1. Introduction
Both circular economy and bioeconomy are considered as alternative economic production models that are crucial to promote sustainable growth and development [
1]. The main goal of both models is to develop and achieve synergies among the economy, environment, and society. In the European Union, both agricultural and agro-industrial production processes produce annually 89 Mtons of biomass as waste [
2]. Amongst these by-products, vital bioactive compounds with high added value and significant potential for utilization as feed additives are discarded as well [
3,
4]. Grape is considered to be the world’s largest fruit crop, with an annual production exceeding 67 Mtons. Within the Mediterranean basin area, the by-products of the winery industry are broadly available, as 62% of global wine production is located in this area [
5]. Approximately 20% of the entire grapes’ biomass used for wine production is wasted. The waste is composed of stems, the woody part of the grapevine and grape pomace, and the solid residue of vinification consisting of skins, stems and seeds [
6]. These biomaterials are usually discarded into nearby open fields for biodegradation, thus polluting the environment and water reservoirs in the vicinity [
7]. Numerous studies have highlighted the potential of these by-products for use as plant originated feed additives that are predominantly rich in a broad variety of polyphenols [
8,
9], especially flavonoids, such as catechins and procyanidins with varying degrees of polymerization [
10]. In this context, there are many studies and research endeavors demonstrating the powerful antioxidant properties of these polyphenols, as pure compounds and/or extracts, and their ability to act as potent free radical scavengers for the improvement of an organism’s oxidative balance [
8].
Since the use of antibiotics as growth promoters was banned in the EU for poultry production in 2006, several management and nutritional strategies in the poultry industry were proposed in order to maintain high standards of productivity, healthiness, and welfare [
9]. More specifically, in broilers, dietary supplementation with grape pomace appeared to effectively substitute vitamin E, preserving the antioxidant capacity in ileal content, excreta, and muscle tissues [
8]. This perspective of utilizing natural compounds with vast antioxidant potentials for the substitution of synthetic substances has gained significant attention due to various safety issues raised over time [
11]. On the other hand, the use of synthetic molecules such as butylated hydroxyl anisole (BHA) and butylated hydroxytoluene (BHT) has been linked with possible toxicity, which has been correlated to side effects on the liver and carcinogenesis in animal studies [
11,
12]. Thus, there is a growing demand of consumers towards the replacement of synthetic with natural antioxidants. The utilization of vinification by-products for the production of natural antioxidants enables producers to satisfy this trend in an economically affordable process and contributes to environment preservation affording added-value functional foods as cleaner label products.
However, the high crude fiber content in combination with some anti-nutrient factors (condensed tannins) of grape pomace may have negative nutritional effects in broilers [
13]. Indeed, Kumanda et al. [
14] found that the dietary inclusion of grape pomace at higher than 7.5% decreased the average feed intake in broilers. This limitation constitutes a serious drawback for the utilization of grape pomace as animal feed. Aiming to facilitate the supply of beneficial antioxidant compounds of grape pomace in animals, many strategies have been suggested, such as the treating of grape pomace with polyethylene glycol, a well-justified tannin-binding compound [
13]. However, under a valorization perspective, such a process appears to be cost-ineffective and resource-consuming. The extraction of polyphenolic extracts of grape leftovers seems to be a more promising sustainable and effective strategy to supply both animals and their products with antioxidant compounds without adversely affecting growth performance and diet efficiency. Indeed, in the study of Iqbal et al. [
15], the substitution of vitamin E by a polyphenolic extract of grape by-products enhanced the antioxidant status and the immunity of broilers at a lower feed cost without any side effects on the physiology or health of chickens. However, it is still questionable whether such extracts could effectively substitute the antioxidant supply of grape pomace considering both economical and physiological perspectives.
Another by-product resulting after the wine-making process with high potential as a feed additive but lacking scientific attention is wine lees. Wine lees represent approximately 5% of wine production, mainly containing ethanol, tartrate salts, phenolic compounds, and yeast cell walls [
16]. According to the Greek national regulation (Joint Ministerial Decision, No. 50910/2727, 2003), ethanol that has been isolated from wine lees has received a license to be commercialized. Moreover, due to their high antioxidant capacity, wine lees have been introduced as preservatives to replace the most commonly used synthetic additives in meat products as well. Alaracon et al. [
17] reported a decreased lipid and protein oxidation of deer meat that had been treated with 2.5 and 5% wine lees. Without narrowing out on wine lees’ antioxidant properties, their pivotal role as prebiotics portrays an additional important potential as feed additives as well. More specifically, a higher survival rate has been found in
Lactobacillus and
Bifidobacterium genera when they were treated with wine lees [
18]. Although wine lees are a promising by-product for valorization, no information exists regarding their impact on animals’ performance.
Although dietary supplementation with grape by-products has been satisfactorily studied in broilers’ growth performance, scarce information exists regarding their impact on their organisms’ antioxidative status. Considering these issues, the objective of this study was to evaluate the impact of three winery by-products (grape pomace, stem extract, and wine lees) on the oxidative status of broiler chickens.
4. Discussion
Many studies have underlined the importance of vinification by-products as plant materials particularly rich in a broad variety of polyphenols. Grape seeds and skins contained in grape pomace constitute important sources of flavonoids, mainly catechins and procyanidins [
6,
7]. The implementation of LC-MS/MS analysis proved to be a tool of implacable importance that provided us a broad screening of the polyphenolic composition present in grape by-products. Amongst them, procyanidin B1 and B2, gallic acid, caftaric acid, catechin, quercetin, and resveratrol were determined as the prevailing compounds. In a study concerning the analysis of grape seeds from both red and white varieties, they were also found to be rich in catechin, epicatechin, epicatechin, protocatechic acid, procyanidin B1, procyanidin B2, procyanidin B3, and procyanidin B4. Lower concentrations of gallic acid and protocatechic acid were found as well [
49]. Puiggròs et al. [
50] demonstrated that grape seed procyanidin extracts are capable of modulating the expression of antioxidant systems, indicating that procyanidin extracts of grape seeds could act to improve the cellular oxidative status through glutathione synthesis pathways. Additionally, Preuss et al. [
51] have reported that the supplementation of proanthocyanidin extracts from grape seeds significantly decreased the concentrations of total cholesterol, oxidized LDL, and LDL after two months in hypercholesterolemic participants.
Numerous enzymes and cellular processes produce reactive oxygen species (ROS), including the mitochondrial electron transport chain, nitric oxide synthases (NOSs), cytochrome P450 reductase, and xanthine oxidase. Nevertheless, in most of these mechanisms, ROS formation results as a by-product of their catalytic function or from a dysfunctional variant of the enzyme. On the contrary, NADPH oxidases are the only enzymes whose principal role is to generate superoxide and consequently other ROS [
52]. NOX family proteins are the catalytic, electron-transporting subunits of the NADPH oxidase enzyme complex [
53]. In our study, the relative transcript level of NOX2 tended to decrease in the liver of PE-fed birds, indicating a lower production of ROS. Interestingly, it has been found that polyphenols, excepting their well-documented ROS scavenging abilities, downregulate NADPH oxidase in numerous tissues of rats and humans including, but not limited to, vessels and platelets [
54,
55]. Notably, new entry polyphenolic compounds that lack typical superoxide scavenging properties and directly inhibit NOX activity are being studied. Steffen et al. [
56] tested the role of several polyphenols in oxidative stress by establishing a set of structural requirements for scavenging ROS and inhibiting NADPH oxidase function. More specifically, polyphenols such as catechin, epicatechin, quercetin, luteolin, and fisetin scavenge the unstable superoxide due to their lack of additional substitutions in their B ring. However, the intake of the aforementioned compounds was not higher in the PE groups. These sets of evidence support that the polyphenols’ composition (profile) may tightly regulate the in vivo antioxidant mechanisms rather than their absolute level per se.
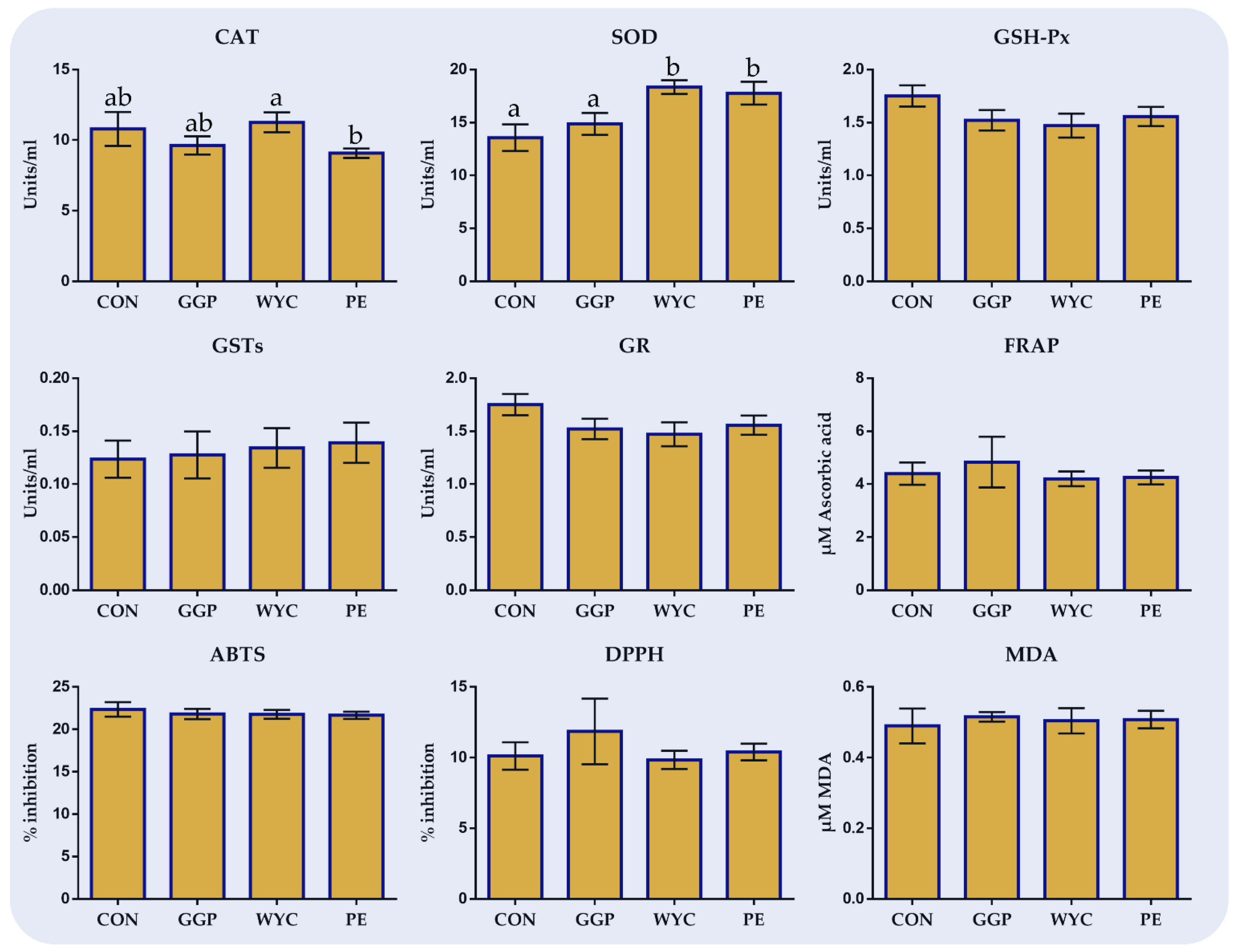
The inclusion of dried wine lees extracts in broilers’ diets tended to increase the relative expression levels of
SOD1 and
GPX1 in the liver of WYC-fed birds. The study of Spanier et al. [
57] showed that resveratrol, an effective polyphenol contained in grapes, reduced NOX4 while increasing SOD1 and GPX1 mRNA levels in human umbilical endothelial cells, which was associated with reduced ROS levels. Furthermore, Hu et al. [
58] reported that dietary supplementation of resveratrol can inhibit lipid peroxidation and improve antioxidant enzymes’ (SOD, GSH-Px, CAT) activity in rats’ hepatocytes as well. Interestingly, the intake of resveratrol in WYC groups was lower compared to other grape by-product treatments, showing the potential involvement of other compounds in SOD1 and GPX1 regulation. The rise of SOD mRNA levels in broilers’ liver caused an increase in the enzymatic level in the blood of WYC- and PE-fed groups compared to the CON. SODs depict the first defense against ROS-mediated injury. SODs catalyze the dismutation of superoxide anion free radical (O
2−) into molecular oxygen and the less harmful ROS, hydrogen peroxide (H
2O
2), decreasing the O
2− level, which damages the cells at extensive concentrations [
59]. In compliance with our findings, supplementation with
Echinacea purpurea L. rich in caftaric and cichoric acids in broilers’ diets increased the activity of SOD in both blood serum and liver [
60].
On the contrary, GSH-Px activity in blood tended to decrease in the WYC group even though the GPX1 mRNA levels in the liver tended to increase. Overlooking the potential involvement of post-transcriptional factors and discrepancies amongst tissue responses [
61], the suppression of GSH-Px activity in blood may lie in SOD upregulation. More specifically, the neutralization of superoxide anion by SOD in blood and subsequently the formation of hydrogen peroxide, the main substrate of both CAT and GSH-Px, may result in an inhibitory feedback. Still, it has been previously found that a high concentration of H
2O
2 inhibits the activity of GSH-Px [
62], while the hydroxyl radical formed by a Fenton reaction using H
2O
2 as a substrate inhibits CAT activity as well [
63].
Total antioxidant capacity was measured in both blood plasma and breast muscles, aiming to investigate the overall oxidative status of tissues. Three methods (FRAP, ABTS, and DPPH) were used, aiming to expand methods’ completability by observing a much broader and dependable perspective. Notably, the FRAP method underestimates the level of the principal exogenous antioxidant, glutathione, compared to the ABTS method [
64]. Concerning the FRAP method, its principal contributors in mammals’ biological samples are uric acid, α-tocopherol, bilirubin, and ascorbic acid, while it does not evaluate the SH-group encompassing non-enzymatic antioxidants, such as glutathione and albumin [
64]. DPPH is a rarely used method in biological samples since the organic compounds are precipitated in their alcoholic medium [
64]. However, DPPH rather complements other methods since it measures substances of non-protein origin substances. Therefore, since there is no optimal assay to evaluate the total antioxidant potential of biological samples, the synergistic use of various assays is strongly suggested, validating changes in the total antioxidant status. In our study, although TAC did not differ in blood plasma, a significant increase in the breast muscle of PE-fed broilers was found according to the FRAP method. This result may be correlated to the overall improvement in the oxidative status in PE groups resulting from
NOX2 downregulation in the liver and an SOD increase in blood plasma.
MDA is one of the main intermediates between lipid peroxidation and oxidative stress [
40]. The rise in MDA levels, which reflects the degree of lipid peroxidation, could be attributed to the increased levels of ROS. Even though blood plasma MDA levels were not affected amongst the dietary treatments, their levels in the breast muscle of WYC- and PE-fed birds were significantly lower. Indeed, it has been previously reported that polyphenols and flavonoids could suppress lipid peroxidation due to their ROS scavenging properties [
58]. MDA is the utmost significant aldehyde formed in the secondary lipid oxidation of polyunsaturated fatty acids (PUFAs) [
40]. In our study, MDA levels in breast muscle amongst the dietary groups ranged between 0.38 and 0.58 mg MDA/kg of tissue. Levels between 0.02 and 2.55 mg MDA/kg have been suggested as acceptable limits for non-rancid meat [
65]. The obtained results indicate an improvement in breast meat oxidative stability. In agreement with our study, the supplementation of grape seeds and skin meal in broilers’ diet significantly decreased the TBARS levels of thigh meat, indicating the synergetic action of γ- and a-tocopherols for stabilizing lipid peroxidation [
66]. Similarly, the inclusion of green tea extract rich in catechins, epigallocatechin-3-gallate, and caffeine in quail diet decreased the MDA concentration in liver and blood plasma [
67].
The nutritional requirements and therefore the composition of broilers’ diets have been extensively studied [
68]. Thus, there is limited plasticity to implement radical changes and substitutions. Aiming to include feed additives rich in antioxidant compounds but poor in nutritional value, soybean meal and/or soybean oil are usually recruited to balance such substitutions. In our study, the proportion of soybean oil was increased in the GGP group to balance the energy content. Soybean oil is not only rich in PUFAs, predominantly linoleic acid (LA), which are prone to oxidation within the organism, but also their inclusion in concentrate mix could enhance its autooxidation [
69]. Thus, the increase in the dietary PUFA level in broilers could induce a cascade of prooxidant incidences [
70]. Without narrowing out on soybean oil’s PUFAs in the GGP group, the grape pomace contained a high proportion of ether extract, with the dominant fatty acid being linoleic acid as well. Considering the aforementioned points, there was strong evidence to assume that the increased PUFA inclusion in the GGP group could disturb the oxidative stability of both broilers’ organism and meat [
71]. Thus, it is plausible to assume that the high content of polyphenols presented in grape pomace may inhibit a likely induced oxidative burst. Although no significant improvement in the oxidative status of the GGP group was observed, it remains unknown what the PUFA increase would induce in the absence of the antioxidant compounds that were included in grape pomace. Considering the above, the discriminant analysis allocated the PE and WYC groups together since both their polyphenolic compositions and levels were quite comparable. On the contrary, the GGP group was mapped away due to the altered response to dietary treatment attributed to both higher ether extract (PUFA) and polyphenol levels.
Finally, increased mortality (5%; 3 out of 60 birds) in the WYC group was observed during the first week of the starting period, indicating the well-documented stressed transition period of chicks in early life [
72]. Even though this proportion tended to differ compared to the CON group, it was within the rational range from an animal scientist’s point of view according to the National Chicken Council [
72].


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}