
BAP1 Downregulates NRF2 Target Genes and Exerts Anti-Tumorigenic Effects by Deubiquitinating KEAP1 in Lung Adenocarcinoma

 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Chemicals, Antibodies, and Plasmids
2.2. Western Blot Analysis
2.3. Immunoprecipitation
2.4. Generation of Stable Cells by Lentival Transduction
2.5. Fractionation of the Nucleus and the Cytosol
2.6. Firefly Luciferase Assay
2.7. Real-time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.8. Wound Healing Assay
2.9. Cell Migration Assay
2.10. TUNEL Assay
2.11. Animal Experiments
2.12. Hematoxylin and Eosin (H&E) Staining and Immunohistochemistry (IHC)
2.13. Statistics
2.13.1. Statistical Comparison of A549 Cells and A549 Tumor Xenografts
2.13.2. Comparison of Bap1 mRNA Expression between LUAD and Normal Lung Tissues of Humans
2.13.3. Survival Analysis of LUAD Cancer Patients
3. Results
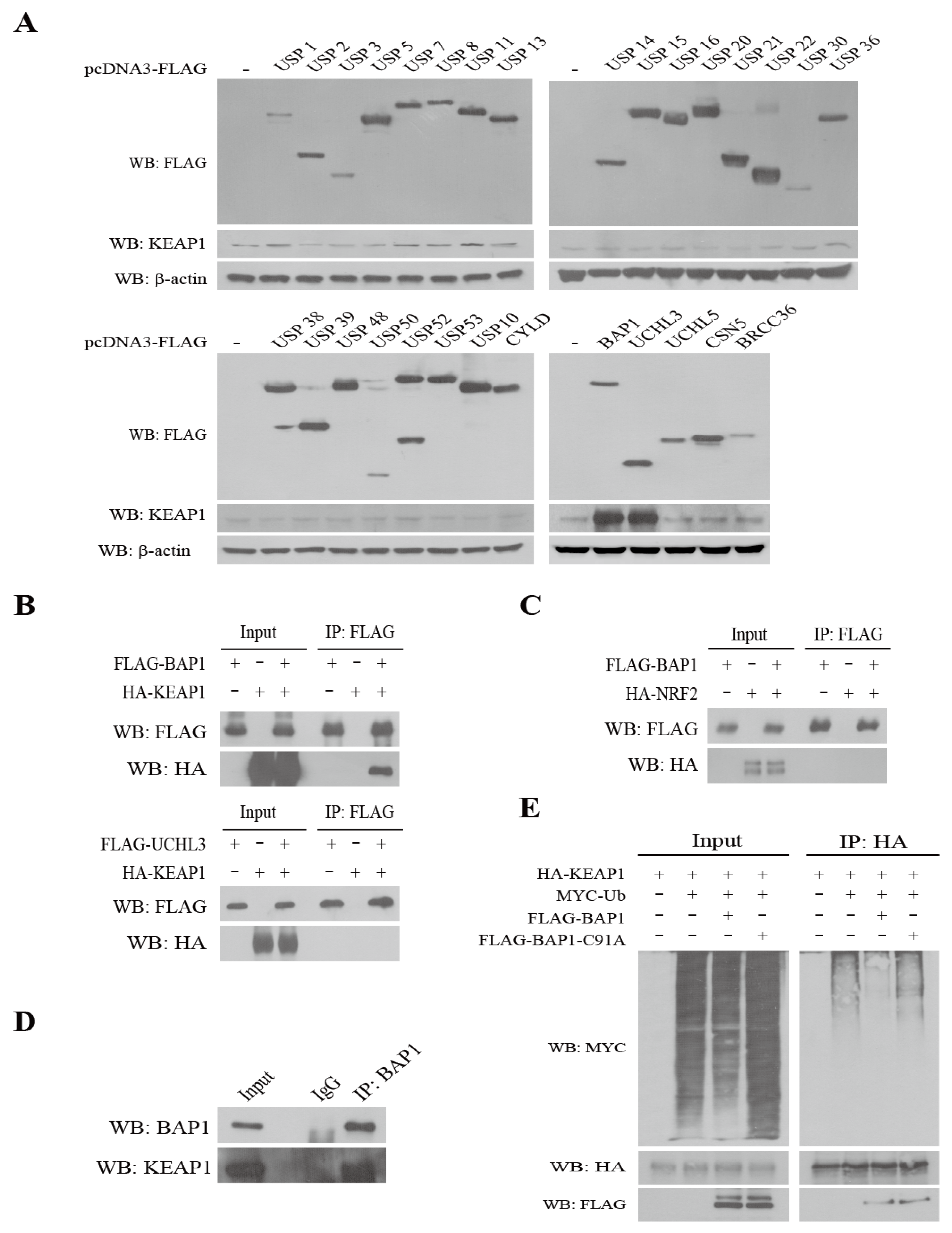
3.1. Identification of BAP1 as a DUB of KEAP1
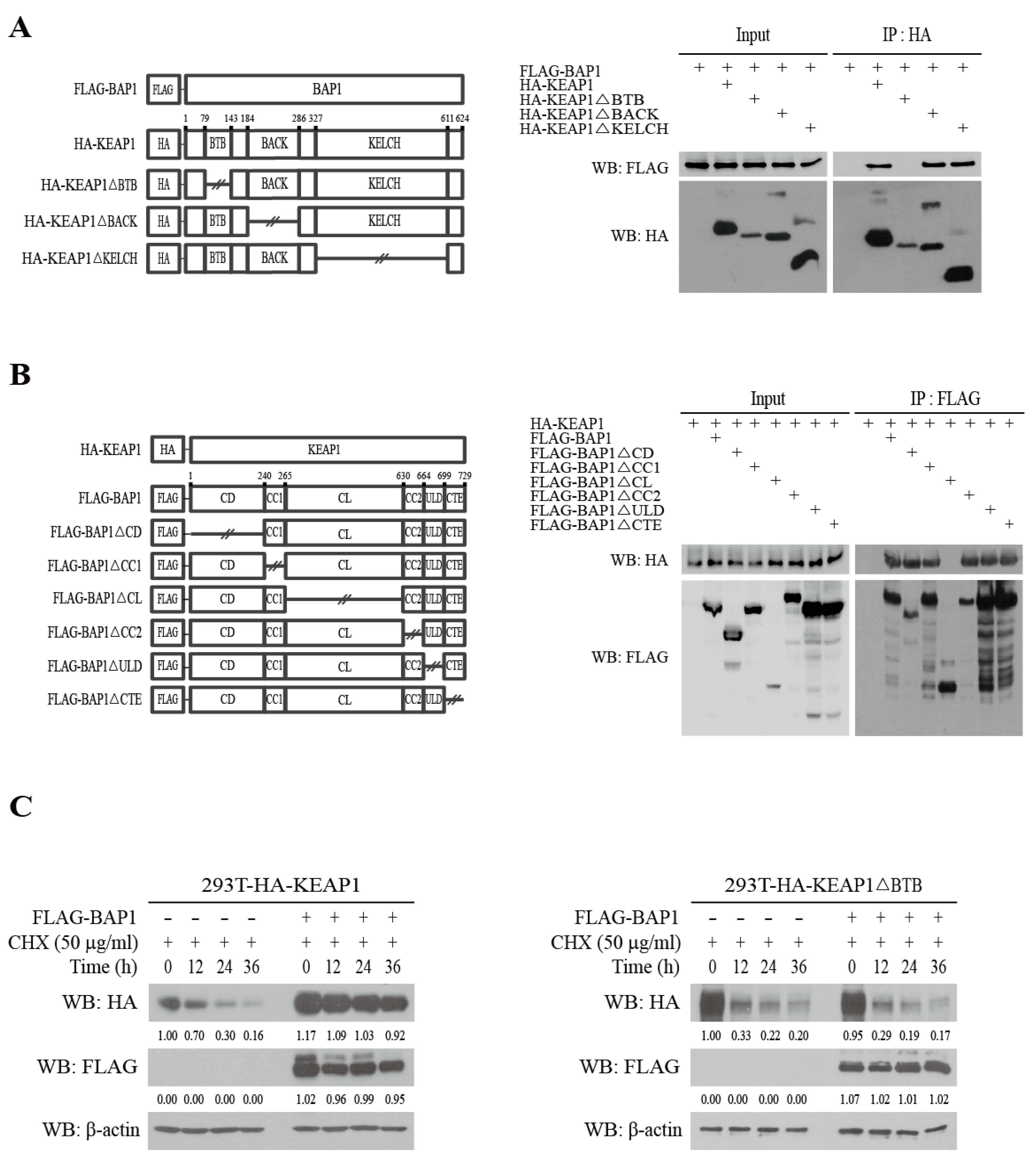
3.2. BAP1 Stabilizes KEAP1 by Binding to the BTB Domain
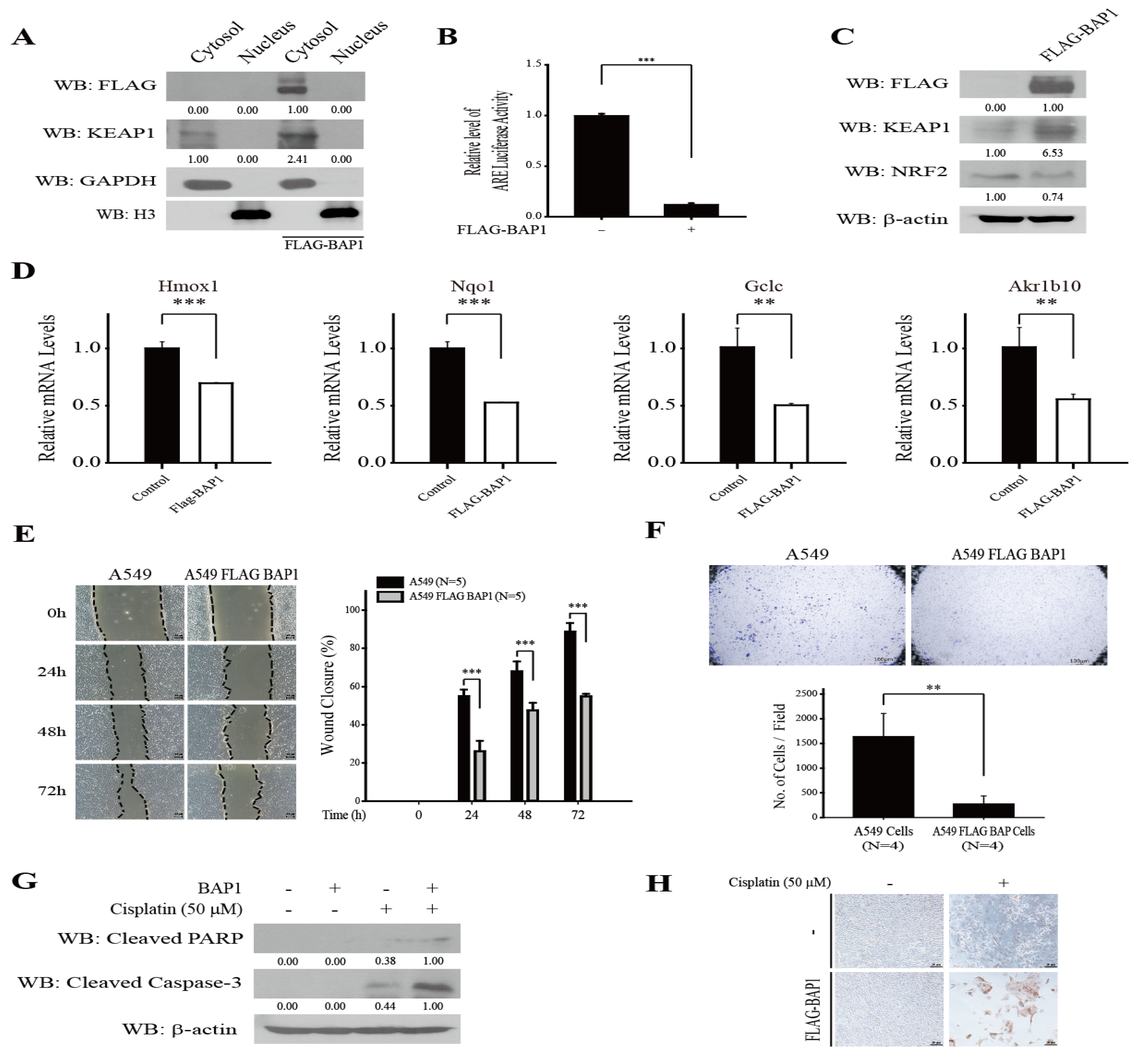
3.3. Overexpression of BAP1 Inhibits NRF2, Suppresses Migration and Invasion, and Promotes Cisplatin-Induced Apoptosis of A549 Cells
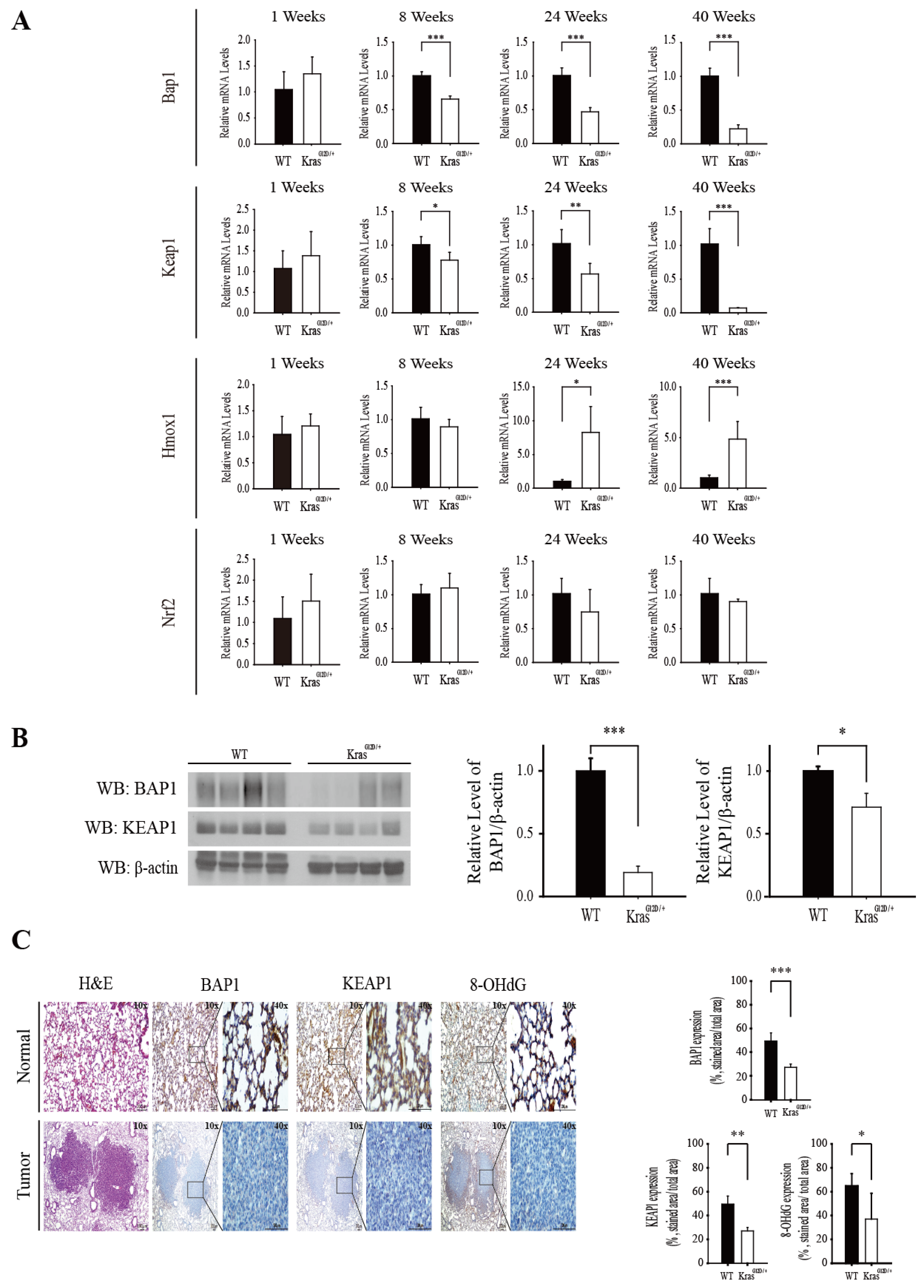
3.4. The Level of Bap1 and Keap1 mRNA Decreases during Progression of Lung Adenocarcinoma in KrasG12D/+ Mice and It Is Positively Associated with NRF2 Activation and Suppression of Oxidative Stress
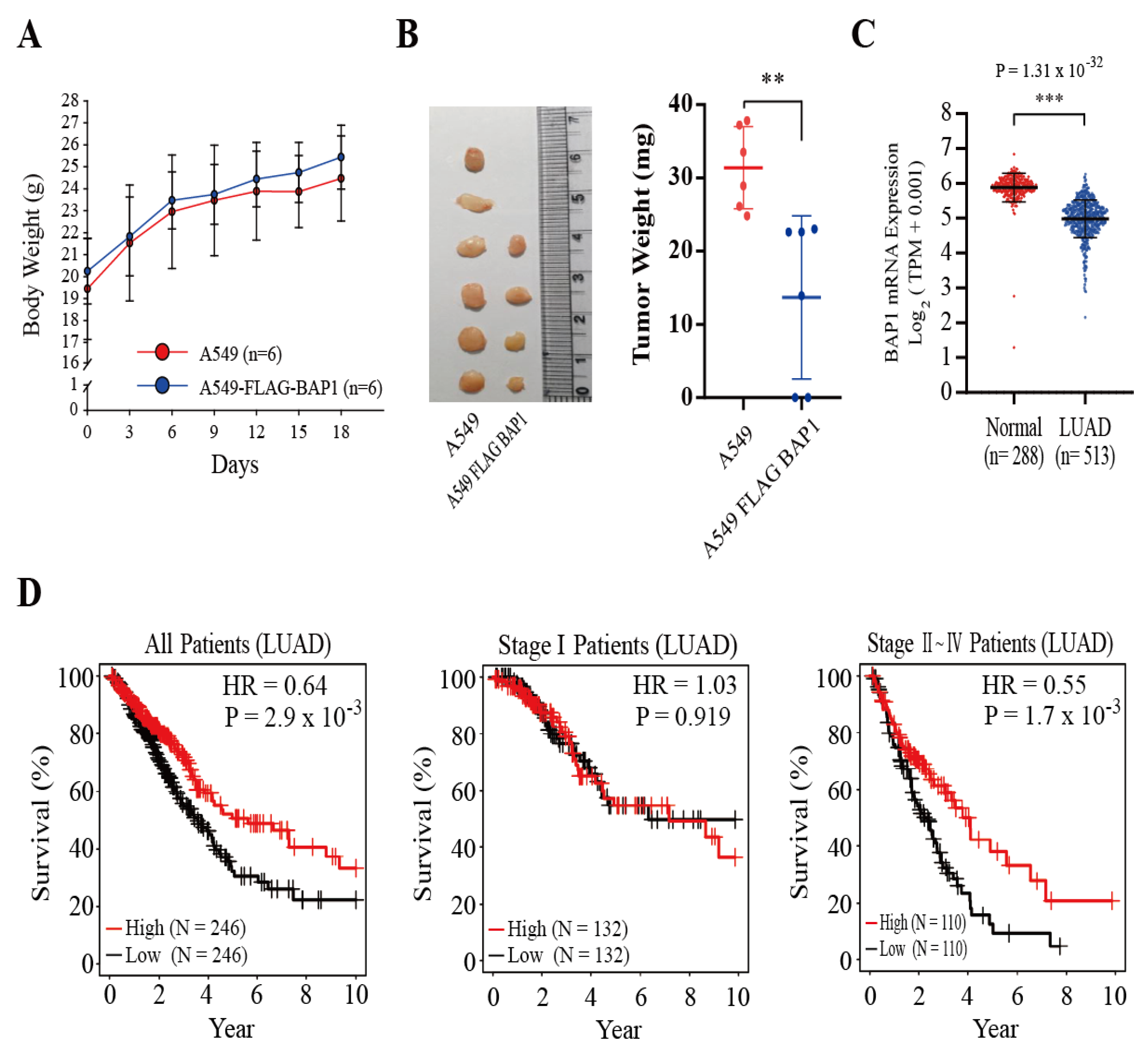
3.5. BAP1 Suppresses the Growth of A549 Xenografts, and the Expression of Bap1 Is Higher in Normal Lung Tissues and Correlated with a Better Survival of LUAD Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Suzuki, T.; Motohashi, H.; Yamamoto, M. Toward clinical application of the Keap1-Nrf2 pathway. Trends Pharmacol. Sci. 2013, 34, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; Leon, R.; Lopez, M.G.; Oliva, B.; et al. Transcription Factor NRF2 as a Therapeutic Target for Chronic Diseases: A Systems Medicine Approach. Pharmacol. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.S.; Nam, L.B.; Yoo, O.K.; Keum, Y.S. Molecular mechanisms and systemic targeting of NRF2 dysregulation in cancer. Biochem. Pharmacol. 2020, 177, 114002. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.J.; Yoo, H.S.; Shin, S.; Park, Y.J.; Jeon, S.M. Dysregulation of NRF2 in Cancer: From Molecular Mechanisms to Therapeutic Opportunities. Biomol. Ther. 2018, 26, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Yamamoto, M. The KEAP1-NRF2 System in Cancer. Front. Oncol. 2017, 7, 85. [Google Scholar] [CrossRef]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Mitsuishi, Y.; Motohashi, H.; Yamamoto, M. The Keap1-Nrf2 system in cancers: Stress response and anabolic metabolism. Front. Oncol. 2012, 2, 200. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.D.; Lo, S.C.; Sun, Z.; Habib, G.M.; Lieberman, M.W.; Hannink, M. Ubiquitination of Keap1, a BTB-Kelch substrate adaptor protein for Cul3, targets Keap1 for degradation by a proteasome-independent pathway. J. Biol. Chem. 2005, 280, 30091–30099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tao, S.; Liao, L.; Li, Y.; Li, H.; Li, Z.; Lin, L.; Wan, X.; Yang, X.; Chen, L. TRIM25 promotes the cell survival and growth of hepatocellular carcinoma through targeting Keap1-Nrf2 pathway. Nat. Commun. 2020, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Clague, M.J.; Urbe, S.; Komander, D. Breaking the chains: Deubiquitylating enzyme specificity begets function. Nat. Rev. Mol. Cell Biol. 2019, 20, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Mevissen, T.E.T.; Komander, D. Mechanisms of Deubiquitinase Specificity and Regulation. Annu. Rev. Biochem. 2017, 86, 159–192. [Google Scholar] [CrossRef] [Green Version]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating enzymes and drug discovery: Emerging opportunities. Nat. Rev. Drug Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Mercer, K.; Greenbaum, D.; Bronson, R.T.; Crowley, D.; Tuveson, D.A.; Jacks, T. Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature 2001, 410, 1111–1116. [Google Scholar] [CrossRef]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repecka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Nam, L.B.; Keum, Y.S. Binding partners of NRF2: Functions and regulatory mechanisms. Arch. Biochem. Biophys. 2019, 678, 108184. [Google Scholar] [CrossRef] [PubMed]
- De, I.; Chittock, E.C.; Grotsch, H.; Miller, T.C.R.; McCarthy, A.A.; Muller, C.W. Structural Basis for the Activation of the Deubiquitinase Calypso by the Polycomb Protein ASX. Structure 2019, 27, 528–536.e524. [Google Scholar] [CrossRef] [Green Version]
- Cloer, E.W.; Goldfarb, D.; Schrank, T.P.; Weissman, B.E.; Major, M.B. NRF2 Activation in Cancer: From DNA to Protein. Cancer Res. 2019, 79, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, A.D.; Fesik, S.W.; Kimmelman, A.C.; Luo, J.; Der, C.J. Drugging the undruggable RAS: Mission possible? Nat. Rev. Drug Discov. 2014, 13, 828–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeNicola, G.M.; Karreth, F.A.; Humpton, T.J.; Gopinathan, A.; Wei, C.; Frese, K.; Mangal, D.; Yu, K.H.; Yeo, C.J.; Calhoun, E.S.; et al. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 2011, 475, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Wang, S.; Moghaddam, S.J.; Ooi, A.; Chapman, E.; Wong, P.K.; Zhang, D.D. Oncogenic KRAS confers chemoresistance by upregulating NRF2. Cancer Res. 2014, 74, 7430–7441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keum, Y.S.; Choi, B.Y. Molecular and chemical regulation of the Keap1-Nrf2 signaling pathway. Molecules 2014, 19, 10074–10089. [Google Scholar] [CrossRef] [Green Version]
- Kris, M.G.; Gaspar, L.E.; Chaft, J.E.; Kennedy, E.B.; Azzoli, C.G.; Ellis, P.M.; Lin, S.H.; Pass, H.I.; Seth, R.; Shepherd, F.A.; et al. Adjuvant Systemic Therapy and Adjuvant Radiation Therapy for Stage I to IIIA Completely Resected Non-Small-Cell Lung Cancers: American Society of Clinical Oncology/Cancer Care Ontario Clinical Practice Guideline Update. J. Clin. Oncol. 2017, 35, 2960–2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murali, R.; Wiesner, T.; Scolyer, R.A. Tumours associated with BAP1 mutations. Pathology 2013, 45, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Yang, H.; Pass, H.I.; Krausz, T.; Testa, J.R.; Gaudino, G. BAP1 and cancer. Nat. Rev. Cancer 2013, 13, 153–159. [Google Scholar] [CrossRef]
- Dey, A.; Seshasayee, D.; Noubade, R.; French, D.M.; Liu, J.; Chaurushiya, M.S.; Kirkpatrick, D.S.; Pham, V.C.; Lill, J.R.; Bakalarski, C.E.; et al. Loss of the tumor suppressor BAP1 causes myeloid transformation. Science 2012, 337, 1541–1546. [Google Scholar] [CrossRef] [Green Version]
- Kadariya, Y.; Cheung, M.; Xu, J.; Pei, J.; Sementino, E.; Menges, C.W.; Cai, K.Q.; Rauscher, F.J.; Klein-Szanto, A.J.; Testa, J.R. Bap1 Is a Bona Fide Tumor Suppressor: Genetic Evidence from Mouse Models Carrying Heterozygous Germline Bap1 Mutations. Cancer Res. 2016, 76, 2836–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, D.E.; Proctor, M.; Marquis, S.T.; Gardner, H.P.; Ha, S.I.; Chodosh, L.A.; Ishov, A.M.; Tommerup, N.; Vissing, H.; Sekido, Y.; et al. BAP1: A novel ubiquitin hydrolase which binds to the BRCA1 RING finger and enhances BRCA1-mediated cell growth suppression. Oncogene 1998, 16, 1097–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, M.; Harbour, J.W.; Brugarolas, J.; Bononi, A.; Pagano, I.; Dey, A.; Krausz, T.; Pass, H.I.; Yang, H.; Gaudino, G. Biological Mechanisms and Clinical Significance of BAP1 Mutations in Human Cancer. Cancer Discov. 2020, 10, 1103–1120. [Google Scholar] [CrossRef] [PubMed]
- Mashtalir, N.; Daou, S.; Barbour, H.; Sen, N.N.; Gagnon, J.; Hammond-Martel, I.; Dar, H.H.; Therrien, M.; Affar, E.B. Autodeubiquitination protects the tumor suppressor BAP1 from cytoplasmic sequestration mediated by the atypical ubiquitin ligase UBE2O. Mol. Cell 2014, 54, 392–406. [Google Scholar] [CrossRef] [Green Version]
- Bononi, A.; Giorgi, C.; Patergnani, S.; Larson, D.; Verbruggen, K.; Tanji, M.; Pellegrini, L.; Signorato, V.; Olivetto, F.; Pastorino, S.; et al. BAP1 regulates IP3R3-mediated Ca(2+) flux to mitochondria suppressing cell transformation. Nature 2017, 546, 549–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villeneuve, N.F.; Tian, W.; Wu, T.; Sun, Z.; Lau, A.; Chapman, E.; Fang, D.; Zhang, D.D. USP15 negatively regulates Nrf2 through deubiquitination of Keap1. Mol. Cell 2013, 51, 68–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Genome Atlas Research, N. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar] [CrossRef]
- Simanshu, D.K.; Nissley, D.V.; McCormick, F. RAS Proteins and Their Regulators in Human Disease. Cell 2017, 170, 17–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobrinski, D.A.; Yang, H.; Kittaneh, M. BAP1: Role in carcinogenesis and clinical implications. Transl. Lung Cancer Res. 2020, 9, S60–S66. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Gene | Primer Sequence | |
|---|---|---|---|
| Mouse | NM_027088 | BAP1 | Forward: 5′-CAGTGAGCCCTTGAGTGGAG-3′ Reverse: 5′-GGTCCTTCGCTGGTCATCAA-3′ |
| NM_016679 | KEAP1 | Forward: 5′-CTCAACCGCTTGCTGTATGC-3′ Reverse: 5′-TTCAACTGGTCCTGCCCATC-3′ | |
| NM_010442 | HMOX1 | Forward: 5′-AGCCCCACCAAGTTCAAACA-3′ Reverse: 5′-TCTCTGCAGGGGCAGTATCT-3′ | |
| NM_010902 | NRF2 | Forward: 5′-CGCTGGAAAAAGAAGTGGGC-3′ Reverse: 5′-GTGACAGGTCACAGCCTTCA-3′ | |
| NM_01289726 | GAPDH | Forward: 5′-GGAGAGTGTTTCCTCGTCCC-3′ Reverse: 5′-ACTGTGCCGTTGAATTTGCC-3′ | |
| Human | NM_002133 | HMOX1 | Forward: 5′-GTGCCACCAAGTTCAAGCAG-3′ Reverse: 5′-CAGCTCCTGCAACTCCTCAA-3′ |
| NM_000903 | NQO1 | Forward: 5′-GGTTTGGAGTCCCTGCCATT-3′ Reverse: 5′-GCCTTCTTACTCCGGAAGGG-3′ | |
| NM_001197115 | GCLC | Forward: 5′-GGAGGAACAATGTCCGAGTT-3′ Reverse: 5′-AGCGAGGGTGCTTGTTTATT-3′ | |
| NM_020299 | AKR1B10 | Forward: 5′-GACTGTGCCTATGTCTATCA-3′ Reverse: 5′-AAGATAGACGTCCAGATAGC-3′ | |
| NM_002046 | GAPDH | Forward: 5′-CATCAATGGAAATCCCATCA-3′ Reverse: 5′-GGCAGAGATGATGACCCTTT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.-S.; Nam, L.B.; Yoo, O.-K.; Lee, K.; Suh, Y.-A.; Kim, D.; Kim, W.K.; Lim, C.-Y.; Lee, H.; Keum, Y.-S. BAP1 Downregulates NRF2 Target Genes and Exerts Anti-Tumorigenic Effects by Deubiquitinating KEAP1 in Lung Adenocarcinoma. Antioxidants 2022, 11, 114. https://doi.org/10.3390/antiox11010114
Kang J-S, Nam LB, Yoo O-K, Lee K, Suh Y-A, Kim D, Kim WK, Lim C-Y, Lee H, Keum Y-S. BAP1 Downregulates NRF2 Target Genes and Exerts Anti-Tumorigenic Effects by Deubiquitinating KEAP1 in Lung Adenocarcinoma. Antioxidants. 2022; 11(1):114. https://doi.org/10.3390/antiox11010114
Chicago/Turabian StyleKang, Jong-Su, Le Ba Nam, Ok-Kyung Yoo, Kyeong Lee, Young-Ah Suh, Dalyong Kim, Woo Kyung Kim, Chi-Yeon Lim, Haeseung Lee, and Young-Sam Keum. 2022. "BAP1 Downregulates NRF2 Target Genes and Exerts Anti-Tumorigenic Effects by Deubiquitinating KEAP1 in Lung Adenocarcinoma" Antioxidants 11, no. 1: 114. https://doi.org/10.3390/antiox11010114
APA StyleKang, J.-S., Nam, L. B., Yoo, O.-K., Lee, K., Suh, Y.-A., Kim, D., Kim, W. K., Lim, C.-Y., Lee, H., & Keum, Y.-S. (2022). BAP1 Downregulates NRF2 Target Genes and Exerts Anti-Tumorigenic Effects by Deubiquitinating KEAP1 in Lung Adenocarcinoma. Antioxidants, 11(1), 114. https://doi.org/10.3390/antiox11010114