
Physical Activity vs. Redox Balance in the Brain: Brain Health, Aging and Diseases

 , , ,
, , ,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
Abstract
1. Introduction
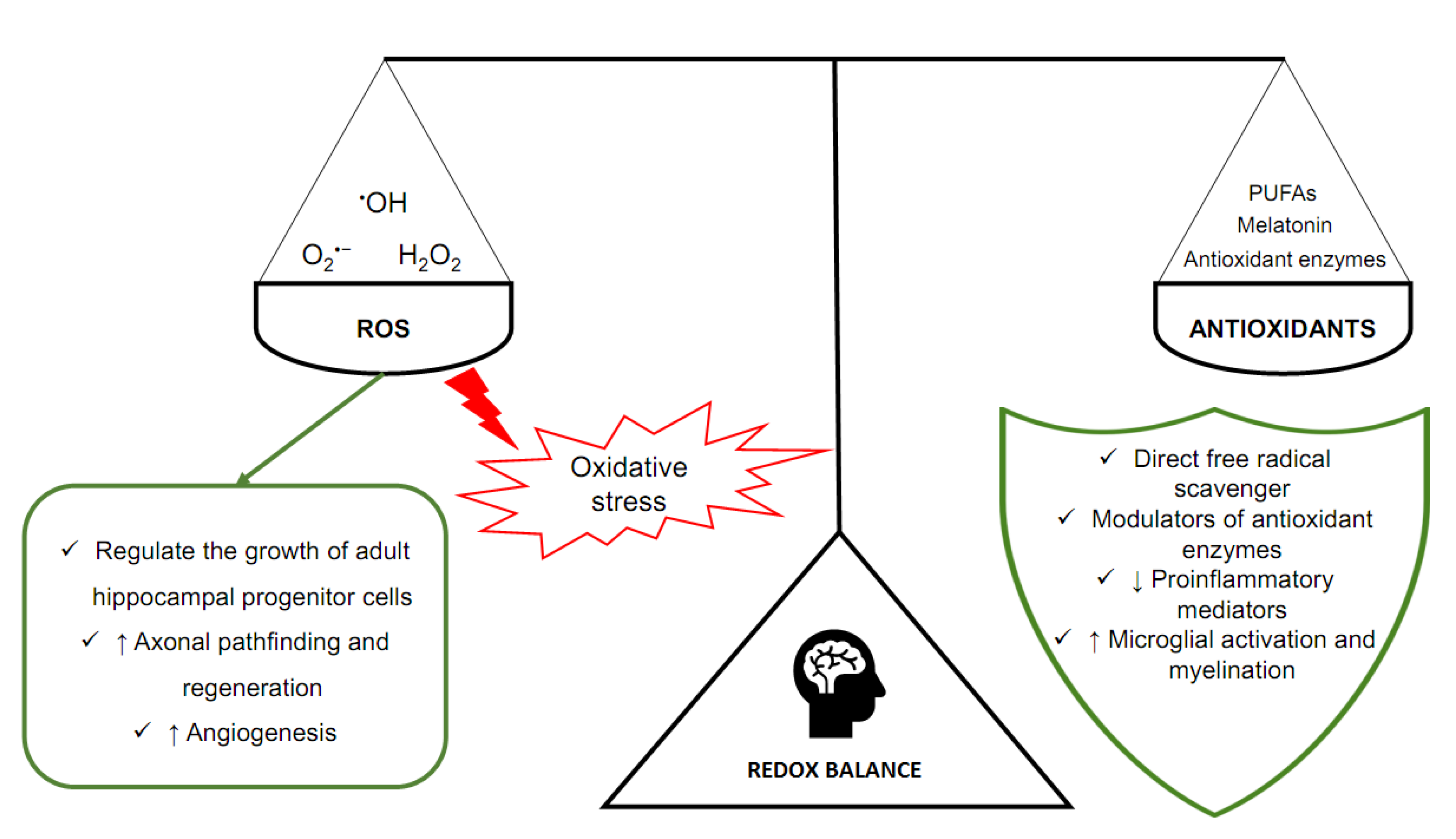
2. The Oxidant–Antioxidant (Redox) Balance
3. The Redox Balance in Brain Disorders
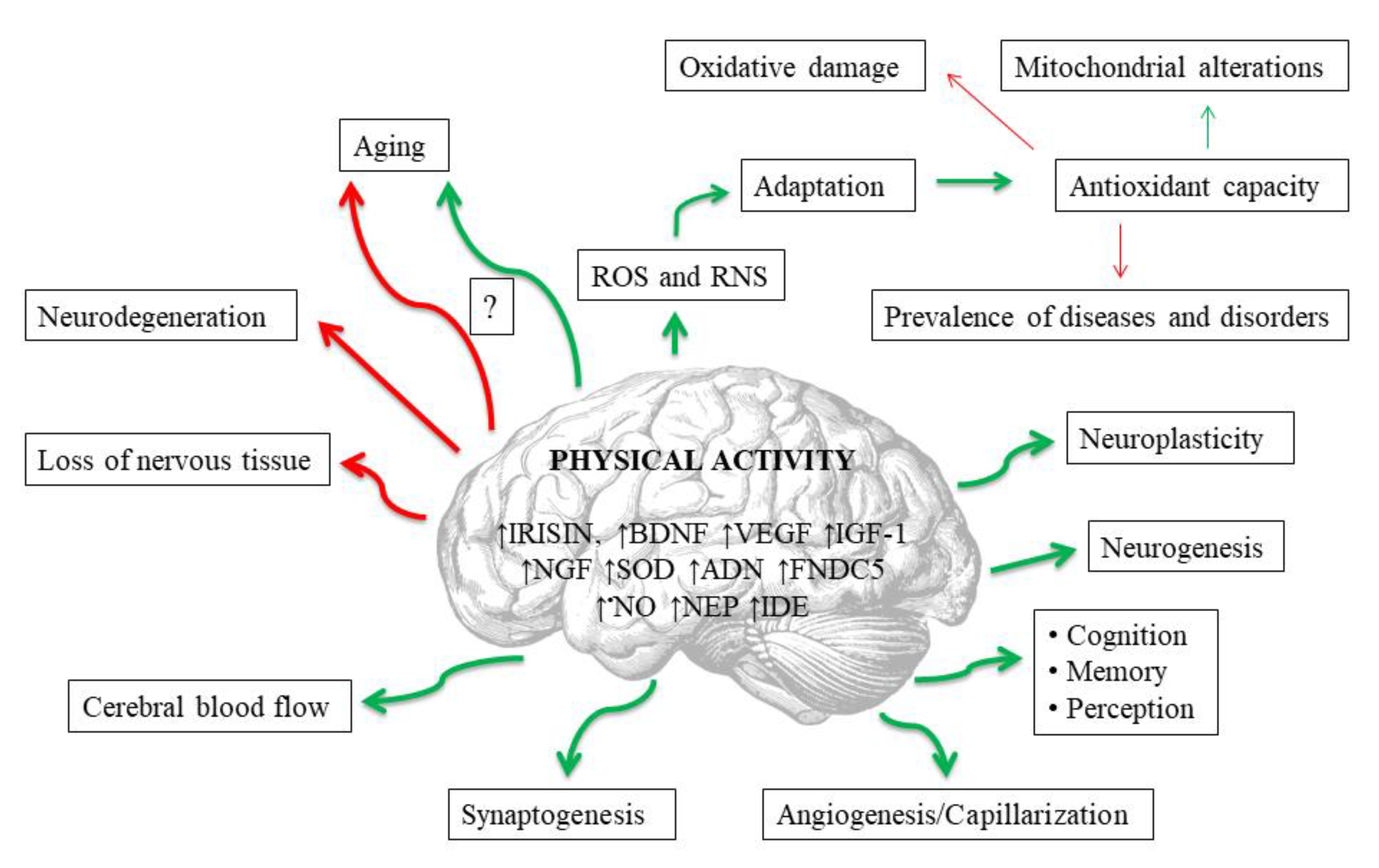
4. The Influence of Physical Exercise on the Redox Balance
5. The Influence of Physical Exercise on Redox Balance in Aging and Brain Diseases
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Aβ | amyloid-β |
| ADN | adiponectin |
| AD | Alzheimer’s disease |
| BDNF | brain-derived neurotrophic factor |
| CAT | catalase |
| CD | conjugated dienes |
| CNS | central nervous system |
| DHA | docosahexaenoic acid |
| FNDC5 | fibronectin type III domain containing 5 |
| GPX | glutathione peroxidase |
| GR | glutathione reductase |
| GSH/GSSG | glutathione (reduced/oxidized form) |
| H2O2 | hydrogen peroxide |
| IDE | insulin-degrading enzyme |
| IGF-1 | insulin-like growth factor 1 |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| ND | neurodegenerative diseases |
| NEP | neprilysin |
| NGF | nerve growth factor |
| •NO | nitric oxide |
| O2•− | superoxide anion radical |
| OFRs | oxygen free radicals |
| •OH | hydroxyl radical |
| PA | physical activity |
| PD | Parkinson’s disease |
| PUFAs | polyunsaturated fatty acids |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| TBARS | thiobarbituric acid reactive substances |
| TRX | thioredoxin |
| VEGF | vascular endothelial growth factor |
References
- Panegyres, K.P.; Panegyres, P.K. The ancient Greek discovery of the nervous system: Alcmaeon, Praxagoras and Herophilus. J. Clin. Neurosci. 2016, 29, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Di Liegro, C.M.; Schiera, G.; Proia, P.; Di Liegro, I. Physical activity and brain health. Genes 2019, 10, 720. [Google Scholar] [CrossRef]
- Erickson, K.I.; Hillman, C.; Stillman, C.M.; Ballard, R.M.; Bloodgood, G.; Conroy, D.E.; Macko, R.; Marquez, D.X.; Petruzzello, S.J.; Powel, K.E.; et al. Physical activity, cognition, and brain outcomes: A review of the 2018 physical activity guidelines. Med. Sci. Sports Exerc. 2019, 51, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Peternelj, T.T.; Coombes, J.S. Antioxidant supplementation during exercise training: Beneficial or detrimental? Sports Med. 2011, 41, 1043–1069. [Google Scholar] [CrossRef]
- Sutkowy, P.; Woźniak, A.; Boraczyński, T.; Boraczyński, M.; Mila-Kierzenkowska, C. The oxidant-antioxidant equilibrium, activities of selected lysosomal enzymes and activity of acute phase protein in peripheral blood of 18-year-old football players after aerobic cycle ergometer test combined with ice-water immersion or recovery at room temperature. Cryobiology 2017, 74, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, C.C.; Chaturvedi, M.M.; Aqqurwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Díaz, M.; Mesa-Herrera, F.; Marín, R. DHA and its elaborated modulation of antioxidant defenses of the brain: Implications in aging and AD neurodegeneration. Antioxidants 2021, 10, 907. [Google Scholar] [CrossRef]
- Rink, C.; Khanna, S. Significance of brain tissue oxygenation and the arachidonic acid cascade in stroke. Antioxid. Redox Signal. 2011, 14, 1889–1903. [Google Scholar] [CrossRef]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef]
- Jardim, F.R.; de Rossi, F.T.; Nascimento, M.X.; da Silva Barros, R.G.; Borges, P.A.; Prescilio, I.C.; de Oliveira, M.R. Resveratrol and brain mitochondria: A review. Mol. Neurobiol. 2018, 55, 2085–2101. [Google Scholar] [CrossRef]
- Vinokurov, A.Y.; Stelmashuk, O.A.; Ukolova, P.A.; Zherebtsov, E.A.; Abramov, A.Y. Brain region specificity in reactive oxygen species production and maintenance of redox balance. Free Radic. Biol. Med. 2021, 174, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.D.; Wang, J.A.; Bosken, J.M.; Singh, I.N. Lipid peroxidation in brain or spinal cord mitochondria after injury. J. Bioenerg. Biomembr. 2016, 48, 169–174. [Google Scholar] [CrossRef]
- Stefanatos, R.; Sanz, A. The role of mitochondrial ROS in the aging brain. FEBS Lett. 2018, 592, 743–758. [Google Scholar] [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Role of mitochondrial ROS in the brain: From physiology to neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.C.; Peltier, J.; Stone, D.; Schaffer, D.V.; Chang, C.J. Nox2 redox signaling maintains essential cell populations in the brain. Nat. Chem. Biol. 2011, 7, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Gauron, C.; Meda, F.; Dupont, E.; Albadri, S.; Quenech’Du, N.; Ipendey, E.; Volovitch, M.; Del Bene, F.; Joliot, A.; Rampon, C.; et al. Hydrogen peroxide (H2O2) controls axon pathfinding during zebrafish development. Dev. Biol. 2016, 414, 133–141. [Google Scholar] [CrossRef]
- Meda, F.; Gauron, C.; Rampon, C.; Teillon, J.; Volovitch, M.; Vriz, S. Nerves control redox levels in mature tissues through schwann cells and hedgehog signaling. Antioxid. Redox Signal. 2016, 24, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Yang, J. The role of reactive oxygen species in angiogenesis and preventing tissue injury after brain ischemia. Microvasc. Res. 2019, 123, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Coimbra-Costa, D.; Alva, N.; Duran, M.; Carbonell, T.; Rama, R. Oxidative stress and apoptosis after acute respiratory hypoxia and reoxygenation in rat brain. Redox Biol. 2017, 12, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Winiarska-Mieczan, A.; Baranowska-Wójcik, E.; Kwiecień, M.; Grela, E.R.; Szwajgier, D.; Kwiatkowska, K.; Kiczorowska, B. The role of dietary antioxidants in the pathogenesis of neurodegenerative diseases and their impact on cerebral oxidoreductive balance. Nutrients 2020, 12, 435. [Google Scholar] [CrossRef] [PubMed]
- Salminen, L.E.; Paul, R.H. Oxidative stress and genetic markers of suboptimal antioxidant defense in the aging brain: A theoretical review. Rev. Neurosci. 2014, 25, 139–148. [Google Scholar] [CrossRef]
- Ehsaei, M.; Khajavi, M.; Arjmand, M.H.; Abuee, M.A.; Ghayour-Mobarhan, M.; Hamidi Alamdari, D. Prooxidant–antioxidant balance in patients with traumatic brain injury. Acta Neurol. Belg. 2015, 115, 69–73. [Google Scholar] [CrossRef]
- Fernández-Gajardo, R.; Matamala, J.M.; Carrasco, R.; Gutiérrez, R.; Melo, R.; Rodrigo, R. Novel therapeutic strategies for traumatic brain injury: Acute antioxidant reinforcement. CNS Drugs 2014, 28, 229–248. [Google Scholar] [CrossRef]
- Onaolapo, O.J.; Onaolapo, A.Y. Melatonin, adolescence, and the brain: An insight into the period-specific influences of a multifunctional signaling molecule. Birth Defects Res. 2017, 109, 1659–1671. [Google Scholar] [CrossRef]
- Hussain, G.; Anwar, H.; Rasul, A.; Imran, A.; Qasim, M.; Zafar, S.; Imran, M.; Kamran, S.K.S.; Aziz, N.; Razzaq, A.; et al. Lipids as biomarkers of brain disorders. Crit. Rev. Food Sci. Nutr. 2020, 60, 351–374. [Google Scholar] [CrossRef] [PubMed]
- Akanji, M.A.; Rotimi, D.E.; Elebiyo, T.C.; Awakan, O.J.; Adeyemi, O.S. Redox Homeostasis and prospects for therapeutic targeting in neurodegenerative disorders. Oxid. Med. Cell. Longev. 2021, 2021, 9971885. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative stress: A key modulator in neurodegenerative diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S. Neuroprotective function of high glycolytic activity in astrocytes: Common roles in stroke and neurodegenerative diseases. Int. J. Mol. Sci. 2021, 22, 6568. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, R.; Sisalli, M.; Sirabella, R.; Della Notte, S.; Borzacchiello, D.; Feliciello, A.; Annunziato, L.; Scorziello, A. Ncx3-Induced Mitochondrial Dysfunction in Midbrain Leads to Neuroinflammation in Striatum of A53t-α-Synuclein Transgenic Old Mice. Int. J. Mol. Sci. 2021, 22, 8177. [Google Scholar] [CrossRef] [PubMed]
- Buccellato, F.R.; D’Anca, M.; Fenoglio, C.; Scarpini, E.; Galimberti, D. Role of oxidative damage in alzheimer’s disease and neurodegeneration: From pathogenic mechanisms to biomarker discovery. Antioxidants 2021, 10, 1353. [Google Scholar] [CrossRef] [PubMed]
- Ionescu-Tucker, A.; Cotman, C.W. Emerging roles of oxidative stress in brain aging and Alzheimer’s disease. Neurobiol. Aging 2021, 107, 86–95. [Google Scholar] [CrossRef]
- Burtscher, J.; Millet, G.P.; Place, N.; Kayser, B.; Zanou, N. The muscle-brain axis and neurodegenerative diseases: The key role of mitochondria in exercise-induced neuroprotection. Int. J. Mol. Sci. 2021, 22, 6479. [Google Scholar] [CrossRef] [PubMed]
- Asanuma, M.; Miyazaki, I. Glutathione and related molecules in parkinsonism. Int. J. Mol. Sci. 2021, 22, 8689. [Google Scholar] [CrossRef]
- Higashi, Y.; Aratake, T.; Shimizu, T.; Shimizu, S.; Saito, M. Protective role of glutathione in the hippocampus after brain ischemia. Int. J. Mol. Sci. 2021, 22, 7765. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Perera, L.M.; Höving, A.L.; Schmidt, K.E.; Cenan, S.; Wohllebe, M.; Greiner, J.F.W.; Kaltschmidt, C.; Simon, M.; Knabbe, C.; Kaltschmidt, B. Neuroprotection mediated by human blood plasma in mouse hippocampal slice cultures and in oxidatively stressed human neurons. Int. J. Mol. Sci. 2021, 22, 9567. [Google Scholar] [CrossRef] [PubMed]
- Mashima, K.; Takahashi, S.; Minami, K.; Izawa, Y.; Abe, T.; Tsukada, N.; Hishiki, T.; Suematsu, M.; Kajimura, M.; Suzuki, N. Neuroprotective role of astroglia in parkinson disease by reducing oxidative stress through dopamine-induced activation of pentose-phosphate pathway. ASN Neuro 2018, 10, 175909141877556. [Google Scholar] [CrossRef]
- Penkowa, M. Metallothioneins are multipurpose neuroprotectants during brain pathology. FEBS J. 2006, 273, 1857–1870. [Google Scholar] [CrossRef]
- Chen, P.-C.; Yu, C.-C.; Chen, Y.-S.; Lu, C.-H.; Chan, S.-H.; Chou, K.-H.; Chen, M.-H.; Chen, M.-H.; Lin, W.C. The potential effects of oxidative stress-related plasma abnormal protein aggregate levels on brain volume and its neuropsychiatric consequences in Parkinson’s disease. Oxid. Med. Cell. Longev. 2021, 2021, 3666327. [Google Scholar] [CrossRef]
- Gonzalez-Riano, C.; Saiz, J.; Barbas, C.; Bergareche, A.; Huerta, J.M.; Ardanaz, E.; Konjevod, M.; Mondragon, E.; Erro, M.E.; Chirlaque, M.D.; et al. Prognostic biomarkers of Parkinson’s disease in the Spanish EPIC cohort: A multiplatform metabolomics approach. NPJ Park. Dis. 2021, 7, 73. [Google Scholar] [CrossRef]
- Perry, E.A.; Castellani, R.J.; Moreira, P.I.; Nunomura, A.; Lui, Q.; Harris, P.L.R.; Sayre, L.M.; Szweda, P.A.; Szweda, L.I.; Zhu, H.; et al. Neurofilaments are the major neuronal target of hydroxynonenal-mediated protein cross-links. Free Radic. Res. 2013, 47, 507–510. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Di Domenico, F.; Barone, E.; Perluigi, M.; Butterfield, D.A. The triangle of death in Alzheimer’s disease brain: The aberrant cross-talk among energy metabolism, mammalian target of rapamycin signaling, and protein homeostasis revealed by redox proteomics. Antioxid. Redox Signal. 2017, 26, 364–387. [Google Scholar] [CrossRef]
- Di Domenico, F.; Pupo, G.; Giraldo, E.; Badìa, M.-C.; Monllor, P.; Lloret, A.; Eugenia Schininà, M.; Giorgi, A.; Cini, C.; Tramutola, A.; et al. Oxidative signature of cerebrospinal fluid from mild cognitive impairment and Alzheimer disease patients. Free Radic. Biol. Med. 2016, 91, 1–9. [Google Scholar] [CrossRef]
- Yamanaka, G.; Suzuki, S.; Morishita, N.; Takeshita, M.; Kanou, K.; Takamatsu, T.; Morichi, S.; Ishida, Y.; Watanabe, Y.; Go, S.; et al. Experimental and clinical evidence of the effectiveness of riboflavin on migraines. Nutrients 2021, 13, 2612. [Google Scholar] [CrossRef]
- Gross, E.C.; Putananickal, N.; Orsini, A.-L.; Vogt, D.R.; Sandor, P.S.; Schoenen, J.; Fischer, D. Mitochondrial function and oxidative stress markers in higher-frequency episodic migraine. Sci. Rep. 2021, 11, 4543. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, G.M.; Kalita, J.; Misra, U.K. A study of oxidative stress in migraine with special reference to prophylactic therapy. Int. J. Neurosci. 2018, 128, 318–324. [Google Scholar] [CrossRef]
- Cegielska, J.; Szmidt-Sałkowska, E.; Domitrz, W.; Gaweł, M.; Radziwoń-Zaleska, M.; Domitrz, I. Migraine and its association with hyperactivity of cell membranes in the course of latent magnesium deficiency—Preliminary study of the importance of the latent tetany presence in the migraine pathogenesis. Nutrients 2021, 13, 2701. [Google Scholar] [CrossRef] [PubMed]
- Abruzzo, P.M.; Panisi, C.; Marini, M. The alteration of chloride homeostasis/GABAergic signaling in brain disorders: Could oxidative stress play a role? Antioxidants 2021, 10, 1316. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Kennedy, K.G.; Grigorian, A.; Fiksenbaum, L.; Freeman, N.; Zai, C.C.; Kennedy, J.L.; MacIntosh, B.J.; Goldstein, B.I. Antioxidative defense genes and brain structure in youth bipolar disorder. Int. J. Neuropsychopharmacol. 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Castillo, L.; Phillips-Farfán, B.V.; Rosas-Mendoza, G.; Baires-López, A.; Toral-Ríos, D.; Campos-Peña, V.; Carvajal, K. Increased oxidative stress contributes to enhance brain amyloidogenesis and blunts energy metabolism in sucrose-fed rat: Effect of AMPK activation. Sci. Rep. 2021, 11, 19547. [Google Scholar] [CrossRef]
- Grubač, Ž.; Šutulović, N.; Šuvakov, S.; Jerotić, D.; Puškaš, N.; Macut, D.; Rašić-Marković, A.; Simić, T.; Stanojlović, O.; Hrnčić, D. Anxiogenic potential of experimental sleep fragmentation is duration-dependent and mediated via oxidative stress state. Oxid. Med. Cell. Longev. 2021, 2021, 2262913. [Google Scholar] [CrossRef]
- Juszczyk, G.; Mikulska, J.; Kasperek, K.; Pietrzak, D.; Mrozek, W.; Herbet, M. Chronic stress and oxidative stress as common factors of the pathogenesis of depression and Alzheimer’s disease: The role of antioxidants in prevention and treatment. Antioxidants 2021, 10, 1439. [Google Scholar] [CrossRef] [PubMed]
- Moosmann, B.; Behl, C. Antioxidants as treatment for neurodegenerative disorders. Expert Opin. Investig. Drugs 2002, 11, 1407–1435. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, S.L.; Stab, B.; Casas, Z.; Sutachan, J.J.; Samudio, I.; Gonzalez, J.; Gonzalo, L.; Capani, F.; Morales, L.; Barreto, G.E. Effects of natural antioxidants in neurodegenerative disease. Nutr. Neurosci. 2012, 15, 1–9. [Google Scholar] [CrossRef]
- Vauzour, D. Dietary polyphenols as modulators of brain functions: Biological actions and molecular mechanisms underpinning their beneficial effects. Oxid. Med. Cell. Longev. 2012, 2012, 914273. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, X. Antioxidant therapies for Alzheimer’s disease. Oxid. Med. Cell. Longev. 2012, 2012, 472932. [Google Scholar] [CrossRef]
- Conti, V.; Izzo, V.; Corbi, G.; Russomanno, G.; Manzo, V.; De Lise, F.; Di Donato, A.; Filipelli, A. Antioxidant supplementation in the treatment of aging-associated diseases. Front. Pharmacol. 2016, 7, 24. [Google Scholar] [CrossRef]
- Velusamy, T.; Panneerselvam, A.S.; Purushottam, M.; Anusuyadevi, M.; Pal, P.K.; Jain, S.; Essa, M.M.; Guillemin, G.J.; Kandasamy, M. Protective effect of antioxidants on neuronal dysfunction and plasticity in Huntington’s disease. Oxid. Med. Cell. Longev. 2017, 2017, 3279061. [Google Scholar] [CrossRef]
- González-Fuentes, J.; Selva, J.; Moya, C.; Castro-Vázquez, L.; Lozano, M.V.; Marcos, P.; Plaza-Oliver, M.; Rodríguez-Robledo, V.; Santander-Ortega, M.; Villaseca-González, N.; et al. Neuroprotective natural molecules, from food to brain. Front. Neurosci. 2018, 12, 721. [Google Scholar] [CrossRef]
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective effect of antioxidants in the brain. Int. J. Mol. Sci. 2020, 21, 7152. [Google Scholar] [CrossRef]
- Scapagnini, G.; Sonya, V.; Nader, A.G.; Calogero, C.; Zella, D.; Fabio, G. Modulation of Nrf2/ARE pathway by food polyphenols: A nutritional neuroprotective strategy for cognitive and neurodegenerative disorders. Mol. Neurobiol. 2011, 44, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Aliev, G.; Obrenovich, M.; Reddy, V.; Shenk, J.; Moreira, P.; Nunomura, A.; Zhu, X.; Smith, M.A.; Perry, G. Antioxidant therapy in Alzheimer’s disease: Theory and practice. Mini-Rev. Med. Chem. 2008, 8, 1395–1406. [Google Scholar] [CrossRef]
- Fernandes, C.; Pinto, M.; Martins, C.; Gomes, M.J.; Sarmento, B.; Oliveira, P.J.; Remião, F.; Borges, F. Development of a PEGylated-based platform for efficient delivery of dietary antioxidants across the blood-brain barrier. Bioconjug. Chem. 2018, 29, 1677–1689. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Galasko, D.R.; Peskind, E.; Clark, C.M.; Quinn, J.F.; Ringman, J.M.; Jicha, G.A.; Cotman, C.; Cottrell, B.; Montine, T.J.; Thomas, R.G.; et al. Antioxidants for Alzheimer disease: A randomized clinical trial with cerebrospinal fluid biomarker measures. Arch. Neurol. 2012, 69, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Shirle, R.; Ord, E.N.J.; Work, L.M. Oxidative stress and the use of antioxidants in stroke. Antioxidants 2014, 3, 472–501. [Google Scholar] [CrossRef]
- Nakase, T.; Yoshioka, S.; Suzuki, A. Free radical scavenger, edaravone, reduces the lesion size of lacunar infarction in human brain ischemic stroke. BMC Neurol. 2011, 11, 35–41. [Google Scholar] [CrossRef]
- Imai, T.; Iwata, S.; Miyo, D.; Nakamura, S.; Shimazawa, M.; Hara, H. A novel free radical scavenger, NSP-116, ameliorated the brain injury in both ischemic and hemorrhagic stroke models. J. Pharmacol. Sci. 2019, 141, 119–126. [Google Scholar] [CrossRef]
- Aly, H.; Elmahdy, H.; El-Dib, M.; Rowisha, M.; Awny, M.; El-Gohary, T.; Elbatch, M.; Hamisa, M.; El-Mashad, A.-R. Melatonin use for neuroprotection in perinatal asphyxia: A randomized controlled pilot study. J Perinatol. 2015, 35, 186–191. [Google Scholar] [CrossRef]
- Ahmad, Q.M.; Chishti, A.L.; Waseem, N. Role of melatonin in management of hypoxic ischaemic encephalopathy in newborns: A randomized control trial. J. Pak. Med. Assoc. 2018, 68, 1233–1237. [Google Scholar] [PubMed]
- Alkhatib, A.; Feng, W.H.; Huang, Y.J.; Kuo, C.H.; Hou, C.W. Anserine reverses exercise-induced oxidative stress and preserves cellular homeostasis in healthy men. Nutrients 2020, 12, 1146. [Google Scholar] [CrossRef]
- Tsao, J.P.; Liu, C.C.; Wang, H.F.; Bernard, J.R.; Huang, C.C.; Cheng, I.S. Oral resveratrol supplementation attenuates exercise-induced interleukin-6 but not oxidative stress after a high cycling challenge in adults. Int. J. Med. Sci. 2021, 18, 2137–2145. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, A. Signs of oxidative stress after exercise. Biol. Sport 2003, 20, 93–112. [Google Scholar]
- Kavazis, A.N.; Talbert, E.E.; Smuder, A.J.; Hudson, M.B.; Nelson, W.B.; Powers, S.K. Mechanical ventilation induces diaphragmatic mitochondrial dysfunction and increased oxidant production. Free Radic. Biol. Med. 2009, 46, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Dominiak, K.; Jarmuszkiewicz, W. The Relationship between mitochondrial reactive oxygen species production and mitochondrial energetics in rat tissues with different contents of reduced coenzyme Q. Antioxidants 2021, 10, 533. [Google Scholar] [CrossRef]
- Powers, S.K.; Nelson, W.B.; Hudson, M.B. Exercise-induced oxidative stress in humans: Cause and consequences. Free Radic. Biol. Med. 2011, 51, 942–950. [Google Scholar] [CrossRef]
- Vollaard, N.B.; Shearman, J.P.; Cooper, C.E. Exercise-induced oxidative stress: Myths, realities and physiological relevance. Sports Med. 2005, 35, 1045–1062. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E.C.; Silva, A.N.; de Oliveira, M.R. Oxidants, antioxidants, and the beneficial roles of exercise-induced production of reactive species. Oxid. Med. Cell. Longev. 2012, 2012, 756132. [Google Scholar] [CrossRef]
- Souza-Junior, T.; Lorenço-Lima, L.; Ganini, D.; Vardaris, C.; Polotow, T.; Barros, M. Delayed uric acid accumulation in plasma provides additional anti-oxidant protection against iron-triggered oxidative stress after a wingate test. Biol. Sport. 2014, 31, 271–276. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Goldfarb, A.H.; Wideman, L.; McKenzie, M.J.; Consitt, L.A. Effects of acute aerobic and anaerobic exercise on blood markers of oxidative stress. J. Strength Cond. Res. 2005, 19, 276–285. [Google Scholar] [CrossRef]
- Bouzid, M.M.; Filaire, E.; McCall, A.; Fabre, C. Radical oxygen species, exercise and aging: An update. Sports Med. 2015, 45, 1245–1261. [Google Scholar] [CrossRef]
- Taito, S.; Sekikawa, K.; Oura, K.; Kamikawa, N.; Matsuki, R.; Kimura, T.; Takahashi, M.; Inamizu, T.; Hamada, H. Plasma oxidative stress is induced by single-sprint anaerobic exercise in young cigarette smokers. Clin. Physiol. Funct. Imaging 2013, 33, 241–244. [Google Scholar] [CrossRef]
- Sutkowy, P.; Woźniak, A.; Boraczyński, T.; Mila-Kierzenkowska, C.; Boraczyński, M. Postexercise impact of ice-cold water bath on the oxidant-antioxidant balance in healthy men. Biomed. Res. Int. 2015, 2015, 706141. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, A.; Mila-Kierzenkowska, C.; Szpinda, M.; Chwalbinska-Moneta, J.; Augustyńska, B.; Jurecka, A. Whole-body cryostimulation and oxidative stress in rowers: The preliminary results. Arch. Med. Sci. 2013, 9, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Rakowski, A.; Jurecka, A.; Rajewski, R. Whole-body cryostimulation in kayaker women: A study of the effect of cryogenic temperatures on oxidative stress after the exercise. J. Sports Med. Phys. Fitness 2009, 49, 201–207. [Google Scholar] [PubMed]
- De Sousa, C.V.; Sales, M.M.; Rosa, T.S.; Lewis, J.E.; De Andrade, R.V.; Simões, H.G. The antioxidant effect of exercise: A systematic review and meta-analysis. Sports Med. 2017, 47, 277–293. [Google Scholar] [CrossRef]
- Cho, S.Y.; So, W.Y.; Roh, H.T. Effect of C242T polymorphism in the gene encoding the NAD(P)H oxidase p22(phox) subunit and aerobic fitness levels on redox state biomarkers and DNA damage responses to exhaustive exercise: A randomized trial. Int. J. Environ. Res. Public Health 2020, 17, 4215. [Google Scholar] [CrossRef]
- Finaud, J.; Lac, G.; Filaire, E. Oxidative stress: Relationship with exercise and training. Sports Med. 2006, 36, 327–358. [Google Scholar] [CrossRef]
- Woźniak, A.; Drewa, G.; Chęsy, G.; Rakowski, A.; Rozwodowska, M.; Olszewska, D. Effect of altitude training on the peroxidation and antioxidant enzymes in sportsmen. Med. Sci. Sports Exerc. 2001, 33, 1109–1113. [Google Scholar] [CrossRef]
- Gomes, M.J.; Pagan, L.U.; Lima, A.R.R.; Reyes, D.R.A.; Martinez, P.F.; Damatto, F.C.; Pontes, T.H.D.; Rodrigues, E.A.; Souza, L.M.; Tosta, I.F.; et al. Effects of aerobic and resistance exercise on cardiac remodelling and skeletal muscle oxidative stress of infarcted rats. J. Cell Mol. Med. 2020, 24, 5352–5362. [Google Scholar] [CrossRef]
- Ye, Y.; Lin, H.; Wan, M.; Qiu, P.; Xia, R.; He, J.; Tao, J.; Chen, L.; Zheng, G. The effects of aerobic exercise on oxidative stress in older adults: A systematic review and meta-analysis. Front Physiol. 2021, 12, 701151. [Google Scholar] [CrossRef]
- Lee, S.J. Effects of preconditioning exercise on nitric oxide and antioxidants in hippocampus of epileptic seizure. J. Exerc. Rehabil. 2019, 15, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Camiletti-Moirón, D.; Aparicio, V.A.; Aranda, P.; Radak, Z. Does exercise reduce brain oxidative stress? A systematic review. Scand. J. Med. Sci. Sports 2013, 23, e202–e212. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Radak, Z.; Ihasz, F.; Koltai, E.; Goto, S.; Taylor, A.W.; Boldogh, I. The redox-associated adaptive response of brain to physical exercise. Free Radic. Res. 2014, 48, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.J.; Cotter, J.D.; Brassard, P.; Bailey, D.M. High-intensity interval exercise and cerebrovascular health: Curiosity, cause, and consequence. J. Cereb. Blood Flow Metab. 2015, 35, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Małkiewicz, M.A.; Szarmach, A.; Sabisz, A.; Cubała, W.J.; Szurowska, E.; Winklewski, P.J. Blood-brain barrier permeability and physical exercise. J. Neuroinflamm. 2019, 16, 15. [Google Scholar] [CrossRef]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Sutkowy, P.; Woźniak, A.; Mila-Kierzenkowska, C. Positive effect of generation of reactive oxygen species on the human organism. Med. Biol. Sci. 2013, 27, 13–17. [Google Scholar] [CrossRef]
- Petrovic, S.; Arsic, A.; Ristic-Medic, D.; Cvetkovic, Z.; Vucic, V. Lipid peroxidation and antioxidant supplementation in neurodegenerative diseases: A review of human studies. Antioxidants 2020, 9, 1128. [Google Scholar] [CrossRef]
- Real, C.C.; Binda, K.H.; Landau, A.M. Treadmill exercise and neuroinflammation: Links with aging. In Factors Affecting Neurological Aging; Academic Press: Cambridge, MA, USA, 2021; pp. 385–401. [Google Scholar] [CrossRef]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef]
- Ziada, A.S.; Smith, M.S.R.; Côté, H.C.F. Updating the free radical theory of aging. Front. Cell Dev. Biol. 2020, 8, 575645. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Mañas, L.; El-Assar, M.; Vallejo, S.; López-Dóriga, P.; Solís, J.; Petidier, R.; Montes, M.; Nevado, J.; Castro, M.; Gómez-Guerrero, C.; et al. Endothelial dysfunction in aged humans is related with oxidative stress and vascular inflammation. Aging Cell. 2009, 8, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Ardiles, Á.O.; Pérez-Espinosa, B.; Núñez-Espinosa, C.; Paula-Lima, A.; González-Billault, C.; Espinosa-Parrilla, Y. Redox modifications in synaptic components as biomarkers of cognitive status, in brain aging and disease. Mech. Ageing Dev. 2020, 189, 111250. [Google Scholar] [CrossRef] [PubMed]
- Paillard, T. Preventive effects of regular physical exercise against cognitive decline and the risk of dementia with age advancement. Sports Med. Open 2015, 1, 20. [Google Scholar] [CrossRef]
- Quan, H.; Koltai, E.; Suzuki, K.; Aguiar, A.S.; Pinho, R.; Boldogh, I.; Berkes, I.; Radak, Z. Exercise, redox system and neurodegenerative diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165778. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.S.; Margaritis, I.; Arnaud, J.; Faure, H.; Roussel, A.M. Physical activity alters antioxidant status in exercising elderly subjects. J. Nutr. Biochem. 2006, 17, 463–470. [Google Scholar] [CrossRef]
- Vanzella, C.; Neves, J.D.; Vizuete, A.F.; Aristimunha, D.; Kolling, J.; Longoni, A.; Gonçalves, C.A.S.; Wyse, A.T.S.; Netto, C.A. Treadmill running prevents age-related memory deficit and alters neurotrophic factors and oxidative damage in the hippocampus of Wistar rats. Behav. Brain Res. 2017, 334, 78–85. [Google Scholar] [CrossRef]
- Bernardo, T.C.; Beleza, J.; Rizo-Roca, D.; Santos-Alves, E.; Leal, C.; Martins, M.J.; Ascensão, A.; Magalhães, J. Physical exercise mitigates behavioral impairments in a rat model of sporadic Alzheimer’s disease. Behav. Brain Res. 2020, 379, 112358. [Google Scholar] [CrossRef]
- Bernardo, T.C.; Marques-Aleixo, I.; Beleza, J.; Oliveira, P.J.; Ascensão, A.; Magalhães, J. Physical exercise and brain mitochondrial fitness: The possible role against Alzheimer’s disease. Brain Pathol. 2016, 26, 648–663. [Google Scholar] [CrossRef]
- Liang, Y.Y.; Zhang, L.D.; Luo, X.; Wu, L.L.; Chen, Z.W.; Wei, G.H.; Zhang, K.Q.; Du, Z.-A.; Li, R.-Z.; So, K.-F.; et al. All roads lead to Rome—a review of the potential mechanisms by which exerkines exhibit neuroprotective effects in Alzheimer’s disease. Neural Regen Res. 2022, 17, 1210–1227. [Google Scholar] [CrossRef]
- Yu, M.; Tsai, S.-F.; Kuo, Y.-M. The therapeutic potential of anti-inflammatory exerkines in the treatment of atherosclerosis. IJMS. 2017, 18, 1260. [Google Scholar] [CrossRef]
- Jin, Y.; Sumsuzzman, D.M.; Choi, J.; Kang, H.; Lee, S.-R.; Hong, Y. Molecular and functional interaction of the myokine irisin with physical exercise and Alzheimer’s disease. Molecules 2018, 23, 3229. [Google Scholar] [CrossRef]
- Pesce, M.; La Fratta, I.; Paolucci, T.; Grilli, A.; Patruno, A.; Agostini, F.; Bernetti, A.; Mangone, M.; Paolini, M.; Invernizzi, M.; et al. From exercise to cognitive performance: Role of irisin. Appl. Sci. 2021, 11, 7120. [Google Scholar] [CrossRef]
- Dun, S.L.; Lyu, R.-M.; Chen, Y.-H.; Chang, J.-K.; Luo, J.J.; Dun, N.J. Irisin-immunoreactivity in neural and non-neural cells of the rodent. Neuroscience 2013, 240, 155–162. [Google Scholar] [CrossRef]
- Sellami, M.; Bragazzi, N.; Prince, M.S.; Denham, J.; Elrayess, M. Regular, intense exercise training as a healthy aging lifestyle strategy: Preventing DNA damage, telomere shortening and adverse DNA methylation changes over a lifetime. Front. Genet. 2021, 12, 652497. [Google Scholar] [CrossRef] [PubMed]
- Pérez, V.I.; van Remmen, H.; Bokov, A.; Epstein, C.J.; Vijg, J.; Richardson, A. The overexpression of major antioxidant enzymes does not extend the lifespan of mice. Aging Cell 2009, 8, 73–75. [Google Scholar] [CrossRef]
- Melo, K.P.; Silva, C.M.; Almeida, M.F.; Chaves, R.S.; Marcourakis, T.; Cardoso, S.M.; Demasi, M.; Netto, L.E.S.; Ferrari, M.F.R. Mild exercise differently affects proteostasis and oxidative stress on motor areas during neurodegeneration: A comparative study of three treadmill running protocols. Neurotox. Res. 2019, 35, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.F.; Silva, C.M.; Chaves, R.S.; Lima, N.C.R.; Almeida, R.S.; Melo, K.P.; Demasi, M.; Fernandes, T.; Oliveira, E.M.; Netto, L.E.S.; et al. Effects of mild running on substantia nigra during early neurodegeneration. J. Sports Sci. 2018, 36, 1363–1370. [Google Scholar] [CrossRef]
- Sessa, F.; Messina, G.; Russo, R.; Salerno, M.; Castruccio Castracani, C.; Distefano, A.; Li Volti, G.; Calogero, A.E.; Cannarella, L.; Mongioi’, L.M.; et al. Consequences on aging process and human wellness of generation of nitrogen and oxygen species during strenuous exercise. Aging Male 2020, 23, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Bast, A.; Haenen, G.R.M.M. Ten misconceptions about antioxidants. Trends Pharmacol. Sci. 2013, 34, 430–436. [Google Scholar] [CrossRef]
- Sarangarajan, R.; Meera, S.; Rukkumani, R.; Sankar, P.; Anuradha, G. Antioxidants: Friend or foe? Asian Pac. J. Trop. Med. 2017, 10, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Watson, J. Oxidants, antioxidants and the current incurability of metastatic cancers. Open Biol. 2013, 3, 120144. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Martorell, M.; Arbiser, J.L.; Sureda, A.; Martins, N.; Maurya, P.K.; Sharifi-Rad, M.; Kumar, P.; Sharifi-Rad, J. Antioxidants: Positive or negative actors? Biomolecules 2018, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Mentor, S.; Fisher, D. Aggressive antioxidant reductive stress impairs brain endothelial cell angiogenesis and blood brain barrier function. Curr. Neurovasc. Res. 2017, 14, 71–81. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutkowy, P.; Woźniak, A.; Mila-Kierzenkowska, C.; Szewczyk-Golec, K.; Wesołowski, R.; Pawłowska, M.; Nuszkiewicz, J. Physical Activity vs. Redox Balance in the Brain: Brain Health, Aging and Diseases. Antioxidants 2022, 11, 95. https://doi.org/10.3390/antiox11010095
Sutkowy P, Woźniak A, Mila-Kierzenkowska C, Szewczyk-Golec K, Wesołowski R, Pawłowska M, Nuszkiewicz J. Physical Activity vs. Redox Balance in the Brain: Brain Health, Aging and Diseases. Antioxidants. 2022; 11(1):95. https://doi.org/10.3390/antiox11010095
Chicago/Turabian StyleSutkowy, Paweł, Alina Woźniak, Celestyna Mila-Kierzenkowska, Karolina Szewczyk-Golec, Roland Wesołowski, Marta Pawłowska, and Jarosław Nuszkiewicz. 2022. "Physical Activity vs. Redox Balance in the Brain: Brain Health, Aging and Diseases" Antioxidants 11, no. 1: 95. https://doi.org/10.3390/antiox11010095
APA StyleSutkowy, P., Woźniak, A., Mila-Kierzenkowska, C., Szewczyk-Golec, K., Wesołowski, R., Pawłowska, M., & Nuszkiewicz, J. (2022). Physical Activity vs. Redox Balance in the Brain: Brain Health, Aging and Diseases. Antioxidants, 11(1), 95. https://doi.org/10.3390/antiox11010095