
Dietary Curcumin Improves Energy Metabolism, Brain Monoamines, Carcass Traits, Muscle Oxidative Stability and Fatty Acid Profile in Heat-Stressed Broiler Chickens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Birds and Management
2.2. Carcass Traits
2.3. Determination of Muscle Fatty Acid Profiles
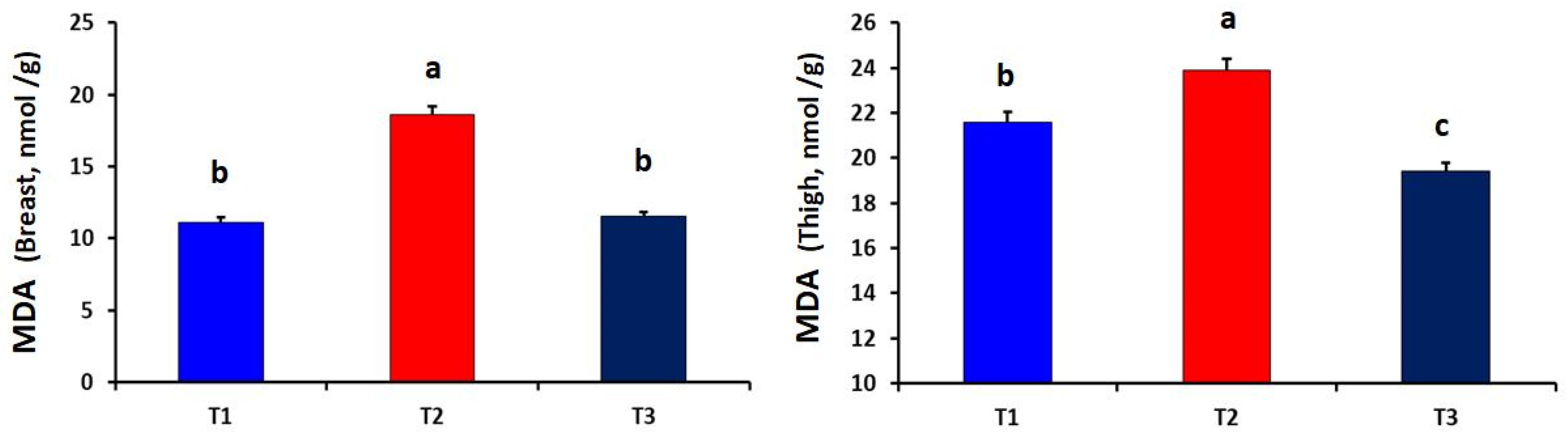
2.4. Determination of Muscle Malondialdehyde
2.5. Determination of Brain Monoamines
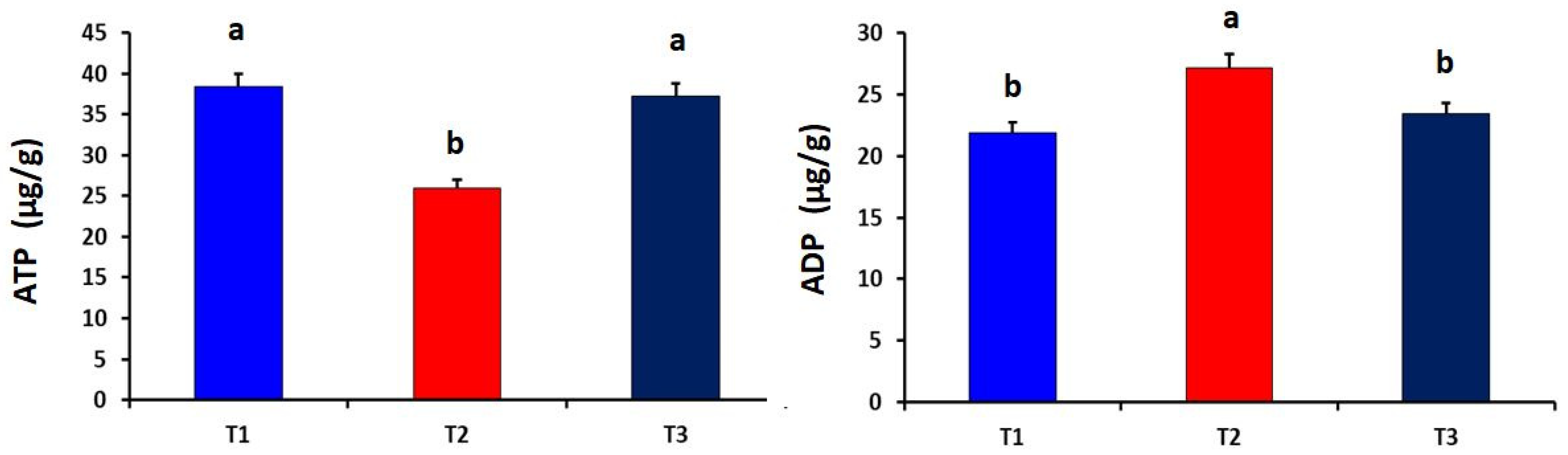
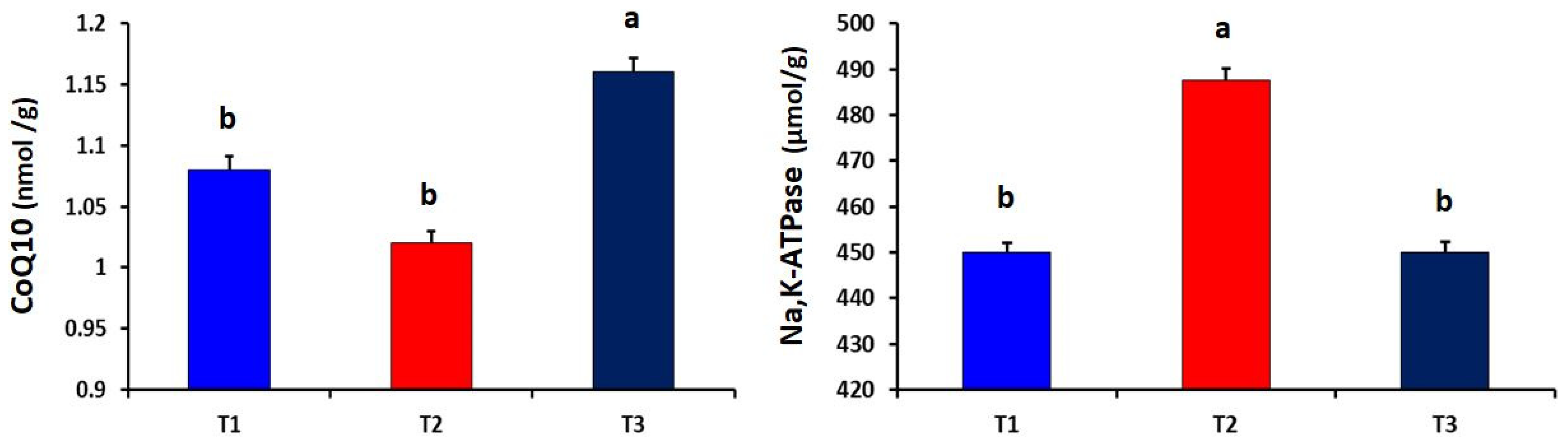
2.6. Determination of CoQ10 and Energy Biomarkers in Liver Tissues
2.7. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Tarabany, M.S. Effect of thermal stress on fertility and egg quality of Japanese quail. J. Therm. Biol. 2016, 61, 38–43. [Google Scholar] [CrossRef]
- Hansen, J.; Ruedy, R.; Sato, M.; Lo, K. Global surface temperature change. Rev. Geophys. 2010, 48, RG4004. [Google Scholar] [CrossRef] [Green Version]
- Bogin, E.; Avidar, Y.; Pech-Waffenschmidt, V.; Doron, Y.; Israeli, B.A.; Kevkhayev, E. The relationship between heat stress, survivability and blood composition of the domestic hen. Eur. J. Clin. Chem. Clin. Biochem. 1996, 34, 463–469. [Google Scholar]
- Swennen, Q.; Verhulst, P.J.; Collin, A.; Bordas, A.; Verbeke, K.; Vansant, G.; Decuypere, E.; Buyse, J. Further investigations on the role of dietinduced thermogenesis in the regulation of feed intake in chickens: Comparison of adult cockerels E62 Bottje and Carstens of lines selected for high or low residual feed intake. Poult. Sci. 2007, 86, 1960–1971. [Google Scholar] [CrossRef]
- Sahin, K.; Kucuk, O. Heat Stress and Dietary Vitamin Supplementation of Poultry Diets. In Nutrition Abstracts and Reviews. Series B, Livestock Feeds and Feeding; CAB International: Wallingford, UK, 2003; Volume 73, pp. 41–50. [Google Scholar]
- Roushdy, E.M.; Zaglool, A.W.; El-Tarabany, M.S. Effects of chronic thermal stress on growth performance, carcass traits, antioxidant indices and the expression of HSP70, growth hormone and superoxide dismutase genes in two broiler strains. J. Therm. Biol. 2018, 74, 337–343. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Sakai, M.; Sá, L.R.M.; Ferreira, A.J.P.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef]
- Lu, Q.; Wen, J.; Zhang, H. Effect of chronic heat exposure on fat deposition and meat quality in two genetic types of chicken. Poult. Sci. 2007, 86, 1059–1064. [Google Scholar] [CrossRef]
- Gous, R.M. Nutritional limitations on growth and development in poultry. Livest. Sci. 2010, 130, 25–32. [Google Scholar] [CrossRef]
- Iwagami, Y. Changes in the ultrasonic of human cells related to certain biological responses under hyperthermic culture conditions. Hum. Cell 1996, 9, 353–366. [Google Scholar]
- Akbarian, A.; Golian, A.; Kermanshahi, H.; De Smet, S.; Michiels, J. Antioxidant enzyme activities, plasma hormone levels and serum metabolites of finishing broiler chickens reared under high ambient temperature and fed lemon and orange peel extracts and C urcuma xanthorrhiza essential oil. J. Anim. Physiol. Anim. Nutr. 2015, 99, 150–162. [Google Scholar] [CrossRef]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Mujahid, A.; Pumford, N.R.; Bottje, W.; Nakagawa, K.; Miyazawa, T.; Akiba, Y.; Toyomizu, M. Mitochondrial oxidative damage in chicken skeletal muscle induced by acute heat stress. J. Poult. Sci. 2007, 44, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.; Warner, R.D.; Dunshea, F.R. Growth performance and characterization of meat quality of broiler chickens supplemented with betaine and antioxidants under cyclic heat stress. Antioxidants 2019, 8, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Chronic heat stress Impairs the quality of breast-muscle meat in broilers by affecting redox status and energy-substance metabolism. J. Agric. Food Chem. 2017, 65, 11251–11258. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M.R.; El-Hack, M.A.; Dhama, K.; Fowler, J. Use of acetylsalicylic acid as a feed additive in poultry nutrition. Worlds Poult. Sci. J. 2017, 73, 633–642. [Google Scholar] [CrossRef]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “curecumin”. From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.U.; Naz, S.; Javdani, M.; Nikousefat, Z.; Selvaggi, M.; Tufarelli, V.; Laudadio, V. The use of turmeric (Curcuma longa) in poultry feed. Worlds Poult. Sci. J. 2012, 68, 97–103. [Google Scholar] [CrossRef]
- Griesser, M.; Pistis, V.; Suzuki, T.; Tejera, N.; Pratt, D.A.; Schneider, C. Autoxidative and cyclooxygenase-2 catalyzed transformation of the dietary chemopreventive agent curcumin. J. Biol. Chem. 2011, 286, 1114–1124. [Google Scholar] [CrossRef] [Green Version]
- Khalil, O.A.K.; De Faria Oliveira, O.M.M.; Vellosa, J.C.R.; De Quadros, A.U.; Dalposso, L.M.; Karam, T.K.; Mainardes, R.M.; Khalil, N.M. Curcumin antifungal and antioxidant activities are increased in the presence of ascorbic acid. Food Chem. 2012, 133, 1001–1005. [Google Scholar] [CrossRef]
- Negi, P.S.; Jayaprakasha, G.K.; Jaganmohan, R.L.; Sakariah, K.K. Antibacterial activity of turmeric oil: A byproduct from curcumin manufacture. J. Agric. Food Chem. 1999, 47, 4297–4300. [Google Scholar] [CrossRef]
- Rajput, N.; Muhammah, N.; Yan, R.; Zhong, X.; Wang, T. Effect of dietary supplementation of curcumin on growth performance, intestinal morphology and nutrients utilization of broiler chicks. Poult. Sci. J. 2013, 50, 44–52. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- JAKIM (Department of Islamic Development Malaysia). Malaysian Protocol for the Halal Meat and Poultry Productions; Standards Malaysia: Selangor, Malaysia, 2011; pp. 1–30.
- Radwan, O.K.; Ahmed, R.F. Amendment effect of resveratrol on diclofenac idiosyncratic toxicity: Augmentation of the anti-inflammatory effect by assessment of Arachidonic acid and IL-1β levels. J. Appl. Pharm. Sci. 2016, 6, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Karalas, F.; Karatepe, M.; Baysar, A. Determination of free malondialdehyde in human serum by high performance liquid chromatography. Anal. Biochem. 2002, 311, 76–79. [Google Scholar] [CrossRef]
- Arafa, N.M.S.; Salem, S.M.A.; Farid, O.A.H.A. Influence of echinacea extract pre- or postnatal supplementation on immune and oxidative status of growing rabbits. Ital. J. Anim. Sci. 2010, 9, 338–343. [Google Scholar]
- Pagel, P.; Blome, J.; Wolf, H.U. High-performance liquid chromatographic separation and measurement of various biogenic compounds possibly involved in the pathomechanism of Parkinson’s disease. J. Chromatogr. B 2000, 746, 297–304. [Google Scholar] [CrossRef]
- Teerlink, T.; Hennekes, M.; Bussemaker, J.; Groeneveld, J. Simultaneous determination of creatine compounds and adenine nucleotides in myocardial tissue by high-performance liquid chromatography. Anal. Biochem. 1993, 214, 278–283. [Google Scholar] [CrossRef]
- Niklowitz, P.; Doring, F.; Paulussen, M.; Menke, T. Determination of coenzyme Q10 tissue status viahigh-performance liquid chromatography with electrochemical detection in swine tissues (Sus scrofadomestica). Anal. Biochem. 2013, 437, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Geraert, P.A.; Guillaumin, S.; Leclercq, B. Are genetically lean broilers more resistant to hot climate? Br. Poult. Sci. 1993, 34, 643–653. [Google Scholar] [CrossRef]
- Ruan, D.; Wang, W.C.; Lin, C.X.; Fouad, A.M.; Chen, W.; Xia, W.G.; Wang, S.; Luo, X.; Zhang, W.H.; Yan, S.J.; et al. Effects of curcumin on performance, antioxidation, intestinal barrier and mitochondrial function in ducks fed corn contaminated with ochratoxin A. Animal 2019, 13, 42–52. [Google Scholar] [CrossRef]
- Delesalle, C.; van de Walle, G.R.; Nolten, C.; Ver Donck, L.; Van Hemelrijck, A.; Drinkenburg, W.; De Bosschere, H.; Claes, P.; Deprez, P.; Lefere, L.; et al. Determination of the source of increased serotonin (5-HT) concentrations in blood and peritoneal fluid of colic horses with compromised bowel. Equine Vet. J. 2008, 40, 326–331. [Google Scholar] [CrossRef]
- Gowda, N.K.; Ledoux, D.R.; Rottinghaus, G.E.; Bermudez, A.J.; Chen, Y.C. Antioxidant efficacy of curcuminoids from Turmeric (Curcuma longa L.) powder in broiler chickens fed diets containing aflatoxin B1. Br. J. Nutr. 2009, 102, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Rajput, N.; Ali, S.; Naeem, M.; Khan, M.A.; Wang, T. The effect of dietary supplementation with the natural carotenoids curcumin and lutein on pigmentation, oxidative stability and quality of meat from broiler chickens affected by a coccidiosis challenge. Br. Poult. Sci. 2014, 55, 501–509. [Google Scholar] [CrossRef]
- Hang, T.T.T.; Molee, W.; Khempaka, S.; Paraksa, N. Supplementation with curcuminoids and tuna oil influenced skin yellowness, carcass composition, oxidation status, and meat fatty acids of slow-growing chickens. Poult. Sci. 2018, 97, 901–909. [Google Scholar] [CrossRef]
- Abou-Elkhair, R.H.A.; Ahmed Selim, S. Effects of black pepper (Piper nigrum), turmeric powder (Curcuma longa) and coriander seeds (Coriandrum sativum) and their combinations as feed additives on growth performance, carcass traits, some blood parameters and humoral immune response of broiler chickens. Asian-Australas. J. Anim. Sci. 2014, 27, 847–854. [Google Scholar]
- Bai, S.; Wang, G.; Zhang, W.; Zhang, S.; Rice, B.B.; Cline, M.A.; Gilbert, E.R. Broiler chicken adipose tissue dynamics during the first two weeks post-hatch. Comp. Biochem. Physiol. A 2015, 189, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Nouzarian, R.; Tabeidian, S.; Toghyani, M.; Ghalamkari, G. Effect of turmeric powder on performance, carcass traits, humoral immune responses, and serum metabolites in broiler chickens. J. Anim. Feed Sci. 2011, 487, 47. [Google Scholar] [CrossRef]
- Zaboli, G.; Huang, X.; Feng, X.; Ahn, D.U. How can heat stress affect chicken meat quality?—A review. Poult. Sci. 2019, 98, 1551–1556. [Google Scholar] [CrossRef]
- Pita, M.C.G.; Piber Neto, E.; Nakaoka, L.M.; Mendonça, J.C.X. Efeito da adição de ácidos graxos insaturados e de vitamina E à dieta de galinhas e seu reflex na composição lipídica e incorporação de α-tocoferol na gema do ovo. Braz. J. Vet. Res. Anim. Sci. 2004, 41, 25–31. [Google Scholar] [CrossRef]
- Botsoglou, N.; Florou-Paneri, P.; Botsoglou, E.; Dotas, V.; Giannenas, I.; Koidis, A.; Mitrakos, P. The effect of feeding rosemary, oregano, saffron and a-tocopheryl acetate on hen performance and oxidative stability of eggs. S. Afr. J. Anim. Sci. 2005, 35, 143–151. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, K.W.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin attenuates hepatic mitochondrial dysfunction through the maintenance of thiol pool, inhibition of mtDNA damage, and stimulation of the mitochondrial thioredoxin system in heat-stressed broilers. J. Anim. Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Daneshyar, M.; Ghandkanlo, M.A.; Bayeghra, F.S.; Farhangpajhoh, F.; Aghaei, M. Effects of dietary turmeric supplementation on plasma lipoproteins, meat quality and fatty acid composition in broilers. S. Afr. J. Anim. Sci. 2011, 41, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Gnoni, G.; Paglialonga, G.; Siculella, L. Quercetin inhibits fatty acid and triacylglycerol synthesis in rat liver cells. Eur. J. Clin. Investig. 2009, 39, 761–768. [Google Scholar] [CrossRef]
- Martínez-Morúa, A.; Soto-Urquieta, M.G.; Franco-Robles, E.; Zúñiga-Trujillo, I.; Campos-Cervantes, A.; Pérez-Vázquez, V.; Ramírez-Emiliano, J. Curcumin decreases oxidative stress in mitochondria isolated from liver and kidneys of high-fat diet-induced obese mice. J. Asian Nat. Prod. Res. 2013, 15, 905–915. [Google Scholar] [CrossRef]
- Soto, P.; Smith, L.C. BH4 peptide derived from Bcl-xL and Bax-inhibitor peptide suppresses apoptotic mitochondrial changes in heat stressed bovine oocytes. Mol. Reprod. Dev. 2009, 76, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xie, J.; Wang, B.; Tang, J. Effect of gammaaminobutyric acid on digestive enzymes, absorption function, and immune function of intestinal mucosa in heatstressed chicken. Poult. Sci. 2014, 93, 2490–2500. [Google Scholar] [CrossRef] [PubMed]
- Gopi, M.; Purushothaman, M.R.; Chandrasekaran, D. Effect of dietary coenzyme Q10 supplementation on the growth rate, carcass characters and cost effectiveness of broiler fed with three energy levels. SpringerPlus 2014, 3, 518. [Google Scholar] [CrossRef] [Green Version]
- Geng, A.L.; Guo, Y.M.; Yang, Y. Reduction of ascites mortality in broilers by coenzyme Q10. Poult. Sci. 2004, 83, 1587–1593. [Google Scholar] [CrossRef]
- Xu, J.; Tang, S.; Yin, B.; Sun, J.R.; Song, E.B.; Bao, E.D. Co-enzyme Q10 and Acetyl salicylic acid enhance Hsp70 expression in primary chicken myocardial cells to protect the cells during heat stress. Mol. Cell. Biochem. 2017, 435, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yin, B.; Huang, B.; Tang, S.; Zhang, X.; Sun, J.; Bao, E. Co-enzyme Q10 protects chicken hearts from in vivo heat stress via inducing HSF1 binding activity and Hsp70 expression. Poult. Sci. 2019, 98, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Raybould, H.E. Visceral perception: Sensory transduction in visceral afferents and nutrients. Gut 2002, 51 (Suppl. 1), i11–i14. [Google Scholar] [CrossRef]
- Buraczewska, L.; Nguyen, C.V.; Mieczkowska, A. Effects of acidifier added to diets containing graded levels of crystalline tryptophan on growth performance, protein digestibility, and on brain serotonin level in broiler chickens. J. Anim. Feed Sci. 2004, 13, 289–300. [Google Scholar]
- Denbow, D.M.; Hobbs, F.C.; Hulet, R.M.; Graham, P.P.; Potter, L.M. Supplemental dietary L-tryptophan effects on growth, meat quality, and brain catecholamine and indolamine concentrations in turkeys. Br. Poult. Sci. 1993, 34, 715–724. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calculated Analysis | Starter Period (1–21 Days) | Grower-Finisher Period (22–42 Days) |
|---|---|---|
| ME (KJ/kg) | 12,342 | 12,949 |
| Crude protein (%) | 22.40 | 19.75 |
| Calcium (%) | 1.05 | 1.05 |
| Available phosphorus (%) | 0.45 | 0.45 |
| Lysine (%) | 1.18 | 1.14 |
| Methionine (%) | 0.48 | 0.45 |
| Parameters (%) | Experimental Groups | ||||
|---|---|---|---|---|---|
| 1 T1 | 2 T2 | 3 T3 | SEM 4 | p-Value | |
| 5 ADFI (g) | 117.5 a | 102.2 b | 116.7 a | 2.87 | 0.048 |
| Dressing percentage | 75.4 a | 68.5 b | 75.2 a | 1.13 | 0.010 |
| Breast | 42.1 a | 35.2 b | 40.9 a | 0.98 | 0.004 |
| Legs | 32.8 | 37.7 | 34.4 | 0.93 | 0.070 |
| Liver | 2.26 | 2.12 | 2.32 | 0.07 | 0.099 |
| Heart | 0.38 | 0.36 | 0.43 | 0.02 | 0.296 |
| Abdominal fat | 0.63 b | 0.95 a | 0.56 b | 0.05 | 0.017 |
| Fatty Acids | Experimental Groups | ||||
|---|---|---|---|---|---|
| 1 T1 | 2 T2 | 3 T3 | SEM 4 | p-Value | |
| Myristic (C14:0) | 0.76 b | 0.97 a | 0.80 b | 0.04 | 0.001 |
| Palmitic (C16:0) | 30.8 b | 37.6 a | 33.2 b | 1.64 | 0.007 |
| Stearic (C18:0) | 12.28 | 12.98 | 12.31 | 0.59 | 0.159 |
| Myristoleic acid (C14:1) | 1.13 a | 0.83 b | 1.18 a | 0.04 | 0.009 |
| Palmitoleic (C16:1) | 1.22 a | 0.92 b | 1.15 a | 0.08 | 0.001 |
| Oleic (C18:1) | 20.7 b | 15.5 c | 23.4 a | 1.06 | 0.001 |
| Linoleic (C18:2n6) | 15.85 a | 11.31 b | 16.22 a | 0.74 | 0.016 |
| α-linolenic acid (C18:3n3) | 0.91 | 0.85 | 0.95 | 0.02 | 0.093 |
| Docosahexaenoic acid (C22:6n3) | 0.63 a | 0.40 b | 0.61 a | 0.02 | 0.001 |
| Eicosapentaenoic acid (C20:5n3) | 0.71 a | 0.49 b | 0.72 a | 0.03 | 0.035 |
| Fatty Acids | Experimental Groups | ||||
|---|---|---|---|---|---|
| 1 T1 | 2 T2 | 3 T3 | SEM 4 | p-Value | |
| Myristic (C14:0) | 1.23 | 1.34 | 1.25 | 0.04 | 0.143 |
| Palmitic (C16:0) | 27.9 c | 31.7 a | 30.2 b | 1.77 | 0.001 |
| Stearic (C18:0) | 10.59 b | 11.85 a | 10.78 b | 0.38 | 0.048 |
| Myristoleic acid (C14:1) | 1.09 a | 0.83 b | 1.16 a | 0.04 | 0.001 |
| Palmitoleic (C16:1) | 1.98 a | 1.57 b | 2.08 a | 0.12 | 0.009 |
| Oleic (C18:1) | 28.9 a | 21.8 b | 28.4 a | 1.04 | 0.013 |
| Linoleic (C18:2n6) | 20.24 a | 13.09 c | 18.52 b | 0.45 | 0.006 |
| α-linolenic acid (C18:3n3) | 0.59 | 0.65 | 0.60 | 0.02 | 0.086 |
| Docosahexaenoic acid (C22:6n3) | 0.51 a | 0.36 c | 0.48 b | 0.02 | 0.001 |
| Eicosapentaenoic acid (C20:5n3) | 0.88 a | 0.57 c | 0.81 b | 0.04 | 0.026 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salah, A.S.; Ahmed-Farid, O.A.; Nassan, M.A.; El-Tarabany, M.S. Dietary Curcumin Improves Energy Metabolism, Brain Monoamines, Carcass Traits, Muscle Oxidative Stability and Fatty Acid Profile in Heat-Stressed Broiler Chickens. Antioxidants 2021, 10, 1265. https://doi.org/10.3390/antiox10081265
Salah AS, Ahmed-Farid OA, Nassan MA, El-Tarabany MS. Dietary Curcumin Improves Energy Metabolism, Brain Monoamines, Carcass Traits, Muscle Oxidative Stability and Fatty Acid Profile in Heat-Stressed Broiler Chickens. Antioxidants. 2021; 10(8):1265. https://doi.org/10.3390/antiox10081265
Chicago/Turabian StyleSalah, Ayman S., Omar A. Ahmed-Farid, Mohamed Abdo Nassan, and Mahmoud S. El-Tarabany. 2021. "Dietary Curcumin Improves Energy Metabolism, Brain Monoamines, Carcass Traits, Muscle Oxidative Stability and Fatty Acid Profile in Heat-Stressed Broiler Chickens" Antioxidants 10, no. 8: 1265. https://doi.org/10.3390/antiox10081265
APA StyleSalah, A. S., Ahmed-Farid, O. A., Nassan, M. A., & El-Tarabany, M. S. (2021). Dietary Curcumin Improves Energy Metabolism, Brain Monoamines, Carcass Traits, Muscle Oxidative Stability and Fatty Acid Profile in Heat-Stressed Broiler Chickens. Antioxidants, 10(8), 1265. https://doi.org/10.3390/antiox10081265