

Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid


Abstract
1. Introduction
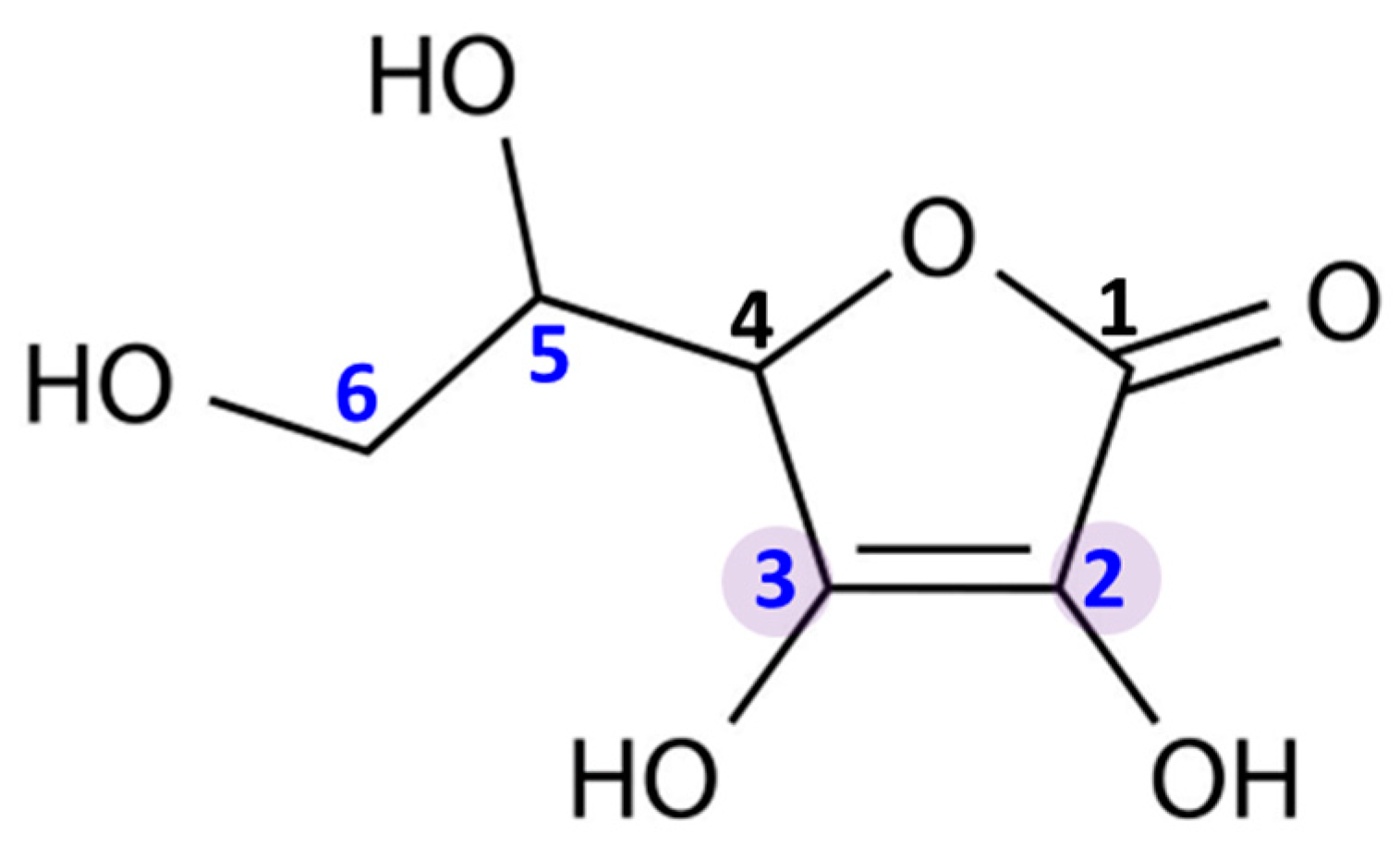
2. Bioavailability and Main Functions
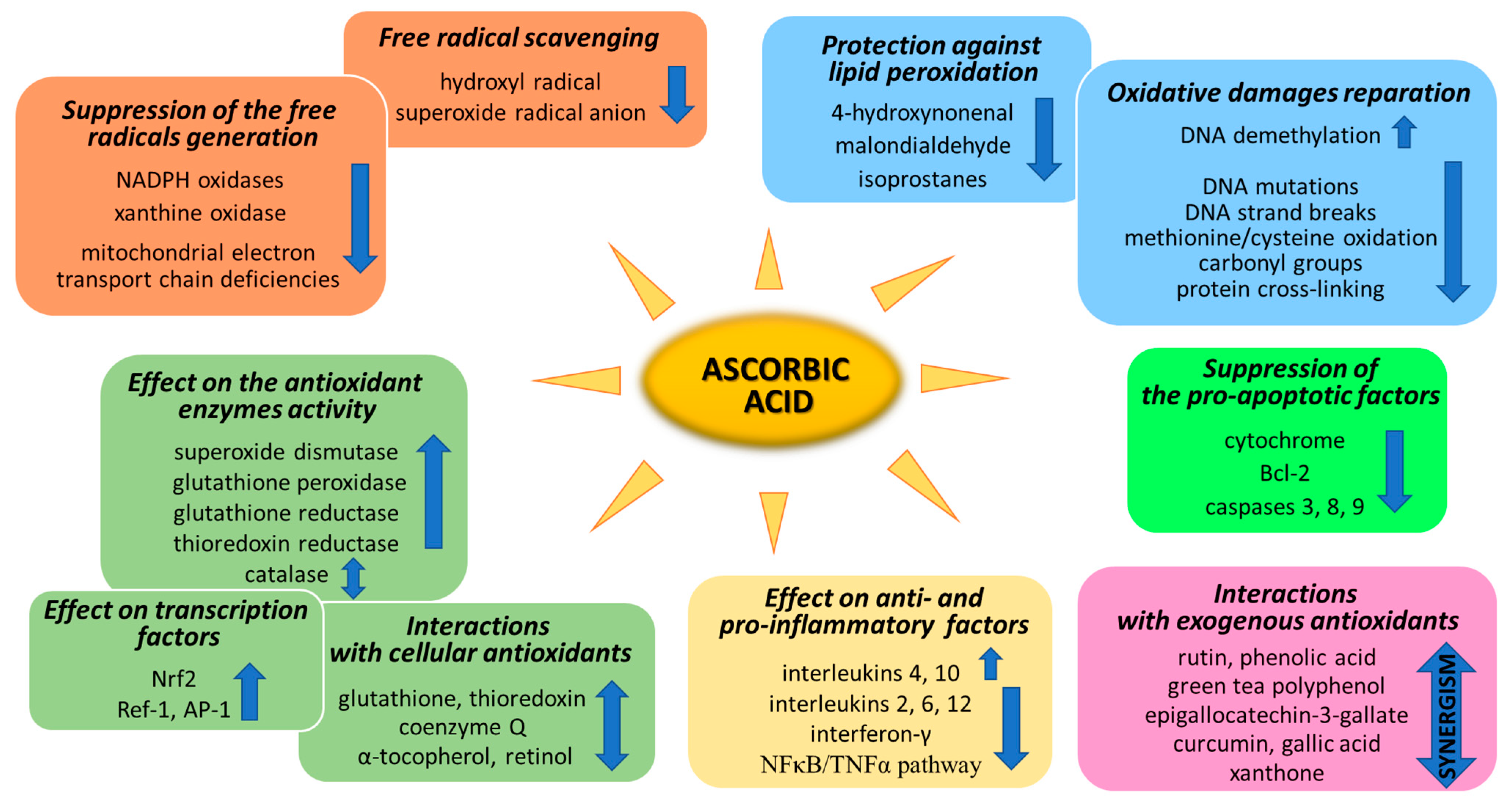
3. Antioxidant Properties
3.1. Suppression of Generation of Free Radicals
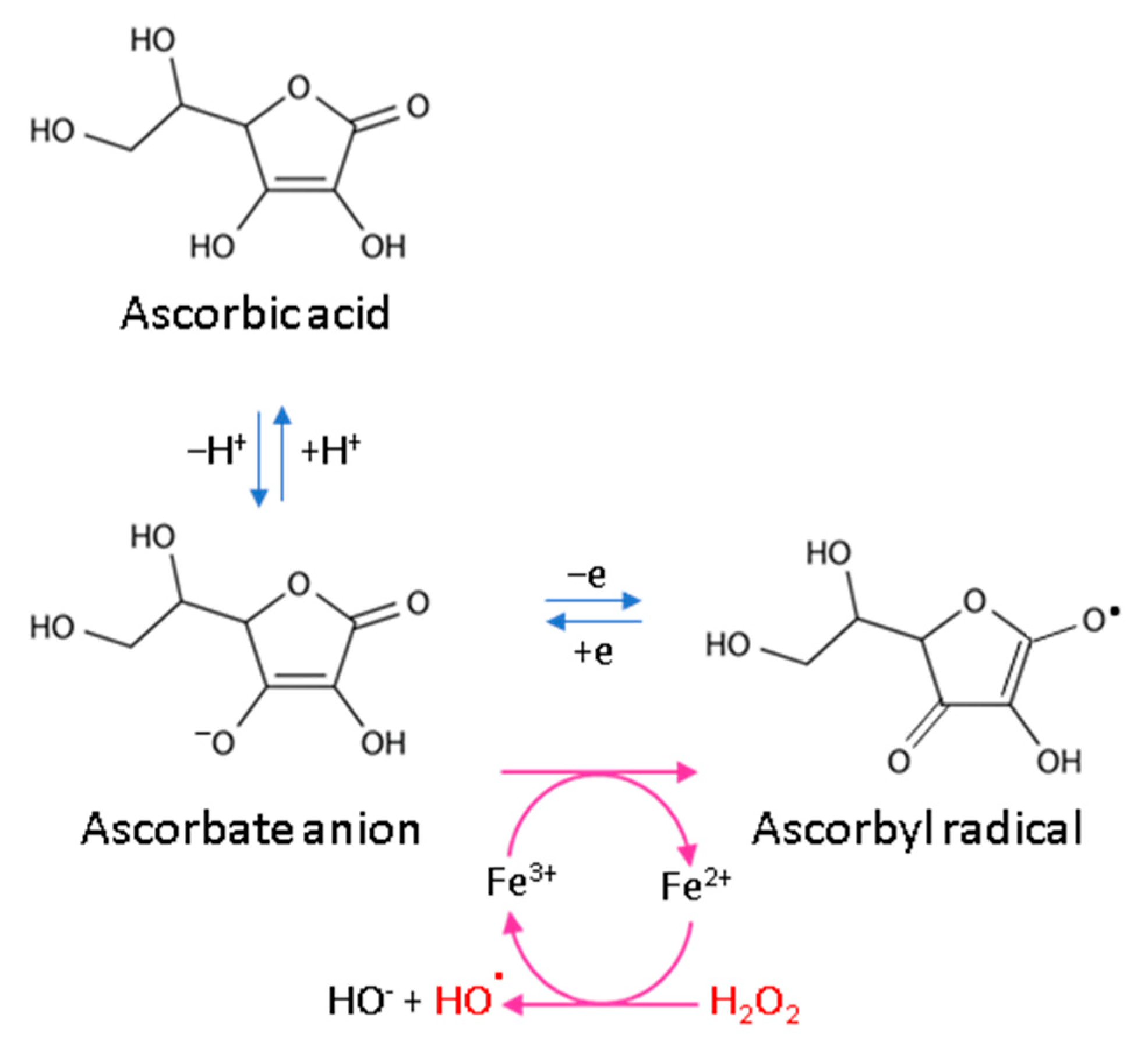
3.2. ROS Scavenging by Ascorbic Acid
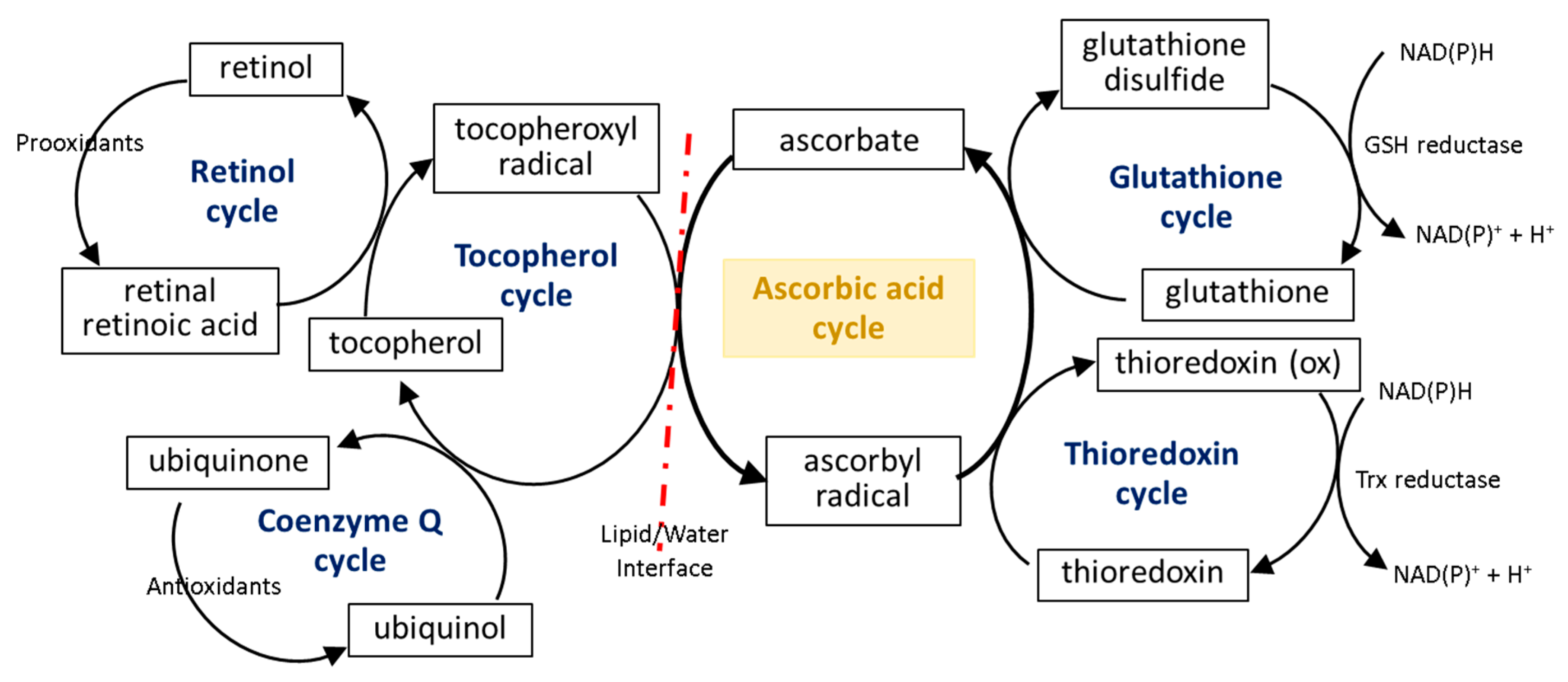
3.3. Ascorbic Acid Interaction with the Cellular Antioxidant System
3.4. Effect of Ascorbic Acid on Cytoprotective Gene Transcription
4. Ascorbic Acid and Oxidative Modifications
4.1. Oxidative Damage Repair
4.2. Prevention Lipid Peroxidation
5. Anti-Inflammatory Properties

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Biological Material | Conditions | Effect of Ascorbic Acid | Refs. |
|---|---|---|---|---|
| CRP | Plasma | Physiological conditions | Downregulation | [126] |
| CRP | Plasma | Inflammation (cardiopulmonary bypass graft surgery) | No effect | [127] |
| IFN-γ | Plasma | Physiological conditions | Downregulation | [128] |
| IL-4 | Plasma | Oxidative stress (Alzheimer’s disease) | Upregulation | [129] |
| IL-6 | Plasma | Oxidative stress (Alzheimer’s disease) | Downregulation | [129] |
| IL-6 IL-8 | Plasma | Oxidative stress (cardiopulmonary bypass initiation) | No effect | [130] |
| NFκB | Skin cells | Oxidative stress (UV irradiation) | Downregulation | [33,134] |
| NFκB | Cell lines ECV304, HUVEC, HeLa, U937, HL-60, MCF7 | Inflammation (induced experimentally/tumor proliferation) | Activation of kinases involved in IκK phosphorylation | [135,136] |
| NFκB | Acute myeloid leukemia | Inflammation (tumor proliferation) | Suppression of NFκB binding to DNA | [137] |
| TNF-α | Brain tissue | Physiological conditions neurotoxicity (induced experimentally) | Downregulation | [138] |
| TNF-α | Splenocytes | Inflammation (induced experimentally) | Downregulation | [140] |
| TNF-α | Endometrial tissue | Oxidative stress (endometritis) | No effect | [143] |
| IL-6 | Brain tissue | Physiological conditions neurotoxicity (induced experimentally) | Downregulation | [138] |
| IL-6 | Skeletal muscle | Contracting skeletal muscle | Downregulation | [139] |
| IL-6 IL-12 | Splenocytes | Inflammation (induced experimentally) | Downregulation | [140] |
| IL-4 | Splenocytes | Inflammation (induced experimentally) | Upregulation | [140] |
| IL-2 IL-6 | Peripheral blood mononuclear cells | Animals with hereditary deficiency in ascorbate synthesis | Downregulation | [141,142] |
| IL-10 | Splenocytes | Inflammation (induced experimentally) | Upregulation | [140] |
| IL-10 | Endometrial tissue | Oxidative stress (endometritis) | Upregulation | [143] |
6. Ascorbic Acid and Apoptosis
7. Ascorbic Acid Cooperation with Other Antioxidants
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Moss, R.W. Free Radical: Albert Szent-Gyorgyi and the Battle over Vitamin C. J. Hist. Biol. 1989, 22, 180–181. [Google Scholar]
- Davey, M.W.; Montagu, M.V.; Inz, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.; Strain, J.J.; Favell, D.; Fletcher, J. PlantL-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Nishikimi, M.; Yagi, K. Molecular basis for the deficiency in humans of gulonolactone oxidase, a key enzyme for ascorbic acid biosynthesis. Am. J. Clin. Nutr. 1991, 54, 1203S–1208S. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US) Panel on Dietary Antioxidants and Related Compounds. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academies Press (US): Washington, DC, USA, 2000. [Google Scholar]
- Mazurek, A.; Pankiewicz, U. Changes of dehydroascorbic acid content in relation to total content of vitamin C in selected fruits and vegetables. Acta Sci. Pol. Hortorum Cultus 2012, 6, 169–177. [Google Scholar]
- Valente, A.; Albuquerque, T.G.; Sanches-Silva, A.; Costa, H.S. Ascorbic acid content in exotic fruits: A contribution to produce quality data for food composition databases. Food Res. Int. 2011, 44, 2237–2242. [Google Scholar] [CrossRef]
- Vanderslice, J.T.; Higgs, D.J.; Hayes, J.M.; Block, G. Ascorbic acid and dehydroascorbic acid content of foods-as-eaten. J. Food Compos. Anal. 1990, 3, 105–118. [Google Scholar] [CrossRef]
- Hill, C.H. 20 Foods That Are High in Vitamin C. Available online: https://www.healthline.com/nutrition/vitamin-c-foods#TOC_TITLE_HDR_2 (accessed on 12 March 2022).
- García-Closas, R.; Berenguer, A.; Tormo, M.J.; Sánchez, M.J.; Quirós, J.R.; Navarro, C.; Arnaud, R.; Dorronsoro, M.; Chirlaque, M.D.; Barricarte, A.; et al. Dietary sources of vitamin C, vitamin E and specific carotenoids in Spain. Br. J. Nutr. 2004, 91, 1005–1011. [Google Scholar] [CrossRef]
- Douglas, R.; Hemilä, H.; Chalker, E.; D’Souza, R.; Treacy, B. Vitamin C for preventing and treating the common cold. In Cochrane Database of Systematic Reviews; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2004. [Google Scholar]
- U.S. Department of Agriculture, Agricultural Research Service. FoodData Central; 2019. Available online: fdc.nal.usda.gov (accessed on 1 October 2022).
- Levine, M.; Conry-Cantilena, C.; Wang, Y.; Welch, R.W.; Washko, P.W.; Dhariwal, K.R.; Park, J.B.; Lazarev, A.; Graumlich, J.F.; King, J.; et al. Vitamin C pharmacokinetics in healthy volunteers: Evidence for a recommended dietary allowance. Proc. Natl. Acad. Sci. USA 1996, 93, 3704–3709. [Google Scholar] [CrossRef]
- Pinnell, S.R.; Yang, H.; Omar, M.; Riviere, N.M.; DeBuys, H.V.; Walker, L.C.; Wang, Y.; Levine, M. Topical L-Ascorbic Acid: Percutaneous Absorption Studies. Dermatol. Surg. 2001, 27, 137–142. [Google Scholar] [CrossRef]
- Rose, R.C. Transport of ascorbic acid and other water-soluble vitamins. BBA–Rev. Biomembr. 1988, 947, 335–366. [Google Scholar] [CrossRef]
- Wilson, J.X. Regulation of vitamin C transport. Annu. Rev. Nutr. 2005, 25, 105–125. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, L.; Thumser, A.E.; Sharp, P. Decreased expression of the vitamin C transporter SVCT1 by ascorbic acid in a human intestinal epithelial cell line. Br. J. Nutr. 2002, 87, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Ströhle, A.; Hahn, A. Vitamin C und immunfunktion. Med. Monatsschr. Pharm. 2009, 32, 49–54. [Google Scholar]
- May, J.M.; Harrison, F.E. Role of vitamin C in the function of the vascular endothelium. Antioxid. Redox Signal. 2013, 19, 2068–2083. [Google Scholar] [CrossRef] [PubMed]
- Crisan, D.; Roman, I.; Crisan, M.; Scharffetter-Kochanek, K.; Badea, R. The role of vitamin C in pushing back the boundaries of skin aging: An ultrasonographic approach. Clin. Cosmet. Investig. Dermatol. 2015, 8, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Juraschek, S.P.; Guallar, E.; Appel, L.J.; Miller, E.R. Effects of vitamin C supplementation on blood pressure: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 95, 1079–1088. [Google Scholar] [CrossRef]
- Ang, A.; Pullar, J.M.; Currie, M.J.; Vissers, M.C.M. Vitamin C and immune cell function in inflammation and cancer. Biochem. Soc. Trans. 2018, 46, 1147–1159. [Google Scholar] [CrossRef]
- Langlois, M.; Duprez, D.; Delanghe, J.; De Buyzere, M.; Clement, D.L. Serum vitamin C concentration is low in peripheral arterial disease and is associated with inflammation and severity of atherosclerosis. Circulation 2001, 103, 1863–1868. [Google Scholar] [CrossRef]
- Kodentsova, V.M. Gradation in the level of vitamin consumption: Possible risk of excessive consumption. Vopr. Pitan. 2014, 83, 41–51. [Google Scholar]
- Lamarche, J.; Nair, R.; Peguero, A.; Courville, C. Vitamin C-Induced Oxalate Nephropathy. Int. J. Nephrol. 2011, 2011, 146927. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Hille, R.; Nishino, T. Xanthine oxidase and xanthine dehydrogenase. FASEB J. 1995, 9, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Nishikimi, M. Oxidation of ascorbic acid with superoxide anion generated by the xanthine-xanthine oxidase system. Biochem. Biophys. Res. Commun. 1975, 63, 463–468. [Google Scholar] [CrossRef]
- Francis, A.J.; Anderson, D.; Jenkinson, P.C.; Parke, D.V.W. The protective effects of L-ascorbic acid and DL-α-tocopherol on cultured rat embryos treated with xanthine/xanthine oxidase. Mutat. Res.–Fundam. Mol. Mech. Mutagen. 1989, 214, 137–145. [Google Scholar] [CrossRef]
- Alghamdi, A.A.; Althumali, J.S.; Almalki, M.M.M.; Almasoudi, A.S.; Almuntashiri, A.H.; Almuntashiri, A.H.; Mohammed, A.I.; Alkinani, A.A.; Almahdawi, M.S.; Mahzari, M.A.H. An Overview on the Role of Xanthine Oxidase Inhibitors in Gout Management. Arch. Pharm. Pract. 2021, 12, 94–99. [Google Scholar] [CrossRef]
- Linani, A.; Benarous, K.; Bou-Salah, L.; Yousfi, M. The inhibitory kinetics of vitamins B9, C, E, and D3 on bovine xanthine oxidase: Gout treatment. Chem. Biol. Interact. 2022, 359, 109922. [Google Scholar] [CrossRef] [PubMed]
- Moonrungsee, N.; Jakmunee, J.; Peamaroon, N.; Boonmee, A.; Kasemsuk, T.; Seeda, S.; Suwancharoen, S. Phytochemical and Xanthine Oxidase Inhibitory Activity in Nypa fruticans Wurmb. Fruit Extracts. Trends Sci. 2022, 19, 2583. [Google Scholar] [CrossRef]
- Li, H.; Liu, X.; Lee, M.; Li, H. Vitamin C alleviates hyperuricemia nephropathy by reducing inflammation and fibrosis. J. Food Sci. 2021, 86, 3265–3276. [Google Scholar] [CrossRef]
- Gęgotek, A.; Ambrożewicz, E.; Jastrząb, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Rutin and ascorbic acid cooperation in antioxidant and antiapoptotic effect on human skin keratinocytes and fibroblasts exposed to UVA and UVB radiation. Arch. Dermatol. Res. 2019, 311, 203–219. [Google Scholar] [CrossRef]
- Gęgotek, A.; Bielawska, K.; Biernacki, M.; Zaręba, I.; Surażyński, A.; Skrzydlewska, E. Comparison of protective effect of ascorbic acid on redox and endocannabinoid systems interactions in in vitro cultured human skin fibroblasts exposed to UV radiation and hydrogen peroxide. Arch. Dermatol. Res. 2017, 309, 285–303. [Google Scholar] [CrossRef]
- Dwenger, A.; Funck, M.; Lueken, B.; Schweizer, G.; Lehmann, U. Effect of Ascorbic Acid on Neutrophil Functions and Hypoxanthine/Xanthine Oxidase-Generated, Oxygen-Derived Radicals. Clin. Chem. Lab. Med. 1992, 30, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef]
- Cobley, J.N.; McHardy, H.; Morton, J.P.; Nikolaidis, M.G.; Close, G.L. Influence of vitamin C and vitamin e on redox signaling: Implications for exercise adaptations. Free Radic. Biol. Med. 2015, 84, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Schuster, D.P.; Tyml, K.; Wilson, J.X. Ascorbate inhibits NADPH oxidase subunit p47phox expression in microvascular endothelial cells. Free Radic. Biol. Med. 2007, 42, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.X. Mechanism of action of vitamin C in sepsis: Ascorbate modulates redox signaling in endothelium. BioFactors 2009, 35, 5–13. [Google Scholar] [CrossRef]
- Bartsch, C.; Bekhite, M.M.; Wolheim, A.; Richter, M.; Ruhe, C.; Wissuwa, B.; Marciniak, A.; Müller, J.; Heller, R.; Figulla, H.R.; et al. NADPH oxidase and eNOS control cardiomyogenesis in mouse embryonic stem cells on ascorbic acid treatment. Free Radic. Biol. Med. 2011, 51, 432–443. [Google Scholar] [CrossRef]
- Li, X.; Cobb, C.E.; May, J.M. Mitochondrial recycling of ascorbic acid from dehydroascorbic acid: Dependence on the electron transport chain. Arch. Biochem. Biophys. 2002, 403, 103–110. [Google Scholar] [CrossRef]
- Martinovich, G.G.; Golubeva, E.N.; Martinovich, I.V.; Cherenkevich, S.N. Redox Regulation of Calcium Signaling in Cancer Cells by Ascorbic Acid Involving the Mitochondrial Electron Transport Chain. J. Biophys. 2012, 2012, 921653. [Google Scholar] [CrossRef]
- Sharma, P.; Morgan, P.D. Ascorbate reduces superoxide production and improves mitochondrial respiratory chain function in human fibroblasts with electron transport chain deficiencies. Mitochondrion 2001, 1, 191–198. [Google Scholar] [CrossRef]
- Bakalova, R.; Zhelev, Z.; Miller, T.; Aoki, I.; Higashi, T. Vitamin C versus Cancer: Ascorbic Acid Radical and Impairment of Mitochondrial Respiration? Oxid. Med. Cell. Longev. 2020, 2020, 1504048. [Google Scholar] [CrossRef]
- Tariq, S.A. Role of ascorbic acid in scavenging free radicals and lead toxicity from biosystems. Mol. Biotechnol. 2007, 37, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, I.P.; Trofimova, S.V.; Piskarev, I.M. Evaluation of prooxidant properties of ascorbic acid. Biophysics 2013, 58, 453–456. [Google Scholar] [CrossRef]
- Yen, G.C.; Duh, P.D.; Tsai, H.L. Antioxidant and pro-oxidant properties of ascorbic acid and gallic acid. Food Chem. 2002, 79, 307–313. [Google Scholar] [CrossRef]
- Du, J.; Wagner, B.A.; Buettner, G.R.; Cullen, J.J. Role of labile iron in the toxicity of pharmacological ascorbate. Free Radic. Biol. Med. 2015, 84, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Weber, S.U.; Rimbach, G. Molecular aspects of α-tocotrienol antioxidant action and cell signalling. J. Nutr. 2001, 131, 369S–373S. [Google Scholar] [CrossRef]
- Peake, J.M. Vitamin C: Effects of exercise and requirements with training. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 125–151. [Google Scholar] [CrossRef]
- Li, X.; Qu, Z.C.; May, J.M. GSH is required to recycle ascorbic acid in cultured liver cell Lines. Antioxid. Redox Signal. 2001, 3, 1089–1097. [Google Scholar] [CrossRef]
- Sünder, A.; Flachowsky, G. Influence of high vitmin E dosages on retinol and carotinoid concentration in body tissues and eggs of laying hens. Arch. Anim. Nutr. 2001, 55, 43–52. [Google Scholar]
- Young, A.M.; Gregoriadis, G. Photolysis of Retinol in Liposomes and Its Protection with Tocopherol and Oxybenzone. Photochem. Photobiol. 1996, 63, 344–352. [Google Scholar] [CrossRef]
- Beyer, R.E. The role of ascorbate in antioxidant protection of biomembranes: Interaction with vitamin E and coenzyme Q. J. Bioenerg. Biomembr. 1994, 26, 349–358. [Google Scholar] [CrossRef]
- Nemati, M.; Shahir, M.; Harakinezhad, M.; Lotfalhian, H. Cold-Induced Ascites in Broilers: Effects of Vitamin C and Coenzyme Q10. Rev. Bras. Ciência Avícola 2017, 19, 537–544. [Google Scholar] [CrossRef]
- Shargorodsky, M.; Debby, O.; Matas, Z.; Zimlichman, R. Effect of long-term treatment with antioxidants (vitamin C, vitamin E, coenzyme Q10 and selenium) on arterial compliance, humoral factors and inflammatory markers in patients with multiple cardiovascular risk factors. Nutr. Metab. 2010, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Kobori, Y.; Ota, S.; Sato, R.; Yagi, H.; Soh, S.; Arai, G.; Okada, H. Antioxidant cosupplementation therapy with vitamin C, vitamin E, and coenzyme Q10 in patients with oligoasthenozoospermia. Arch. Ital. Urol. Androl. 2014, 86, 1–4. [Google Scholar] [CrossRef]
- Pavlović, S.Z.; Ognjanović, B.I.; Štajn, A.Š.; Źikić, R.V.; Saičić, Z.S.; Petrović, V.M. The effect of coenzyme Q10 on blood ascorbic acid, vitamin E, and lipid peroxide in chronic cadmium intoxication. J. Environ. Pathol. Toxicol. Oncol. 2001, 20, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Moretti, M.; Budni, J.; Dos Santos, D.B.; Antunes, A.; Daufenbach, J.F.; Manosso, L.M.; Farina, M.; Rodrigues, A.L.S. Protective effects of ascorbic acid on behavior and oxidative status of restraint-stressed mice. J. Mol. Neurosci. 2013, 49, 68–79. [Google Scholar] [CrossRef]
- Muthuvel, R.; Venkataraman, P.; Krishnamoorthy, G.; Gunadharini, D.N.; Kanagaraj, P.; Jone Stanley, A.; Srinivasan, N.; Balasubramanian, K.; Aruldhas, M.M.; Arunakaran, J. Antioxidant effect of ascorbic acid on PCB (Aroclor 1254) induced oxidative stress in hypothalamus of albino rats. Clin. Chim. Acta 2006, 365, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Adikwu, E.; Deo, O. Hepatoprotective Effect of Vitamin C (Ascorbic Acid). Pharmacol. Pharm. 2013, 4, 84–92. [Google Scholar] [CrossRef]
- Koc, M.; Imik, H.; Odabasoglu, F. Gastroprotective and anti-oxidative properties of ascorbic acid on indomethacin-induced gastric injuries in rats. Biol. Trace Elem. Res. 2008, 126, 222–236. [Google Scholar] [CrossRef]
- Hu, J.H.; Tian, W.Q.; Zhao, X.L.; Zan, L.S.; Wang, H.; Li, Q.W.; Xin, Y.P. The cryoprotective effects of ascorbic acid supplementation on bovine semen quality. Anim. Reprod. Sci. 2010, 121, 72–77. [Google Scholar] [CrossRef]
- Krishnan, N.; Kodrík, D.; Kłudkiewicz, B.; Sehnal, F. Glutathione-ascorbic acid redox cycle and thioredoxin reductase activity in the digestive tract of Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 2009, 39, 180–188. [Google Scholar] [CrossRef]
- Choe, J.Y.; Kim, S.K. Quercetin and Ascorbic Acid Suppress Fructose-Induced NLRP3 Inflammasome Activation by Blocking Intracellular Shuttling of TXNIP in Human Macrophage Cell Lines. Inflammation 2017, 40, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Yang, J.; Ji, S.; Li, T.; Gao, S.; Sun, C.; Xu, H. Cancer therapy by targeting thioredoxin reductase based on selenium-containing dynamic covalent bond. CCS Chem. 2020, 2, 225–235. [Google Scholar] [CrossRef]
- Ishaq, G.M.; Saidu, Y.; Bilbis, L.S.; Muhammad, S.A.; Jinjir, N.; Shehu, B.B. Effects of α-tocopherol and ascorbic acid in the severity and management of traumatic brain injury in albino rats. J. Neurosci. Rural Pract. 2013, 4, 292–297. [Google Scholar] [CrossRef]
- Erdogan, Z.; Erdogan, S.; Celik, S.; Unlu, A. Effects of ascorbic acid on cadmium-induced oxidative stress and performance of broilers. Biol. Trace Elem. Res. 2005, 104, 19–31. [Google Scholar] [CrossRef]
- May, J.M.; Qu, Z.C.; Meredith, M.E. Mechanisms of ascorbic acid stimulation of norepinephrine synthesis in neuronal cells. Biochem. Biophys. Res. Commun. 2012, 426, 148–152. [Google Scholar] [CrossRef]
- Saffi, J.; Sonego, L.; Varela, Q.D.; Salvador, M. Antioxidant activity of L-ascorbic acid in wild-type and superoxide dismutase deficient strains of Saccharomyces cerevisiae. Redox Rep. 2006, 11, 179–184. [Google Scholar] [CrossRef]
- Tamari, Y.; Nawata, H.; Inoue, E.; Yoshimura, A.; Yoshii, H.; Kashino, G.; Seki, M.; Enomoto, T.; Watanabe, M.; Tano, K. Protective roles of ascorbic acid in oxidative stress induced by depletion of superoxide dismutase in vertebrate cells. Free Radic. Res. 2013, 47, 1–7. [Google Scholar] [CrossRef]
- Washio, K.; Inagaki, M.; Tsuji, M.; Morio, Y.; Akiyama, S.; Gotoh, H.; Gotoh, T.; Gotoh, Y.; Oguchi, K. Oral Vitamin C Supplementation in Hemodialysis Patients and Its Effect on the Plasma Level of Oxidized Ascorbic Acid and Cu/Zn Superoxide Dismutase, an Oxidative Stress Marker. Nephron Clin. Pract. 2008, 109, c49–c54. [Google Scholar] [CrossRef]
- Botanicae Horti Agrobotanici Cluj-Napoca, N.; Dolatabadian, A.; Ali Mohammad Modarres Sanavy, S. Effect of the Ascorbic Acid, Pyridoxine and Hydrogen Peroxide Treatments on Germination, Catalase Activity, Protein and Malondialdehyde Content of Three Oil Seeds. Hort. Agrobot. Cluj 2008, 36, 61–66. [Google Scholar]
- Sun, Y.; Zheng, Q.; LI, G.; Guo, D.; Wang, Z. Mechanism of ascorbic acid-induced reversion against malignant phenotype in human gastric cancer cells. Biomed. Environ. Sci. 2006, 19, 385–391. [Google Scholar]
- Klingelhoeffer, C.; Kämmerer, U.; Koospal, M.; Mühling, B.; Schneider, M.; Kapp, M.; Kübler, A.; Germer, C.T.; Otto, C. Natural resistance to ascorbic acid induced oxidative stress is mainly mediated by catalase activity in human cancer cells and catalase-silencing sensitizes to oxidative stress. BMC Complement. Altern. Med. 2012, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Skrzydlewska, E. The role of transcription factor Nrf2 in skin cells metabolism. Arch. Dermatol. Res. 2015, 307, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.J.; Jiang, W.D.; Feng, L.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.Y.; Tang, L.; Tang, W.N.; Zhang, Y.A.; et al. Dietary vitamin C deficiency depresses the growth, head kidney and spleen immunity and structural integrity by regulating NF-κB, TOR, Nrf2, apoptosis and MLCK signaling in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2016, 52, 111–138. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.J.; Jiang, W.D.; Feng, L.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.Y.; Tang, L.; Tang, W.N.; Zhang, Y.A.; et al. Dietary vitamin C deficiency depressed the gill physical barriers and immune barriers referring to Nrf2, apoptosis, MLCK, NF-κB and TOR signaling in grass carp (Ctenopharyngodon idella) under infection of Flavobacterium columnare. Fish Shellfish Immunol. 2016, 58, 177–192. [Google Scholar] [CrossRef]
- Vineetha, R.C.; Binu, P.; Arathi, P.; Nair, R.H. L-ascorbic acid and α-tocopherol attenuate arsenic trioxide-induced toxicity in H9c2 cardiomyocytes by the activation of Nrf2 and Bcl2 transcription factors. Toxicol. Mech. Methods 2018, 28, 353–360. [Google Scholar] [CrossRef]
- Li, Y.; Darwish, W.S.; Chen, Z.; Hui, T.; Wu, Y.; Hirotaka, S.; Chiba, H.; Hui, S.P. Identification of lead-produced lipid hydroperoxides in human HepG2 cells and protection using rosmarinic and ascorbic acids with a reference to their regulatory roles on Nrf2-Keap1 antioxidant pathway. Chem. Biol. Interact. 2019, 314, 108847. [Google Scholar] [CrossRef]
- Wagner, A.E.; Ernst, I.; Iori, R.; Desel, C.; Rimbach, G. Sulforaphane but not ascorbigen, indole-3-carbinole and ascorbic acid activates the transcription factor Nrf2 and induces phase-2 and antioxidant enzymes in human keratinocytes in culture. Exp. Dermatol. 2010, 19, 137–144. [Google Scholar] [CrossRef]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Cytoprotective Effect of Ascorbic Acid and Rutin against Oxidative Changes in the Proteome of Skin Fibroblasts Cultured in a Three-Dimensional System. Nutrients 2020, 12, 1074. [Google Scholar] [CrossRef]
- Wagner, A.E.; Boesch-Saadatmandi, C.; Breckwoldt, D.; Schrader, C.; Schmelzer, C.; Döring, F.; Hashida, K.; Hori, O.; Matsugo, S.; Rimbach, G. Ascorbic acid partly antagonizes resveratrol mediated heme oxygenase-1 but not paraoxonase-1 induction in cultured hepatocytes—Role of the redox-regulated transcription factor Nrf2. BMC Complement. Altern. Med. 2011, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi-Pour, Z.; Ramezani, F.; Keshavarzi, F.; Samadi, N. The role of quercetin and vitamin c in NRF2-dependent oxidative stress production in breast cancer cells. Oncol. Lett. 2017, 13, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, A.; Mostafavi-Pour, Z.; Amiri, A.; Keshavarzi, F.; Nejabat, N.; Ramezani, F.; Sardarian, A.; Zal, F. Chemoprevention of Prostate Cancer Cells by Vitamin C plus Quercetin: Role of Nrf2 in Inducing Oxidative Stress. Nutr. Cancer 2021, 73, 2003–2013. [Google Scholar] [CrossRef] [PubMed]
- Xanthoudakis, S.; Curran, T. Identification and characterization of Ref-1, a nuclear protein that facilitates AP-1 DNA-binding activity. EMBO J. 1992, 11, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.; Angel, P.; Schorpp-Kistner, M. AP-1 subunits: Quarrel and harmony among siblings. J. Cell Sci. 2004, 117, 5965–5973. [Google Scholar] [CrossRef]
- Huang, C.; Ma, W.; Dong, Z. Inhibitory effects of ascorbic acid on AP-1 activity and transformation of JB6 cells. Int. J. Oncol. 1996, 8, 389–393. [Google Scholar] [CrossRef]
- Catani, M.V.; Savini, I.; Rossi, A.; Melino, G.; Avigliano, L. Biological Role of Vitamin C in Keratinocytes. Nutr. Rev. 2005, 63, 81–90. [Google Scholar] [CrossRef]
- Lee, A.J.; Lim, J.W.; Kim, H. Ascorbic Acid Suppresses House Dust Mite-Induced Expression of Interleukin-8 in Human Respiratory Epithelial Cells. J. Cancer Prev. 2021, 26, 64–70. [Google Scholar] [CrossRef]
- Lu, A.L.; Li, X.; Gu, Y.; Wright, P.M.; Chang, D.Y. Repair of oxidative DNA damage: Mechanisms and functions. Cell Biochem. Biophys. 2001, 35, 141–170. [Google Scholar] [CrossRef]
- Lan, L.; Nakajima, S.; Oohata, Y.; Takao, M.; Okano, S.; Masutani, M.; Wilson, S.H.; Yasui, A. In situ analysis of repair processes for oxidative DNA damage in mammalian cells. Proc. Natl. Acad. Sci. USA 2004, 101, 13738–13743. [Google Scholar] [CrossRef] [PubMed]
- Lutsenko, E.A.; Cárcamo, J.M.; Golde, D.W. Vitamin C prevents DNA mutation induced by oxidative stress. J. Biol. Chem. 2002, 277, 16895–16899. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Y.; Häder, D.P. UV-B-induced formation of reactive oxygen species and oxidative damage of the cyanobacterium Anabaena sp.: Protective effects of ascorbic acid and N-acetyl-L-cysteine. J. Photochem. Photobiol. B Biol. 2002, 66, 115–124. [Google Scholar] [CrossRef]
- Yin, R.; Mao, S.Q.; Zhao, B.; Chong, Z.; Yang, Y.; Zhao, C.; Zhang, D.; Huang, H.; Gao, J.; Li, Z.; et al. Ascorbic acid enhances tet-mediated 5-methylcytosine oxidation and promotes DNA demethylation in mammals. J. Am. Chem. Soc. 2013, 135, 10396–10403. [Google Scholar] [CrossRef]
- Blaschke, K.; Ebata, K.T.; Karimi, M.M.; Zepeda-Martínez, J.A.; Goyal, P.; Mahapatra, S.; Tam, A.; Laird, D.J.; Hirst, M.; Rao, A.; et al. Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature 2013, 500, 222–226. [Google Scholar] [CrossRef]
- Shenoy, N.; Bhagat, T.; Nieves, E.; Stenson, M.; Lawson, J.; Choudhary, G.S.; Habermann, T.; Nowakowski, G.; Singh, R.; Wu, X.; et al. Upregulation of TET activity with ascorbic acid induces epigenetic modulation of lymphoma cells. Blood Cancer J. 2017, 7, e587. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, N.; Bhagat, T.D.; Cheville, J.; Lohse, C.; Bhattacharyya, S.; Tischer, A.; Machha, V.; Gordon-Mitchell, S.; Choudhary, G.; Wong, L.F.; et al. Ascorbic acid–induced TET activation mitigates adverse hydroxymethylcytosine loss in renal cell carcinoma. J. Clin. Investig. 2019, 129, 1612–1625. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.P.; Miles, S.L. Silencing HIF-1α induces TET2 expression and augments ascorbic acid induced 5-hydroxymethylation of DNA in human metastatic melanoma cells. Biochem. Biophys. Res. Commun. 2017, 490, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R.; Levine, R.L. Protein Oxidation. Ann. N. Y. Acad. Sci. 2006, 899, 191–208. [Google Scholar] [CrossRef]
- Friguet, B. Oxidized protein degradation and repair in ageing and oxidative stress. FEBS Lett. 2006, 580, 2910–2916. [Google Scholar] [CrossRef]
- Perrone, G.; Hideshima, T.; Ikeda, H.; Okawa, Y.; Calabrese, E.; Gorgun, G.; Santo, L.; Cirstea, D.; Raje, N.; Chauhan, D.; et al. Ascorbic acid inhibits antitumor activity of bortezomib in vivo. Leukemia 2009, 23, 1679–1686. [Google Scholar] [CrossRef]
- Zou, W.; Yue, P.; Lin, N.; He, M.; Zhou, Z.; Lonial, S.; Khuri, F.R.; Wang, B.; Sun, S.-Y.Y. Vitamin C inactivates the proteasome inhibitor PS-341 in human cancer cells. AACR 2006, 12, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Nauser, T.; Gebicki, J.M. Physiological concentrations of ascorbate cannot prevent the potentially damaging reactions of protein radicals in humans. Chem. Res. Toxicol. 2017, 30, 1702–1710. [Google Scholar] [CrossRef] [PubMed]
- Domazou, A.S.; Koppenol, W.H.; Gebicki, J.M. Efficient repair of protein radicals by ascorbate. Free Radic. Biol. Med. 2009, 46, 1049–1057. [Google Scholar] [CrossRef]
- Gebicki, J.M.; Nauser, T. Initiation and prevention of biological damage by radiation-generated protein radicals. Int. J. Mol. Sci. 2022, 23, 396. [Google Scholar] [CrossRef] [PubMed]
- Guéraud, F.; Atalay, M.; Bresgen, N.; Cipak, A.; Eckl, P.M.; Huc, L.; Jouanin, I.; Siems, W.; Uchida, K. Chemistry and biochemistry of lipid peroxidation products. Free Radic. Res. 2010, 44, 1098–1124. [Google Scholar] [CrossRef]
- Ramana, K.V.; Srivastava, S.; Singhal, S.S. Lipid peroxidation products in human health and disease. Oxid. Med. Cell. Longev. 2013, 2013, 583438. [Google Scholar] [CrossRef]
- Ranjan, R.; Swarup, D.; Naresh, R.; Patra, R.C. Enhanced erythrocytic lipid peroxides and reduced plasma ascorbic acid, and alteration in blood trace elements level in dairy cows with mastitis. Vet. Res. Commun. 2005, 29, 27–34. [Google Scholar] [CrossRef]
- Marchlewicz, M.; Wiszniewska, B.; Gonet, B.; Baranowska-Bosiacka, I.; Safranow, K.; Kolasa, A.; Głąbowski, W.; Kurzawa, R.; Jakubowska, K.; Rać, M.E. Increased lipid peroxidation and ascorbic acid utilization in testis and epididymis of rats chronically exposed to lead. BioMetals 2007, 20, 13–19. [Google Scholar] [CrossRef]
- Surapaneni, K.; Venkataramana, G. Status of lipid peroxidation, glutathione, ascorbic acid, vitamin E and antioxidant enzymes in patients with osteoarthritis. Indian J. Med. Sci. 2007, 61, 9–14. [Google Scholar] [CrossRef]
- Sönmez, M.; Türk, G.; Yüce, A. The effect of ascorbic acid supplementation on sperm quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology 2005, 63, 2063–2072. [Google Scholar] [CrossRef]
- Krishna Mohan, S.; Venkataramana, G. Status of lipid peroxidation, glutathione, ascorbic acid, vitamin E and antioxidant enzymes in patients with pregnancy—Induced hypertension. Indian J. Physiol. Pharmacol. 2007, 51, 284–288. [Google Scholar] [PubMed]
- Chen, W.; Guo, J.; Guo, H.; Kong, X.; Bai, J.; Long, P. Protective Effect of Vitamin C against Infancy Rat Corneal Injury Caused by Acute UVB Irradiation. Biomed Res. Int. 2020, 2020, 8089273. [Google Scholar] [CrossRef] [PubMed]
- Akbulut, S.; Elbe, H.; Eris, C.; Dogan, Z.; Toprak, G.; Otan, E.; Erdemli, E.; Turkoz, Y. Cytoprotective effects of amifostine, ascorbic acid and N-acetylcysteine against methotrexate-induced hepatotoxicity in rats. World J. Gastroenterol. 2014, 20, 10158–10165. [Google Scholar] [CrossRef] [PubMed]
- Raina, R.; Baba, N.A.; Verma, P.K.; Sultana, M.; Singh, M. Hepatotoxicity induced by subchronic exposure of fluoride and chlorpyrifos in Wistar rats: Mitigating effect of ascorbic acid. Biol. Trace Elem. Res. 2015, 166, 157–162. [Google Scholar] [CrossRef]
- Ray, S.; Roy, K.; Sengupta, C. Cisplatin-induced lipid peroxidation and its inhibition with ascorbic acid. Indian J. Pharm. Sci. 2006, 68, 199–204. [Google Scholar]
- Hernández-Guerra, M.; García-Pagán, J.C.; Turnes, J.; Bellot, P.; Deulofeu, R.; Abraldes, J.G.; Bosch, J. Ascorbic acid improves the intrahepatic endothelial dysfunction of patients with cirrhosis and portal hypertension. Hepatology 2006, 43, 485–491. [Google Scholar] [CrossRef]
- Das, K.K.; Gupta, A.D.; Dhundasi, S.A.; Patil, A.M.; Das, S.N.; Ambekar, J.G. Protective role of L-ascorbic acid on antioxidant defense system in erythrocytes of albino rats exposed to nickel sulfate. BioMetals 2007, 20, 177–184. [Google Scholar] [CrossRef]
- Tonon, E.; Ferretti, R.; Shiratori, J.H.; Santo Neto, H.; Marques, M.J.; Minatel, E. Ascorbic acid protects the diaphragm muscle against myonecrosis in mdx mice. Nutrition 2012, 28, 686–690. [Google Scholar] [CrossRef]
- Mason, S.A.; Rasmussen, B.; van Loon, L.J.C.; Salmon, J.; Wadley, G.D. Ascorbic acid supplementation improves postprandial glycaemic control and blood pressure in individuals with type 2 diabetes: Findings of a randomized cross-over trial. Diabetes Obes. Metab. 2019, 21, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Dohi, Y.; Kojima, M.; Miyagawa, K.; Takase, H.; Katada, E.; Suzuki, S. Effects of Ascorbic Acid on Ambulatory Blood Pressure in Elderly Patients with Refractory Hypertension. Arzneimittelforschung 2011, 56, 535–540. [Google Scholar] [CrossRef]
- Jialal, I.; Singh, U. Is vitamin C an antiinflammatory agent? Am. J. Clin. Nutr. 2006, 83, 525–526. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wannamethee, S.G.; Lowe, G.D.O.; Rumley, A.; Bruckdorfer, K.R.; Whincup, P.H. Associations of vitamin C status, fruit and vegetable intakes, and markers of inflammation and hemostasis. Am. J. Clin. Nutr. 2006, 83, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Colby, J.A.; Chen, W.T.; Baker, W.L.; Coleman, C.I.; Reinhart, K.; Kluger, J.; White, C.M. Effect of ascorbic acid on inflammatory markers after cardiothoracic surgery. Am. J. Health Pharm. 2011, 68, 1632–1639. [Google Scholar] [CrossRef]
- García-Bailo, B.; Roke, K.; Mutch, D.M.; El-Sohemy, A.; Badawi, A. Association between circulating ascorbic acid, -tocopherol, 25-hydroxyvitamin D, and plasma cytokine concentrations in young adults: A cross-sectional study. Nutr. Metab. 2012, 9, 102. [Google Scholar] [CrossRef]
- de Oliveira, B.F.; Veloso, C.A.; Nogueira-Machado, J.A.; de Moraes, E.N.; dos Santos, R.R.; Cintra, M.T.G.; Chaves, M.M. Ascorbic acid, alpha-tocopherol, and betacarotene reduce oxidative stress and proinflammatory cytokines in mononuclear cells of Alzheimer’s disease patients. Nutr. Neurosci. 2012, 15, 244–251. [Google Scholar] [CrossRef]
- Jouybar, R.; Kabgani, H.; Hamid, K.; Shahbazi, S.; Allahyary, E.; Rasouli, M.; Akhlagh, S.H.; Shafa, M.; Ghazinoor, M.; Moeinvaziri, M.T.; et al. The Perioperative Effect of Ascorbic Acid on Inflammatory Response in Coronary Artery Bypass Graft Surgery. Cardiovasc. Res. J. 2012, 6, 13–17. [Google Scholar]
- Tan, P.H.; Sagoo, P.; Chan, C.; Yates, J.B.; Campbell, J.; Beutelspacher, S.C.; Foxwell, B.M.J.; Lombardi, G.; George, A.J.T. Inhibition of NF-κB and Oxidative Pathways in Human Dendritic Cells by Antioxidative Vitamins Generates Regulatory T Cells. J. Immunol. 2005, 174, 7633–7644. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. WIREs Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef]
- Peng, Y.; Kwok, K.H.H.; Yang, P.H.; Ng, S.S.M.; Liu, J.; Wong, O.G.; He, M.L.; Kung, H.F.; Lin, M.C.M. Ascorbic acid inhibits ROS production, NF-κB activation and prevents ethanol-induced growth retardation and microencephaly. Neuropharmacology 2005, 48, 426–434. [Google Scholar] [CrossRef]
- Gegotek, A.; Biernacki, M.; Ambrozewicz, E.; Surazyński, A.; Wroński, A.; Skrzydlewska, E. The cross-talk between electrophiles, antioxidant defence and the endocannabinoid system in fibroblasts and keratinocytes after UVA and UVB irradiation. J. Dermatol. Sci. 2016, 81, 107–117. [Google Scholar] [CrossRef]
- Bowie, A.G.; O’Neill, L.A.J. Vitamin C Inhibits NF-κB Activation by TNF Via the Activation of p38 Mitogen-Activated Protein Kinase. J. Immunol. 2000, 165, 7180–7188. [Google Scholar] [CrossRef]
- Cárcamo, J.M.; Pedraza, A.; Bórquez-Ojeda, O.; Golde, D.W. Vitamin C suppresses TNFα-induced NFκB activation by inhibiting IκBα phosphorylation. Biochemistry 2002, 41, 12995–13002. [Google Scholar] [CrossRef]
- Han, S.-S.; Kim, K.; Hahm, E.-R.; Lee, S.J.; Surh, Y.-J.; Park, H.K.; Kim, W.S.; Jung, C.W.; Lee, M.H.; Park, K.; et al. L-ascorbic acid represses constitutive activation of NF-?B and COX-2 expression in human acute myeloid leukemia, HL-60. J. Cell. Biochem. 2004, 93, 257–270. [Google Scholar] [CrossRef]
- Alhusaini, A.M.; Fadda, L.M.; Alsharafi, H.; Alshamary, A.F.; Hasan, I.H. L-Ascorbic Acid and Curcumin Prevents Brain Damage Induced via Lead Acetate in Rats: Possible Mechanisms. Dev. Neurosci. 2022, 44, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.P.; Hiscock, N.J.; Penkowa, M.; Basu, S.; Vessby, B.; Kallner, A.; Sjöberg, L.-B.; Pedersen, B.K. Supplementation with vitamins C and E inhibits the release of interleukin-6 from contracting human skeletal muscle. J. Physiol. 2004, 558, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Kong, E.H.; Ma, S.Y.; Jeong, J.Y.; Kim, K.H. Effects of L-ascorbic acid on the production of pro-inflammatory and anti-inflammatory cytokines in C57BL/6 mouse splenocytes. Kosin Med. J. 2015, 30, 41–49. [Google Scholar] [CrossRef]
- Schwager, J.; Schulze, J. Modulation of interleukin production by ascorbic acid. Vet. Immunol. Immunopathol. 1998, 64, 45–57. [Google Scholar] [CrossRef]
- Schwager, J.; Schulze, J. Influence of ascorbic acid on the response to mitogens and interleukin production of porcine lymphocytes. Int. J. Vitam. Nutr. Res. 1997, 67, 10–16. [Google Scholar]
- Prabudi, M.O.; Siregar, M.F.G.; Nasution, I.P.A.; Ilyas, S. The Effect of Ascorbic Acid on Interleukin-10 and Tumor Necrosis Factor-α Cytokines in Rattus norvegicus with Endometritis. Open Access Maced. J. Med. Sci. 2021, 9, 798–801. [Google Scholar] [CrossRef]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NFκB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef]
- Tian, H.; Zhang, B.F.; Di, J.H.; Jiang, G.; Chen, F.F.; Li, H.Z.; Li, L.T.; Pei, D.S.; Zheng, J.N. Keap1: One stone kills three birds Nrf2, IKKβ and Bcl-2/Bcl-xL. Cancer Lett. 2012, 325, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Sorice, A.; Guerriero, E.; Capone, F.; Colonna, G.; Castello, G.; Costantini, S. Ascorbic Acid: Its Role in Immune System and Chronic Inflammation Diseases. Mini Rev. Med. Chem. 2014, 14, 444–452. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef]
- Chandra, J.; Samali, A.; Orrenius, S. Triggering and modulation of apoptosis by oxidative stress. Free Radic. Biol. Med. 2000, 29, 323–333. [Google Scholar] [CrossRef]
- Wenzel, U. Ascorbic acid suppresses drug-induced apoptosis in human colon cancer cells by scavenging mitochondrial superoxide anions. Carcinogenesis 2003, 25, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, K.; Anusuyadevi, M.; Shila, S.; Panneerselvam, C. Ascorbic acid and α-tocopherol as potent modulators of apoptosis on arsenic induced toxicity in rats. Toxicol. Lett. 2005, 156, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Abu Zeid, E.H.; Hussein, M.M.A.; Ali, H. Ascorbic acid protects male rat brain from oral potassium dichromate-induced oxdative DNA damage and apoptotic changes: The expression patterns of caspase-3, P 53, Bax, and Bcl-2 genes. Environ. Sci. Pollut. Res. 2018, 25, 13056–13066. [Google Scholar] [CrossRef]
- Park, S.; Han, S.S.; Park, C.H.; Hahm, E.R.; Lee, S.J.; Park, H.K.; Lee, S.H.; Kim, W.S.; Jung, C.W.; Park, K.; et al. L-Ascorbic acid induces apoptosis in acute myeloid leukemia cells via hydrogen peroxide-mediated mechanisms. Int. J. Biochem. Cell Biol. 2004, 36, 2180–2195. [Google Scholar] [CrossRef]
- Chen, X.Y.; Chen, Y.; Qu, C.J.; Pan, Z.H.; Qin, Y.; Zhang, X.; Liu, W.J.; Li, D.F.; Zheng, Q. Vitamin C induces human melanoma A375 cell apoptosis via Bax- And Bcl-2-mediated mitochondrial pathways. Oncol. Lett. 2019, 18, 3880–3886. [Google Scholar] [CrossRef]
- Jung, S.A.; Lee, D.H.; Moon, J.H.; Hong, S.W.; Shin, J.S.; Hwang, I.Y.; Shin, Y.J.; Kim, J.H.; Gong, E.Y.; Kim, S.M.; et al. L-Ascorbic acid can abrogate SVCT-2-dependent cetuximab resistance mediated by mutant KRAS in human colon cancer cells. Free Radic. Biol. Med. 2016, 95, 200–208. [Google Scholar] [CrossRef]
- Frajese, G.V.; Benvenuto, M.; Fantini, M.; Ambrosin, E.; Sacchetti, P.; Masuelli, L.; Giganti, M.G.; Modesti, A.; Bei, R. Potassium increases the antitumor effects of ascorbic acid in breast cancer cell lines in vitro. Oncol. Lett. 2016, 11, 4224–4234. [Google Scholar] [CrossRef] [PubMed]
- Bourbour, M.; Khayam, N.; Noorbazargan, H.; Tavakkoli Yaraki, M.; Asghari Lalami, Z.; Akbarzadeh, I.; Eshrati Yeganeh, F.; Dolatabadi, A.; Mirzaei Rad, F.; Tan, Y.N. Evaluation of anti-cancer and anti-metastatic effects of folate-PEGylated niosomes for co-delivery of letrozole and ascorbic acid on breast cancer cells. Mol. Syst. Des. Eng. 2022, 7, 1102–1118. [Google Scholar] [CrossRef]
- Wu, T.-M.; Liu, S.-T.; Chen, S.-Y.; Chen, G.-S.; Wu, C.-C.; Huang, S.-M. Mechanisms and Applications of the Anti-cancer Effect of Pharmacological Ascorbic Acid in Cervical Cancer Cells. Front. Oncol. 2020, 10, 1483. [Google Scholar] [CrossRef]
- Pires, A.S.; Marques, C.R.; Encarnação, J.C.; Abrantes, A.M.; Marques, I.A.; Laranjo, M.; Oliveira, R.; Casalta-Lopes, J.E.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; et al. Ascorbic Acid Chemosensitizes Colorectal Cancer Cells and Synergistically Inhibits Tumor Growth. Front. Physiol. 2018, 9, 911. [Google Scholar] [CrossRef] [PubMed]
- Putchala, M.C.; Ramani, P.; Sherlin, H.J.; Premkumar, P.; Natesan, A. Ascorbic acid and its pro-oxidant activity as a therapy for tumours of oral cavity-A systematic review. Arch. Oral Biol. 2013, 58, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Martin, S.M.; Levine, M.; Wagner, B.A.; Buettner, G.R.; Wang, S.H.; Taghiyev, A.F.; Du, C.; Knudson, C.M.; Cullen, J.J. Mechanisms of ascorbate-induced cytotoxicity in pancreatic cancer. Clin. Cancer Res. 2010, 16, 509–520. [Google Scholar] [CrossRef]
- Shenoy, N.; Creagan, E.; Witzig, T.; Levine, M. Ascorbic Acid in Cancer Treatment: Let the Phoenix Fly. Cancer Cell 2018, 34, 700–706. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; El-Ghoneimy, A. Synergistic protective effects of ceftriaxone and ascorbic acid against subacute deltamethrin-induced nephrotoxicity in rats. Ren. Fail. 2015, 37, 297–304. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M. Synergistic protective role of ceftriaxone and ascorbic acid against subacute diazinon-induced nephrotoxicity in rats. Cytotechnology 2016, 68, 279–289. [Google Scholar] [CrossRef]
- Naksuriya, O.; Okonogi, S. Comparison and combination effects on antioxidant power of curcumin with gallic acid, ascorbic acid, and xanthone. Drug Discov. Ther. 2015, 9, 136–141. [Google Scholar] [CrossRef]
- Enko, J.; Gliszczyńska-Świgło, A. Influence of the interactions between tea (Camellia sinensis) extracts and ascorbic acid on their antioxidant activity: Analysis with interaction indexes and isobolograms. Food Addit. Contam.—Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wu, J.X.; Tu, Y.Y. Synergistic effects of tea polyphenols and ascorbic acid on human lung adenocarcinoma SPC-A-1 cells. J. Zhejiang Univ. Sci. B 2010, 11, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Luca, A.; Alexa, T.; Dondaş, A.; Andron, G.; Bǎdescu, M.; Alexa, I.D.; Bohotin, C. Pain modulation by curcumin and ascorbic acid in mice. Rev. Med. Chir. Soc. Med. Nat. Iasi. 2014, 118, 346–351. [Google Scholar] [PubMed]
- Andarwulan, N.; Kurniasih, D.; Apriady, R.A.; Rahmat, H.; Roto, A.V.; Bolling, B.W. Polyphenols, carotenoids, and ascorbic acid in underutilized medicinal vegetables. J. Funct. Foods 2012, 4, 339–347. [Google Scholar] [CrossRef]
- Li, L.; Shao, J.; Zhu, X.; Zhou, G.; Xu, X. Effect of plant polyphenols and ascorbic acid on lipid oxidation, residual nitrite and N-nitrosamines formation in dry-cured sausage. Int. J. Food Sci. Technol. 2013, 48, 1157–1164. [Google Scholar] [CrossRef]
- Gęgotek, A.; Domingues, P.; Skrzydlewska, E. Natural Exogenous Antioxidant Defense against Changes in Human Skin Fibroblast Proteome Disturbed by UVA Radiation. Oxid. Med. Cell. Longev. 2020, 2020, 3216415. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Herrero, J.A.; Frutos, M.J. Influence of rutin and ascorbic acid in colour, plum anthocyanins and antioxidant capacity stability in model juices. Food Chem. 2015, 173, 495–500. [Google Scholar] [CrossRef]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Synergistic cytoprotective effects of rutin and ascorbic acid on the proteomic profile of 3D-cultured keratinocytes exposed to UVA or UVB radiation. Nutrients 2019, 11, 2672. [Google Scholar] [CrossRef]
- Gęgotek, A.; Jastrząb, A.; Dobrzyńska, M.; Biernacki, M.; Skrzydlewska, E. Exogenous Antioxidants Impact on UV-Induced Changes in Membrane Phospholipids and the Effectiveness of the Endocannabinoid System in Human Skin Cells. Antioxidants 2021, 10, 1260. [Google Scholar] [CrossRef]




| Food Products | Amount [mg] Per 100 g |
|---|---|
| Plant origin | |
| Kakadu plum | 5300 |
| Acerola cherries | 1600–1700 |
| Wild rose | 250–800 |
| Blackcurrant | 150–300 |
| Guava | 230 |
| Peppers | 125–200 |
| Brussels | 65–145 |
| Broccoli | 65–100 |
| Grapefruit | 30–70 |
| Pomelo | 61 |
| Lemon | 40–60 |
| Orange | 50 |
| Lime | 29 |
| Animal origin | |
| Liver | 22–30 |
| Cod | 2 |
| Trout | 1 |
| Cow’s milk | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gęgotek, A.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid. Antioxidants 2022, 11, 1993. https://doi.org/10.3390/antiox11101993
Gęgotek A, Skrzydlewska E. Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid. Antioxidants. 2022; 11(10):1993. https://doi.org/10.3390/antiox11101993
Chicago/Turabian StyleGęgotek, Agnieszka, and Elżbieta Skrzydlewska. 2022. "Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid" Antioxidants 11, no. 10: 1993. https://doi.org/10.3390/antiox11101993
APA StyleGęgotek, A., & Skrzydlewska, E. (2022). Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid. Antioxidants, 11(10), 1993. https://doi.org/10.3390/antiox11101993