCQAs, such as 5-CQA, 5-FQA, 5-p-CoQA from simple carbon sources. The 5-CQA titer of approximately 0.8 g/l has been achieved in fed-batch fermentation using Saccharomyces cerevisiae. Although the biotechnological production of HCQAs requires a significant advancement to make it suitable for industrial application, the utilization of micro-organisms shows great promise in the recycling and recovery of HCQAs from organic waste.
Author Contributions
Conceptualization, E.V. and N.M.; original draft preparation, E.V.; review and editing, E.V. and N.M.; funding acquisition and project administration, N.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Regional Development Fund (project no. 01.2.2-LMT-K-718-02-0023) under grant agreement with the Research Council of Lithuania (LMTLT).
Acknowledgments
The authors are indebted to the Research Council of Lithuania (LMTLT). This research was funded by the European Regional Development Fund (project no. 01.2.2-LMT-K-718-02-0023) under grant agreement with the LMTLT.
Conflicts of Interest
The authors declare no conflict of interest. The funding source had no role in the study’s design; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Silva, N.; Mazzafera, P.; Cesarino, I. Should I stay or should I go: Are chlorogenic acids mobilized towards lignin biosynthesis? Phytochemistry 2019, 166, 112063. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.; Jaganath, I.; Ludwig, I.; Crozier, A. Chlorogenic acids and the acyl-quinic acids: Discovery, biosynthesis, bioavailability and bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef] [PubMed]
- Abrankó, L.; Clifford, M. An Unambiguous Nomenclature for the Acyl-Quinic Acids Commonly Known as Chlorogenic Acids. J. Agric. Food Chem. 2017, 65, 3602–3608. [Google Scholar] [CrossRef] [PubMed]
- DataIntelo. Global Chlorogenic Acid Market Report, History and Forecast 2014–2025, Breakdown Data by Manufacturers, Key Regions, Types and Application. Available online: https://dataintelo.com/report/chlorogenic-acid-market (accessed on 15 March 2022).
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Gouthamchandra, K.; Sudeep, H.; Venkatesh, B.; Shyam Prasad, K. Chlorogenic acid complex (CGA7), standardized extract from green coffee beans exerts anticancer effects against cultured human colon cancer HCT-116 cells. Food Sci. Hum. Wellness 2017, 6, 147–153. [Google Scholar] [CrossRef]
- Cao, X.; Islam, M.N.; Duan, Z.; Pan, X.; Xu, W.; Wei, X.; Zhong, S. Chlorogenic acid osmosis of snakehead fish: A novel approach to maintain quality and suppress deterioration during storage. Int. J. Food Prop. 2020, 23, 387–399. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Cho, A.-S.; Jeon, S.-M.; Kim, M.-J.; Yeo, J.; Seo, K.-I.; Choi, M.-S.; Lee, M.-K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef]
- Ali, F.; Hassan, N.; Abdrabou, R. Hepatoprotective and antiproliferative activity of moringinine, chlorogenic acid and quercetin. Int. J. Med. Sci. 2016, 4, 1147–1153. [Google Scholar] [CrossRef]
- Li, Y.; Yang, D.; Jia, Y.; He, L.; Li, J.; Yu, C.; Liao, C.; Yu, Z.; Zhang, C. Research Note: Anti-inflammatory effects and antiviral activities of baicalein and chlorogenic acid against infectious bursal disease virus in embryonic eggs. Poult. Sci. 2021, 100, 100987. [Google Scholar] [CrossRef]
- Hakkou, Z.; Maciuk, A.; Leblais, V.; Bouanani, N.; Mekhfi, H.; Bnouham, M.; Aziz, M.; Ziyyat, A.; Rauf, A.; Hadda, T.B.; et al. Antihypertensive and vasodilator effects of methanolic extract of Inula viscosa: Biological evaluation and POM analysis of cynarin, chlorogenic acid as potential hypertensive. Biomed. Pharmacother. 2017, 93, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Heitman, E.; Ingram, D. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Narita, Y.; Inouye, K. Inhibitory effects of chlorogenic acids from green coffee beans and cinnamate derivatives on the activity of porcine pancreas α-amylase isozyme I. Food Chem. 2011, 127, 1532–1539. [Google Scholar] [CrossRef]
- Narita, Y.; Inouye, K. Kinetic analysis and mechanism on the inhibition of chlorogenic acid and its components against porcine pancreas α-amylase isozymes I and II. J. Agric. Food Chem. 2009, 57, 9218–9225. [Google Scholar] [CrossRef] [PubMed]
- Bajko, E.; Kalinowska, M.; Borowski, P.; Siergiejczyk, L.; Lewandowski, W. 5-O-Caffeoylquinic acid: A spectroscopic study and biological screening for antimicrobial activity. LWT Food Sci. Technol. 2016, 65, 471–479. [Google Scholar] [CrossRef]
- Buron, N.; Coton, M.; Desmarais, C.; Ledauphin, J.; Guichard, H.; Barillier, D.; Coton, E. Screening of representative cider yeasts and bacteria for volatile phenol-production ability. Food Microbiol. 2011, 28, 1243–1251. [Google Scholar] [CrossRef]
- Whiting, G.C.; Carr, J.G. Chlorogenic Acid Metabolism in Cider Fermentation. Nature 1957, 180, 1479. [Google Scholar] [CrossRef]
- Torres-Mancera, M.T.; Baqueiro-Peña, I.; Figueroa-Montero, A.; Rodríguez-Serrano, G.; González-Zamora, E.; Favela-Torres, E.; Saucedo-Castañeda, G. Biotransformation and improved enzymatic extraction of chlorogenic acid from coffee pulp by filamentous fungi. Biotechnol. Prog. 2013, 29, 337–345. [Google Scholar] [CrossRef]
- Gauthier, L.; Bonnin-Verdal, M.-N.; Marchegay, G.; Pinson-Gadais, L.; Ducos, C.; Richard-Forget, F.; Atanasova-Penichon, V. Fungal biotransformation of chlorogenic and caffeic acids by Fusarium graminearum: New insights in the contribution of phenolic acids to resistance to deoxynivalenol accumulation in cereals. Int. J. Food Microbiol. 2016, 221, 61–68. [Google Scholar] [CrossRef]
- Kulik, T.; Stuper-Szablewska, K.; Bilska, K.; Buśko, M.; Ostrowska-Kołodziejczak, A.; Załuski, D.; Perkowski, J. Trans-Cinnamic and Chlorogenic Acids Affect the Secondary Metabolic Profiles and Ergosterol Biosynthesis by Fusarium culmorum and F. graminearum Sensu Stricto. Toxins 2017, 9, 198. [Google Scholar] [CrossRef]
- Bel-Rhlid, R.; Thapa, D.; Kraehenbuehl, K.; Fischer, L. Biotransformation of caffeoyl quinic acids from green coffee extracts by Lactobacillus johnsonii NCC 533. AMB Express 2013, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, A.A. Natural products may interfere with SARS-CoV-2 attachment to the host cell. J. Biomol. Struct. Dyn. 2021, 39, 3194–3203. [Google Scholar] [CrossRef] [PubMed]
- Parkar, S.G.; Trower, T.M.; Stevenson, D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe 2013, 23, 12–19. [Google Scholar] [CrossRef]
- Clifford, M.N.; Copeland, E.L.; Bloxsidge, J.P.; Mitchell, L.A. Hippuric acid as a major excretion product associated with black tea consumption. Xenobiotica 2000, 30, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Spencer, J.P.; Kuhnle, G.; Hahn, U.; Rice-Evans, C.A. Novel biomarkers of the metabolism of caffeic acid derivatives in vivo. Free Radic. Biol. Med. 2001, 30, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Gonthier, M.-P.; Verny, M.-A.; Besson, K.; Rémésy, C.; Scalbert, A. Chlorogenic Acid Bioavailability Largely Depends on Its Metabolism by the Gut Microflora in Rats. J. Nutr. 2003, 133, 1853–1859. [Google Scholar] [CrossRef]
- Hwang, S.J.; Jun, S.H.; Park, Y.; Cha, S.-H.; Yoon, M.; Cho, S.; Lee, H.-J.; Park, Y. Green synthesis of gold nanoparticles using chlorogenic acid and their enhanced performance for inflammation. Nanotechnol. Biol. Med. 2015, 11, 1677–1688. [Google Scholar] [CrossRef]
- Chen, R.; Wu, S.; Meng, C. Size-tunable green synthesis of platinum nanoparticles using chlorogenic acid. Res. Chem. Intermed. 2021, 47, 1775–1787. [Google Scholar] [CrossRef]
- Salimi, S.; Hallaj, R.; Ghadermazi, M. Modification of carbon ceramic electrode prepared with sol–gel technique by a thin film of chlorogenic acid: Application to amperometric detection of NADH. Talanta 2005, 65, 888–894. [Google Scholar] [CrossRef]
- Moccia, F.; Martín, M.; Ramos, S.; Goya, L.; Marzorati, S.; DellaGreca, M.; Panzella, L.; Napolitano, A. A new cyanine from oxidative coupling of chlorogenic acid with tryptophan: Assessment of the potential as red dye for food coloring. Food Chem. 2021, 348, 129152. [Google Scholar] [CrossRef]
- Jiao, W.; Shu, C.; Li, X.; Cao, J.; Fan, X.; Jiang, W. Preparation of a chitosan-chlorogenic acid conjugate and its application as edible coating in postharvest preservation of peach fruit. Postharvest Biol. Technol. 2019, 154, 129–136. [Google Scholar] [CrossRef]
- Abdullah, A.; Ismail, Z.; Zainal Abidin, A.; Yusoh, K. Green sonochemical synthesis of few-layer graphene in instant coffee. Mater. Chem. Phys. 2019, 222, 11–19. [Google Scholar] [CrossRef]
- Flourat, A.; Combes, J.; Bailly-Maitre-Grand, C.; Magnien, K.; Haudrechy, A.; Renault, J.-H.; Allais, F. Accessing p-Hydroxycinnamic Acids: Chemical Synthesis, Biomass Recovery, or Engineered Microbial Production? ChemSusChem 2021, 14, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Can, T.H.; Tufekci, E.F.; Altunoglu, Y.C.; Baloglu, M.C.; Llorent-Martínez, E.J.; Stefanucci, A.; Mollica, A.; Cichelli, A.; Zengin, G. Chemical characterization, computational analysis and biological views on Daphne gnidioides Jaub. Spach extracts: Can a new raw material be provided for biopharmaceutical applications? Comput. Biol. Chem. 2020, 87, 107273. [Google Scholar]
- Zhang, X.; Liu, D.-Y.; Shang, H.; Jia, Y.; Xu, X.-D.; Tian, Y.; Guo, P. Amino acid ester-coupled caffeoylquinic acid derivatives as potential hypolipidemic agents: Synthesis and biological evaluation. RSC Adv. 2021, 11, 1654–1661. [Google Scholar] [CrossRef]
- COSMOS. 27 April 2021. Available online: http://www.cosmos-standard-rm.org/verifmp.php (accessed on 15 September 2021).
- Svetol. 27 April 2021. Available online: http://www.svetol.com/what-is-svetol/ (accessed on 16 September 2021).
- Valiñas, M.A.; Lanteri, M.L.; ten Have, A.; Andreu, A.B. Chlorogenic Acid Biosynthesis Appears Linked with Suberin Production in Potato Tuber (Solanum tuberosum). J. Agric. Food Chem. 2015, 63, 4902–4913. [Google Scholar] [CrossRef]
- Lee, D.; Douglas, C.J. Two Divergent Members of a Tobacco 4-Coumarate:Coenzyme A Ligase (4CL) Gene Family (cDNA Structure, Gene Inheritance and Expression, and Properties of Recombinant Proteins). Plant Physiol. 1996, 112, 193–205. [Google Scholar] [CrossRef]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef]
- Mahesh, V.; Million-Rousseau, R.; Ullmann, P.; Chabrillange, N.; Bustamante, J.; Mondolot, L.; Morant, M.; Noirot, M.; Hamon, S.; De Kochko, A.; et al. Functional characterization of two p-coumaroyl ester 3′-hydroxylase genes from coffee tree: Evidence of a candidate for chlorogenic acid biosynthesis. Plant Mol. Biol. 2007, 64, 145–159. [Google Scholar] [CrossRef]
- Joet, T.; Salmona, J.; Laffargue, A.; Descroix, F.; Dussert, S. Use of the growing environment as a source of variation to identify the quantitative trait transcripts and modules of co-expressed genes that determine chlorogenic acid accumulation. Plant Cell Environ. 2010, 33, 1220–1233. [Google Scholar] [CrossRef]
- LLallemand, L.A.; Zubieta, C.; Lee, S.G.; Wang, Y.; Acajjaoui, S.; Timmins, J.; McSweeney, S.; Jez, J.M.; McCarthy, J.G.; McCarthy, A.A. A Structural Basis for the Biosynthesis of the Major Chlorogenic Acids Found in Coffee. Plant Physiol. 2012, 160, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Moglia, A.; Lanteri, S.; Comino, C.; Hill, L.; Knevitt, D.; Cagliero, C.; Rubiolo, P.; Bornemann, S.; Martin, C. Dual Catalytic Activity of Hydroxycinnamoyl-Coenzyme A Quinate Transferase from Tomato Allows It to Moonlight in the Synthesis of Both Mono- and Dicaffeoylquinic Acids. Plant Physiol. 2014, 166, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Sissi, M.; Guillaume, L.; Léonor, D.; Delporte, M.; Menin, B.; Michel, C.; Alexandre, O.; Gabrielle, C.; Aleksander, S.; Joakim, B.; et al. A GDSL lipase-like from Ipomoea batatas catalyzes efficient production of 3,5-diCQA when expressed in Pichia pastoris. Commun. Biol. 2020, 3, 673. [Google Scholar]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Madala, N.E.; Dubery, I.A. Metabolomic fingerprinting of primed tobacco cells provide the first evidence for the biological origin of cis-chlorogenic acid. Biotechnol. Lett. 2015, 37, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Karaköse, H.; Jaiswal, R.; Deshpande, S.; Kuhnert, N. Investigation of the Photochemical Changes of Chlorogenic Acids Induced by Ultraviolet Light in Model Systems and in Agricultural Practice with Stevia rebaudiana Cultivation as an Example. J. Agric. Food Chem. 2015, 63, 3338–3347. [Google Scholar] [CrossRef]
- Hoffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purification, Cloning, and Properties of an Acyltransferase Controlling Shikimate and Quinate Ester Intermediates in Phenylpropanoid Metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, D.; Liu, J.; Yu, X.; Wang, R.; Wei, Y.; Wen, C.; Ouyang, Z. Transcriptomic analysis of key genes involved in chlorogenic acid biosynthetic pathway and characterization of MaHCT from Morus alba L. Protein Expr. Purif. 2019, 156, 25–35. [Google Scholar] [CrossRef]
- Farah, A.; Donangelo, C. Phenolic compounds in coffee. Braz. J. Plant Physiol. 2006, 18, 23–36. [Google Scholar] [CrossRef]
- Gutiérrez Ortiz, A.; Berti, F.; Solano Sánchez, W.; Navarini, L.; Colomban, S.; Crisafulli, P.; Forzato, C. Distribution of p-coumaroylquinic acids in commercial Coffea spp. of different geographical origin and in other wild coffee species. Food Chem. 2019, 286, 459–466. [Google Scholar] [CrossRef]
- Jaiswal, R.; Patras, M.; Eravuchira, P.; Kuhnert, N. Profile and Characterization of the Chlorogenic Acids in Green Robusta Coffee Beans by LC-MSn: Identification of Seven New Classes of Compounds. J. Agric. Food Chem. 2010, 58, 8722–8737. [Google Scholar] [CrossRef]
- Meinhart, A.; Damin, F.; Caldeirão, L.; da Silveira, T.; Filho, J.T.; Godoy, H. Chlorogenic acid isomer contents in 100 plants commercialized in Brazil. Food Res. Int. 2017, 99, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Meinhart, A.D.; Damin, F.M.; Caldeirao, L.; de Jesus Filho, M.; da Silva, L.C.; da Silva Constant, L.; Teixeira Filho, J.; Wagner, R.; Godoy, H.T. Study of new sources of six chlorogenic acids and caffeic acid. J. Food Compos. Anal. 2019, 82, 103244. [Google Scholar] [CrossRef]
- Galani, J.; Mankad, P.M.; Shah, A.K.; Patel, N.J.; Acharya, R.R.; Talati, J.G. Effect of Storage Temperature on Vitamin C, Total Phenolics, UPLC Phenolic Acid Profile and Antioxidant Capacity of Eleven Potato (Solanum tuberosum) Varieties. Hortic. Plant J. 2017, 3, 73–89. [Google Scholar] [CrossRef]
- Meinhart, A.D.; Damin, F.M.; Caldeirao, L.; de Jesus Filho, M.; da Silva, L.C.; da Silva Constant, L.; Teixeira Filho, J.; Wagner, R.; Godoy, H.T. Chlorogenic and caffeic acids in 64 fruits consumed in Brazil. Food Chem. 2019, 286, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Le, P.; Vu, Q.; Nguyen, Q.; Tran, K.A.; Le, K. Spent Coffee Grounds as a Valuable Source of Bioactive Compounds and Bioenergy. Chem. Eng. Trans. 2017, 56, 37–42. [Google Scholar]
- Aires, A.; Carvalho, R.; Saavedra, M. Reuse potential of vegetable wastes (broccoli, green bean and tomato) for the recovery of antioxidant phenolic acids and flavonoids. Int. J. Food Sci. Technol. 2017, 52, 98–107. [Google Scholar] [CrossRef]
- Murthy, P.; Naidu, M. Recovery of Phenolic Antioxidants and Functional Compounds from Coffee Industry By-Products. Food Bioprocess Technol. 2012, 5, 897–903. [Google Scholar] [CrossRef]
- Waldhauser, S.; Baumann, T. Compartmentation of caffeine and related purine alkaloids depends exclusively on the physical chemistry of their vacuolar complex formation with chlorogenic acids. Phytochemistry 1996, 42, 985–996. [Google Scholar] [CrossRef]
- Pardo Torre, J.; Schmidt, G.; Paetz, C.; Reichelt, M.; Schneider, B.; Gershenzon, J.; D’Auria, J. The biosynthesis of hydroxycinnamoyl quinate esters and their role in the storage of cocaine in Erythroxylum coca. Phytochemistry 2013, 91, 177–186. [Google Scholar] [CrossRef]
- dos Santos Scholz, M.; Kitzberger, C.; Durand, N.; Rakocevic, M. From the field to coffee cup: Impact of planting design on chlorogenic acid isomers and other compounds in coffee beans and sensory attributes of coffee beverage. Eur. Food Res. Technol. 2018, 244, 1793–1802. [Google Scholar] [CrossRef]
- Klejdus, B.; Kopecký, J.; Benešová, L.; Vacek, J. Solid-phase/supercritical-fluid extraction for liquid chromatography of phenolic compounds in freshwater microalgae and selected cyanobacterial species. J. Chromatogr. A 2009, 1216, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J. Phenolic Profile and Antioxidant Activity of Crude Extracts from Microalgae and Cyanobacteria Strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef]
- Blagojević, D.; Babić, O.; Rašeta, M.; Šibul, F.; Janjušević, L.; Simeunović, J. Antioxidant activity and phenolic profile in filamentous cyanobacteria: The impact of nitrogen. J. Appl. Phycol. 2018, 30, 2337–2346. [Google Scholar] [CrossRef]
- Onofrejová, L.; Vašíčková, J.; Klejdus, B.; Stratil, P.; Mišurcová, L.; Kráčmar, S.; Kopecký, J.; Vacek, J. Bioactive phenols in algae: The application of pressurized-liquid and solid-phase extraction techniques. J. Pharm. Biomed. Anal. 2010, 51, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Boudet, A.-M. Evolution and current status of research in phenolic compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar] [CrossRef]
- Wianowska, D.; Gil, M. Recent advances in extraction and analysis procedures of natural chlorogenic acids. Phytochem. Rev. 2019, 18, 273–302. [Google Scholar] [CrossRef]
- Mena-García, A.; Rodríguez-Sánchez, S.; Ruiz-Matute, A.I.; Sanz, M.L. Exploitation of artichoke byproducts to obtain bioactive extracts enriched in inositols and caffeoylquinic acids by Microwave Assisted Extraction. J. Chromatogr. A 2020, 1613, 460703. [Google Scholar] [CrossRef]
- Rajha, H.N.; Mhanna, T.; Kantar, S.E.; Khoury, A.E.; Louka, N.; Maroun, R.G. Innovative process of polyphenol recovery from pomegranate peels by combining green deep eutectic solvents and a new infrared technology. LWT 2019, 111, 138–146. [Google Scholar] [CrossRef]
- Verduzco-Oliva, R.; Gutierrez-Uribe, J.A. Beyond Enzyme Production: Solid State Fermentation (SSF) as an Alternative Approach to Produce Antioxidant Polysaccharides. Sustainability 2020, 12, 495. [Google Scholar] [CrossRef]
- Martins, D.A.; Prado, H.F.; Leite, R.S.; Ferreira, H.; Souza Moretti, M.M.; da Silva, R.; Gomes, E. Agroindustrial Wastes as Substrates for Microbial Enzymes Production and Source of Sugar for Bioethanol Production. In Integrated Waste Management; Kumar, S., Ed.; IntechOpen: London, UK, 2011; Volume 2. [Google Scholar]
- Pinelo, M.; Zornoza, B.; Meyer, A. Selective release of phenols from apple skin: Mass transfer kinetics during solvent and enzyme-assisted extraction. Sep. Purif. Technol. 2008, 63, 620–627. [Google Scholar] [CrossRef]
- Liu, T.; Sui, X.; Li, L.; Zhang, J.; Liang, X.; Li, W.; Zhang, H.; Fu, S. Application of ionic liquids based enzyme-assisted extraction of chlorogenic acid from Eucommia ulmoides leaves. Anal. Chim. Acta 2016, 903, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Belwal, T.; Bhatt, I.D.; Rawal, R.S.; Pande, V. Microwave-assisted extraction (MAE) conditions using polynomial design for improving antioxidant phytochemicals in Berberis asiatica Roxb. ex DC. leaves. Ind. Crops Prod. 2017, 95, 393–403. [Google Scholar] [CrossRef]
- Nakkong, K.; Tangpromphan, P.; Jaree, A. The Design of Three-Zone Simulated Moving Bed Process for the Separation of Chlorogenic and Gallic Acids Extracted from Spent Coffee Grounds. Waste Biomass Valoriz. 2020, 12, 2389–2405. [Google Scholar] [CrossRef]
- Náthia-Neves, G.; Alonso, E. Valorization of sunflower by-product using microwave-assisted extraction to obtain a rich protein flour: Recovery of chlorogenic acid, phenolic content and antioxidant capacity. Food Bioprod. Process. 2021, 125, 57–67. [Google Scholar] [CrossRef]
- Li, G.; Wang, W.; Wang, Q.; Zhu, T. Deep Eutectic Solvents Modified Molecular Imprinted Polymers for Optimized Purification of Chlorogenic Acid from Honeysuckle. J. Chromatogr. Sci. 2016, 54, 271–279. [Google Scholar] [CrossRef]
- Daraee, A.; Ghoreishi, S.M.; Hedayati, A. Supercritical CO2 extraction of chlorogenic acid from sunflower (Helianthus annuus) seed kernels: Modeling and optimization by response surface methodology. J. Supercrit. Fluids. 2019, 144, 19–27. [Google Scholar] [CrossRef]
- Sun, Y.; Ding, S.; Huang, H.; Hu, Y. Ionic liquid-based enzyme-assisted extraction of chlorogenic acid from Flos Lonicera Japonicae. Bioresour. Bioprocess. 2017, 4, 45. [Google Scholar] [CrossRef]
- Ge, L.; Wan, H.; Tang, S.; Chen, H.; Li, J.; Zhang, K.; Zhou, B.; Fei, J.; Wu, S.; Zeng, X. Novel caffeoylquinic acid derivatives from Lonicera japonica Thunb. flower buds exert pronounced anti-HBV activities. RSC Adv. 2018, 8, 35374–35385. [Google Scholar] [CrossRef]
- de Souza, A.; Stefanov, S.; Bombardelli, M.; Corazza, M.; Stateva, R. Assessment of composition and biological activity of Arctium lappa leaves extracts obtained with pressurized liquid and supercritical CO2 extraction. J. Supercrit. Fluids. 2019, 152, 104573. [Google Scholar] [CrossRef]
- Granados-Vallejo, M.; Arriola-Guevara, E.; Corona-González, R.I.; Flores-Méndez, D.A.; Padilla-de la Rosa, J.D.; Esquivel-Solis, H.; Pelayo-Ortiz, C.; Guatemala-Morales, G.M. Optimization of 5-CQA Extraction Conditions from Green Coffee By-Product (Coffea arabica) Using a Response-Surface Design and the Study of Its Extraction Kinetics. Molecules 2022, 27, 5704. [Google Scholar] [CrossRef]
- Zeng, G.; Ran, Y.; Huang, X.; Li, Y.; Zhang, M.; Ding, H.; Ma, Y.; Ma, H.; Jin, L.; Sun, D. Optimization of Ultrasonic-Assisted Extraction of Chlorogenic Acid from Tobacco Waste. Int. J. Environ. Res. Public Health 2022, 19, 1555. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, S.; Abid, M.; Wu, T.; Hashim, M.M.; Saeeduddin, M.; Hu, B.; Lei, S.; Zeng, X. Ultrasound Extraction of Bioactives from Carrot. J. Food Process. Preserv. 2015, 39, 1878–1888. [Google Scholar] [CrossRef]
- Castaldo, L.; Izzo, L.; De Pascale, S.; Narváez, A.; Rodriguez-Carrasco, Y.; Ritieni, A. Chemical Composition, In Vitro Bioaccessibility and Antioxidant Activity of Polyphenolic Compounds from Nutraceutical Fennel Waste Extract. Molecules 2021, 26, 1968. [Google Scholar] [CrossRef]
- Riciputi, Y.; Diaz-de-Cerio, E.; Akyol, H.; Capanoglu, E.; Cerretani, L.; Caboni, M.F.; Verardo, V. Establishment of ultrasound-assisted extraction of phenolic compounds from industrial potato by-products using response surface methodology. Food Chem. 2018, 269, 258–263. [Google Scholar] [CrossRef]
- Wang, L.-T.; Gao, M.-Z.; Yang, Q.; Cui, Q.; Jian, Y.; Fan, X.-H.; Yao, L.-P.; Fu, Y.-J. An Efficient Strategy Based on Liquid–Liquid Extraction with Acid Condition and HSCCC for Rapid Enrichment and Preparative Separation of Three Caffeoylquinic Acid Isomers From Mulberry Leaves. J. Chromatogr. Sci. 2019, 57, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Du, X.; Zu, Y.; Yang, L. A new approach for preparation of essential oil, followed by chlorogenic acid and hyperoside with microwave-assisted simultaneous distillation and dual extraction (MSDDE) from Vaccinium uliginosum leaves. Ind. Crops Prod. 2015, 77, 809–826. [Google Scholar] [CrossRef]
- Puga, H.; Alves, R.; Costa, A.; Vinha, A.; Oliveira, M. Multi-frequency multimode modulated technology as a clean, fast, and sustainable process to recover antioxidants from a coffee by-product. J. Clean. Prod. 2017, 168, 14–21. [Google Scholar] [CrossRef]
- Santos da Silveira, J.; Durand, N.; Lacour, S.; Belleville, M.-P.; Perez, A.; Loiseau, G.; Dornier, M. Solid-state fermentation as a sustainable method for coffee pulp treatment and production of an extract rich in chlorogenic acids. Food Bioprod. Process. 2019, 115, 175–184. [Google Scholar] [CrossRef]
- Alves Filho, E.G.; Sousa, V.M.; Rodrigues, S.; de Brito, E.S.; Fernandes, F.A. Green ultrasound-assisted extraction of chlorogenic acids from sweet potato peels and sonochemical hydrolysis of caffeoylquinic acids derivatives. Ultrason. Sonochem. 2020, 63, 104911. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, J.; Jia, J.; Liu, J.; Ling, X.; Lu, D. Enrichment and Purification of Total Chlorogenic Acids from Tobacco Waste Extract with Macroporous Resins. Sep. Sci. Technol. 2010, 45, 794–800. [Google Scholar]
- Wang, D.; Du, N.; Wen, L.; Zhu, H.; Liu, F.; Wang, X.; Du, J.; Li, S. An Efficient Method for the Preparative Isolation and Purification of Flavonoid Glycosides and Caffeoylquinic Acid Derivatives from Leaves of Lonicera japonica Thunb. Using High Speed Counter-Current Chromatography (HSCCC) and Prep-HPLC Guided by DPPH-HPL. Molecules 2017, 22, 229. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, F.; Dai, G.; Zhang, P.; Tang, K. Preparative separation of chlorogenic acid from Eucommia ulmoides extract via fractional extraction. J. Chem. Technol. Biotechnol. 2020, 95, 2139–2148. [Google Scholar] [CrossRef]
- Han, R.; Zeng, J.; Han, L.; Wang, Y.; Chang, Q.; Zhang, X.; Zhou, J. Application of integrated membrane technology in purification of chlorogenic acid. Desalination Water Treat. 2015, 55, 2165–2170. [Google Scholar] [CrossRef]
- Li, G.; Shi, Z.; Li, D. Efficient synthesis of boronate affinity-based chlorogenic acid-imprinted magnetic nanomaterials for the selective recognition of chlorogenic acid in fruit juices. New J. Chem. 2020, 44, 11013–11021. [Google Scholar] [CrossRef]
- Mansour, M.; Abdel-Shafy, H.; Mehaya, F. Valorization of food solid waste by recovery of polyphenols using hybrid molecular imprinted membrane. J. Environ. Chem. Eng. 2018, 6, 4160–4170. [Google Scholar] [CrossRef]
- Tan, Z.; Wang, C.; Yi, Y.; Wang, H.; Li, M.; Zhou, W.; Tan, S.; Li, F. Extraction and purification of chlorogenic acid from ramie (Boehmeria nivea L. Gaud) leaf using an ethanol/salt aqueous two-phase system. Sep. Purif. Technol. 2014, 132, 396–400. [Google Scholar] [CrossRef]
- Dil, E.; Doustimotlagh, A.; Javadian, H.; Asfaram, A.; Ghaedi, M. Nano-sized Fe3O4@SiO2-molecular imprinted polymer as a sorbent for dispersive solid-phase microextraction of melatonin in the methanolic extract of Portulaca oleracea, biological, and water samples. Talanta 2021, 221, 121620. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Zhang, Z.; Liao, H.; Nie, L.; Yao, S. Separation and purification of chlorogenic acid by molecularly imprinted polymer monolithic stationary phase. J. Chromatogr. A 2005, 1098, 66–74. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, S.; He, J.; Thirumdas, R.; Montesano, D.; Barba, F. Enzyme-assisted extraction of polyphenol from edible lotus (Nelumbo nucifera) rhizome knot: Ultra-filtration performance and HPLC-MS2 profile. Food Res. Int. 2018, 111, 291–298. [Google Scholar] [CrossRef]
- Negrel, J.; Javelle, F.; Morandi, D. Detection of a plant enzyme exhibiting chlorogenate-dependant caffeoyltransferase activity in methanolic extracts of arbuscular mycorrhizal tomato roots. Plant Physiol. Biochem. 2013, 66, 77–83. [Google Scholar] [CrossRef]
- Wianowska, D.; Typek, R.; Dawidowicz, A.L. Chlorogenic acid stability in pressurized liquid extraction conditions. J. AOAC Int. 2015, 98, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.; Zheng, W.; Kuhnert, N. Profiling the chlorogenic acids of aster by HPLC–MSn. Phytochem. Anal. 2006, 17, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Şahin, S. Tailor-designed deep eutectic liquids as a sustainable extraction media: An alternative to ionic liquids. J. Pharm. Biomed. Anal. 2019, 174, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.-Z.; Cui, Q.; Wang, L.-T.; Meng, Y.; Yu, L.; Li, Y.-Y.; Fu, Y.-J. A green and integrated strategy for enhanced phenolic compounds extraction from mulberry (Morus alba L.) leaves by deep eutectic solvent. Microchem. J. 2020, 154, 104598. [Google Scholar] [CrossRef]
- Yue, Y.; Huang, Q.; Fu, Y.; Chang, J. A quick selection of natural deep eutectic solvents for the extraction of chlorogenic acid from herba artemisiae scopariae. RCS Adv. 2020, 10, 23403–23409. [Google Scholar] [CrossRef]
- Tan, T.; Lai, C.-J.-S.; Yang, H.O.; He, M.-Z.; Feng, Y. Ionic liquid-based ultrasound-assisted extraction and aqueous two-phase system for analysis of caffeoylquinic acids from Flos Lonicerae japonicae. J. Pharm. Biomed. Anal. 2016, 120, 134–141. [Google Scholar] [CrossRef]
- Burniol-Figols, A.; Cenian, K.; Skiadas, I.V.; Gavala, H.N. Integration of chlorogenic acid recovery and bioethanol production from spent coffee grounds. Biochem. Eng. J. 2016, 111, 54–64. [Google Scholar] [CrossRef]
- Zuorro, A.; Maffei, G.; Lavecchia, R. Reuse potential of artichoke (Cynara scolimus L.) waste for the recovery of phenolic compounds and bioenergy. J. Clean. Prod. 2016, 111, 279–284. [Google Scholar] [CrossRef]
- Pujol, D.; Liu, C.; Gominho, J.; Olivella, M.; Fiol, N.; Villaescusa, I.; Pereira, H. The chemical composition of exhausted coffee waste. Ind. Crops Prod. 2013, 50, 423–429. [Google Scholar] [CrossRef]
- del Pozo, C.; Bartrolí, J.; Alier, S.; Puy, N.; Fàbregas, E. Production of antioxidants and other value-added compounds from coffee silverskin via pyrolysis under a biorefinery approach. Waste Manag. 2020, 109, 19–27. [Google Scholar] [CrossRef]
- Schievano, A.; Adani, F.; Buessing, L.; Botto, A.; Casoliba, E.N.; Rossoni, M.; Goldfarb, J.L. An integrated biorefinery concept for olive mill waste management: Supercritical CO2 extraction and energy recovery. Green Chem. 2015, 17, 2874–2887. [Google Scholar] [CrossRef]
- Mayanga-Torres, P.; Lachos-Perez, D.; Rezende, C.; Prado, J.; Ma, Z.; Tompsett, G.; Timko, M.; Forster-Carneiro, T. Valorization of coffee industry residues by subcritical water hydrolysis: Recovery of sugars and phenolic compounds. J. Supercrit. Fluids 2017, 120, 75–85. [Google Scholar] [CrossRef]
- Janissen, B.; Huynh, T. Chemical composition and value-adding applications of coffee industry by-products: A review. Resour. Conserv. Recycl. 2018, 128, 110–117. [Google Scholar] [CrossRef]
- Kondamudi, N.; Mohapatra, S.; Misra, M. Spent Coffee Grounds as a Versatile Source of Green Energy. J. Agric. Food Chem. 2008, 56, 11757–11760. [Google Scholar] [CrossRef]
- Gupta, N.; Poddar, K.; Sarkar, D.; Kumari, N.; Padhan, B.; Sarkar, A. Fruit waste management by pigment production and utilization of residual as bioadsorbent. J. Environ. Manag. 2019, 244, 138–143. [Google Scholar] [CrossRef]
- Nisar, T.; Wang, Z.-C.; Alim, A.; Iqbal, M.; Yang, X.; Sun, L.; Guo, Y. Citrus pectin films enriched with thinned young apple polyphenols for potential use as bio-based active packaging. CYTA J. Food. 2019, 17, 695–705. [Google Scholar] [CrossRef]
- Kamh, M.; Hedia, R. NPK-Liquid Fertilizer Based on Humic-Like Substances Extracted from Spent Coffee Grounds: Extraction, Preparation and Application to Maize. Alex. Sci. Exch. J. 2018, 39, 260–267. [Google Scholar] [CrossRef]
- Kobylinska, N.; Shakhovsky, A.; Khainakova, O.; Klymchuk, D.; Avdeeva, L.; Ratushnyak, Y.; Duplij, V.; Matvieieva, N. ‘Hairy’ root extracts as source for ‘green’ synthesis of silver nanoparticles and medical applications. RSC Adv. 2020, 10, 39434–39446. [Google Scholar] [CrossRef] [PubMed]
- Al-Farsi, M.; Lee, C.Y. Optimization of phenolics and dietary fibre extraction from date seeds. Food Chem. 2008, 108, 977–985. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.; Mertz, C.; Morel, G.; Lacour, S.; Belleville, M.-P.; Durand, N.; Dornier, M. Alcoholic fermentation as a potential tool for coffee pulp detoxification and reuse: Analysis of phenolic composition and caffeine content by HPLC-DAD-MS/MS. Food Chem. 2020, 319, 126600. [Google Scholar] [CrossRef]
- Cai, W.; Chen, T.; Lei, M.; Wan, X. Effective strategy to recycle arsenic-accumulated biomass of Pteris vittata with high benefits. Sci. Total Environ. 2021, 756, 143890. [Google Scholar] [CrossRef]
- Sefkow, M.; Kelling, A.; Schilde, U. First Efficient Syntheses of 1-, 4-, and 5-Caffeoylquinic Acid. Eur. J. Org. Chem. 2001, 2001, 2735–2742. [Google Scholar] [CrossRef]
- Sefkow, M. First Efficient Synthesis of Chlorogenic Acid. Eur. J. Org. Chem. 2001, 2001, 1137–1141. [Google Scholar] [CrossRef]
- Kuhnert, N.; Jaiswal, R.; Matei, M.; Sovdat, T.; Deshpande, S. How to distinguish between feruloyl quinic acids and isoferuloyl quinic acids by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Smarrito, C.M.; Munari, C.; Roberta, F.; Barron, D. A novel efficient and versatile route to the synthesis of 5-O-feruloylquinic acids. Org. Biomol. Chem. 2008, 6, 986–987. [Google Scholar] [CrossRef] [PubMed]
- Kadidae, L.; Usami, A.; Koyama, T.; Honda, M.; Kunimoto, K.-K. New route for synthesis of 3- and 5-caffeoylquinic acids via protected quinic acids. Eur. J. Chem. 2015, 6, 367–373. [Google Scholar] [CrossRef]
- Dokli, I.; Navarini, L.; Hameršak, Z. Syntheses of 3-, 4-, and 5-O-feruloylquinic acids. Tetrahedron Asymmetry 2013, 24, 785–790. [Google Scholar] [CrossRef]
- Gutiérrez Ortiz, A.; Berti, F.; Navarini, L.; Monteiro, A.; Resmini, M.; Forzato, C. Synthesis of p-coumaroylquinic acids and analysis of their interconversion. Tetrahedron Asymmetry 2017, 28, 419–427. [Google Scholar] [CrossRef]
- De Pooter, H.; De Brucker, J.; van Sumere, C.F. Synthesis of 3-0-o.coumaryl-, 4-0-o.coumaryl-and 3-0-ferulyl-d-(-)-quinic acid. Improved synthesis of 3-0-sinapyl-d-(-)-quinic acid. Bull. Soc. Chim. Belg. 1976, 85, 663–671. [Google Scholar] [CrossRef]
- Nagaoka, T.; Baskota, A.H.; Xiong, Q.; Tezuka, Y.; Kadota, S. Synthesis and antihepatotoxic and antiproliferative activities of di- and tri-O-caffeoylquinic acid derivatives. J. Tradit. Med. 2001, 18, 183–190. [Google Scholar]
- Miyamae, Y.; Kurisu, M.; Han, J.; Isoda, H.; Shigemori, H. Structure-activity relationship of caffeoylquinic acids on the accelerating activity on ATP production. Chem. Pharm. Bull. 2011, 59, 502–507. [Google Scholar] [CrossRef]
- Brummond, K.; DeForrest, J. Synthesis of the Naturally Occurring (-)-1,3,5-Tri-O-Caffeoylquinic Acid. Synlett 2009, 9, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- Raheem, K.S.; Botting, N.P.; Williamson, G.; Barron, D. Total synthesis of 3,5-O-dicaffeoylquinic acid and its derivatives. Tetrahedron Lett. 2011, 52, 7175–7177. [Google Scholar] [CrossRef]
- ChemicalBook. Chemicalbook.com. 2021. Available online: https://www.chemicalbook.com/ (accessed on 15 April 2021).
- Wang, X.; Qin, L.; Zhou, J.; Fan, X. A novel design to screen chlorogenic acid-producing microbial strains from the environment. Sci. Rep. 2018, 8, 14756. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sang, X.; Li, S.; Zhang, S.; Bai, L. Studies on a chlorogenic acid-producing endophytic fungi isolated from Eucommia ulmoides Oliver. J. Ind. Microbiol. Biotechnol. 2010, 37, 447–454. [Google Scholar] [CrossRef]
- Gonçalves Tavares, D.; Viana Lessa Barbosa, B.; Lopes Ferreira, R.; Ferreira Duarte, W.; Gomes Cardoso, P. Antioxidant activity and phenolic compounds of the extract from pigment-producing fungi isolated from Brazilian caves. Biocatal. Agric. Biotechnol. 2018, 16, 148–154. [Google Scholar] [CrossRef]
- Ahmad, B.; Rizwan, M.; Rauf, A.; Raza, M.; Bashir, S.; Molnár, J.; Csonka, A.; Szabo, D.; Mubarak, M.S.; Noor, M.; et al. Isolation of Chlorogenic Acid from Soil Borne Fungi Screlotium rolfsii, their Reversal of Multidrug Resistance and Anti-Proliferative in Mouse Lymphoma Cells. Med. Chem. 2017, 13, 721–726. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, Z.; Tian, Z.; Sun, J.; Li, Y.; Fan, X. Validation of spectrophotometric determination of chlorogenic acid in fermentation broth and fruits. Food Chem. 2019, 278, 170–177. [Google Scholar] [CrossRef]
- Buraimoh, O.; Ilori, M.; Amund, O.; Isanbor, C.; Michel, F. The degradation of coniferyl alcohol and the complementary production of chlorogenic acids in the growth culture of Streptomyces albogriseolus KF977548 isolated from decaying wood residues. Process Biochem. 2017, 52, 22–29. [Google Scholar] [CrossRef]
- Kim, B.G.; Jung, W.D.; Mok, H.; Ahn, J.-H. Production of hydroxycinnamoyl-shikimates and chlorogenic acid in Escherichia coli: Production of hydroxycinnamic acid conjugates. Microb. Cell Factories 2013, 12, 15. [Google Scholar] [CrossRef]
- Lee, H.; Song, M.; Kim, B.-G.; Ahn, J.-H. Synthesis of chlorogenic acid and p-coumaroyl shikimate by expressing shikimate gene modules in Escherichia coli. J. Appl. Microbiol. 2022, 132, 1166–1175. [Google Scholar] [CrossRef]
- Cha, M.N.; Kim, H.J.; Kim, B.G.; Ahn, J.-H. Synthesis of Chlorogenic Acid and p-Coumaroyl Shikimates from Glucose Using Engineered Escherichia coli. J. Microbiol. Biotechnol. 2014, 24, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liang, C.; Liu, G.; Jin, J.-M.; Tao, Y.; Tang, S.-Y. De Novo Biosynthesis of Chlorogenic Acid Using an Artificial Microbial Community. J. Agric. Food Chem. 2021, 69, 2816–2825. [Google Scholar] [CrossRef] [PubMed]
- Moglia, A.; Comino, C.; Lanteri, S.; de Vos, R.; de Waard, P.; van Beek, T.A.; Goitre, L.; Retta, S.F.; Beekwilder, J. Production of novel antioxidative phenolic amides through heterologous expression of the plant’s chlorogenic acid biosynthesis genes in yeast. Metab. Eng. 2010, 12, 223–232. [Google Scholar]
- Eudes, A.; Mouille, M.; Robinson, D.S.; Benites, V.T.; Wang, G.; Roux, L.; Tsai, Y.-L.; Baidoo, E.E.K.; Chiu, T.-Y.; Heazlewood, J.L.; et al. Exploiting members of the BAHD acyltransferase family to synthesize multiple hydroxycinnamate and benzoate conjugates in yeast. Microb. Cell Factories 2016, 15, 198. [Google Scholar] [CrossRef]
- Xiao, F.; Lian, J.; Tu, S.; Xie, L.; Li, J.; Zhang, F.; Linhardt, R.J.; Huang, H.; Zhong, W. Metabolic Engineering of Saccharomyces cerevisiae for High-Level Production of Chlorogenic Acid from Glucose. ACS Synth. Biol. 2022, 11, 800–811. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).