
Avocado Seeds Relieve Oxidative Stress-Dependent Nephrotoxicity but Enhance Immunosuppression Induced by Cyclosporine in Rats

 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant and Chemicals
2.2. Diet Preparation
2.3. Detection of Bioactive Compounds by GC-MS
2.4. Experimental Design
2.5. Growth-Related Parameters
2.6. Sampling
2.7. Biochemical Analysis
2.8. Comet Assay
2.9. Molecular Analysis by Real-Time PCR
2.10. Histopathology Examinations
2.11. Statistical Analysis
3. Results
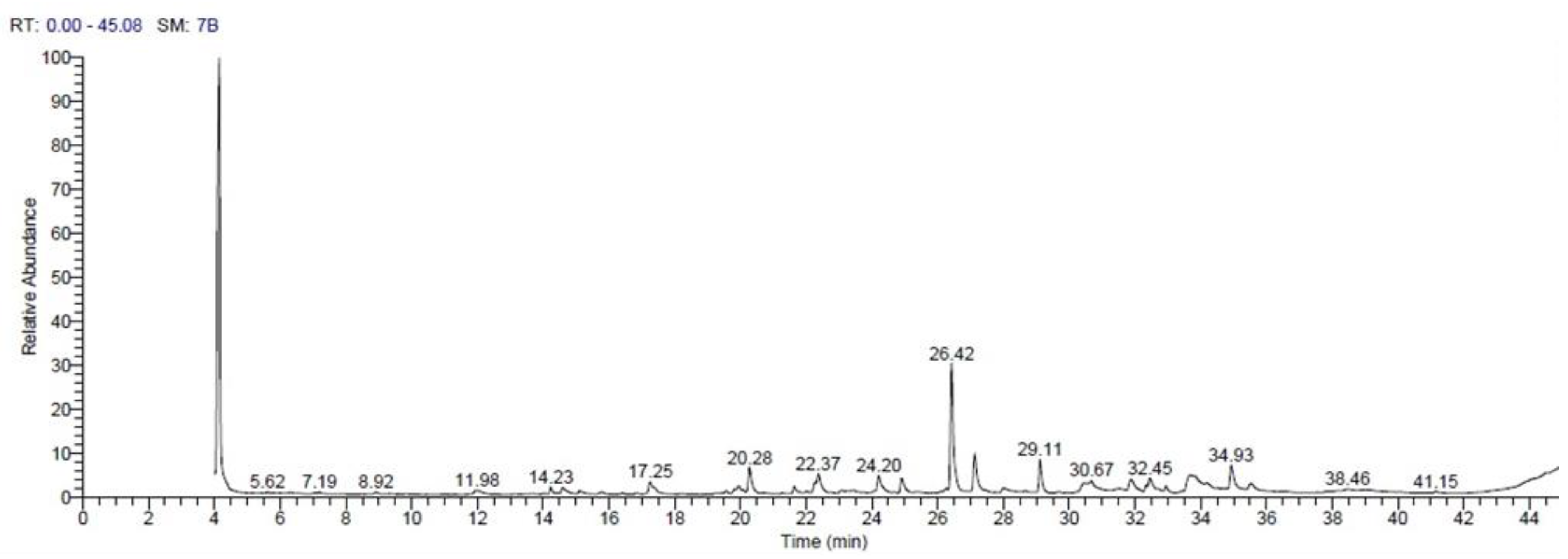
3.1. GC-MS Analysis of Avocado Seed Extract
3.2. Growth-Related Parameters and Biochemical Analysis
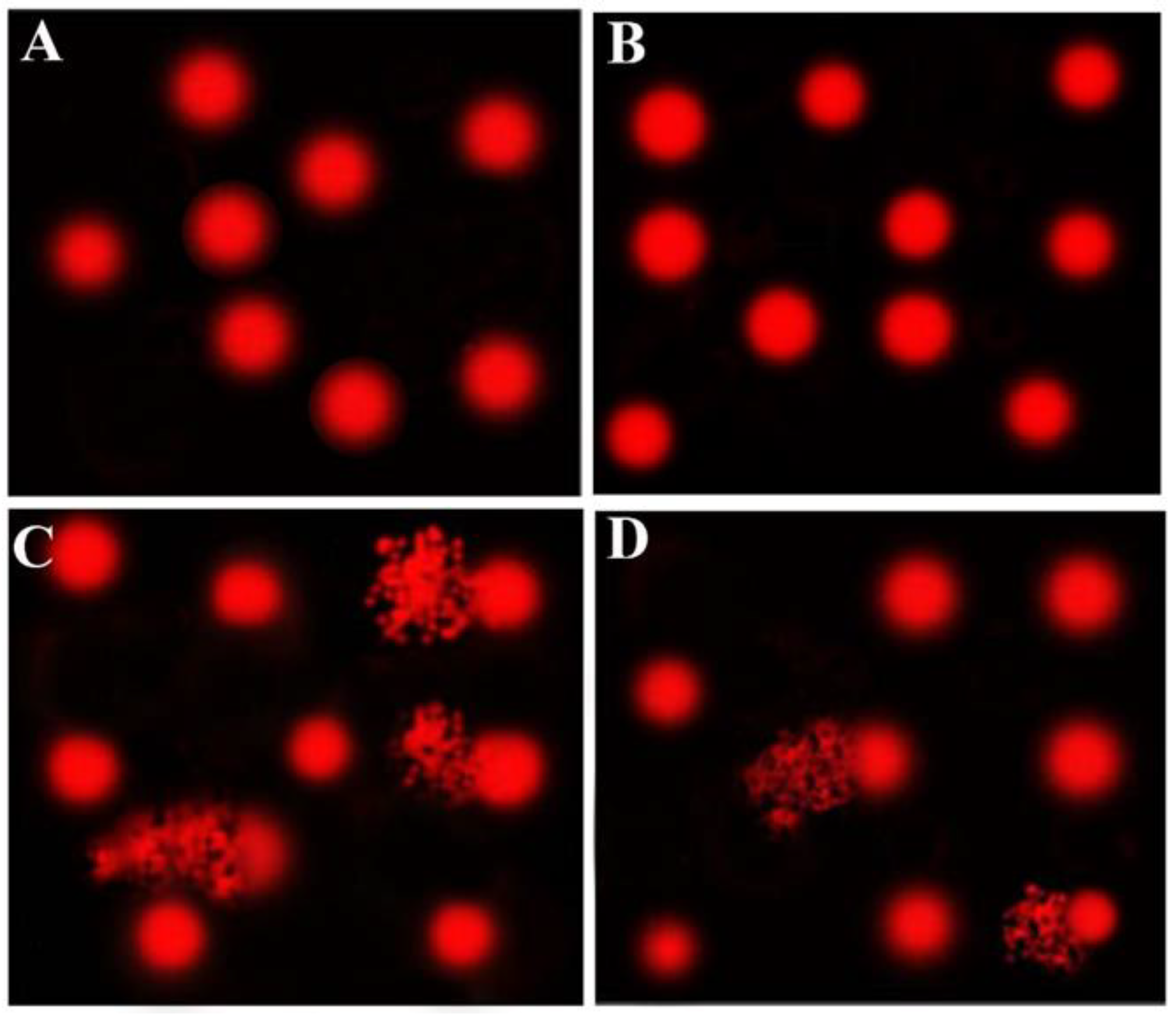
3.3. Comet Assay Results
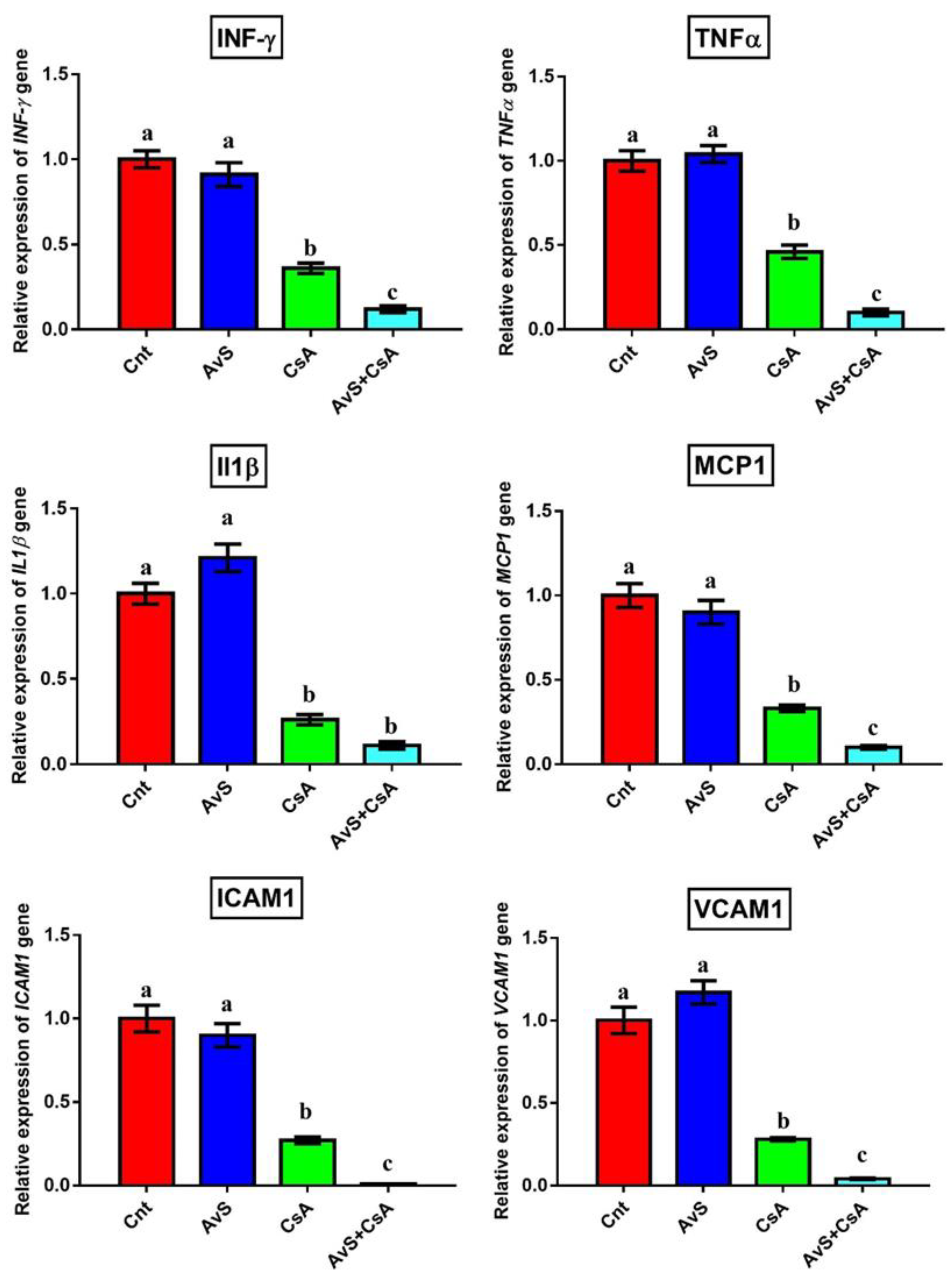
3.4. Molecular Analysis
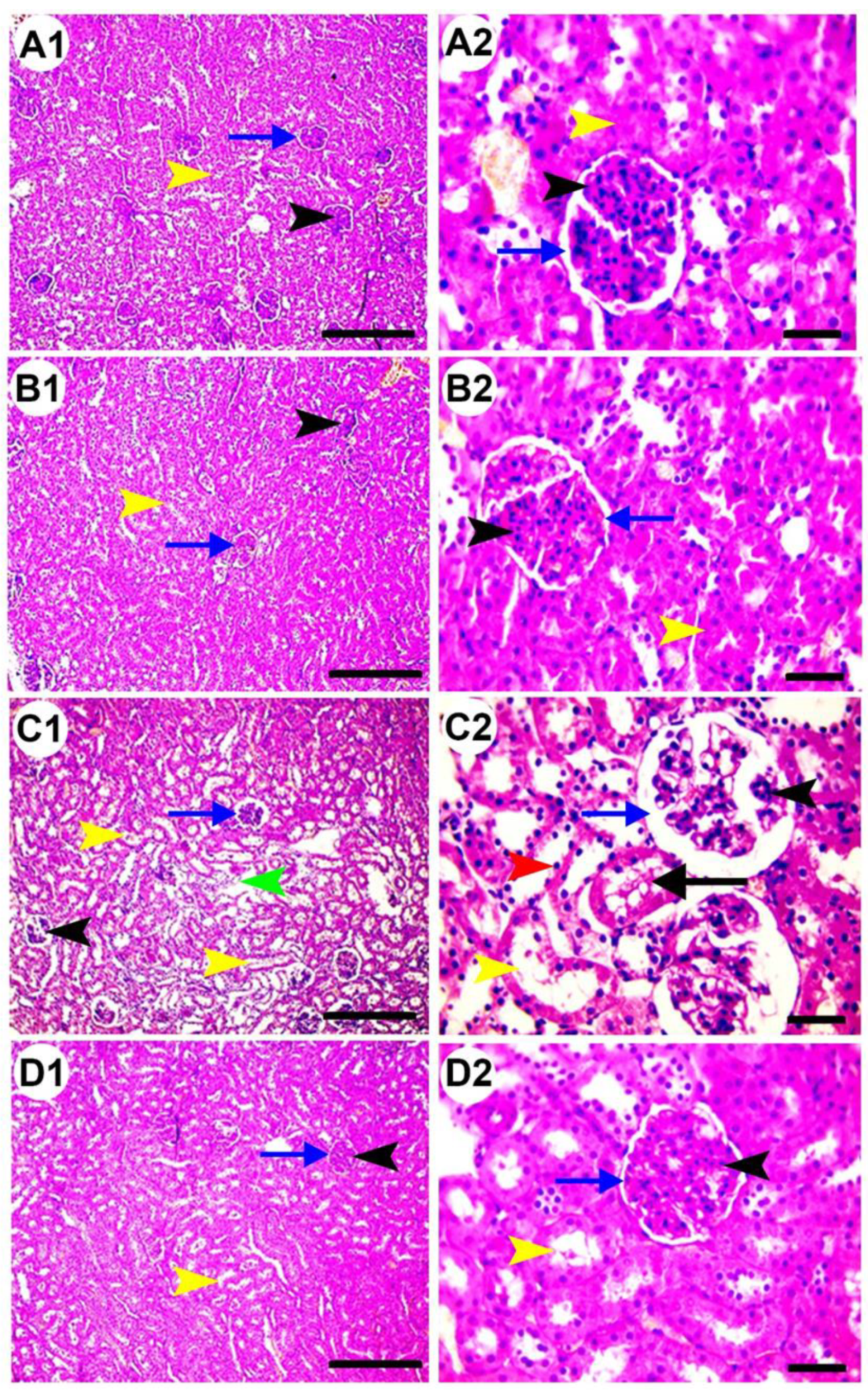
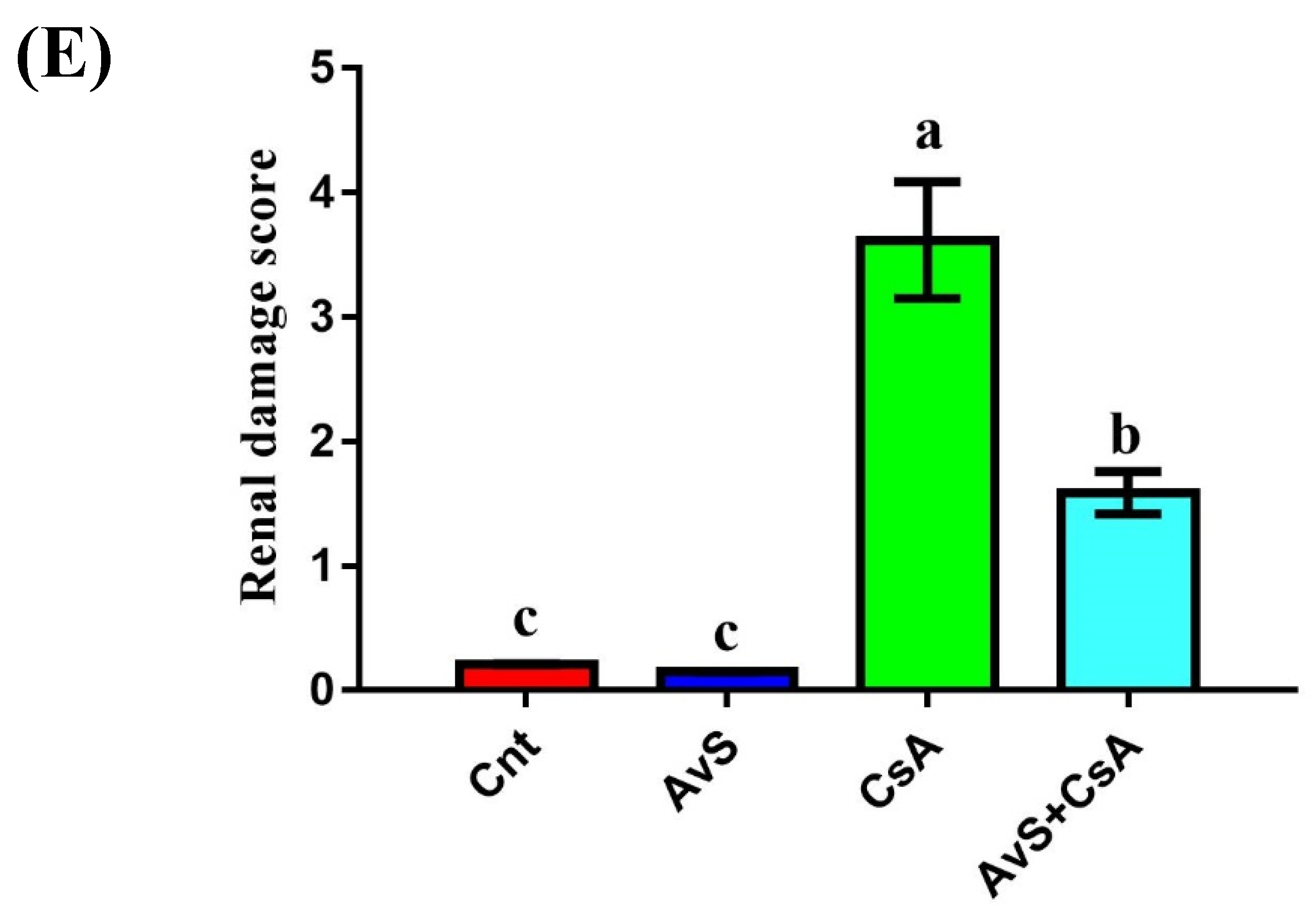
3.5. Histopathological Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Türk, G.; Ateşşahin, A.; Sönmez, M.; Yüce, A.; Ceribaşi, A.O. Lycopene protects against cyclosporine a-induced testicular toxicity in rats. Theriogenology 2007, 67, 778–785. [Google Scholar] [CrossRef]
- Zimmermann, S.; Sekula, P.; Venhoff, M.; Motschall, E.; Knaus, J.; Schumacher, M.; Mockenhaupt, M. Systemic immunomodulating therapies for stevens-johnson syndrome and toxic epidermal necrolysis: A systematic review and meta-analysis. JAMA Dermatol. 2017, 153, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, D.; Haragsim, L. Cyclosporine: A review. J. Transplant. 2012, 2012, 230386. [Google Scholar] [CrossRef] [PubMed]
- Türk, G.; Sönmez, M.; Ceribaşi, A.O.; Yüce, A.; Ateşşahin, A. Attenuation of cyclosporine a-induced testicular and spermatozoal damages associated with oxidative stress by ellagic acid. Int. Immunopharmacol. 2010, 10, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Ghazipour, A.M.; Shirpoor, A.; Ghiasi, R.; Pourheydar, B.; Khalaji, N.; Naderi, R. Cyclosporine a induces testicular injury via mitochondrial apoptotic pathway by regulation of mir-34a and sirt-1 in male rats: The rescue effect of curcumin. Chem. Biol. Interact. 2020, 327, 109180. [Google Scholar] [CrossRef]
- Josephine, A.; Amudha, G.; Veena, C.K.; Preetha, S.P.; Varalakshmi, P. Oxidative and nitrosative stress mediated renal cellular damage induced by cyclosporine a: Role of sulphated polysaccharides. Biol. Pharm. Bull. 2007, 30, 1254–1259. [Google Scholar] [CrossRef][Green Version]
- da Silva, J.B.; de Melo Lima, M.H.; Secoli, S.R. Influence of cyclosporine on the occurrence of nephrotoxicity after allogeneic hematopoietic stem cell transplantation: A systematic review. Rev. Bras. Hematol. Hemoter. 2014, 36, 363–368. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, X.; Nepovimova, E.; Wang, Y.; Yang, H.; Kuca, K. Mechanism of cyclosporine a nephrotoxicity: Oxidative stress, autophagy, and signalings. Food Chem. Toxicol. 2018, 118, 889–907. [Google Scholar] [CrossRef]
- Rao, S.R.; Sundararajan, S.; Subbarayan, R.; Murugan Girija, D. Cyclosporine-a induces endoplasmic reticulum stress and influences pro-apoptotic factors in human gingival fibroblasts. Mol. Cell. Biochem. 2017, 429, 179–185. [Google Scholar] [CrossRef]
- Chanderbali, A.S.; Albert, V.A.; Ashworth, V.E.; Clegg, M.T.; Litz, R.E.; Soltis, D.E.; Soltis, P.S. Persea americana (avocado): Bringing ancient flowers to fruit in the genomics era. BioEssays 2008, 30, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carpena, J.G.; Morcuende, D.; Andrade, M.J.; Kylli, P.; Estévez, M. Avocado (Persea americana mill.) phenolics, in vitro antioxidant and antimicrobial activities, and inhibition of lipid and protein oxidation in porcine patties. J. Agric. Food Chem. 2011, 59, 5625–5635. [Google Scholar] [CrossRef]
- Segovia, F.J.; Corral-Pérez, J.J.; Almajano, M.P. Avocado seed: Modeling extraction of bioactive compounds. Ind. Crop. Prod. 2016, 85, 213–220. [Google Scholar] [CrossRef]
- Plaza, L.; Sánchez-Moreno, C.; de Pascual-Teresa, S.; de Ancos, B.; Cano, M.P. Fatty acids, sterols, and antioxidant activity in minimally processed avocados during refrigerated storage. J. Agric. Food Chem. 2009, 57, 3204–3209. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.A.; Dorantes, A.L.; Gallndez, M.J.; Cardenas, S.E. Effect of a novel oil extraction method on avocado (Persea americana mill) pulp microstructure. Plant Foods Hum. Nutr. 2004, 59, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Camberos, E.; Martínez-Velázquez, M.; Flores-Fernández, J.M.; Villanueva-Rodríguez, S. Acute toxicity and genotoxic activity of avocado seed extract (Persea americana mill., c.V. Hass). Sci. World J. 2013, 2013, 245828. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Bostic, T.R.; Gu, L. Antioxidant capacities, procyanidins and pigments in avocados of different strains and cultivars. Food Chem. 2010, 122, 1193–1198. [Google Scholar] [CrossRef]
- Jiménez-Arellanes, A.; Luna-Herrera, J.; Ruiz-Nicolás, R.; Cornejo-Garrido, J.; Tapia, A.; Yépez-Mulia, L. Antiprotozoal and antimycobacterial activities of Persea americana seeds. BMC Complementary Altern. Med. 2013, 13, 109. [Google Scholar] [CrossRef]
- Abubakar, A.N.F.; Achmadi, S.S.; Suparto, I.H. Triterpenoid of avocado (Persea americana) seed and its cytotoxic activity toward breast mcf-7 and liver hepg2 cancer cells. Asian Pac. J. Trop. Biomed. 2017, 7, 397–400. [Google Scholar] [CrossRef]
- Uchenna, U.E.; Shori, A.B.; Baba, A.S. Inclusion of avocado (Persea american) seeds in the diet to improve carbohydrate and lipid metabolism in rats. Rev. Argent. Endocrinol. Metab. 2017, 54, 140–148. [Google Scholar]
- Dao, P.T.A.; An, N.T.H.; Thuy, N.T.T.; Tuyet, N.T.A.; Truc, T.T.M. Screening on Antioxidant Activities of by-Products from Vegetables and Fruits in Tay Nguyen Region and Applying for Shrimp cold Storage. In Proceedings of the 2016 3rd International Conference on Green Technology and Sustainable Development (GTSD), Kaohsiung, Taiwan, 24–25 November 2016; pp. 93–97. [Google Scholar]
- Saad, S.A. The antioxidant activity of avocado seeds on immunosuppression induced by cyclosporine in rats. Home Econ. J. 2020, 36, 93–109. [Google Scholar]
- Reeves, P.G.; Rossow, K.L.; Lindlauf, J. Development and testing of the ain-93 purified diets for rodents: Results on growth, kidney calcification and bone mineralization in rats and mice. J. Nutr. 1993, 123, 1923–1931. [Google Scholar] [CrossRef] [PubMed]
- Fossati, P.; Prencipe, L.; Berti, G. Use of 3,5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clin. Chem. 1980, 26, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Patton, C.J.; Crouch, S.R. Spectrophotometric and kinetics investigation of the berthelot reaction for the determination of ammonia. Anal. Chem. 1977, 49, 464–469. [Google Scholar] [CrossRef]
- Bartels, H.; Böhmer, M.; Heierli, C. Serum creatinine determination without protein precipitation. Clin. Chim. Acta Int. J. Clin. Chem. 1972, 37, 193–197. [Google Scholar] [CrossRef]
- Bontin, S. Presence of enzyme system of mammalian tissue. Wiley Inter. Sci. 1970, 197, 257–363. [Google Scholar]
- Mesbah, L.; Soraya, B.; Narimane, S.; Jean, P.F. Protective effect of flavonides against the toxicity of vinblastine cyclophosphamide and paracetamol by inhibition of lipid-peroxydation and increase of liver glutathione. Haematology 2004, 7, 59–67. [Google Scholar]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. In Methods in Enzymology, 3rd ed.; Lippincott-Raven Publishers: Philadelphia, PA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Hafeman, D.G.; Sunde, R.A.; Hoekstra, W.G. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutr. 1974, 104, 580–587. [Google Scholar] [CrossRef]
- El-Demerdash, F.M.; El-Magd, M.A.; El-Sayed, R.A. Panax ginseng modulates oxidative stress, DNA damage, apoptosis, and inflammations induced by silicon dioxide nanoparticles in rats. Environ. Toxicol. 2001, 36, 362–1374. [Google Scholar]
- Chang, C.L.; Sung, P.H.; Chen, K.H.; Shao, P.L.; Yang, C.C.; Cheng, B.C.; Lin, K.C.; Chen, C.H.; Chai, H.T.; Chang, H.W.; et al. Adipose-derived mesenchymal stem cell-derived exosomes alleviate overwhelming systemic inflammatory reaction and organ damage and improve outcome in rat sepsis syndrome. Am. J. Transl. Res. 2018, 10, 1053–1070. [Google Scholar] [PubMed]
- Zahran, R.; Ghozy, A.; Elkholy, S.S.; El-Taweel, F.; El-Magd, M.A. Combination therapy with melatonin, stem cells and extracellular vesicles is effective in limiting renal ischemia–reperfusion injury in a rat model. Int. J. Urol. 2020, 27, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Ciarcia, R.; Damiano, S.; Florio, A.; Spagnuolo, M.; Zacchia, E.; Squillacioti, C.; Mirabella, N.; Florio, S.; Pagnini, U.; Garofano, T.; et al. The protective effect of apocynin on cyclosporine a-induced hypertension and nephrotoxicity in rats. J. Cell. Biochem. 2015, 116, 1848–1856. [Google Scholar] [CrossRef]
- Ouyang, Z.; Cao, W.; Zhu, S.; Liu, X.; Zhong, Z.; Lai, X.; Deng, H.; Jiang, S.; Wang, Y. Protective effects of 2-deoxy-d-glucose on nephrotoxicity induced by cyclosporine a in rats. Int. J. Clin. Exp. Pathol. 2014, 7, 4587–4595. [Google Scholar] [PubMed]
- Bellwon, P.; Culot, M.; Wilmes, A.; Schmidt, T.; Zurich, M.G.; Schultz, L.; Schmal, O.; Gramowski-Voss, A.; Weiss, D.G.; Jennings, P.; et al. Cyclosporine a kinetics in brain cell cultures and its potential of crossing the blood-brain barrier. Toxicol. Vitr. 2015, 30, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Haleagrahara, N.; Yee, T.; Chakravarthi, S.; Lee, N. Protective effect of n-acetylcysteine on cyclosporine a-induced changes in lipid hydroperoxide levels and renal dysfunction in rats. Arch. Med. Sci. 2009, 5, 16–22. [Google Scholar]
- Helmy, M.M.; Helmy, M.W.; El-Mas, M.M. Upregulation of cystathionine-γ-lyase/hydrogen sulfide pathway underlies the celecoxib counteraction of cyclosporine-induced hypertension and renal insult in rats. Prostaglandins Other Lipid Mediat. 2019, 141, 1–10. [Google Scholar] [CrossRef]
- Kim, J.S.; Yang, J.W.; Han, B.G.; Kwon, H.J.; Kim, J.H.; Choi, S.O. Protective role of apelin against cyclosporine-induced renal tubular injury in rats. Transplant. Proc. 2017, 49, 1499–1509. [Google Scholar] [CrossRef]
- Adekunle, I.A.; Imafidon, C.E.; Oladele, A.A.; Ayoka, A.O. Ginger polyphenols attenuate cyclosporine-induced disturbances in kidney function: Potential application in adjuvant transplant therapy. Pathophysiology 2018, 25, 101–115. [Google Scholar] [CrossRef]
- Deppe, C.E.; Heering, P.J.; Tinel, H.; Kinne-Saffran, E.; Grabensee, B.; Kinne, R.K. Effect of cyclosporine a on na+/k(+)-atpase, na+/k+/2cl- cotransporter, and h+/k(+)-atpase in mdck cells and two subtypes, c7 and c11. Exp. Nephrol. 1997, 5, 471–480. [Google Scholar]
- Yoon, H.E.; Yang, C.W. Established and newly proposed mechanisms of chronic cyclosporine nephropathy. Korean J. Intern. Med. 2009, 24, 81–92. [Google Scholar] [CrossRef]
- Lee, J. Use of antioxidants to prevent cyclosporine a toxicity. Toxicol. Res. 2010, 26, 163–170. [Google Scholar] [CrossRef]
- Capasso, G.; Di Gennaro, C.I.; Della Ragione, F.; Manna, C.; Ciarcia, R.; Florio, S.; Perna, A.; Pollastro, R.M.; Damiano, S.; Mazzoni, O.; et al. In vivo effect of the natural antioxidant hydroxytyrosol on cyclosporine nephrotoxicity in rats. Nephrol. Dial. Transplant. 2008, 23, 1186–1195. [Google Scholar] [CrossRef]
- Abdelhady, D.H.; El-Magd, M.A.; Elbialy, Z.I.; Saleh, A.A. Bromuconazole-induced hepatotoxicity is accompanied by upregulation of pxr/cyp3a1 and downregulation of car/cyp2b1 gene expression. Toxicol. Mech. Methods 2017, 27, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Abu Gazia, M.; El-Magd, M.A. Effect of pristine and functionalized multiwalled carbon nanotubes on rat renal cortex. Acta Histochem. 2018, 121, 207–217. [Google Scholar] [CrossRef] [PubMed]
- El-Magd, M.A.; Kahilo, K.A.; Nasr, N.E.; Kamal, T.; Shukry, M.; Saleh, A.A. A potential mechanism associated with lead-induced testicular toxicity in rats. Andrologia 2016, 49, e12750. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Ramnath, R.D.; Foster, R.R.; Wylie, E.C.; Friden, V.; Dasgupta, I.; Haraldsson, B.; Welsh, G.I.; Mathieson, P.W.; Satchell, S.C. Reactive oxygen species modulate the barrier function of the human glomerular endothelial glycocalyx. PLoS ONE 2013, 8, e55852. [Google Scholar] [CrossRef]
- Cao, Y.; Roursgaard, M.; Danielsen, P.H.; Moller, P.; Loft, S. Carbon black nanoparticles promote endothelial activation and lipid accumulation in macrophages independently of intracellular ros production. PLoS ONE 2014, 9, e106711. [Google Scholar] [CrossRef]
- Rao, U.S.M.; Ponnusamy, K.; Naidu, J.; Sundaram, S. Modulatory influence of avocado on renal oxido -lipidemic stress and mrna expression of nos in renal artery studied in nephropathy induced rats. Int. Med. J. 2014, 21, 1–7. [Google Scholar]
- Abdel Moneim, A.; Ahmed, O.; Fahim, H.; Mohamed, E. The preventive effects of avocado fruit and seed extracts on cardio-nephrotoxicity induced by diethylnitrosamine/2-acetylaminoflurine in wistar rats. Basic Sci. Med. 2017, 6, 4–13. [Google Scholar]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef]
- Rosero, J.C.; Cruz, S.; Osorio, C.; Hurtado, N. Analysis of phenolic composition of byproducts (seeds and peels) of avocado (Persea americana mill.) cultivated in colombia. Molecules 2019, 24, 3209. [Google Scholar] [CrossRef]
- Anshar, A.; Bahar, M.A.; Ikliptikawati, D. The effect of avocado to the profile of blood urea nitrogen (bun) and creatinine in rats (rattus norvegicus) induced with meloxicam. J. Ris. Vet. Indones. (J. Indones. Vet. Res.) 2018, 2, 1–7. [Google Scholar] [CrossRef]
- Lara-Márquez, M.; Báez-Magaña, M.; Raymundo-Ramos, C.; Spagnuolo, P.A.; Macías-Rodríguez, L.; Salgado-Garciglia, R.; Ochoa-Zarzosa, A.; López-Meza, J.E. Lipid-rich extract from mexican avocado (Persea americana var. Drymifolia) induces apoptosis and modulates the inflammatory response in caco-2 human colon cancer cells. J. Funct. Foods 2020, 64, 103658. [Google Scholar] [CrossRef]
- Tu, H.P.; Chen, Y.T.; Chiu, H.C.; Chin, Y.T.; Huang, S.M.; Cheng, L.C.; Fu, E.; Chiang, C.Y. Cyclosporine a enhances apoptosis in gingival keratinocytes of rats and in oecm1 cells via the mitochondrial pathway. J. Periodontal Res. 2009, 44, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Øzbay, L.A.; Smidt, K.; Mortensen, D.M.; Carstens, J.; Jørgensen, K.A.; Rungby, J. Cyclosporin and tacrolimus impair insulin secretion and transcriptional regulation in ins-1e beta-cells. Br. J. Pharmacol. 2011, 162, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Rincón, J.; Parra, G.; Quiroz, Y.; Benatuil, L.; Rodríguez-Iturbe, B. Cyclosporin a reduces expression of adhesion molecules in the kidney of rats with chronic serum sickness. Clin. Exp. Immunol. 2000, 121, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Daull, P.; Barabino, S.; Feraille, L.; Kessal, K.; Docquier, M.; Parsadaniantz, S.M.; Baudouin, C.; Garrigue, J.S. Modulation of inflammation-related genes in the cornea of a mouse model of dry eye upon treatment with cyclosporine eye drops. Curr. Eye Res. 2019, 44, 476–485. [Google Scholar] [CrossRef]
- Satonaka, H.; Suzuki, E.; Nishimatsu, H.; Oba, S.; Takeda, R.; Goto, A.; Omata, M.; Fujita, T.; Nagai, R.; Hirata, Y. Calcineurin promotes the expression of monocyte chemoattractant protein-1 in vascular myocytes and mediates vascular inflammation. Circ. Res. 2004, 94, 693–700. [Google Scholar] [CrossRef]
- Pflugfelder, S.C.; Stern, M.; Zhang, S.; Shojaei, A. Lfa-1/icam-1 interaction as a therapeutic target in dry eye disease. J. Ocul. Pharmacol. Ther. 2017, 33, 5–12. [Google Scholar] [CrossRef]
- Zhang, W.; Smith, C.; Monette, R.; Hutchison, J.; Stanimirovic, D.B. Indomethacin and cyclosporin a inhibit in vitro ischemia-induced expression of icam-1 and chemokines in human brain endothelial cells. Acta Neurochirurgica Suppl. 2000, 76, 47–53. [Google Scholar]
- Dabas, D.; Elias, R.J.; Ziegler, G.R.; Lambert, J.D. In vitro antioxidant and cancer inhibitory activity of a colored avocado seed extract. Int. J. Food Sci. 2019, 2019, 6509421. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/Kg) | Basal Diet | Basal Diet with 5% Avocado Seed Powder |
|---|---|---|
| Casein (>85% protein) | 120.0 | 120.0 |
| Corn oil | 70.0 | 70.0 |
| Mineral mix | 35.0 | 35.0 |
| Vitamin mix | 10.0 | 10.0 |
| L- cystine | 3.0 | 3.0 |
| Choline bitartrate | 2.5 | 2.5 |
| Wheat bran | 50.0 | 50.0 |
| Sucrose | 100.0 | 100.0 |
| Corn starch | 609.5 | 559.5 |
| Avocado seeds powder | -- | 50.0 |
| Gene | Forward Primer | Reverse Primer | Accession Number |
|---|---|---|---|
| TNFα | GCATGATCCGCGACGTGGAA | AGATCCATGCCGTTGGCCAG | NM_012675.3 |
| IL1β | CACCTCTCAAGCAGAGCACAG | GGGTTCCATGGTGAAGTCAAC | NM_031512.2 |
| MCP1 | TCGCTTCTGACACCATGCA | TGCTACAGGCAGCAAATGTGA | M57441.1 |
| IFN-γ | GATCCAGCACAAAGCTGTCA | GACTCCTTTTCCGCTTCCTT | AF010466.1 |
| ICAM1 | AGATCACATTCACGGTGCTG | CTTCAGAGGCAGGAAACAGG | NM_012967.1 |
| VCAM1 | TGCACGGTCCCTAATGTGTA | TGCCAATTTCCTCCCTTAAA | NM_012889.2 |
| GAPDH | CAACTCCCTCAAGATTGTCAGCAA | GGCATGGACTGTGGTCATGA | NM_017008.4 |
| RT | Compound Name | Area % | Molecular Formula | Molecular Weight |
|---|---|---|---|---|
| 14.23 | 7-Hexadecene | 1.41 | C16H32 | 224 |
| 14.60 | 10-Undecenal | 2.32 | C11H20O | 168 |
| 17.25 | 13-Oxabicyclo[1 0.1.0]tridecan | 4.58 | C12H22O | 182 |
| 19.94 | Tridecanal | 2.22 | C13H26O | 198 |
| 20.28 | Dodecanoic acid, methyl ester | 5.53 | C13H26O2 | 214 |
| 21.64 | 5Oxatricyclo[8.2.0.0(4,6)]dodecane, 12-trimethyl-9-methylene | 1.32 | C15H24O | 220 |
| 22.27 | cis-9-Hexadecenal | 1.71 | C16H30O | 238 |
| 22.37 | Tetradecanal | 4.34 | C14H28O | 212 |
| 24.20 | Cyclopentadecanone | 3.33 | C15H28O | 224 |
| 24.90 | Tetradecanoic acid, methyl ester | 3.37 | C15H30O2 | 242 |
| 26.42 | Z,Z-4,15-Octadecadien-1-ol acetate | 26.06 | C20H36O2 | 308 |
| 27.12 | Aromadendrene oxide-(1) | 8.63 | C15H24O | 220 |
| 29.11 | Hexadecanoic acid, methyl ester | 7.28 | C17H34O2 | 270 |
| 30.67 | Oleic Acid | 1.31 | C18H34O2 | 282 |
| 31.88 | 9-Octadecenoic acid (z) | 3.47 | C18H34O2 | 282 |
| 32.33 | 9,12-Octadecadienoic acid (z,z)-, methyl ester | 1.13 | C19H34O2 | 294 |
| 32.45 | 9-Octadecenoic acid (Z)-, methyl ester | 3.77 | C19H36O2 | 296 |
| 32.94 | Heptadecanoic acid, 16-methyl-, methyl ester | 1.26 | C19H38O2 | 298 |
| 33.67 | 9,12-Octadecadienoic acid | 9.32 | C18H32O2 | 280 |
| 34.92 | 7-Methyl-Z-tetradecen-1-ol acetate | 5.71 | C17H32O2 | 268 |
| 35.53 | Hexadecadienoic acid, methyl ester | 1.94 | C17H30O2 | 266 |
| Groups | FBWG (g) | Total FI (g) | FER |
|---|---|---|---|
| Cnt | 60.00 ± 3.28 b | 501.00 ± 12.79 a | 0.120 ± 0.006 b |
| AvS | 73.33 ± 3.19 a | 475.00 ± 10.43 a | 0.154 ± 0.002 a |
| CsA | 30.33 ± 2.03 d | 351.67 ± 8.67 c | 0.086 ± 0.007 d |
| AvS + CsA | 47.00 ± 2.61 c | 437.00 ± 11.89 b | 0.108 ± 0.004 c |
| Groups | Uric Acid (mg/dL) | Urea (mg/dL) | Creatinine (mg/dL) |
|---|---|---|---|
| Cnt | 2.61 ± 0.10 c | 17.63 ± 0.76 c | 0.72 ± 0.03 c |
| AvS | 2.13 ± 0.12 d | 14.31 ± 0.63 d | 0.64 ± 0.02 d |
| CsA | 6.56 ± 0.21 a | 74.23 ± 2.18 a | 1.46 ± 0.06 a |
| AvS + CsA | 3.62 ± 0.11 b | 32.35 ± 1.06 b | 0.93 ± 0.04 b |
| Groups | MDA (nmol/g Tissue) | SOD (U/g Tissue) | CAT (U/g Tissue) | GPx (U/g Tissue) |
|---|---|---|---|---|
| Cnt | 11.37 ± 0.73 c | 3.75 ± 0.24 a | 3.04 ± 0.19 a | 115.75 ± 7.51 b |
| AvS | 9.70 ± 0.54 d | 3.94 ± 0.28 a | 3.15 ± 0.17 a | 138.11 ± 7.82 a |
| CsA | 35.68 ± 2.72 a | 1.21 ± 0.09 c | 1.46 ± 0.08 c | 37.30 ± 2.36 d |
| AvS + CsA | 18.24 ± 0.91 b | 2.45 ± 0.14 b | 2.25 ± 0.09 b | 92.73 ± 6.00 c |
| Groups | Creatinine Clearance (mL/min/100 g Body wt) | Na+/K+ ATPase (ng/mg) | kidney Weight (g) |
|---|---|---|---|
| Cnt | 1.6 ± 0.04 a | 0.85 ± 0.02 a | 1.66 ± 0.05 bc |
| AvS | 1.14 ± 0.02 c | 0.79 ± 0.005 b | 1.93 ± 0.02 a |
| CsA | 0.82 ± 0.03 d | 0.14 ± 0.004 d | 1.26 ± 0.05 d |
| AvS + CsA | 1.25 ± 0.04 b | 0.55 ± 0.03 c | 1.60 ± 0.03 c |
| Group | Tailed % | Untailed % | Tail Length (µm) | Tail DNA% | Tail Moment * |
|---|---|---|---|---|---|
| Cnt | 2 | 98 | 1.65 ± 0.11 c | 1.10 | 1.82 |
| AvS | 4 | 96 | 1.89 ± 0.12 c | 1.75 | 3.31 |
| CsA | 18 | 82 | 5.63 ± 0.28 a | 4.50 | 25.34 |
| AvS + CsA | 11 | 89 | 3.95 ± 0.19 b | 3.24 | 12.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmoslemany, A.M.; El-Magd, M.A.; Ghamry, H.I.; Alshahrani, M.Y.; Zidan, N.S.; Zedan, A.M.G. Avocado Seeds Relieve Oxidative Stress-Dependent Nephrotoxicity but Enhance Immunosuppression Induced by Cyclosporine in Rats. Antioxidants 2021, 10, 1194. https://doi.org/10.3390/antiox10081194
Elmoslemany AM, El-Magd MA, Ghamry HI, Alshahrani MY, Zidan NS, Zedan AMG. Avocado Seeds Relieve Oxidative Stress-Dependent Nephrotoxicity but Enhance Immunosuppression Induced by Cyclosporine in Rats. Antioxidants. 2021; 10(8):1194. https://doi.org/10.3390/antiox10081194
Chicago/Turabian StyleElmoslemany, Amira M., Mohammed A. El-Magd, Heba I. Ghamry, Mohammad Y. Alshahrani, Nahla S. Zidan, and Amina M. G. Zedan. 2021. "Avocado Seeds Relieve Oxidative Stress-Dependent Nephrotoxicity but Enhance Immunosuppression Induced by Cyclosporine in Rats" Antioxidants 10, no. 8: 1194. https://doi.org/10.3390/antiox10081194
APA StyleElmoslemany, A. M., El-Magd, M. A., Ghamry, H. I., Alshahrani, M. Y., Zidan, N. S., & Zedan, A. M. G. (2021). Avocado Seeds Relieve Oxidative Stress-Dependent Nephrotoxicity but Enhance Immunosuppression Induced by Cyclosporine in Rats. Antioxidants, 10(8), 1194. https://doi.org/10.3390/antiox10081194