
Potential Implications of Rimonabant on Age-Related Oxidative Stress and Inflammation






 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Protocol
2.2. Determination of Cardiac HO Activity
2.3. Determination of Cardiac ROS, HO-1, NF-κB, and TNF-α Concentrations
2.4. Measurement of Cardiac Total (GSH+GSSG) Level
2.5. Determination of Serum Superoxide Dismutase Activity
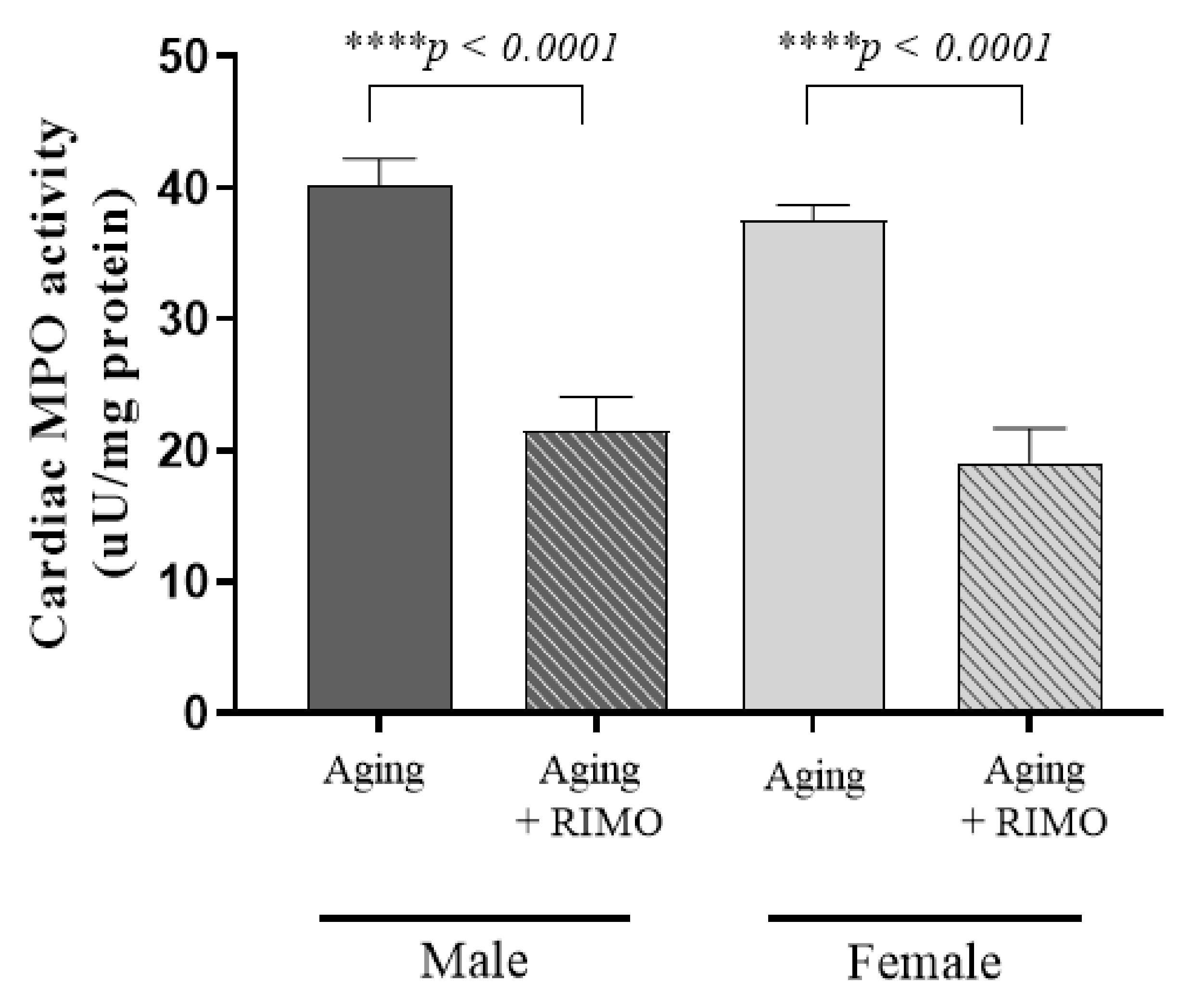
2.6. Determination of Cardiac MPO Activity
2.7. Protein Determination
2.8. Statistical Analysis
3. Results
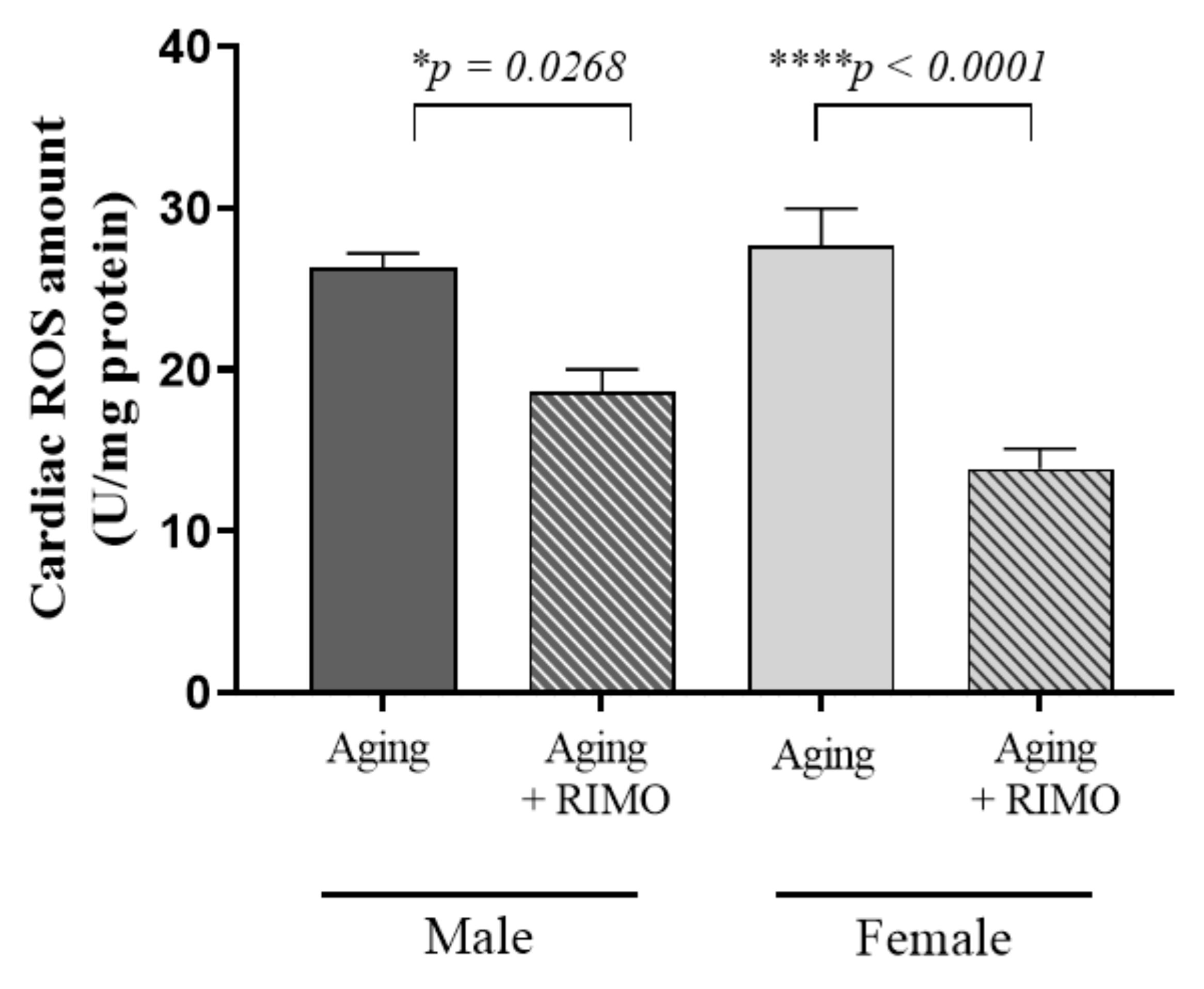
3.1. Determination of Cardiac ROS Concentration
3.2. Measurement of Cardiac HO-1 Concentration and HO Activity
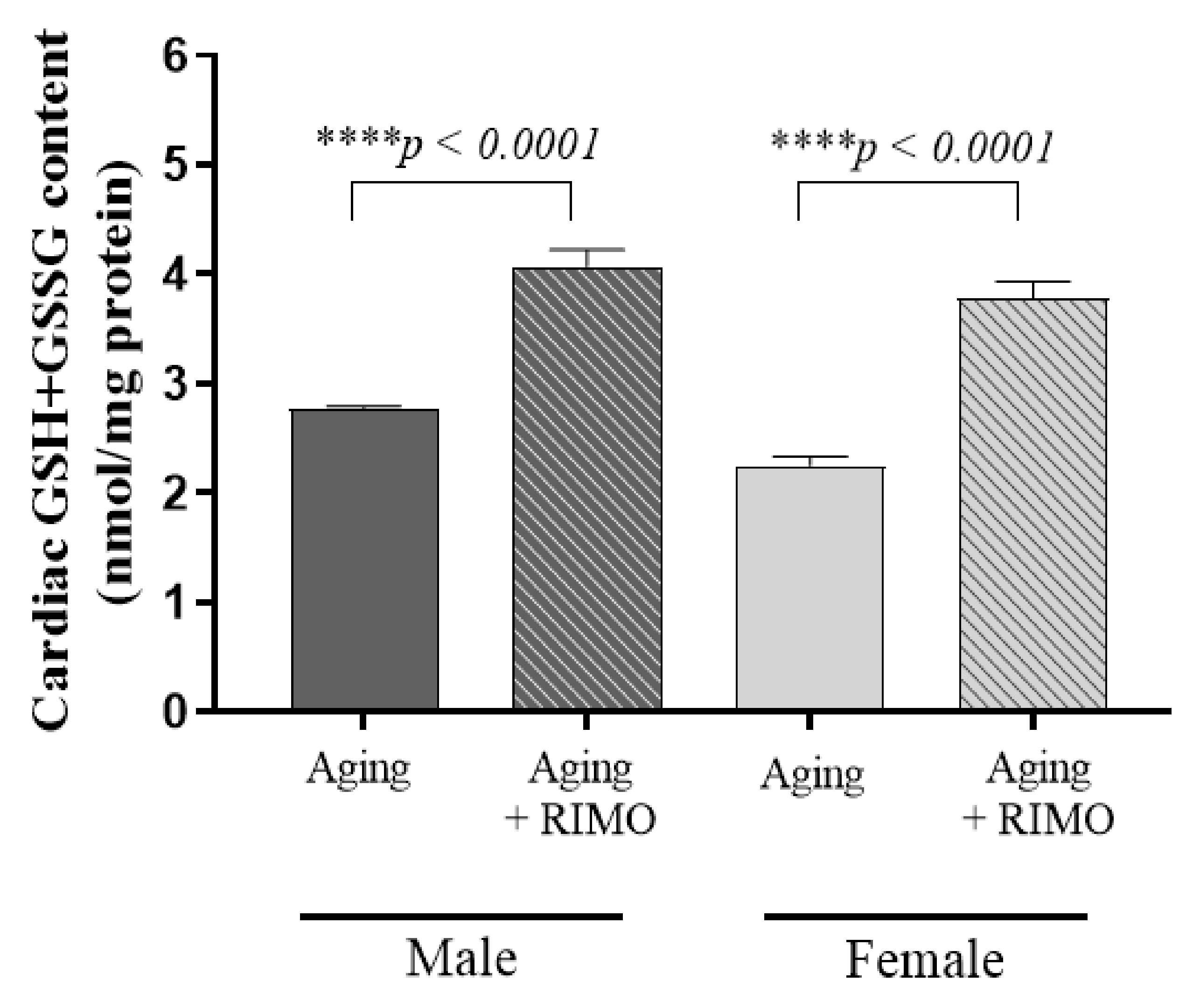
3.3. Determination of Cardiac Total (GSH+GSSG) Content
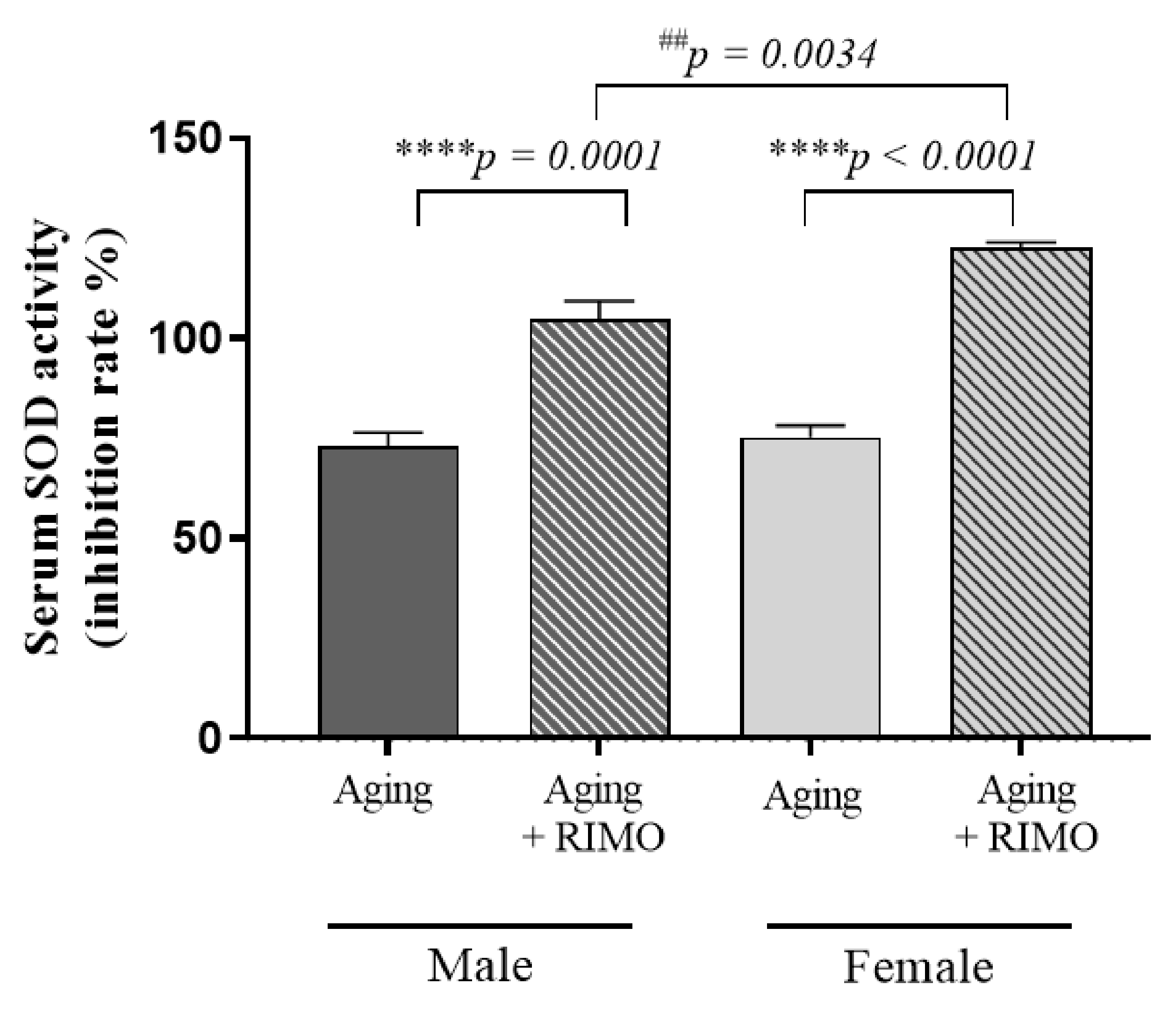
3.4. Determination of Serum SOD Activity
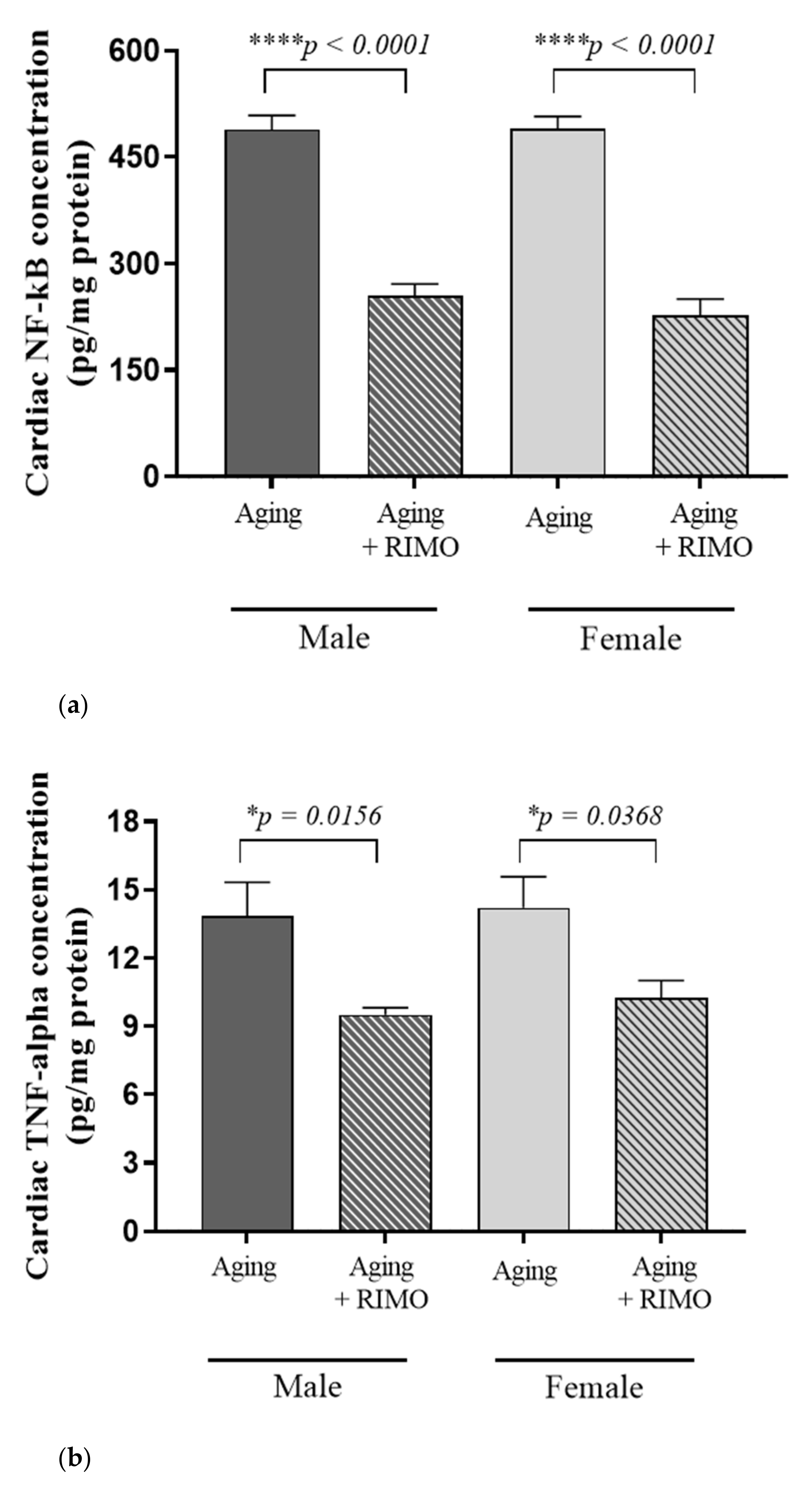
3.5. Measurement of Cardiac NF-κB and TNF-α Concentrations
3.6. Determination of Cardiac MPO Activity
3.7. Assessment of Body Weight Changes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, A.J.P.O.; de Almeida Rezende, M.S.; Dantas, S.H.; de Lima Silva, S.; de Oliveira, J.C.P.L.; de Lourdes Assuncao Araujo de Azevedo, F.; Alves, R.; de Menezes, G.M.S.; Dos Santos, P.F.; Goncalves, T.A.F.; et al. Unveiling the Role of Inflammation and Oxidative Stress on Age-Related Cardiovascular Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 1954398. [Google Scholar] [CrossRef]
- Moris, D.; Spartalis, M.; Spartalis, E.; Karachaliou, G.S.; Karaolanis, G.I.; Tsourouflis, G.; Tsilimigras, D.I.; Tzatzaki, E.; Theocharis, S. The role of reactive oxygen species in the pathophysiology of cardiovascular diseases and the clinical significance of myocardial redox. Ann. Transl. Med. 2017, 5, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredenburgh, L.E.; Merz, A.A.; Cheng, S. Haeme oxygenase signalling pathway: Implications for cardiovascular disease. Eur. Heart J. 2015, 36, 1512–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puhl, S.L. Cannabinoid-sensitive receptors in cardiac physiology and ischaemia. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118462. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Mohanraj, R.; Batkai, S.; Pacher, P. CB1 cannabinoid receptor inhibition: Promising approach for heart failure? Congest. Heart Fail. 2008, 14, 330–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dol-Gleizes, F.; Paumelle, R.; Visentin, V.; Mares, A.M.; Desitter, P.; Hennuyer, N.; Gilde, A.; Staels, B.; Schaeffer, P.; Bono, F. Rimonabant, a selective cannabinoid CB1 receptor antagonist, inhibits atherosclerosis in LDL receptor-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Tam, J.; Vemuri, V.K.; Liu, J.; Batkai, S.; Mukhopadhyay, B.; Godlewski, G.; Osei-Hyiaman, D.; Ohnuma, S.; Ambudkar, S.V.; Pickel, J.; et al. Peripheral CB1 cannabinoid receptor blockade improves cardiometabolic risk in mouse models of obesity. J. Clin. Investig. 2010, 120, 2953–2966. [Google Scholar] [CrossRef] [Green Version]
- Jorgacevic, B.; Mladenovic, D.; Ninkovic, M.; Veskovic, M.; Dragutinovic, V.; Vatazevic, A.; Vucevic, D.; Jesic Vukicevic, R.; Radosavljevic, T. Rimonabant Improves Oxidative/Nitrosative Stress in Mice with Nonalcoholic Fatty Liver Disease. Oxid. Med. Cell. Longev. 2015, 2015, 842108. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, P.; Rajesh, M.; Batkai, S.; Patel, V.; Kashiwaya, Y.; Liaudet, L.; Evgenov, O.V.; Mackie, K.; Hasko, G.; Pacher, P. CB1 cannabinoid receptors promote oxidative stress and cell death in murine models of doxorubicin-induced cardiomyopathy and in human cardiomyocytes. Cardiovasc. Res. 2010, 85, 773–784. [Google Scholar] [CrossRef]
- Paradisi, A.; Oddi, S.; Maccarrone, M. The endocannabinoid system in ageing: A new target for drug development. Curr. Drug Targets 2006, 7, 1539–1552. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, S.; Putalun, W.; Vimolmangkang, S.; Phoolcharoen, W.; Shoyama, Y.; Tanaka, H.; Morimoto, S. Enzyme-linked immunosorbent assay for the quantitative/qualitative analysis of plant secondary metabolites. J. Nat. Med. 2018, 72, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, R.; Borzsei, D.; Hoffmann, A.; Lesi, Z.N.; Gesztelyi, R.; Juhasz, B.; Szebeni, G.J.; Osman, J.; Sebestyen, J.; Nagy, A.; et al. Lifestyle-Induced Redox-Sensitive Alterations: Cross-Talk among the RAAS, Antioxidant/Inflammatory Status, and Hypertension. Oxid. Med. Cell. Longev. 2021, 2021, 3080863. [Google Scholar] [CrossRef]
- Borzsei, D.; Priksz, D.; Szabo, R.; Bombicz, M.; Karacsonyi, Z.; Puskas, L.G.; Feher, L.Z.; Radak, Z.; Kupai, K.; Berko, A.M.; et al. Exercise-mitigated sex-based differences in aging: From genetic alterations to heart performance. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H854–H866. [Google Scholar] [CrossRef]
- Szabo, R.; Karacsonyi, Z.; Borzsei, D.; Juhasz, B.; Al-Awar, A.; Torok, S.; Berko, A.M.; Takacs, I.; Kupai, K.; Varga, C.; et al. Role of Exercise-Induced Cardiac Remodeling in Ovariectomized Female Rats. Oxid. Med. Cell. Longev. 2018, 2018, 6709742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, R.; Borzsei, D.; Kupai, K.; Hoffmann, A.; Gesztelyi, R.; Magyarine Berko, A.; Varga, C.; Posa, A. Spotlight on a New Heme Oxygenase Pathway: Testosterone-Induced Shifts in Cardiac Oxidant/Antioxidant Status. Antioxidants 2019, 8, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, A.; Ribeiro, T.P.; de Medeiros, I.A. Aging: Molecular Pathways and Implications on the Cardiovascular System. Oxid. Med. Cell. Longev. 2017, 2017, 7941563. [Google Scholar] [CrossRef] [Green Version]
- Posa, A.; Szabo, R.; Csonka, A.; Veszelka, M.; Berko, A.M.; Barath, Z.; Menesi, R.; Pavo, I.; Gyongyosi, M.; Laszlo, F.; et al. Endogenous Estrogen-Mediated Heme Oxygenase Regulation in Experimental Menopause. Oxid. Med. Cell. Longev. 2015, 2015, 429713. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, X.; Zheng, S.; Khanabdali, R.; Kalionis, B.; Wu, J.; Wan, W.; Tai, X. An Update on Inflamm-Aging: Mechanisms, Prevention, and Treatment. J. Immunol. Res. 2016, 2016, 8426874. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; DELLA-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Duvigneau, J.C.; Esterbauer, H.; Kozlov, A.V. Role of Heme Oxygenase as a Modulator of Heme-Mediated Pathways. Antioxidants 2019, 8, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csiszar, A. Anti-inflammatory effects of resveratrol: Possible role in prevention of age-related cardiovascular disease. Ann. N. Y. Acad. Sci. 2011, 1215, 117–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pósa, A.; Szabó, R.; Kupai, K.; Berkó, A.M.; Veszelka, M.; Szűcs, G.; Börzsei, D.; Gyongyosi, M.; Pávó, I.; Deim, Z.; et al. Cardioprotective Effect of Selective Estrogen Receptor Modulator Raloxifene Are Mediated by Heme Oxygenase in Estrogen-Deficient Rat. Oxid. Med. Cell. Longev. 2017, 2017, 2176749. [Google Scholar] [CrossRef] [Green Version]
- Costa, B. Rimonabant: More than an anti-obesity drug? Br. J. Pharmacol. 2007, 150, 535–537. [Google Scholar] [CrossRef] [Green Version]
- Pagotto, U.; Pasquali, R. Fighting obesity and associated risk factors by antagonising cannabinoid type 1 receptors. Lancet 2005, 365, 1363–1364. [Google Scholar] [CrossRef]
- Rajesh, M.; Batkai, S.; Kechrid, M.; Mukhopadhyay, P.; Lee, W.S.; Horvath, B.; Holovac, E.; Cinar, R.; Liaudet, L.; Mackie, K.; et al. Cannabinoid 1 receptor promotes cardiac dysfunction, oxidative stress, inflammation, and fibrosis in diabetic cardiomyopathy. Diabetes 2012, 61, 716–727. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.; Kim, D.H.; Yang, H.; Lee, D.H.; Bae, S.H.; Park, C.Y. CB1 receptor blockade ameliorates hepatic fat infiltration and inflammation and increases Nrf2-AMPK pathway in a rat model of severely uncontrolled diabetes. PLoS ONE 2018, 13, e0206152. [Google Scholar] [CrossRef]
- Funes, S.; Rios, M.; Fernández-Fierro, A.; Covián, C.; Bueno, S.M.; Riedel, C.A.; Mackern-Oberti, J.P.; Kalergis, A.M. Naturally Derived Heme-Oxygenase 1 Inducers and Their Therapeutic Application to Immune-Mediated Diseases. Front. Immunol. 2020, 11, 1467. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of inflammation by the antioxidant haem oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Satta, S.; Mahmoud, A.M.; Wilkinson, F.L.; Alexander, M.Y.; White, S.J. The Role of Nrf2 in Cardiovascular Function and Disease. Oxid. Med. Cell. Longev. 2017, 2017, 9237263. [Google Scholar] [CrossRef]
- Harvey, C.J.; Thimmulappa, R.K.; Singh, A.; Blake, D.J.; Ling, G.; Wakabayashi, N.; Fujii, J.; Myers, A.; Biswal, S. Nrf2-regulated glutathione recycling independent of biosynthesis is critical for cell survival during oxidative stress. Free Radic. Biol. Med. 2009, 46, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, R.; Hoffmann, A.; Borzsei, D.; Kupai, K.; Veszelka, M.; Berko, A.M.; Pavo, I.; Gesztelyi, R.; Juhasz, B.; Turcsan, Z.; et al. Hormone Replacement Therapy and Aging: A Potential Therapeutic Approach for Age-Related Oxidative Stress and Cardiac Remodeling. Oxid. Med. Cell. Longev. 2021, 2021, 8364297. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.L.; Juang, J.M.; Wang, Y.H.; Hsueh, C.H.; Liang, Y.J.; Lin, J.L.; Tsai, C.T.; Lai, L.P. Rimonabant inhibits TNF-alpha-induced endothelial IL-6 secretion via CB1 receptor and cAMP-dependent protein kinase pathway. Acta Pharmacol. Sin. 2010, 31, 1447–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.M.; Maltagliati, A.J. Nrf2 at the heart of oxidative stress and cardiac protection. Physiol. Genom. 2018, 50, 77–97. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | p Value |
|---|---|
| Interaction | 0.0862 |
| Treatment | 0.3285 |
| Sex | <0.0001 |
| (a) | |
| Source of Variation | p Value |
| Interaction | 0.7257 |
| Sex | 0.551 |
| Treatment | 0.0005 |
| (b) | |
| Source of Variation | p Value |
| Interaction | 0.6506 |
| Sex | 0.9535 |
| Treatment | 0.0074 |
| Source of Variation | p Value |
|---|---|
| Interaction | 0.3079 |
| Sex | 0.0036 |
| Treatment | <0.0001 |
| Source of Variation | p Value |
|---|---|
| Interaction | 0.0217 |
| Sex | 0.0047 |
| Treatment | <0.0001 |
| (a) | |
| Source of Variation | p Value |
| Interaction | 0.4548 |
| Sex Treatment | 0.4931 <0.0001 |
| (b) | |
| Source of Variation | p Value |
| Interaction | 0.8328 |
| Sex | 0.5682 |
| Treatment | 0.0002 |
| Source of Variation | p Value |
|---|---|
| Interaction | 0.9821 |
| Sex | 0.2522 |
| Treatment | <0.0001 |
| Body Weight Changes | |||
|---|---|---|---|
| Groups | Mean Initial Body Weight (g) | Mean Final Body Weight (g) | Changes between the Final and Initial Body Weights (%) |
| Aging male | 644.5 | 635.1 | 1.5 % ↓ |
| Aging male + RIMO | 641.3 | 622.4 | 3 % ↓ |
| Aging female | 317.3 | 321.0 | 1.2 % ↓ |
| Aging female + RIMO | 307.4 | 289.3 | 5.9 % ↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabó, R.; Szabó, Z.; Börzsei, D.; Hoffmann, A.; Lesi, Z.N.; Pálszabó, P.; Pálszabó, A.; Dvorácskó, S.; Gesztelyi, R.; Kupai, K.; et al. Potential Implications of Rimonabant on Age-Related Oxidative Stress and Inflammation. Antioxidants 2022, 11, 162. https://doi.org/10.3390/antiox11010162
Szabó R, Szabó Z, Börzsei D, Hoffmann A, Lesi ZN, Pálszabó P, Pálszabó A, Dvorácskó S, Gesztelyi R, Kupai K, et al. Potential Implications of Rimonabant on Age-Related Oxidative Stress and Inflammation. Antioxidants. 2022; 11(1):162. https://doi.org/10.3390/antiox11010162
Chicago/Turabian StyleSzabó, Renáta, Zsuzsanna Szabó, Denise Börzsei, Alexandra Hoffmann, Zelma Nadin Lesi, Patrícia Pálszabó, Andrea Pálszabó, Szabolcs Dvorácskó, Rudolf Gesztelyi, Krisztina Kupai, and et al. 2022. "Potential Implications of Rimonabant on Age-Related Oxidative Stress and Inflammation" Antioxidants 11, no. 1: 162. https://doi.org/10.3390/antiox11010162
APA StyleSzabó, R., Szabó, Z., Börzsei, D., Hoffmann, A., Lesi, Z. N., Pálszabó, P., Pálszabó, A., Dvorácskó, S., Gesztelyi, R., Kupai, K., Priksz, D., Juhász, B., Altmayer, A., Varga, C., & Pósa, A. (2022). Potential Implications of Rimonabant on Age-Related Oxidative Stress and Inflammation. Antioxidants, 11(1), 162. https://doi.org/10.3390/antiox11010162