
Role of Nutrients and Foods in Attenuation of Cardiac Remodeling through Oxidative Stress Pathways

 , , , ,
, , , , 
Abstract
1. Introduction
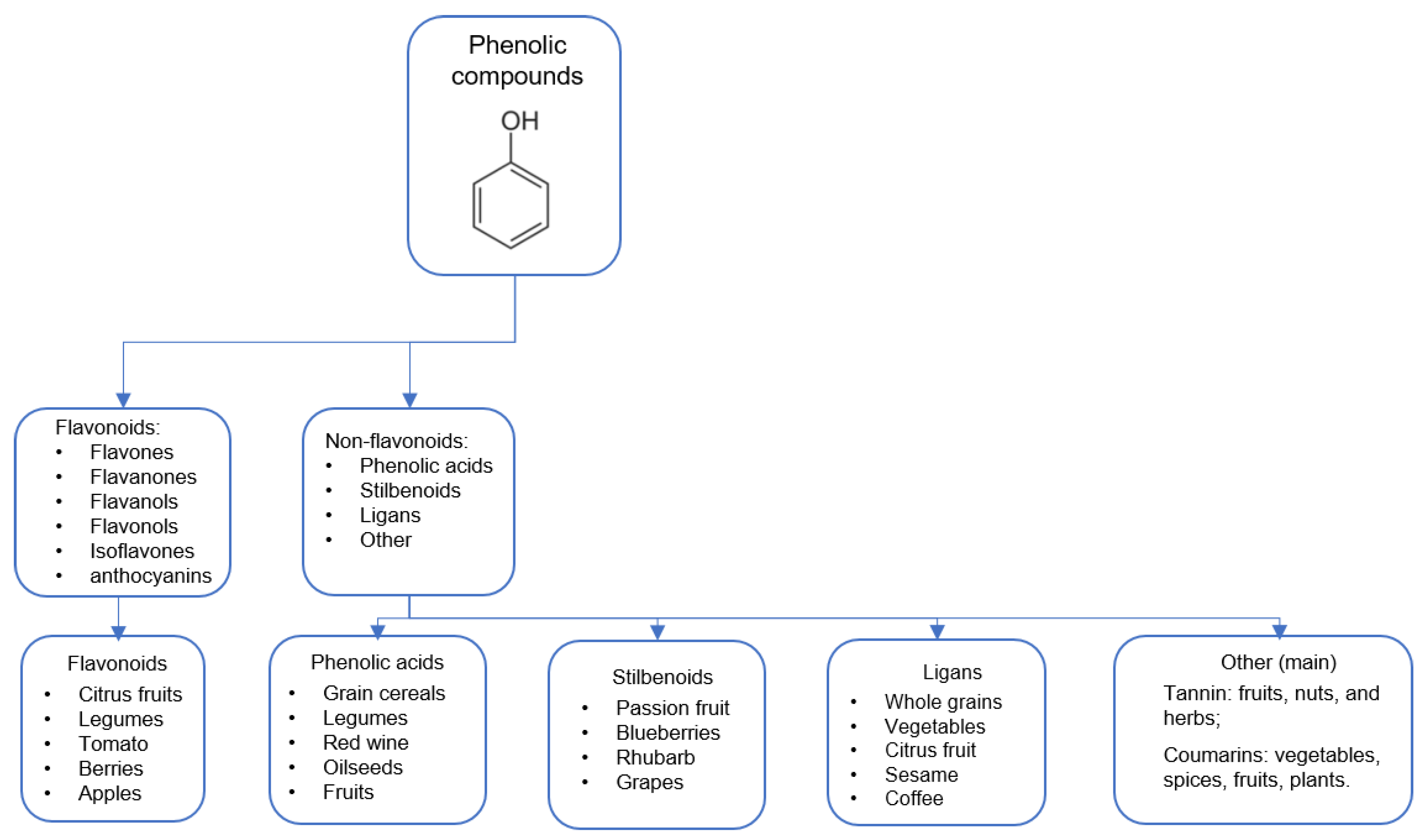
2. Diets
3. Macronutrients
3.1. Taurine
3.2. Fatty Acids
4. Micronutrients
4.1. All-Trans-Retinoic Acid
4.2. Beta-Carotene
4.3. Folic Acid
4.4. Vitamin E
4.5. Combinations of Vitamins
4.6. Zinc
4.7. Selenium
4.8. Magnesium
5. Phytochemicals/Phytonutrients
5.1. Tomato
5.2. Spondias
5.3. Açaí
5.4. Jaboticaba
5.5. Bergamot
5.6. Orange Juice
5.7. Raspberry
5.8. Blueberry
5.9. Cranberry
5.10. Cocoa
5.11. Rosemary
5.12. Camellia sinensis
5.13. Moringa
5.14. Citronella
5.15. Ginkgo biloba
5.16. Roselle
5.17. Atractylodis macrocephalae Rhizoma
5.18. Herba Houttuynia

{kind=link}
| Investigated Compound | Model | Cardiac Remodeling Effect | Redox Effect | Reference |
|---|---|---|---|---|
| Hypocaloric diet | Rats (obesity) Rats (metabolic syndrome) | ↓ hypertrophy ↓ hypertrophy ↓ fibrosis ↓ diastolic dysfunction | ↓ HO-1, ↓NQO1 ↓ NOX | [8] [9] |
| Ketogenic diet | Rats (underlying) | ↓ hypertrophy ↓ fibrosis | ↑ SOD, ↑ GPx, ↑ CAT, ↓ MDA ↓ 3-NT | [10] |
| Sardine oil-loaded microparticle | Cardiomyoblasts (doxorubicin toxicity) | ↓ apoptosis | ↓ ROS | [12] |
| Omega-3 and lignan | Rats (aortic stenosis) | improved LV contractile dysfunction, ↓ apoptosis, ↓ fibrosis | ↑ SOD, ↑ GPx, ↑ GST, ↑ MPO, ↓ MDA | [14] |
| Omega-3 | Rats (doxorubicin toxicity) | ↓ cardiac histopathological damages, ↓ apoptosis | ↑ SOD, ↑ GPx, ↓ MDA | [15] |
| α-Linolenic acid | Rats (doxorubicin toxicity) | improved cardiac function, ↓ cardiac histopathological damages | ↑ SOD, ↑ GPx, ↑ CAT, ↑ Nrf2 ↓ MDA | [16] |
| ATRA | Cardiomyocytes (mechanical stretch) Cardiomyocytes (doxorubicin) Rats (pressure overload) Rats (doxorubicin) | ↓ apoptosis ↓ cells death ↓ hypertrophic, fibrosis, and apoptosis ↓ myocardial histoarchitecture damages | ↓ ROS, ↑ SOD ↓ ROS, ↑ HO-1, ↑ SOD, ↑ Nrf2 ↑ SOD ↑SOD, ↑ GPx, ↑ CAT, ↑GSH, ↓ MDA | [19] [20] [21] [22] |
| Beta-carotene | Rats (cigarette smoking) | ↓ hypertrophy, preserved fibers morphological aspects | ↓ LH | [23] |
| Folic acid | Rats (monocrotaline) Rats (aging process) Rats (high-fat diet–obesity) | ↓ myocyte proliferation ↓ hypertrophy, ↓ fibrosis, ↓ apoptosis preserved LV function ↓cardiac dilatation, ↓ systolic dysfunction, ↓fibrosis | ↓ SOD ↓ MDA, ↓ 8-OHDG ↑ CAT, ↑ GSH, ↓MDA | [25] [26] [27] |
| Vitamin E | Rats (diabetic cardiomyopathy) | improved LV systolic and diastolic function | ↓ 8-iso PGF2α, ↓ GSSG | [28] |
| Combination of vitamins | Rabbits (tachycardia induced) Rabbits (myocardial infarction) | attenuated the increase of EDD and a decrease of FS attenuated LV dilation and dysfunction | ↑ GSH/GSSG ↓ 8-oxo-dG and ↑ GSH/GSSG ↓ 8-OHDG, ↓ 4-HNE | [29] [30] |
| Zinc | Rats (myocardial infarction) Rats (diabetic cardiomyopathy) | ↓ systolic and diastolic disfunction ↓ systolic and diastolic disfunction And ↓ fibrosis | ↑ CAT, ↓ SOD ↑ GSH ↑ MT | [32] [34] |
| Selenium | Rats (myocardial infarction) | ↓ necrosis, ↓ dilation of LV | ↑ GPx, ↑TrxR | [36] |
| Magnesium | Rats (L-NAME hypertension) | ↓ hypertrophy, restoring contractile dysfunction | ↓ O2−, ↓ H2O2, ↓ Carbonyl | [39] |
| Investigated Compound | Model | Cardiac Remodeling Effect | Redox Effect | Reference |
|---|---|---|---|---|
| Tomato | Rats (myocardial infarction) | improved diastolic dysfunction, ↓ interstitial fibrosis, ↓ hypertrophy | ↓ LH | [45] |
| Spondias | Rats (tobacco smoke) Rats (myocardial infarction) Rats (ISP— cardiotoxicity) | ↓ hypertrophy ↓hypertrophy, ↓ fibrosis ↓ hypertrophy, ↓ disruption and fragmentation myofibrils | ↑ SOD, ↑ GPx, ↓ LH ↓ LH ↑ SOD, ↑ CAT ↓ MDA, ↑ GSH | [48] [49] [50] |
| Açaí | Rats (high-fat-diet) Rats (Ischemia-Reperfusion) Rats (doxorubicin cardiotoxicity) | ↓ hypertrophy Poorer diastolic function Improved systolic function | ↑ GPx ↑ CAT, ↑ Nrf2 ↑ CAT, ↑SOD, ↑ GPx, ↓ LH ↓ LH | [52] [53] [54] |
| Jaboticaba | Rabbits (doxorubicin cardiotoxicity) Rats (myocardial infarction) | ↓ hypertrophy Improved diastolic function, ↓ fibrosis | ↓ NT, ↓ MDA ↑ GPx, ↓ LH | [55] [57] |
| Bergamot | Rats (doxorubicin cardiotoxicity) | Improved systolic function ↓ hypertrophy, ↓ apoptosis | ↓ ROS, ↓ MDA ↓ 8-OHDG | [58] |
| Orange juice | Rats (myocardial infarction) Rats (doxorubicin cardiotoxicity) | improved systolic and diastolic function, ↓ hypertrophy improved systolic and diastolic function, ↓hypertrophy | ↓GPx, ↑ HO-1 ↑ SOD, ↑GPx, ↑ CAT ↑ LH | [59] [60] |
| Red raspberry | Rats (obese diabetic) | ↓ levels of proteins associated involved in CR | ↓ NQO1, ↓GSTA4, ↓GSTP1 | [81] |
| Blueberry | Rats’ Cardiomyocytes (norepinephrine) | ↓ hypertrophy, ↓cell death ↑ contractile function | ↑ SOD, ↑ CAT | [62] |
| Cranberry | Rats (doxorubicin toxicity) | ↓histopathological lesions | ↑ GPx ↓ MPO, ↑ GSH, ↓ MDA | [63] |
| Cocoa–Carob blend | Rats (diabetic fatty) | mitigated cardiac dysfunction, hypertrophy, and fibrosis | ↓ NOX, ↓ ROS ↑ SIRT1, ↑Nrf2 | [65] |
| Rosemary | Rats (myocardial infarction) | Increased diastolic function ↓ hypertrophy | ↓ SOD, ↓ LH | [66] |
| Camella sinensis | Humans (dialysis patients) Rats (doxorubicin cardiotoxicity Rats (myocardial infarction) Rats (doxorubicin cardiotoxicity) | ↓ hypertrophy Preserved microarchitecture improved systolic and diastolic dysfunction, ↓ hypertrophy ↓ hypertrophy | ↓ p22 phox, ↑ HO-1, ↓ fosforilaion ERK1/2 ↑ GPx, ↑ GSH, ↑ GST, ↑ SOD, ↑ CAT, ↓ MDA ↓ SOD, ↓ CAT, ↑ Nrf-2, ↓ Carbonyl ↓ LH ↑ SOD and GPx | [68] [82] [83] [84] |
| Moringa | Rats (myocardial infarction) Rats (isoproterenol cardiotoxicity) Rats (isoproterenol cardiotoxicity) | attenuating contraction dysfunction, ↓ infarction area, apoptosis, and fibrosis ↓ necrosis, inflammatory infiltrate, and preservation of the myofibrillar structure ↓ improved systolic and diastolic function, preserved microarchitecture, ↓ myonecrosis | ↓gp91 phox ↑ SOD, ↑ CAT, ↑ GPx, ↑ GSH, ↓ MDA, ↓ ROS ↑ SOD, ↑ CAT, and GPx ↓ MDA | [85] [86] [71] |
| Citronellal | Rats (doxorubicin cardiotoxicity) | improvement of systolic function, ↓ apoptosis ↓ fibrosis | ↑ SOD, ↑ GSH ↓ MDA ↓ NHE1 | [73] |
| Ginkgolide A | Rats (aortic constriction) | ↓ hypertrophy improved systolic cardiac function, ↓ apoptosis | ↑ SOD, ↓ MDA, ↓NOX | [75] |
| Roselle | Rats (ISO myocardial infarction) | ameliorates cardiac diastolic dysfunction, ↓ hypertrophy, ↓ fibrosis | ↑ SOD, ↑GSH, ↓NOX, ↓8-iso PGF2α | [76] |
| Atractylodis macrocephalae rhizoma | Rats (ISO myocardial infarction) | ↓ hypertrophy, ↓ fibrosis | ↓ MDA | [78] |
| Herba Houttuynia | Rats (hyperlipidemic induced) | ↓ hypertrophy | ↑ SOD, ↑ GST, ↑ GPx, ↑ Nrf2, ↑NQO1, ↑HO-1, ↓ carbonyl | [80] |
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Azevedo, P.S.; Polegato, B.F.; Minicucci, M.F.; Paiva, S.A.; Zornoff, L.A. Cardiac remodeling: Concepts, clinical impact, pathophysiological mechanisms and pharmacologic treatment. Arq. Bras. De Cardiol. 2016, 106, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Ramachandra, C.J.A.; Cong, S.; Chan, X.; Yap, E.P.; Yu, F.; Hausenloy, D.J. Oxidative stress in cardiac hypertrophy: From molecular mechanisms to novel therapeutic targets. Free Radic. Biol. Med. 2021, 166, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, H.; Kinugawa, S.; Matsushima, S. Oxidative stress and heart failure. Am. J. Physiol. -Heart Circ. Physiol. 2011, 301, H2181–H2190. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.; Cross, C.E. Free radicals, antioxidants, and human disease: Where are we now? J. Lab. Clin. Med. 1992, 119, 598–620. [Google Scholar] [PubMed]
- Riley, P.A. Free radicals in biology: Oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Biol. 1994, 65, 27–33. [Google Scholar] [CrossRef]
- Valaei, K.; Taherkhani, S.; Arazi, H.; Suzuki, K. Cardiac oxidative stress and the therapeutic approaches to the intake of antioxidant supplements and physical activity. Nutrients 2021, 13, 3483. [Google Scholar] [CrossRef]
- An, H.S.; Lee, J.Y.; Choi, E.B.; Jeong, E.A.; Shin, H.J.; Kim, K.E.; Park, K.A.; Jin, Z.; Lee, J.E.; Koh, J.S.; et al. Caloric restriction reverses left ventricular hypertrophy through the regulation of cardiac iron homeostasis in impaired leptin signaling mice. Sci. Rep. 2020, 10, 7176. [Google Scholar] [CrossRef]
- Takatsu, M.; Nakashima, C.; Takahashi, K.; Murase, T.; Hattori, T.; Ito, H.; Murohara, T.; Nagata, K. Calorie restriction attenuates cardiac remodeling and diastolic dysfunction in a rat model of metabolic syndrome. Hypertension 2013, 62, 957–965. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, F.; Wang, J.; Zhang, D.; Zhao, X. Ketogenic diet attenuates aging-associated myocardial remodeling and dysfunction in mice. Exp. Gerontol. 2020, 140, 111058. [Google Scholar] [CrossRef]
- Baliou, S.; Adamaki, M.; Ioannou, P.; Pappa, A.; Panayiotidis, M.; Spandidos, D.; Christodoulou, I.; Kyriakopoulos, A.M.; Zoumpourlis, V. Protective role of taurine against oxidative stress (Review). Mol. Med. Rep. 2021, 24, 605. [Google Scholar] [CrossRef] [PubMed]
- Vishnu, K.V.; Kumar, K.K.A.; Chatterjee, N.S.; Lekshmi, R.G.K.; Sreerekha, P.R.; Mathew, S.; Ravishankar, C.N. Sardine oil loaded vanillic acid grafted chitosan microparticles, a new functional food ingredient: Attenuates myocardial oxidative stress and apoptosis in cardiomyoblast cell lines (H9c2). Cell Stress Chaperones 2018, 23, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Ardisson, L.P.; Rafacho, B.P.M.; Santos, P.P.; Assalin, H.; Gonçalves, A.F.; Azevedo, P.S.; Minicucci, M.F.; Polegato, B.F.; Okoshi, K.; Marchini, J.S.; et al. Taurine attenuates cardiac remodeling after myocardial infarction. Int. J. Cardiol. 2013, 168, 4925–4926. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ghule, A.E.; Kandhare, A.D.; Jadhav, S.S.; Zanwar, A.A.; Bodhankar, S.L. Omega-3-fatty acid adds to the protective effect of flax lignan concentrate in pressure overload-induced myocardial hypertrophy in rats via modulation of oxidative stress and apoptosis. Int. Immunopharmacol. 2015, 28, 751–763. [Google Scholar] [CrossRef]
- Uygur, R.; Aktas, C.; Tulubas, F.; Alpsoy, S.; Topcu, B.; Ozen, O. Cardioprotective effects of fish omega-3 fatty acids on doxorubicin-induced cardiotoxicity in rats. Hum. Exp. Toxicol. 2014, 33, 435–445. [Google Scholar] [CrossRef]
- Yu, X.; Cui, L.; Zhang, Z.; Zhao, Q.; Li, S. α-Linolenic acid attenuates doxorubicin-induced cardiotoxicity in rats through suppression of oxidative stress and apoptosis. Acta Biochim. Et Biophys. Sin. 2013, 45, 817–826. [Google Scholar] [CrossRef]
- Petiz, L.L.; Girardi, C.S.; Bortolin, R.C.; Kunzler, A.; Gasparotto, J.; Rabelo, T.K.; Matté, C.; Moreira, J.C.F.; Gelain, D.P. Vitamin A supplementation induces oxidative stress and suppresses IL-10 and HSP70 in the skeletal muscle of trained rats. Nutrients 2017, 9, 353. [Google Scholar] [CrossRef]
- Novo, R.; Azevedo, P.S.; Minicucci, M.F.; Zornoff, L.A.M.; Paiva, S.A.R. Effect of beta-carotene on oxidative stress and expression of cardiac connexin 43. Arq. Bras. De Cardiol. 2013, 101, 233–239. [Google Scholar] [CrossRef]
- Choudhary, R.; Baker, K.M.; Pan, J. All-trans retinoic acid prevents angiotensin II- and mechanical stretch-induced reactive oxygen species generation and cardiomyocyte apoptosis. J. Cell. Physiol. 2008, 215, 172–181. [Google Scholar] [CrossRef]
- Yang, L.; Luo, C.; Chen, C.; Wang, X.; Shi, W.; Liu, J. All-trans retinoic acid protects against doxorubicin-induced cardiotoxicity by activating the ERK2 signalling pathway. Br. J. Pharmacol. 2016, 173, 357–371. [Google Scholar] [CrossRef]
- Choudhary, R.; Palm-Leis, A.; Scott, R.C.; Guleria, R.S.; Rachut, E.; Baker, K.M.; Pan, J. All-trans retinoic acid prevents development of cardiac remodeling in aortic banded rats by inhibiting the renin-angiotensin system. Am. J. Physiol. -Heart Circ. Physiol. 2008, 294, H633–H644. [Google Scholar] [CrossRef] [PubMed]
- Khafaga, A.F.; El-Sayed, Y.S. All-trans-retinoic acid ameliorates doxorubicin-induced cardiotoxicity: In vivo potential involvement of oxidative stress, inflammation, and apoptosis via caspase-3 and p53 down-expression. Aunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Zornoff, L.A.M.; Matsubara, L.S.; Matsubara, B.B.; Okoshi, M.P.; Okoshi, K.; Dal Pai-Silva, M.; Carvalho, R.F.; Cicogna, A.C.; Padovani, C.R.; Novelli, E.L.; et al. Beta-carotene supplementation attenuates cardiac remodeling induced by one-month tobacco-smoke exposure in rats. Toxicol. Sci. 2006, 90, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Asbaghi, O.; Ghanavati, M.; Ashtary-Larky, D.; Bagheri, R.; Kelishadi, M.R.; Nazarian, B.; Nordvall, M.; Wong, A.; Dutheil, F.; Suziki, K.; et al. Effects of folic acid supplementation on oxidative stress markers: A systematic review and meta-analysis of randomized controlled trials. Antioxidants 2021, 10, 871. [Google Scholar] [CrossRef]
- Uzelac, J.J.; Djukic, T.; Radic, T.; Mutavdzin, S.; Stankovic, S.; Rakocevic, J.K.; Borovic, M.L.; Milic, N.; Simic, T.; Savic-Radojevic, A.; et al. Folic acid affects cardiometabolic, oxidative stress, and immunohistochemical parameters in monocrotaline-induced rat heart failure. Can. J. Physiol. Pharmacol. 2020, 98, 708–716. [Google Scholar] [CrossRef]
- Ye, S.; Zhou, X.; Chen, P.; Lin, J.F. Folic acid attenuates remodeling and dysfunction in the aging heart through the ER stress pathway. Life Sci. 2021, 264, 118718. [Google Scholar] [CrossRef]
- Li, W.; Tang, R.; Ouyang, S.; Ma, F.; Liu, Z.; Wu, J. Folic acid prevents cardiac dysfunction and reduces myocardial fibrosis in a mouse model of high-fat diet-induced obesity. Nutr. Metab. 2017, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.; Smith, H.M.; Hill, M.F. Dietary supplementation with vitamin E ameliorates cardiac failure in type I diabetic cardiomyopathy by suppressing myocardial generation of 8-iso-prostaglandin F2alpha and oxidized glutathione. J. Card. Fail. 2007, 13, 884–892. [Google Scholar] [CrossRef]
- Shite, J.; Qin, F.; Mao, W.; Kawai, H.; Stevens, S.Y.; Liang, C. Antioxidant vitamins attenuate oxidative stress and cardiac dysfunction in tachycardia-induced cardiomyopathy. J. Am. Coll. Cardiol. 2001, 38, 1734–1740. [Google Scholar] [CrossRef]
- Qin, F.; Yan, C.; Patel, R.; Liu, W.; Dong, E. Vitamins C and E attenuate apoptosis, β-adrenergic receptor desensitization, and sarcoplasmic reticular Ca2+ ATPase downregulation after myocardial infarction. Free Radic. Biol. Med. 2006, 40, 1827–1842. [Google Scholar] [CrossRef]
- Eide, D.J. The oxidative stress of zinc deficiency. Metallomics 2011, 3, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.F.; Polegato, B.F.; Fernandes, A.A.; Ishikawa, L.L.; Okoshi, K.; Bazan, S.G.Z.; Minicucci, M.F.; Azevedo, P.S.; Ikoma, M.R.; Penitenti, M.; et al. Zinc supplementation attenuates cardiac remodeling after experimental myocardial infarction. Cell. Physiol. Biochem. 2018, 50, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Ludbrook, P.; Karliner, J.S.; London, A.; Peterson, K.L.; Leopold, G.R.; O’Rourke, R.A. Posterior wall velocity: An unreliable index of total left ventricular performance in patients with coronary artery disease. Am. J. Cardiol. 1974, 33, 475–482. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Elsherif, L.; Song, Z.; Zhou, G.; Prabhu, S.D.; Saari, J.T.; Cai, L. Cardiac metallothionein induction plays the major role in the prevention of diabetic cardiomyopathy by zinc supplementation. Circulation 2006, 113, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Silvestrini, A.; Mordente, A.; Martino, G.; Bruno, C.; Vergani, E.; Meucci, E.; Mancini, A. The role of selenium in oxidative stress and non-thyroidal illness syndrome (NTIS): An overview. Curr. Med. Chem. 2020, 27, 423–449. [Google Scholar] [CrossRef] [PubMed]
- Tanguy, S.; Rakotovao, A.; Jouan, M.G.; Ghezzi, C.; de Leiris, J.; Boucher, F. Dietary selenium intake influences Cx43 dephosphorylation, TNF-α expression and cardiac remodeling after reperfused infarction. Mol. Nutr. Food Res. 2011, 55, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Metes-Kosik, N.; Luptak, I.; DiBello, P.M.; Handy, D.E.; Tang, S.S.; Zhi, H.; Qin, F.; Jacobsen, D.W.; Loscalzo, J.; Joseph, J. Both selenium deficiency and modest selenium supplementation lead to myocardial fibrosis in mice via effects on redox-methylation balance. Mol. Nutr. Food Res. 2012, 56, 1812–1824. [Google Scholar] [CrossRef]
- Wyparlo-Wszelaki, M.; Wasik, M.; Machón-Grecka, A.; Kasperczyk, A.; Bellanti, F.; Kasperczyk, S.; Dobrakpwski, M. Blood magnesium level and selected oxidative stress indices in lead-exposed workers. Biol. Trace Elem. Res. 2021, 199, 465–472. [Google Scholar] [CrossRef]
- Ozturk, N.; Olgar, Y.; Aslan, M.; Ozdemir, S. Effects of magnesium supplementation on electrophysiological remodeling of cardiac myocytes in L-NAME induced hypertensive rats. J. Bioenerg. Biomembr. 2016, 48, 425–436. [Google Scholar] [CrossRef]
- Yu, X.; Yang, T.; Qi, Q.; Du, Y.; Shi, J.; Liu, X.; Liu, Y.; Zhang, H.; Zhang, Z.; Yan, N. Comparison of the contents of phenolic compounds including flavonoids and antioxidant activity of rice (Oryza sativa) and Chinese wild rice (Zizania latifolia). Food Chem. 2021, 344, 128600. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Future J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Res. Int. 2020, 132, 109114. [Google Scholar] [CrossRef] [PubMed]
- Akinwumi, B.C.; Bordun, K.M.; Anderson, H.D. Biological Activities of Stilbenoids. Int. J. Mol. Sci. 2018, 19, 792. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.; Dwyer, J.; Adlercreutz, H.; Scalbert, A.; Jacques, P.; McCullough, M.L. Dietary lignans: Physiology and potential for cardiovascular disease risk reduction. Nutr. Rev. 2010, 68, 571–603. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.L.B.; Reis, P.P.; Severino, F.E.; Felix, T.F.; Braz, M.G.; Nogueira, F.R.; Silva, R.A.; Cardoso, A.C.; Lourenço, M.A.M.; Figueiredo, A.M.; et al. Tomato (Lycopersicon esculentum) or lycopene supplementation attenuates ventricular remodeling after myocardial infarction through different mechanistic pathways. J. Nutr. Biochem. 2017, 46, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Antunes, K.A.; Monteiro-Alfredo, T.; Cunha, J.S.M.; Espindola, P.P.T.; Oliveira, A.S.; de Oliveira, C.F.R.; de Carvalho, J.T.G.; Domingues, N.L.C.; Silva, D.B.; Olinto, S.C.F.; et al. Spondias purpurea L. bark extract protects against oxidative stress and reduces hypercholesterolemia in mice fed high-fat diet. Oxidative Med. Cell. Longev. 2022, 2022, 3046483. [Google Scholar]
- Tiburski, J.H.; Rosenthal, A.; Deliza, R.; de Oliveira Godoy, R.L.; Pacheco, S. Nutritional properties of yellow mombin (Spondias mombin L.) pulp. Food Res. Int. 2011, 44, 2326–2331. [Google Scholar] [CrossRef]
- Lourenço, M.A.M.; Braz, M.G.; Aun, A.G.; Pereira, B.L.B.; Figueiredo, A.M.; da Silva, R.A.C.; Kazmarek, E.M.; Alegre, P.H.C.; Bachiega, T.F.; Zanati, S.G.; et al. Spondias mombin supplementation attenuated cardiac remodelling process induced by tobacco smoke. J. Cell. Mol. Med. 2018, 22, 3996–4004. [Google Scholar] [CrossRef]
- Pereira, B.L.B.; Rodrigue, A.; Arruda, F.C.O.; Bachiega, T.F.; Lourenço, M.A.M.; Correa, C.R.; Azevedo, P.S.; Polegato, B.F.; Okoshi, K.; Fernandes, A.A.H.; et al. Spondias mombin L. attenuates ventricular remodelling after myocardial infarction associated with oxidative stress and inflammatory modulation. J. Cell. Mol. Med. 2020, 24, 7862–7872. [Google Scholar] [CrossRef]
- Akinmoladun, A.C.; Obuotor, E.M.; Barthwal, M.K.; Dikshit, M.; Farombi, E.O. Ramipril-like activity of Spondias mombin linn against no-flow ischemia and isoproterenol-induced cardiotoxicity in rat heart. Cardiovasc. Toxicol. 2010, 10, 295–305. [Google Scholar] [CrossRef]
- Yamaguchi, K.K.L.; Pereira, L.F.R.; Lamarão, C.V.; Lima, E.S.; da Veiga-Junior, V.F. Amazon acai: Chemistry and biological activities: A review. Food Chem. 2015, 179, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Alegre, P.; Mathias, L.; Lourenço, M.A.; Santos, P.P.D.; Gonçalves, A.; Fernandes, A.A.; Gaiolla, P.S.A.; Minicucci, M.F.; Zornoff, L.; Paiva, S.A.R.; et al. Euterpe oleracea Mart. (Açaí) reduces oxidative stress and improves energetic metabolism in myocardial ischemia-reperfusion injury in rats. Arq. Bras. De Cardiol. 2020, 114, 78–86. [Google Scholar]
- Arnoso, B.J.M.; Magliaccio, F.M.; de Araújo, C.A.; Soares, R.A.; Santos, I.B.; de Bem, G.F.; Fernandes-Santos, C.; Ognibene, D.T.; de Moura, R.S.; Resende, A.C.; et al. Açaí seed extract (ASE) rich in proanthocyanidins improves cardiovascular remodeling by increasing antioxidant response in obese high-fat diet-fed mice. Chem.-Biol. Interact. 2022, 351, 109721. [Google Scholar] [CrossRef]
- Mathias, L.M.B.S.; Alegre, P.H.C.; Dos Santos, I.O.F.; Bachiega, T.; Figueiredo, A.M.; Chiuso-Minicucci, F.; Fernandes, A.A.; Bazan, S.G.Z.; Minicucci, M.F.; Azevedo, P.S.; et al. Euterpe oleracea Mart. (Açai) Supplementation attenuates acute doxorubicin-induced cardiotoxicity in rats. Cell. Physiol. Biochem. 2019, 53, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Romão, P.V.M.; Palozi, R.A.C.; Guarnier, L.P.; Silva, A.O.; Lorençone, B.R.; Nocchi, S.R.; Moura, C.C.F.S.; Lourenço, E.L.B.; Silva, D.B.; Junior, A.G. Cardioprotective effects of Plinia cauliflora (Mart.) Kausel in a rabbit model of doxorubicin-induced heart failure. J. Ethnopharmacol. 2019, 242, 112042. [Google Scholar] [CrossRef]
- Da Silva, R.C.; Polegato, B.F.; Azevedo, P.S.; Fernandes, A.A.; Okoshi, K.; de Paiva, S.A.R.; Minicucci, M.F.; Zornoff, L.A.M. Jaboticaba (Myrciaria jaboticaba) attenuates ventricular remodeling after myocardial infarction in rats. Antioxidants 2022, 11, 249. [Google Scholar] [CrossRef]
- Maiuolo, J.; Carresi, C.; Gliozzi, M.; Musolino, V.; Scarano, F.; Coppoletta, A.R.; Guarnieri, L.; Nucera, S.; Scicchitano, M.; Bosco, F.; et al. Effects of bergamot polyphenols on mitochondrial dysfunction and sarcoplasmic reticulum stress in diabetic cardiomyopathy. Nutrients 2021, 13, 2476. [Google Scholar] [CrossRef]
- Carresi, C.; Musolino, V.; Gliozzi, M.; Maiuolo, J.; Mollace, R.; Nucera, S.; Maretta, A.; Sergi, D.; Muscoli, S.; Gratteri, S.; et al. Anti-oxidant effect of bergamot polyphenolic fraction counteracts doxorubicin-induced cardiomyopathy: Role of autophagy and c-kitposCD45negCD31neg cardiac stem cell activation. J. Mol. Cell. Cardiol. 2018, 119, 10–18. [Google Scholar] [CrossRef]
- Oliveira, B.C.; Santos, P.P.; Figueiredo, A.M.; Rafacho, B.P.M.; Ishikawa, L.; Zanati, S.G.; Fernandes, A.A.H.; Azevedo, P.S.; Polegato, B.F.; Zornoff, L.A.M.; et al. Influência do consumo de suco de laranja (Citrus sinensis) na remodelação cardíaca de ratos submetidos a infarto do miocárdio. Arq. Bras. De Cardiol. 2021, 116, 1127–1136. [Google Scholar] [CrossRef]
- Ribeiro, A.P.D.; Pereira, A.G.; Todo, M.C.; Fujimori, A.S.S.; Dos Santos, P.P.; Dantas, D.; Fernandes, A.A.; Zanati, S.G.; Hassimotto, N.M.A.; Zornoff, L.A.M.; et al. Pera orange (Citrus sinensis) and Moro orange (Citrus sinensis (L.) Osbeck) juices attenuate left ventricular dysfunction and oxidative stress and improve myocardial energy metabolism in acute doxorubicin-induced cardiotoxicity in rats. Nutrition 2021, 91–92, 111350. [Google Scholar] [CrossRef]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Red raspberries and their bioactive polyphenols: Cardiometabolic and neuronal health links. Adv. Nutr. 2016, 7, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Louis, X.L.; Thandapilly, S.J.; Kalt, W.; Vinqvist-Tymchuk, M.; Aloud, B.M.; Raj, P.; Yu, L.; Le, H.; Netticadan, T. Blueberry polyphenols prevent cardiomyocyte death by preventing calpain activation and oxidative stress. Food Funct. 2014, 5, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Elberry, A.A.; Abdel-Naim, A.B.; Abdel-Sattar, E.A.; Nagy, A.A.; Mosli, H.A.; Mohamadin, A.M.; Ashour, O.M. Cranberry (Vaccinium macrocarpon) protects against doxorubicin-induced cardiotoxicity in rats. Food Chem. Toxicol. 2010, 48, 1178–1184. [Google Scholar] [CrossRef]
- Akdeniz, E.; Yakışık, E.; Rasouli Pirouzian, H.; Akkın, S.; Turan, B.; Tipigil, E.; Toker, O.S.; Ozcan, O. Carob powder as cocoa substitute in milk and dark compound chocolate formulation. J. Food Sci. Technol. 2021, 58, 4558–4566. [Google Scholar] [CrossRef] [PubMed]
- García-Díez, E.; López-Oliva, M.E.; Caro-Vadillo, A.; Pérez-Vizcaíno, F.; Pérez-Jiménez, J.; Ramos, S.; Martín, M.A. Supplementation with a Cocoa–Carob blend, alone or in combination with metformin, attenuates diabetic cardiomyopathy, cardiac oxidative stress and inflammation in Zucker diabetic rats. Antioxidants 2022, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Rafacho, B.P.M.; dos Santos, P.P.; Gonçalves, A.F.; Fernandes, A.A.H.; Okoshi, K.; Chiuso-Minicucci, F.; Azevedo, P.S.; Zornoff, L.A.M.; Minicucci, M.F.; Wang, X.D.; et al. Rosemary supplementation (Rosmarinus oficinallis L.) attenuates cardiac remodeling after myocardial infarction in rats. PLoS ONE 2017, 12, e0177521. [Google Scholar]
- Coimbra, S.; Castro, E.; Rocha-Pereira, P.; Rebelo, I.; Rocha, S.; Santos-Silva, A. The effect of green tea in oxidative stress. Clin. Nutr. 2006, 25, 790–796. [Google Scholar] [CrossRef]
- Calo, L.A.; Vertolli, U.; Davis, P.A.; Maso, L.D.; Pagnin, E.; Ravarotto, V.; Maiolino, G.; Lupia, M.; Seccia, T.M.; Rossi, G.P. Molecular biology based assessment of green tea effects on oxidative stress and cardiac remodelling in dialysis patients. Clin. Nutr. 2014, 33, 437–442. [Google Scholar] [CrossRef]
- Ercan, K.; Gecesefa, O.F.; Taysi, M.E.; Ali, O.A.A.; Taysi, S. Moringa Oleifera: A review of its occurrence, pharmacological importance and oxidative stress. Mini Rev. Med. Chem. 2021, 21, 380–396. [Google Scholar] [CrossRef]
- Tarafdar, A.; Pula, G. The role of NADPH oxidases and oxidative stress in neurodegenerative disorders. Int. J. Mol. Sci. 2018, 19, 3824. [Google Scholar] [CrossRef]
- Panda, S. Butanolic fraction of Moringa oleifera Lam. (Moringaceae) attenuates isoprotrenol-induced cardiac necrosis and oxidative stress in rats: An EPR study. EXCLI J. 2015, 14, 64–74. [Google Scholar] [PubMed]
- Randriamboavonjy, J.I.; Loirand, G.; Vaillant, N.; Lauzier, B.; Derbré, S.; Michalet, S.; Pacaud, P.; Tesse, A. Cardiac protective Effects of Moringa oleifera seeds in spontaneous hypertensive rats. Am. J. Hypertens. 2016, 29, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qiu, Y.; Huang, N.; Liu, Y.; Wang, H.; Yu, Y.; Song, Y.T.; Wan, G.R.; Wang, S.X.; Li, P.; et al. Citronellal alleviates doxorubicin-induced cardiotoxicity by suppressing oxidative stress and apoptosis via Na+/H+ exchanger-1 inhibition. J. Biochem. Mol. Toxicol. 2022, 36, e22971. [Google Scholar] [CrossRef] [PubMed]
- Souza, G.A.; de Marqui, S.V.; Matias, J.N.; Guiguer, E.L.; Barbalho, S.M. Effects of Ginkgo biloba on diseases related to oxidative stress. Planta Med. 2020, 86, 376–386. [Google Scholar]
- You, W.; Wu, Z.; Ye, F.; Wu, X. Ginkgolide A protects adverse cardiac remodeling through enhancing antioxidation and nitric oxide utilization in mice with pressure overload. Die Pharm. -Int. J. Pharm. Sci. 2019, 74, 698–702. [Google Scholar]
- Si, L.Y.N.; Ramalingam, A.; Ali, S.S.; Aminuddin, A.; Ng, P.Y.; Latip, J.; Kamisah, Y.; Budin, S.B.; Zainalabidin, S. Roselle attenuates cardiac hypertrophy after myocardial infarction in vivo and in vitro. EXCLI J. 2019, 18, 876–892. [Google Scholar]
- Li, X.; Lin, J.; Han, W.; Mai, W.; Wang, L.; Li, Q.; Lin, M.; Bai, M.; Zhang, L.; Chen, D. Antioxidant ability and mechanism of rhizoma Atractylodes macrocephala. Molecules 2012, 17, 13457–13472. [Google Scholar]
- Cui, X.H.; Wang, H.L.; Wu, R.; Yao, P.A.; Wei, K.Z.; Gao, J.P. Effect of Atractylodes macrocephala rhizoma on isoproterenol-induced ventricular remodeling in rats. Mol. Med. Rep. 2018, 17, 2607–2613. [Google Scholar]
- Shingnaisui, K.; Dey, T.; Manna, P.; Kalita, J. Therapeutic potentials of Houttuynia cordata Thunb. against inflammation and oxidative stress: A review. J. Ethnopharmacol. 2018, 220, 35–43. [Google Scholar] [CrossRef]
- Cao, K.; Lv, W.; Liu, X.; Fan, Y.; Wang, K.; Feng, Z.; Jianshu, L.; Zang, W.; Xing, L.; Liu, J. Herba Houttuyniae extract benefits hyperlipidemic mice via activation of the AMPK/PGC-1α/Nrf2 cascade. Nutrients 2020, 12, 164. [Google Scholar] [CrossRef]
- Noratto, G.; Chew, B.P.; Ivanov, I. Red raspberry decreases heart biomarkers of cardiac remodeling associated with oxidative and inflammatory stress in obese diabetic db/db mice. Food Funct. 2016, 7, 4944–4955. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.; Haque, S.E.; Anwer, T.; Ahsan, M.N.; Safhi, M.M.; Alam, M.F. Cardioprotective effect of green tea extract on doxorubicin-induced cardiotoxicity in rats. Acta Pol. Pharm. 2014, 71, 861–868. [Google Scholar] [PubMed]
- Lustosa, B.B.; Polegato, B.; Minicucci, M.; Rafacho, B.; Santos, P.P.; Fernandes, A.A.; Okoshi, K.; Batista, D.; Modesto, P.; Gonçalves, A.; et al. Green tea (Cammellia sinensis) attenuates ventricular remodeling after experimental myocardial infarction. Int. J. Cardiol. 2016, 225, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Modesto, P.N.; Polegato, B.F.; dos Santos, P.P.; Grassi, L.D.V.; Molina, L.C.C.; Bazan, S.G.Z.; Pereira, E.J.; Fernandes, A.A.H.; Fabro, A.T.; Androcioli, V.N.; et al. Green Tea (Camellia sinensis) extract increased topoisomerase IIβ, improved antioxidant defense, and attenuated cardiac remodeling in an acute doxorubicin toxicity model. Oxidative Med. Cell. Longev. 2021, 2021, 8898919. [Google Scholar] [CrossRef]
- Li, Y.J.; Ji, Q.Q.; Wang, Z.; Shen, L.H.; He, B. Moringa oleifera seeds mitigate myocardial injury and prevent ventricular failure induced by myocardial infarction. Am. J. Transl. Res. 2020, 12, 4511–4521. [Google Scholar]
- Nandave, M.; Ojha, S.K.; Joshi, S.; Kumari, S.; Arya, D.S. Moringa oleifera leaf extract prevents isoproterenol-induced myocardial damage in rats: Evidence for an antioxidant, antiperoxidative, and cardioprotective intervention. J. Med. Food 2009, 12, 47–55. [Google Scholar] [CrossRef]
- Steinhubl, S.R. Why have antioxidants failed in clinical trials? Am. J. Cardiol. 2008, 101, 14D–19D. [Google Scholar] [CrossRef]
- Xie, M.; Burchfield, J.S.; Hill, J.A. Pathological ventricular remodeling: Therapies: Part 2 of 2. Circulation 2013, 128, 1021–1030. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazzarin, T.; Garcia, L.R.; Martins, D.; Queiroz, D.A.R.; Tonon, C.R.; Balin, P.d.S.; Polegato, B.F.; Paiva, S.A.R.d.; Azevedo, P.S.; Minicucci, M.; et al. Role of Nutrients and Foods in Attenuation of Cardiac Remodeling through Oxidative Stress Pathways. Antioxidants 2022, 11, 2064. https://doi.org/10.3390/antiox11102064
Lazzarin T, Garcia LR, Martins D, Queiroz DAR, Tonon CR, Balin PdS, Polegato BF, Paiva SARd, Azevedo PS, Minicucci M, et al. Role of Nutrients and Foods in Attenuation of Cardiac Remodeling through Oxidative Stress Pathways. Antioxidants. 2022; 11(10):2064. https://doi.org/10.3390/antiox11102064
Chicago/Turabian StyleLazzarin, Taline, Leonardo Rufino Garcia, Danilo Martins, Diego Aparecido Rios Queiroz, Carolina Rodrigues Tonon, Paola da Silva Balin, Bertha Furlan Polegato, Sergio Alberto Rupp de Paiva, Paula Schmidt Azevedo, Marcos Minicucci, and et al. 2022. "Role of Nutrients and Foods in Attenuation of Cardiac Remodeling through Oxidative Stress Pathways" Antioxidants 11, no. 10: 2064. https://doi.org/10.3390/antiox11102064
APA StyleLazzarin, T., Garcia, L. R., Martins, D., Queiroz, D. A. R., Tonon, C. R., Balin, P. d. S., Polegato, B. F., Paiva, S. A. R. d., Azevedo, P. S., Minicucci, M., & Zornoff, L. (2022). Role of Nutrients and Foods in Attenuation of Cardiac Remodeling through Oxidative Stress Pathways. Antioxidants, 11(10), 2064. https://doi.org/10.3390/antiox11102064