
Molecular Responses of Lactobacilli to Plant Phenolic Compounds: A Comparative Review of the Mechanisms Involved



Abstract
:1. Introduction
2. Comparative Transcriptomic Responses of L. plantarum WCFS1 to Different PPCs
3. Oxidative Stress Responses Modulated by PPCs
4. Genotoxic Stress Responses
5. General Stress Pathways Activated in Response to PPCs
6. Detoxification Mechanisms of PPCs in Lactobacillus
6.1. Metabolic Pathways for the Detoxification of PPCs
6.2. Drug Efflux and ABC-Transport Systems
7. Metabolic Adaptations of Lactobacillus spp. to PPCs
7.1. Nitrogen Metabolism
7.2. Sugar and Energy Metabolism
7.3. Gallic Acid: A Source of Chemiosmotic Energy to Lactobacillus
8. Membrane and Cell-Wall Modifications in Response to PPCs
8.1. Cell-Wall Modifications
8.2. Membrane Modifications
9. Role of PPCs in the Molecular Adaptation of Lactobacillus to Host Environments
9.1. Regulation of Molecular Functions Involved in the Survival of Lactobacillus to Gastrointestinal (GI) Tract Stress
9.1.1. Gallic Acid
9.1.2. Tannic Acid
9.1.3. p-Coumaric Acid (p-CA)
9.1.4. Resveratrol
9.2. Remodeling of the Cell Envelope Induced by PPCs: Potential Impact on the Communication Capacities of Lactobacillus with the Host
9.2.1. Capsular Polysaccharides
9.2.2. Teichoic Acids
9.2.3. Surface and Moonlighting Proteins
10. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bonaccio, M.; Pounis, G.; Cerletti, C.; Donati, M.B.; Iacoviello, L.; de Gaetano, G. MOLI-SANI Study Investigators. Mediterranean diet, dietary polyphenols and low grade inflammation: Results from the MOLI-SANI study. Br. J. Clin. Pharmacol. 2017, 83, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Puupponen-Pimia, R.; Dauer, A.; Aura, A.M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Res. 2009, 53, 310–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 781–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hervert-Hernandez, D.; Goñi, I. Dietary polyphenols and human gut microbiota: A review. Food Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Valdés, L.; Cuervo, A.; Salazar, N.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. The relationship between phenolic compounds from diet and microbiota: Impact on human health. Food Funct. 2015, 6, 2424–2439. [Google Scholar] [CrossRef]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly) phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar]
- Curiel, J.A.; Muñoz, R.; López de Felipe, F. pH and dose-dependent effects of quercetin on the fermentation capacity of Lact. plantarum. LWT Food Sci. Technol. 2010, 43, 926–933. [Google Scholar] [CrossRef]
- Choy, Y.Y.; Quifer-Rada, P.; Holstege, D.M.; Frese, S.A.; Calvert, C.C.; Mills, D.A.; Lamuela-Raventos, R.M.; Waterhouse, A.L. Phenolic metabolites and substantial microbiome changes in pig feces by ingesting grape seed proanthocyanidins. Food Funct. 2014, 5, 2298–2308. [Google Scholar] [CrossRef] [PubMed]
- López de Felipe, F.; Curiel, J.A.; Muñoz, R. Improvement of the fermentation performance of Lactobacillus plantarum by the flavanol catechin is uncoupled from its degradation. J. Appl. Microbiol. 2010, 109, 687–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberto, M.R.; Farías, M.E.; Manca De Nadra, M.C. Effect of gallic acid and catechin on Lactobacillus hilgardii 5w growth and metabolism of organic compounds. J. Agric Food Chem. 2001, 49, 4359–4363. [Google Scholar] [CrossRef] [PubMed]
- Parkar, S.G.; Stevenson, D.E.; Skinner, M.A. The potential influence of fruit polyphenols on colonic microflora and human gut health. Int. J. Food Microbiol. 2008, 124, 295–298. [Google Scholar] [CrossRef]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- Bialonska, D.; Ramnani, P.; Kasimsetty, S.G.; Muntha, K.R.; Gibson, G.R.; Ferreira, D. The influence of pomegranate by-product and punicalagins on selected groups of human intestinal microbiota. Int. J. Food Microbiol. 2010, 140, 175–182. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef]
- Rechner, A.R.; Kuhnle, G.; Bremner, P.; Hubbard, G.P.; Moore, K.P.; Rice-Evans, C.A. The metabolic fate of dietary polyphenols in humans. Free Rad. Biol. Med. 2002, 33, 220–235. [Google Scholar] [CrossRef]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Hammes, W.; Hertel, C. The genera Lactobacillus and Carnobacterium. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 320–403. [Google Scholar] [CrossRef]
- Fhoula, I.; Najjari, A.; Turki, Y.; Jaballah, S.; Boudabous, A.; Ouzari, H. Diversity and antimicrobial properties of lactic acid bacteria isolated from rhizosphere of olive trees and desert truffles of Tunisia. Biome. Res. Int. 2013, 2013, 405708. [Google Scholar] [CrossRef] [Green Version]
- Pontonio, E.; Di Cagno, R.; Tarraf, W.; Filannino, P.; De Mastro, G.; Gobbetti, M. Dynamic and assembly of epiphyte and endophyte lactic acid bacteria during the life cycle of Origanum vulgare L. Front. Microbiol. 2018, 9, 1372. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Kao, K.C. Transcriptional analysis of Lactobacillus brevis to N-butanol and ferulic acid stress responses. PLoS ONE 2011, 6, e21438. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Sendra, A.; Landete, J.M.; Alcántara, C.; Zúñiga, M. Response of Lactobacillus casei BL23 to phenolic compounds. J. Appl. Microbiol. 2011, 111, 1473–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverón, I.; de las Rivas, B.; Muñoz, R.; López de Felipe, F. Genome-wide transcriptomic responses of a human isolate of Lactobacillus plantarum exposed to p-coumaric acid stress. Mol. Nutr. Food Res. 2012, 56, 1848–1859. [Google Scholar] [CrossRef] [PubMed]
- Reverón, I.; de las Rivas, B.; Matesanz, R.; Muñoz, R.; López de Felipe, F. Molecular adaptation of Lactobacillus plantarum WCFS1 to gallic acid revealed by genome-scale transcriptomic signature and physiological analysis. Microb. Cell Fact. 2015, 14, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverón, I.; Plaza-Vinuesa, L.; Franch, M.; de las Rivas, B.; Muñoz, R.; López de Felipe, F. Transcriptome-based analysis in Lactobacillus plantarum WCFS1 reveals new insights into resveratrol effects at system level. Mol. Nutr. Food Res. 2018, 62, e1700992. [Google Scholar] [CrossRef]
- Santamaría, L.; Reverón, I.; Plaza-Vinuesa, L.; Oliveros, J.C.; de las Rivas, B.; Muñoz, R.; López de Felipe, F. Oleuropein transcriptionally primes Lactobacillus plantarum to interact with plant hosts. Front. Microbiol. 2019, 10, 2177. [Google Scholar] [CrossRef]
- Reverón, I.; Plaza-Vinuesa, L.; Santamaría, L.; Oliveros, J.C.; de las Rivas, B.; Muñoz, R.; López de Felipe, F. Transcriptomic evidence of molecular mechanisms underlying the response of Lactobacillus plantarum WCFS1 to hydroxytyrosol. Antioxidants 2020, 9, 442. [Google Scholar] [CrossRef]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. (2007–2015). Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 25 July 2020).
- Simić, A.; Manojlović, D.; Segan, D.; Todorović, M. Electrochemical behavior and antioxidant and prooxidant activity of natural phenolics. Molecules 2007, 12, 2327–2340. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Schindeldecker, M.; Moosmann, B. Protein-borne methionine residues as structural antioxidants in mitochondria. Amino Acids 2015, 47, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Gatz, S.; Wiesmüller, L. Take a break-resveratrol in action on DNA. Carcinogenesis 2008, 29, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yin, J.; Liu, J.; Xu, Q.; Lan, T.; Ren, F.; Hao, Y. The copper homeostasis transcription factor CopR is involved in H2O2 stress in Lactobacillus plantarum CAUH2. Front. Microbiol. 2017, 8, 2015. [Google Scholar] [CrossRef] [Green Version]
- Groot, M.N.; Klaassens, E.; de Vos, W.M.; Delcour, J.; Hols, P.; Kleerebezem, M. Genome-based in silico detection of putative manganese transport systems in Lactobacillus plantarum and their genetic analysis. Microbiology 2005, 151, 1229–1238. [Google Scholar] [CrossRef]
- Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Anglade, P.; Baraige, F.; Zagorec, M.; Champomier-Vergès, M.; Muñoz, R.; López de Felipe, F. Response of a Lactobacillus plantarum human isolate to tannic acid challenge assessed by proteomic analyses. Mol. Nutr. Food Res. 2011, 55, 1454–1465. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, A.R.; Alkema, W.; Starrenburg, M.J.; Hugenholtz, J.; van Hijum, S.A.; Bron, P.A. Strain-dependent transcriptome signatures for robustness in Lactococcus lactis. PLoS ONE 2016, 11, e0167944. [Google Scholar] [CrossRef]
- Shatalin, K.; Shatalina, E.; Mironov, A.; Nudler, E. H2S: A universal defense against antibiotics in bacteria. Science 2011, 334, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Matoba, Y.; Yoshida, T.; Izuhara-Kihara, H.; Noda, M.; Sugiyama, M. Crystallographic and mutational analyses of cystathionine β-synthase in the H2S-synthetic gene cluster in Lactobacillus plantarum. Protein Sci. 2017, 26, 763–783. [Google Scholar] [CrossRef]
- Matoba, Y.; Noda, M.; Yoshida, T.; Oda, K.; Ezumi, Y.; Yasutake, C.; Izuhara-Kihara, H.; Danshiitsoodol, N.; Kumaga, T.; Sugiyama, M. Catalytic specificity of the Lactobacillus plantarum cystathionine γ-lyase presumed by the crystallographic analysis. Sci. Rep. 2020, 10, 14886. [Google Scholar] [CrossRef]
- Roca, A.I.; Cox, M.M. The RecA protein, structure and function. Crit. Rev. Bioch. Mol. Biol. 1990, 25, 415–456. [Google Scholar] [CrossRef]
- Cecconi, D.; Cristofoletti, M.; Milli, A.; Antonioli, P.; Rinalducci, S.; Zolla, L.; Zapparoli, G. Effect of tannic acid on Lactobacillus plantarum wine strain during starvation: A proteomic study. Electrophoresis 2009, 30, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Bossi, A.; Rinalducci, S.; Zolla, L.; Antonioli, P.; Righetti, P.G.; Zapparoli, G. Effect of tannic acid on Lactobacillus hilgardii analysed by a proteomic approach. J. Appl. Microbiol. 2007, 102, 787–795. [Google Scholar] [CrossRef]
- Mazzeo, M.F.; Lippolis, R.; Sorrentino, A.; Liberti, S.; Fragnito, F.; Siciliano, R.A. Lactobacillus acidophilus-rutin interplay investigated by proteomics. PLoS ONE 2015, 10, e0142376. [Google Scholar]
- Zolkiewski, M. A camel passes through the eye of a needle: Protein unfolding activity of Clp ATPases. Mol. Microbiol. 2006, 61, 1094–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Cao, C.; Zhang, J.; Kwok, L.Y.; Zhang, H.; Chen, Y. Lactobacillus casei asp23 gene contributes to gentamycin resistance via regulating specific membrane-associated proteins. J. Dairy Sci. 2018, 101, 1915–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauryliuk, V.; Atkinson, G.C.; Murakami, K.S.; Tenson, T.; Gerdes, K. Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nat. Rev. Microbiol. 2015, 13, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Kriel, A.; Bittner, A.N.; Kim, S.H.; Liu, K.; Tehranchi, A.K.; Zou, W.Y.; Rendon, S.; Chen, R.; Tu, B.P.; Wang, J.D. Direct regulation of GTP homeostasis by (p)ppGpp: A critical component of viability and stress resistance. Mol. Cell 2012, 48, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Landete, J.M.; Rodríguez, H.; Curiel, J.A.; de las Rivas, B.; López de Felipe, F.; Muñoz, M. Degradation of phenolic compounds found in olive products by Lactobacillus plantarum strains. In Olives and Olive Oil in Health and Disease Prevention, 2nd ed.; Victor, R., Preedy, R.R.W., Eds.; Academic Press: London, UK, 2021; pp. 133–144. ISBN 9780128195284. [Google Scholar]
- van Veen, H.W.; Venema, K.; Bolhuis, H.; Oussenko, I.; Kok, J.; Poolman, B.; Driessen, A.J.; Konings, W.N. Multidrug resistance mediated by a bacterial homolog of the human multidrug transporter MDR1. Proc. Natl. Acad. Sci. USA 1996, 93, 10668–10672. [Google Scholar] [CrossRef] [Green Version]
- Revilla-Guarinos, A.; Gebhard, S.; Mascher, T.; Zúñiga, M. Defence against antimicrobial peptides: Different strategies in Firmicutes. Environ. Microbiol. 2014, 16, 1225–1237. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, J.; Tian, X.L.; Versey, J.; Wishart, A.; Li, Y.H. The BceABRS four-component system regulates the bacitracin-induced cell envelope stress response in Streptococcus mutans. Antimicrob. Agents Chemother. 2010, 54, 3895–3906. [Google Scholar] [CrossRef] [Green Version]
- Groot Kormelink, T.; Koenders, E.; Hagemeijer, Y.; Overmars, L.; Siezen, R.J.; de Vos, W.M.; Francke, C. Comparative genome analysis of central nitrogen metabolism and its control by GlnR in the class Bacilli. BMC Genom. 2012, 13, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boogerd, F.C.; Ma, H.; Bruggeman, F.J.; van Heeswijk, W.C.; García-Contreras, R.; Molenaar, D.; Krab, K.; Westerhoff, H.V. AmtB-mediated NH3 transport in prokaryotes must be active and as a consequence regulation of transport by GlnK is mandatory to limit futile cycling of NH4(+)/NH3. FEBS Lett. 2011, 585, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.C.; van der Donk, W.A. The many roles of glutamate in metabolism. J. Ind. Microbiol. Biotechnol. 2016, 43, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, J.D.; Herskovits, A.A.; O’ Riordan, M.X.D. Metabolism of the Gram-Positive bacterial Pathogen Listeria monocytogenes. Microbiol. Spectr. 2019, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Rajendran, R.; Zhao, Y.; Tan, B.; Wu, G.; Bazer, F.W.; Zhu, G.; Peng, Y.; Huang, X.; Deng, J.; et al. Amino acids as mediators of metabolic cross talk between host and pathogen. Front. Immunol. 2018, 9, 319. [Google Scholar] [CrossRef] [PubMed]
- Plumed-Ferrer, C.; Koistinen, K.M.; Tolonen, T.L.; Lehesranta, S.J.; Kärenlampi, S.O.; Mäkimattila, E.; Joutsjoki, V.; Virtanen, V.; von Wright, A. Comparative study of sugar fermentation and protein expression patterns of two Lactobacillus plantarum strains grown in three different media. Appl. Environ. Microbiol. 2008, 74, 5349–5358. [Google Scholar] [CrossRef] [Green Version]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; de Angelis, M.; Gobbetti, M.; di Cagno, R. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Appl. Environ. Microbiol. 2014, 80, 2206–2215. [Google Scholar] [CrossRef] [Green Version]
- Kvint, K.; Nachin, L.; Diez, A.; Nystrom, T. The bacterial universal stress protein: Function and regulation. Curr. Opin. Microbiol. 2003, 6, 140–145. [Google Scholar] [CrossRef]
- Chapot-Chartier, M.P.; Kulakauskas, S. Cell wall structure and function in lactic acid bacteria. Microb. Cell Fact. 2014, 29, S9. [Google Scholar] [CrossRef] [Green Version]
- Rohde, M. The Gram-Positive bacterial cell wall. Microbiol. Spectrum 2019, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Reverón, I.; Rodríguez, H.; Campos, G.; Curiel, J.A.; Ascaso, C.; Carrascosa, A.V.; Prieto, A.; de las Rivas, B.; Muñoz, R.; López de Felipe, F. Tannic acid-dependent modulation of selected Lactobacillus plantarum traits linked to gastrointestinal survival. PLoS ONE 2013, 11, e66473. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. p-Coumaric acid kills bacteria through dual damage mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Mainardi, J.L.; Villet, R.; Bugg, T.D.; Mayer, C. Evolution of peptidoglycan biosynthesis under the selective pressure of antibiotics in Gram-positive bacteria. FEMS Microbiol. Rev. 2008, 32, 386–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boekhorst, J.; Wels, M.; Kleerebezem, M.; Siezen, R.J. The predicted secretome of Lactobacillus plantarum WCFS1 sheds light on interactions with its environment. Microbiology 2006, 152, 3175–3183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bron, P.A.; Tomita, S.; van Swam, I., I; Remus, D.M.; Meijerink, M.; Wels, M.; Okada, S.; Wells, J.M.; Kleerebezem, M. Lactobacillus plantarum possesses the capability for wall teichoic acid backbone alditol switching. Microb. Cell Fact. 2012, 11, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.; Taverne., N.I.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M. Impact of 4 Lactobacillus plantarum capsular polysaccharide clusters on surface glycan composition and host cell signaling. Microb. Cell Fact. 2012, 11, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Peng, Z.; Hu, M.; Xiao, Y.S.; Liu, Z.G.; Guan, Q.Q.; Xie, M.Y.; Xiong, T. Interactions between Lactobacillus plantarum NCU116 and its environments based on extracellular proteins and polysaccharides prediction by comparative analysis. Genomics 2020, 112, 3579–3587. [Google Scholar] [CrossRef]
- Parlindungan, E.; May, B.K.; Jones, O.A.H. Metabolic insights into the effects of nutrient stress on Lactobacillus plantarum B21. Front. Mol. Biosci. 2019, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Fozo, E.M.; Kajfasz, J.K.; Quivey, R.G., Jr. Low pH-induced membrane fatty acid alterations in oral bacteria. FEMS Microbiol. Lett. 2004, 238, 291–295. [Google Scholar] [CrossRef]
- van Bokhorst-van de Veen, H.; Abee, T.; Tempelaars, M.; Bron, P.A.; Kleerebezem, M.; Marco, M.L. Short- and long-term adaptation to ethanol stress and its cross-protective consequences in Lactobacillus plantarum. Appl. Environ. Microbiol. 2011, 77, 5247–5256. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; He, J.; Pan, D.; Wu, Z.; Guo., Y.; Zeng, X.; Lian, L. Metabolomics analysis of Lactobacillus plantarum ATCC 14917 adhesion activity under initial acid and alkali stress. PLoS ONE 2018, 13, e0196231. [Google Scholar] [CrossRef] [PubMed]
- Wu, V.C.; Qiu, X.; de los Reyes, B.G.; Lin, C.S.; Pan, Y. Application of cranberry concentrate (Vaccinium macrocarpon) to control Escherichia coli O157:H7 in ground beef and its antimicrobial mechanism related to the down-regulated slp, hdeA and cfa. Food Microbiol. 2009, 26, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Zeng, X.A.; Wang, M.S.; Brennan, C.S.; Gong, D. Modification of membrane properties and fatty acids biosynthesis-related genes in Escherichia coli and Staphylococcus aureus: Implications for the antibacterial mechanism of naringenin. Biochim. Biophys. Acta Biomembr. 2018, 1860, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Rowlett, V.W.; Mallampalli, V.K.P.S.; Karlstaedt, A.; Dowhan, W.; Taegtmeyer, H.; Margolin, W.; Vitrac, H. Impact of membrane phospholipid alterations in Escherichia coli on cellular function and bacterial stress adaptation. J. Bacteriol. 2017, 199, e00849-16. [Google Scholar] [CrossRef] [Green Version]
- Duong, T.; Barrangou, R.; Russell, W.M.; Klaenhammer, T.R. Characterisation of the tre locus and analysis of trehalose cryoprotection in Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2006, 72, 1218–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, N.; Liu, L. Microbial response to acid stress: Mechanisms and applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustos, A.Y.; Font de Valdez, G.; Fadda, S.; Taranto, M.P. New insights into bacterial bile resistance mechanisms: The role of bile salt hydrolase and its impact on human health. Food Res. Int. 2018, 112, 250–262. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef] [Green Version]
- Koskenniemi, K.; Laakso, K.; Koponen, J.; Kankainen, M.; Greco, D.; Auvinen, P.; Savijoki, K.; Nyman, T.A.; Surakka, A.; Salusjärvi, T.; et al. Proteomics and transcriptomics characterization of bile stress response in probiotic Lactobacillus rhamnosus GG. Mol. Cell. Proteomics 2011, 10, M110.002741. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.; Mathesius, U. The role of flavonoids in root–rhizosphere signalling: Opportunities and challenges for improving plant–microbe interactions. J. Exp. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef] [Green Version]
- Oldroyd, G.E.D. Speak, friend, and enter: Signaling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Schandry, N.; Becker, C. Allelopathic plants: Models for studying plant-interkingdom interactions. Trends Plant Sci. 2020, 25, 176–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bove, P.; Russo, P.; Capozzi, V.; Gallone, A.; Spano, G.; Fiocco, D. Lactobacillus plantarum passage through an oro-gastro-intestinal tract simulator: Carrier matrix effect and transcriptional analysis of genes associated to stress and probiosis. Microbiol. Res. 2013, 168, 351–359. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; de Keersmaecker, S.C. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [Green Version]
- Bron, P.A.; Marco, M.L.; Hoffer, S.M.; van Mullekom, E.; de Vos, W.M.; Kleerebezem, M. Genetic characterization of the bile salt response in Lactobacillus plantarum and analysis of responsive promoters in vitro and in situ in the gastrointestinal tract. J. Bacteriol. 2004, 186, 7829–7835. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bron, P.A.; Grangette, C.; Mercenier, A.; de Vos, W.M.; Kleerebezem, M. Identification of Lactobacillus plantarum genes that are induced in the gastrointestinal tract of mice. J. Bacteriol. 2004, 186, 5721–5729. [Google Scholar] [CrossRef] [Green Version]
- Bron, P.A.; Molenaar, D.; de Vos, W.M.; Kleerebezem, M. DNA micro-array-based identification of bile-responsive genes in Lactobacillus plantarum. J. Appl. Microbiol. 2006, 100, 728–738. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.C. Analyzing Global Gene Expression of Lactobacillus plantarum in the Human Gastrointestinal Tract. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2006; p. 147. [Google Scholar]
- Kleerebezem, M.; Hols, P.; Bernard, E.; Rolain, T.; Zhou, M.; Siezen, R.J.; Bron, P.A. The extracellular biology of the lactobacilli. FEMS Microbiol. Rev. 2010, 34, 199–230. [Google Scholar] [CrossRef]
- Van den Nieuwboer, M.; van Hemert, S.; Claassen, E.; de Vos, W.M. Lactobacillus plantarum WCFS1 and its host interaction: A dozen years after the genome. Microb. Biotechnol. 2016, 9, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Marco, M.L.; de Vries, M.C.; Wels, M.; Molenaar, D.; Mangell, P.; Ahrne, S.; de Vos, W.M.; Vaughan, E.E.; Kleerebezem, M. Convergence in probiotic Lactobacillus gut-adaptive responses in humans and mice. ISME J. 2010, 4, 1481–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bron, P.A.; Meijer, M.; Bongers, R.S.; de Vos, W.M.; Kleerebezem, M. Dynamics of competitive population abundance of Lactobacillus plantarum ivi gene mutants in faecal samples after passage through the gastrointestinal tract of mice. J. Appl. Microbiol. 2007, 103, 1424–1434. [Google Scholar] [CrossRef] [PubMed]
- Van Bokhorst-van de Veen, H.; Lee, I.C.; Marco, M.L.; Wels, M.; Bron, P.A.; Kleerebezem, M. Modulation of Lactobacillus plantarum gastrointestinal robustness by fermentation conditions enables identification of bacterial robustness markers. PLoS ONE 2012, 7, e39053. [Google Scholar]
- Heng, J.; Zhao, Y.; Liu, M.; Liu, Y.; Fan, J.; Wang, X.; Zhao, Y.; Zhang., X.C. Substrate-bound structure of the E. coli multidrug resistance transporter MdfA. Cell Res. 2015, 25, 1060–1073. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.M.; Bongers, R.S.; de Vos, W.M.; Kleerebezem, M. Functional analysis of four bile salt hydrolase and penicillin acylase family members in Lactobacillus plantarum WCFS1. Appl. Environ. Microbiol. 2008, 74, 4719–4726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, M.H.; O’Flaherty, S.; Allen, G.; Rivera, A.J.; Stewart, A.K.; Barrangou, R.; Theriot, C.M. Lactobacillus bile salt hydrolase substrate specificity governs bacterial fitness and host colonization. Proc. Natl. Acad. Sci. USA 2021, 118, e2017709118. [Google Scholar] [CrossRef]
- Yao, L.; Seaton, S.C.; Ndousse-Fetter, S.; Adhikari, A.A.; DiBenedetto, N.; Mina, A.I.; Banks, A.S.; Bry, L.; Devlin, A.S. A selective gut bacterial bile salt hydrolase alters host metabolism. eLife 2018, 7, e37182. [Google Scholar] [CrossRef]
- Van Bokhorst-van de Veen, H.; van Swam, I., I; Wels, M.; Bron, P.A.; Kleerebezem, M. Congruent strain specific intestinal persistence of Lactobacillus plantarum in an intestine-mimicking in vitro system and in human volunteers. PLoS ONE 2012, 7, e44588. [Google Scholar] [CrossRef] [Green Version]
- Lebeer, S.; Vanderleyden, J.; de Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Lebeer, S.; Verhoeven, T.L.; Francius, G.; Schoofs, G.; Lambrichts, I.; Dufrêne, Y.; Vanderleyden, J.; de Keersmaecker, S.C. Identification of a gene cluster for the biosynthesis of a long, galactose-rich exopolysaccharide in Lactobacillus rhamnosus GG and functional analysis of the priming glycosyltransferase. Appl. Environ. Microbiol. 2009, 75, 3554–3563. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.C.; Caggianiello, G.; van Swam, I., I; Taverne, N.; Meijerink, M.; Bron, P.A.; Spano, G.; Kleerebezem, M. Strain-specific features of extracellular polysaccharides and their impact on Lactobacillus plantarum-host interactions. Appl. Environ. Microbiol. 2016, 82, 3959–3970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidenmaier, C.; Peschel, A. Teichoic acids and related cell-wall glycopolymers in Gram-positive physiology and host interactions. Nat. Rev. Microbiol. 2008, 6, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, M.; van Hemert, S.; Taverne, N.; Wels, M.; de Vos, P.; Bron, P.A.; Savelkoul, H.F.; van Bilsen, J.; Kleerebezem, M.; Wells, J.M. Identification of genetic loci in Lactobacillus plantarum that modulate the immune response of dendritic cells using comparative genome hybridization. PLoS ONE 2010, 5, e10632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hemmert, S.; Meijerink, M.; Molenaar, D.; Bron, P.A.; de Vos, P.; Kleerebezem, M.; Wells, J.M.; Marco, M.L. Identification of Lactobacillus plantarum genes modulating the cytokine response of human peripheral blood mononuclear cells. BMC Microbiol. 2010, 10, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celebioglu, H.U.; Delsoglio, M.; Brix, S.; Pessione, E.; Svensson, B. Plant polyphenols stimulate adhesion to intestinal mucosa and induce proteome changes in the probiotic Lactobacillus acidophilus NCFM. Mol. Nutr. Food Res. 2018, 62, 4. [Google Scholar] [CrossRef]
- Cui, G.; Wang, J.; Qi, X.; Su, J. Transcription elongation factor GreA plays a key role in cellular invasion and virulence of Francisella tularensis subsp. novicida. Sci. Rep. 2018, 8, 6895. [Google Scholar] [CrossRef]
- Buck, B.L.; Azcarate-Peril, M.A.; Klaenhammer, T.R. Role of autoinducer-2 on the adhesion ability of Lactobacillus acidophilus. J. Appl. Microbiol. 2009, 107, 269–279. [Google Scholar] [CrossRef]
- Wang, W.; Jeffery, C.J. An analysis of surface proteomics results reveals novel candidates for intracellular/surface moonlighting proteins in bacteria. Mol. Biosyst. 2016, 12, 1420–1431. [Google Scholar] [CrossRef]
- Celebioglu, H.U.; Olesen, S.V.; Prehn, K.; Lahtinen, S.J.; Brix, S.; Hachem, M.A.; Svensson, B. Mucin- and carbohydrate-stimulated adhesion and subproteome changes of the probiotic bacterium Lactobacillus acidophilus NCFM. Proteomics 2017, 163, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, K.; Ochiai, A.; Tsubokawa., D.; Ishihara, K.; Yamamoto, Y. Identification and characterization of sulphated carbohydrate-binding protein from Lactobacillus reuteri. PLoS ONE 2013, 8, e83703. [Google Scholar] [CrossRef]
- Espinosa-Urgel, M.; Salido, A.; Ramos, J.L. Genetic analysis of functions involved in adhesion of Pseudomonas putida to seeds. J. Bacteriol. 2000, 182, 2363–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Lu, Y.; Xie, J.; Fei, Y.; Zheng, G.; Wang, Z.; Liu, J.; Lv, L.; Ling, Z.; Berglund, B.; et al. Probiotic gastrointestinal transit and colonization after oral administration: A long journey. Front. Cell. Infect. Microbiol. 2021, 11, 609722. [Google Scholar] [CrossRef] [PubMed]
- Bergonzelli, G.E.; Granato, D.; Pridmore, R.D.; Marvin-Guy, L.F.; Donnicola, D.; Corthesy-Theulaz, I.E. GroEL of Lactobacillus johnsonii La1 (NCC 533) is cell surface associated: Potential role in interactions with the host and the gastric pathogen Helicobacter pylori. Infect. Immun. 2006, 74, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, A.; Russo, P.; Capozzi, V.; Spano, G.; Fiocco, D. Knock out of sHSP genes determines some modifications in the probiotic attitude of Lactiplantibacillus plantarum. Biotechnol Lett. 2020, 43, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Glenting, J.; Holmstrøm, K.; Israelsen, H.; Vrang, A.; Antonsson, M.; Ahrné, S.; Madsen, S.M. Molecular switch controlling expression of the mannose-specific adhesin, msa, in Lactobacillus plantarum. Appl. Environ. Microbiol. 2019, 85, e02954-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
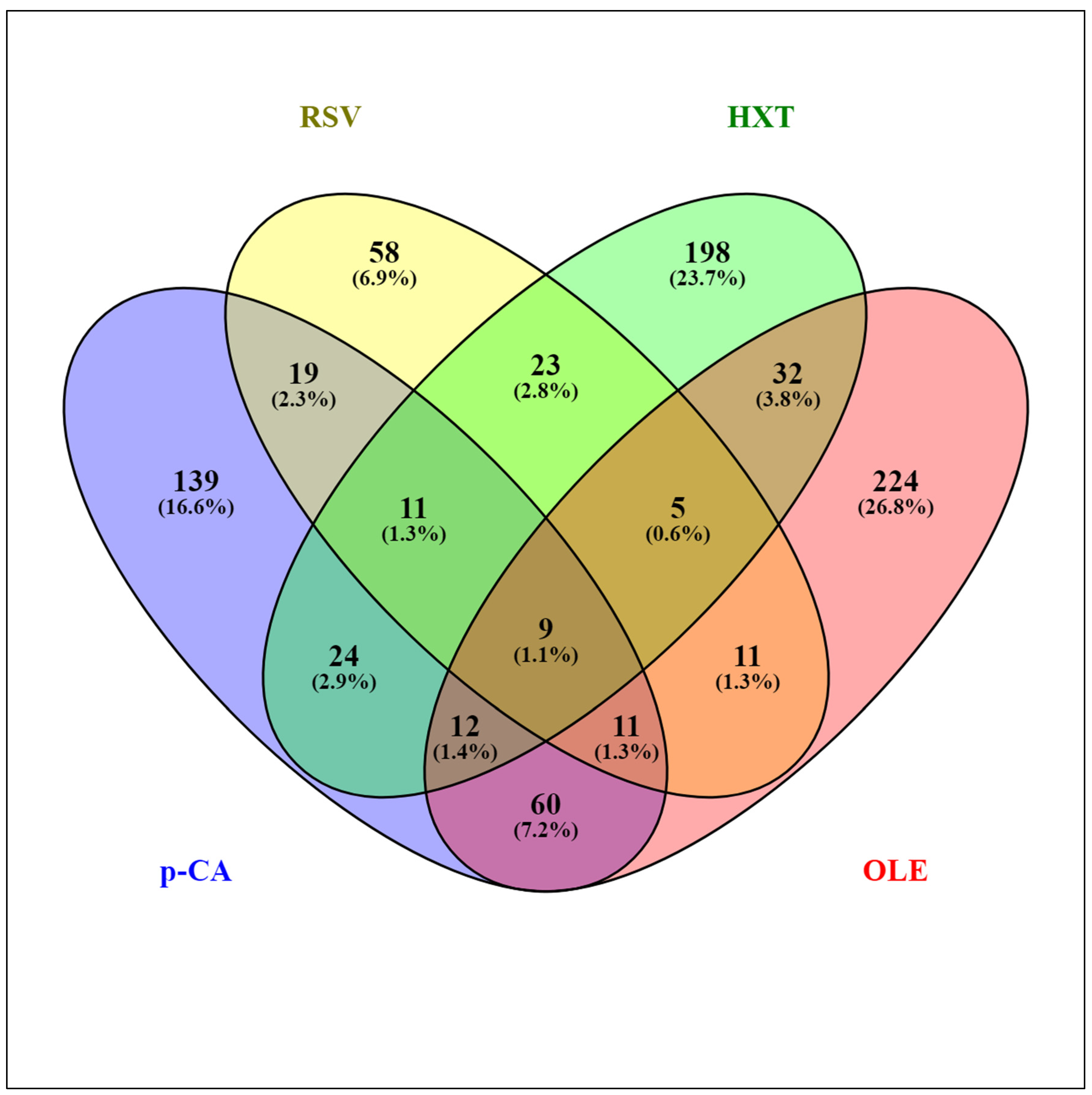
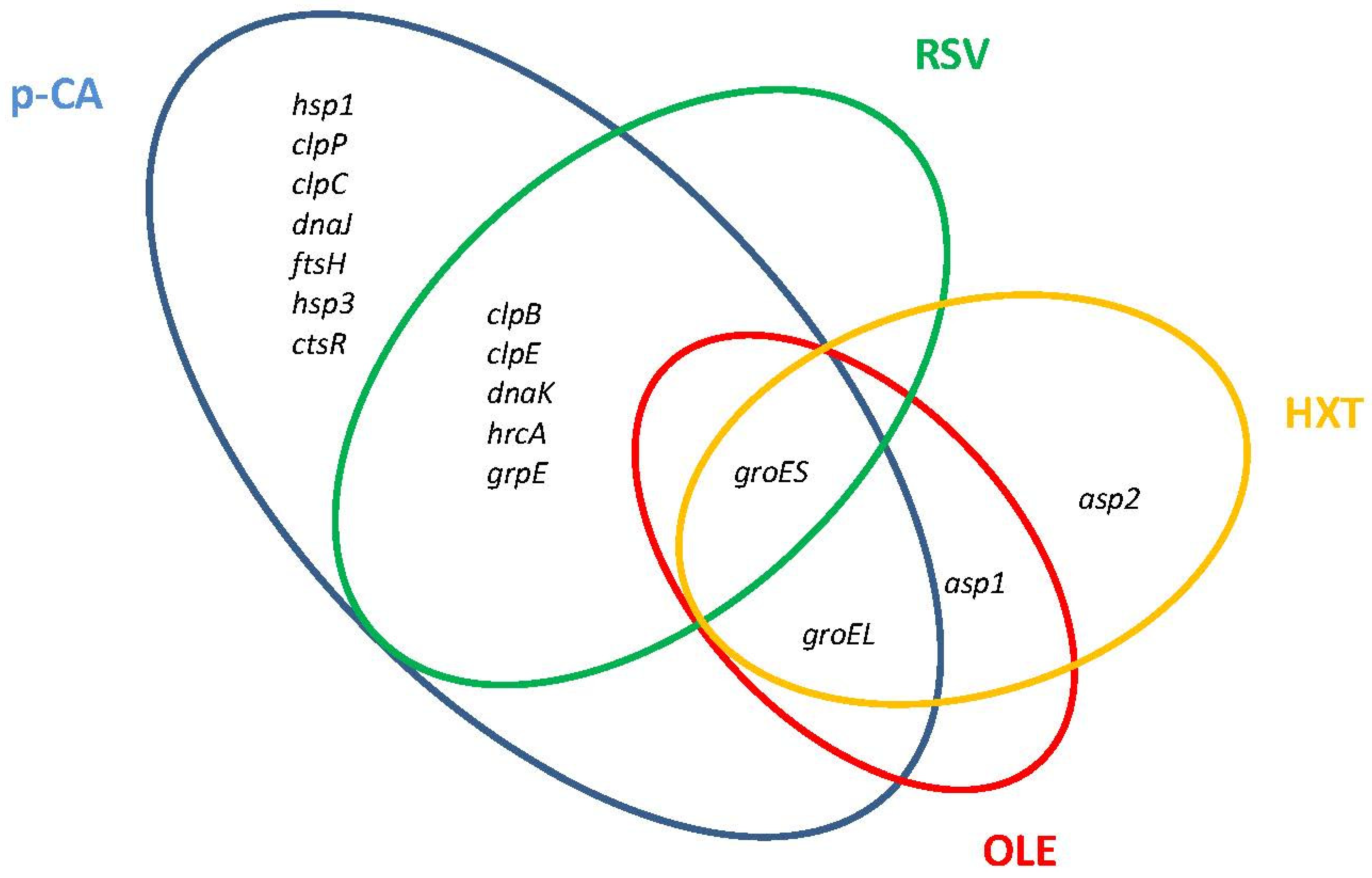
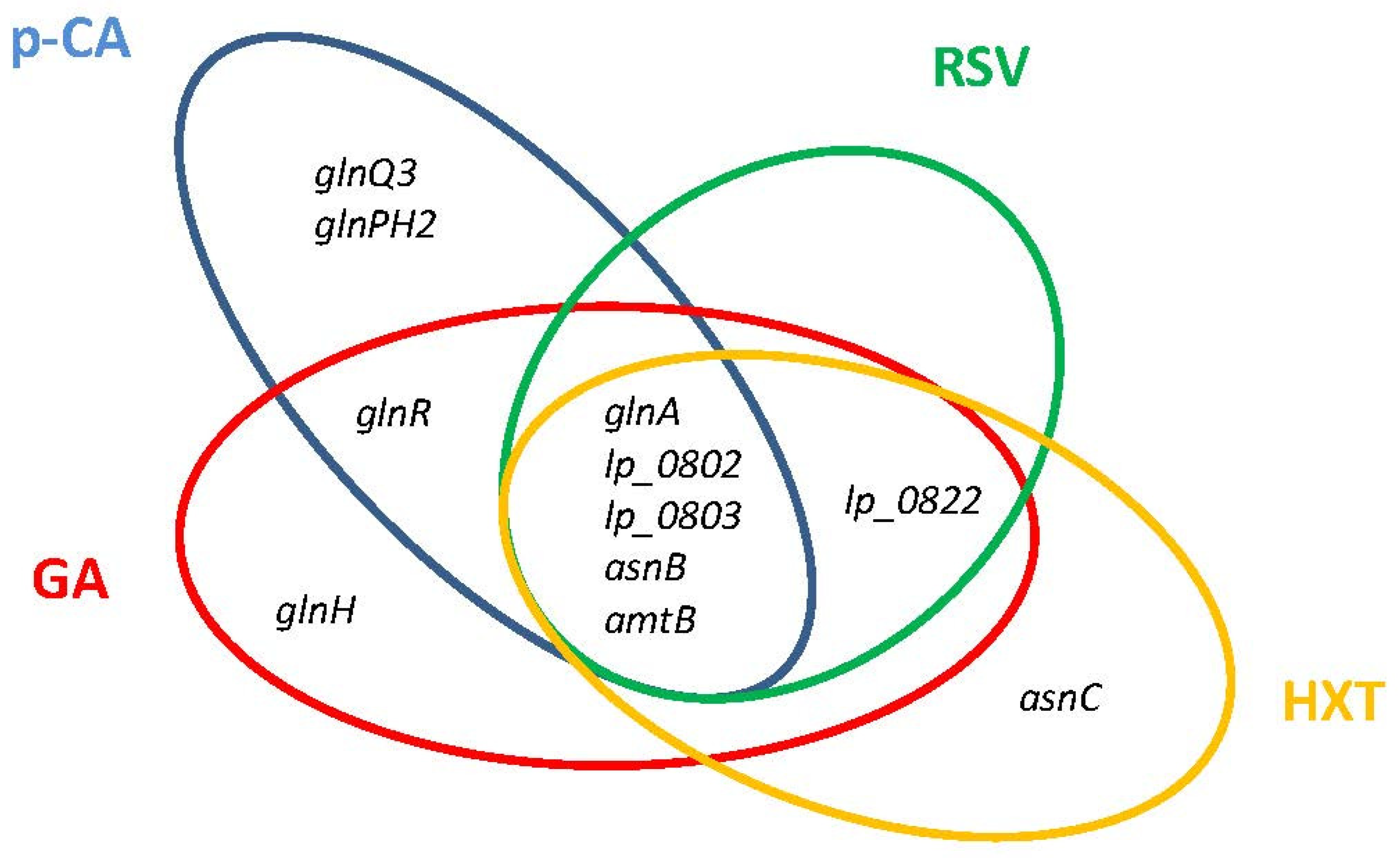

{kind=link}
{kind=link}
{kind=link}
| Phenolic Compound | Molecular Mechanism and Actors Involved | References | ||
|---|---|---|---|---|
| Enzymatic Inactivation of O2 Radicals | Repair of O2 Radical-Induced Damage | Modulation of Metal Transport Participating in the Fenton-Like Chemistry | ||
| Tannic acid | Npr2 | RecA | [37] | |
| p-coumaric acid | gshR2 trxA1, trxA3, trxB1 msrA2, msrA3, msrA4 tpx CBS, CSE | [25] | ||
| Resveratrol | xerC mfd pth CBS, CSE | copR lp_0355 (multicopper oxidase) lp_3302, lp_0463 (zinc ABC transporters) | [27] | |
| Oleuropein | npr2 pox5 (lp_3589) | gshR2 msrA4 lexA CBS, CSE | lp_0463, lp_0464 (zinc ABC transporter) | [28] |
| Hydroxytyrosol | npr2 kat (lp_3578) pox4 (lp_3587) | msrA3 gshR4 lp_0858 CBS, CSE | lp_2992 (mntH3) (iron transporter) | [29] |
| Phenolic Compound | Transporter Type | Locus Tags a and Fold Change b,c | References | |||
|---|---|---|---|---|---|---|
| ABC (BceAB-like) | ||||||
| lp_2739 | lp_2740 | |||||
| GA | −7.2 | −6.7 | [26] | |||
| p-CA | −6.4 | 2.0 | [25] | |||
| RSV | −12.4 | −2.7 | [27] | |||
| OLE | −20.4 | −16.6 | [28] | |||
| HXT | −4.7 | −3.8 | [29] | |||
| MFS | ||||||
| lp_0989 | lp_0990 | lp_0991 | lp_0992 | |||
| p-CA | 3.0 | 19.5 | 17.5 | 16.7 | [25] | |
| RSV | 5.8 | 5.6 | 5.5 | 5.7 | [27] | |
| LmRA-like | ||||||
| lp_2393 | lp_2394 | lp_2395 | ||||
| p-CA | 10.2 | 8.7 | 9.8 | [25] | ||
| RSV | 3.8 | 4.6 | 6.5 | [27] | ||
| Locus Tags | PPC Effector (Ref.) a | Effect | Phenotype Involved (Ref.) b | |
|---|---|---|---|---|
| In vitro orogastric–intestinal (OGI) survival | ||||
| lp_1903 | clpB: ATP-dependent Clp protease ATP-binding subunit | p-CA [24], RSV [27] | + | In vitro OGI survival [86] |
| lp_1269 | clpE: ATP-dependent Clp protease ATP-binding subunit | p-CA, RSV | + | In vitro OGI survival |
| lp_0786 | clpP: ATP-dependent Clp protease, protease subunit | p-CA | + | In vitro OGI survival |
| lp_0547 | ftsH: cell division protease | p-CA | + | |
| lp_2029 | hrcA: heat-inducible transcriptional repressor | p-CA | + | In vitro OGI survival |
| lp_0728 | groEL: chaperonin groEL | p-CA, HXT [29], OLE [28] | + | In vitro OGI survival |
| lp_0727 | groES: chaperonin groES | p-CA, HXT, RSV, OLE, | + | In vitro OGI survival |
| lp_2027 | dnaK: molecular chaperone | p-CA, RSV | + | In vitro OGI survival |
| lp_0129 | hsp1: small heat-shock protein 1 | p-CA | + | In vitro OGI survival |
| lp_3352 | hsp3: small heat-shock protein 3 | p-CA | + | In vitro OGI survival |
| Bile resistance | ||||
| lp_2365 lp_2367 lp_2368 | atpG: F-type H+-transporting ATPase subunit gamma atpH: F-type H+-transporting ATPase subunit delta atpF: F-type H+-transporting ATPase subunit b | HXT | + | Bile resistance [91] |
| lp_1253 | gshR2: glutathione reductase | p-CA, OLE | + | Bile resistance |
| lp_0254 | cysE: serine O-acetyltransferase | HXT, OLE | + | Bile resistance |
| lp_0255 | metC1: cystathionine beta-lyase | HXT, OLE | + | Bile resistance |
| lp_0256 | cysK: cysteine synthase | HXT, OLE | + | Bile resistance |
| Gastro-intestinal tract (GIT) passage | ||||
| lp_2940 | cell surface protein precursor, LPXTG-motif cell-wall anchor | TA [64], GA [26] | + | Human/mouse GIT passage [95] |
| lp_3055 | copA: copper-transporting ATPase | TA [64] | + | Human/mouse GIT passage [95] |
| lp_0775 | argG: argininosuccinate synthase | TA [64] | + | Mouse GIT passage [88,90] |
| lp_3473 | ram2: alpha-l-rhamnosidase | TA [64] | + | Mouse GIT passage [90] |
| PBP2A | transpeptidase–transglycosylase (penicillin binding protein 2A) | TA [64] | - | GIT-robustness [97] |
| lp_1835 | msrA2 protein-methionine-S-oxide reductase | p-CA | + | Maintenance in gut ecosystems [95] |
| lp_1836 | msrA3 protein-methionine-S-oxide reductase | HXT, p-CA | + | Maintenance in gut ecosystems [95] |
| lp_1979 | msrA4 protein-methionine-S-oxide reductase | OLE, p-CA | + | Maintenance in gut ecosystems [95] |
| lp_0991 | multidrug transport protein, major facilitator superfamily (MFS) | p-CA, RSV | + | Human small intestine passage [92] |
| lp_0930 | asp2: alkaline shock protein | p-CA | + | Human small intestine passage [92] |
| lp_2034 lp_2035 lp_2037 | tyrA; prephenate dehydrogenase aroE; 3-phosphoshikimate 1-carboxyvinyltransferase aroF; chorismate synthase | p-CA p-CA p-CA | + + + | Human small intestine passage [92] |
| lp_1744 lp_1745 lp_1746 | d-methionine ABC transporter, ATP binding protein d-methionine ABC transporter permease d-methionine ABC transporter, substrate-binding protein | p-CA p-CA p-CA | + + + | Human small intestine passage [92] |
| Other mechanisms | ||||
| Oxidative stress | ||||
| npr2 kat pox trxA1, trxA; trxB1 tpx gshR4 lp_0858 | NADH-peroxidase catalase pyruvate oxidase thioredoxin; thioredoxin reductase thiol peroxidase glutathione reductase redox protein, peroxiredoxin | p-CA, OLE, HXT HXT OLE, HXT p-CA p-CA HXT HXT | + + + + + + | Resistance against oxidative stress at mucosal surface of the colon (putative) [94] |
| Peptidoglycan remodeling | ||||
| LDH-D | d-lactate dehydrogenase | TA [37] Rutin [45] | + + | Maintaining integrity of the cell envelope under GIT stress (bile, osmotic and acid stresses) (putative) [87] |
| DAPF | diaminopimelate (DAP) epimerase | TA [37] | + | |
| GLMU | UDP-N-acetylglucosamine pyrophosphorylase | Rutin [45] | + | |
| aad | d-alanyl-d-alanine dipeptidase | p-CA | + | |
| hicD2 | d-hydroxyisocaproate dehydrogenase | p-CA | + | |
| lp_0302;lp_3014; lp_3015 | lytic transglycosylases | p-CA | + − | |
| Compatible solutes | ||||
| lp_3324 glpF4 pts5ABC | glycine betaine/carnitine/choline transport protein glycerol uptake facilitator protein PTS system specific- trehalose transporter | p-CA, HXT p-CA HXT | − − − | Osmoprotection against osmotic stress in the GIT (putative) [87,94] |
| Locus Tags | Species | PPC Effector (Ref.) a | Effect | Phenotype Involved (Ref.) b | |
|---|---|---|---|---|---|
| Surface polysaccharide biosynthesis | |||||
| lp_1177 through lp_1185 | cps1A-I: polysaccharide biosynthesis gene cluster 1 proteins | L. plantarum | p-CA [25] | − | Reduced SPS levels [69,104,105] |
| lp_1220-lp_1221 | cps3DE: polysaccharide biosynthesis gene cluster 3 proteins | p-CA | − | Reduced SPS levels | |
| lp_2101-lp_2103; lp_2106-lp_2108 | cps4HGF: polysaccharide biosynthesis gene cluster 4 proteins | p-CA | − | Reduced SPS levels | |
| lp_2101-lp_2102; lp_2106 | cps4HG; cps4C: polysaccharide biosynthesis gene cluster 4 proteins | RSV [27] | − | Reduced SPS levels | |
| lp_1180; lp_1184-lp_1185 | cps1D; cps1H-I: polysaccharide biosynthesis gene cluster 1 proteins | OLE [28] | − | Reduced SPS levels | |
| lp_1197- lp_1198 | cps2AB: polysaccharide biosynthesis gene cluster 2 proteins | OLE | − | Reduced SPS levels | |
| lp_1201- lp_1203; lp_1207 | cps2EFG; cps2K: polysaccharide biosynthesis gene cluster 2 proteins | OLE | − | Reduced SPS levels | |
| lp_1215; lp_1222-lp_1224 | cps3A; cps3FG: polysaccharide biosynthesis gene cluster 3 proteins | OLE | − | Reduced SPS levels | |
| lp_2099-lp_2104; lp_2106-lp_2108 | cps4JIH; cps4GFE: polysaccharide biosynthesis gene cluster 4 proteins | OLE | − | Reduced SPS levels | |
| Teiochoic acid biosynthesis | |||||
| lp_2844 | tagE6: poly(glycerol-phosphate) alpha-glucosyltransferase | L. plantarum | HXT [29], OLE | + | Glycosyl substitution of WTA [68] |
| TagE6 | Poly(glycerol-phosphate) alpha-glucosyltransferase | TA [37] | + | Glycosyl substitution of WTA [68] | |
| lp_1816 | tarI: d-ribitol-5-phosphate cytidylyltransferase | OLE | + | Synthesis of alternative WTA variants [68] | |
| lp_1817 | tarJ: ribitol-5-phosphate 2-dehydrogenase | OLE | + | Synthesis of alternative WTA variants [68] | |
| lp_1818 | tarK: ribitolphosphotransferase | OLE, HXT | + | Synthesis of alternative WTA variants [99] | |
| lp_1372 | gtca1: teichoic acid glycosylation protein | HXT | − | LTA glycosylation [107] | |
| Adhesion | |||||
| EF-P | Elongation factor EF-P | L. acidophilus | RSV [109] | + | Adhesion [112] |
| PK | Pyruvate kinase | RSV 109 | + | Adhesion [113] | |
| EF-Tu | Elongation factor EF-Tu | RSV [109] | + | Adhesion [114] | |
| EF-P | Elongation factor EF-P | Caffeic acid [109] | − | Adhesion [112] | |
| PYK | Pyruvate kinase | Caffeic acid [109] | − | Adhesion [113] | |
| EF-Tu | Elongation factor EF-Tu | Caffeic acid [109] | − | Adhesion [114] | |
| lp_0728 | groEL: chaperonin groEL | L. plantarum | p-CA, HXT, OLE | + | Adhesion [117] |
| EF-GreA | Elongation factor EF-GreA | L. plantarum | TA [37] | + | Adhesion [110] |
| LuxS | Autoinducer production protein LuxS | TA [37] | − | Adhesion [111] | |
| lp_1229 | msa: mannose-specific adhesin | L. plantarum | RSV [27] | + | Mannose-specific adhesion [119] |
| lp_1261 | oppA: oligopeptide ABC transporter substrate binding protein | p-CA, OLE | + | Adhesion [115] | |
| lp_0129 | hsp1: small heat-shock protein 1 | p-CA | + | Adhesion [118] | |
| lp_3352 | hsp3: small heat-shock protein 3 | p-CA | + | Adhesion [118] | |
| Immnunomodulation | |||||
| lp_3581a | lamD: auto-inducing peptide (AIP) precursor | L. plantarum | OLE | − | Immunomodulatory capacity [107] |
| lp_3581 | lamC: two-component system, LytTR family | OLE | − | Immunomodulatory capacity [107] | |
| lp_0422 | plnE: bacteriocin precursor peptide | OLE | − | Immunomodulatory capacity [107] | |
| lp_0421 | plnF: bacteriocin precursor peptide | OLE | Immunomodulatory capacity [107] | ||
| lp_0419 | plnI: bacteriocin immunity protein | OLE | Immunomodulatory capacity [107] | ||
| lp_0423 | plnG: bacteriocin ABC transporter, ATP-binding and permease protein | OLE | − | Immunomodulatory capacity [107] | |
| lp_0424 | plnH: bacteriocin ABC transporter, accessory factor | OLE | − | Immunomodulatory capacity [107] | |
| lp_0428 | plnV: membrane protein | OLE | − | Immunomodulatory capacity [107] | |
| lp_2647 | pts19A: PTS system,N-acetylglucosamine/galactosamine-specific EIIA component | OLE | − | Immunomodulatory capacity [108] | |
| lp_2650 | pts19B: PTS system, N-acetylglucosamine-specific EIIB component | OLE | − | Immunomodulatory capacity [108] | |
| lp_2460 | prophage P2b protein 21 | OLE | − | Immunomodulatory capacity [108] | |
| Phenolic Compound | Technique | Species | References |
|---|---|---|---|
| Hydroxycinnamic acids | |||
p-coumaric acid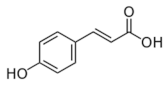 | Transcriptomics Proteomics | L. plantarum L.casei | [25] [24] |
Ferulic acid | Transcriptomics Proteomics | L. brevis L. acidophilus | [23] [109] |
Caffeic acid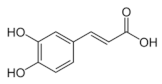 | Proteomics | L. acidophilus | [109] |
| Hydroxybenzoic acids | |||
Gallic acid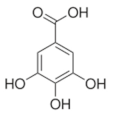 | Transcriptomics | L. plantarum | [26] |
| Stilbenoids | |||
Resveratrol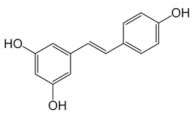 | Transcriptomics Proteomics | L. plantarum L. acidophilus | [27] [109] |
| Phenylethanoid | |||
Hydroxytyrosol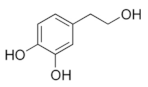 | Transcriptomics | L. plantarum | [29] |
| Flavonoids | |||
Rutin | Proteomics | L. acidophilus | [45] |
| Secoiridoids | |||
Oleuropein | Transcriptomics | L. plantarum | [28] |
| Tannins | |||
Tannic acid | Proteomics Differential gene expression | L. plantarum L. plantarum | [37,43,44] [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López de Felipe, F.; de las Rivas, B.; Muñoz, R. Molecular Responses of Lactobacilli to Plant Phenolic Compounds: A Comparative Review of the Mechanisms Involved. Antioxidants 2022, 11, 18. https://doi.org/10.3390/antiox11010018
López de Felipe F, de las Rivas B, Muñoz R. Molecular Responses of Lactobacilli to Plant Phenolic Compounds: A Comparative Review of the Mechanisms Involved. Antioxidants. 2022; 11(1):18. https://doi.org/10.3390/antiox11010018
Chicago/Turabian StyleLópez de Felipe, Félix, Blanca de las Rivas, and Rosario Muñoz. 2022. "Molecular Responses of Lactobacilli to Plant Phenolic Compounds: A Comparative Review of the Mechanisms Involved" Antioxidants 11, no. 1: 18. https://doi.org/10.3390/antiox11010018
APA StyleLópez de Felipe, F., de las Rivas, B., & Muñoz, R. (2022). Molecular Responses of Lactobacilli to Plant Phenolic Compounds: A Comparative Review of the Mechanisms Involved. Antioxidants, 11(1), 18. https://doi.org/10.3390/antiox11010018