
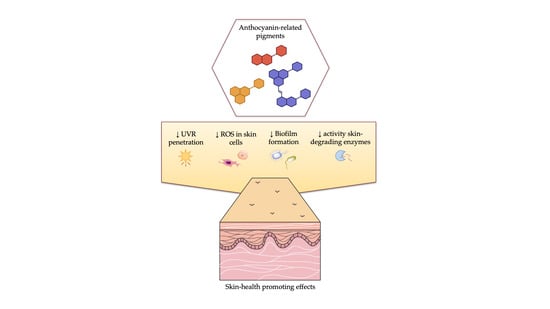
Anthocyanin-Related Pigments: Natural Allies for Skin Health Maintenance and Protection

 ,
, 

 , ,
, ,  , ,
, ,  , ,
, ,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
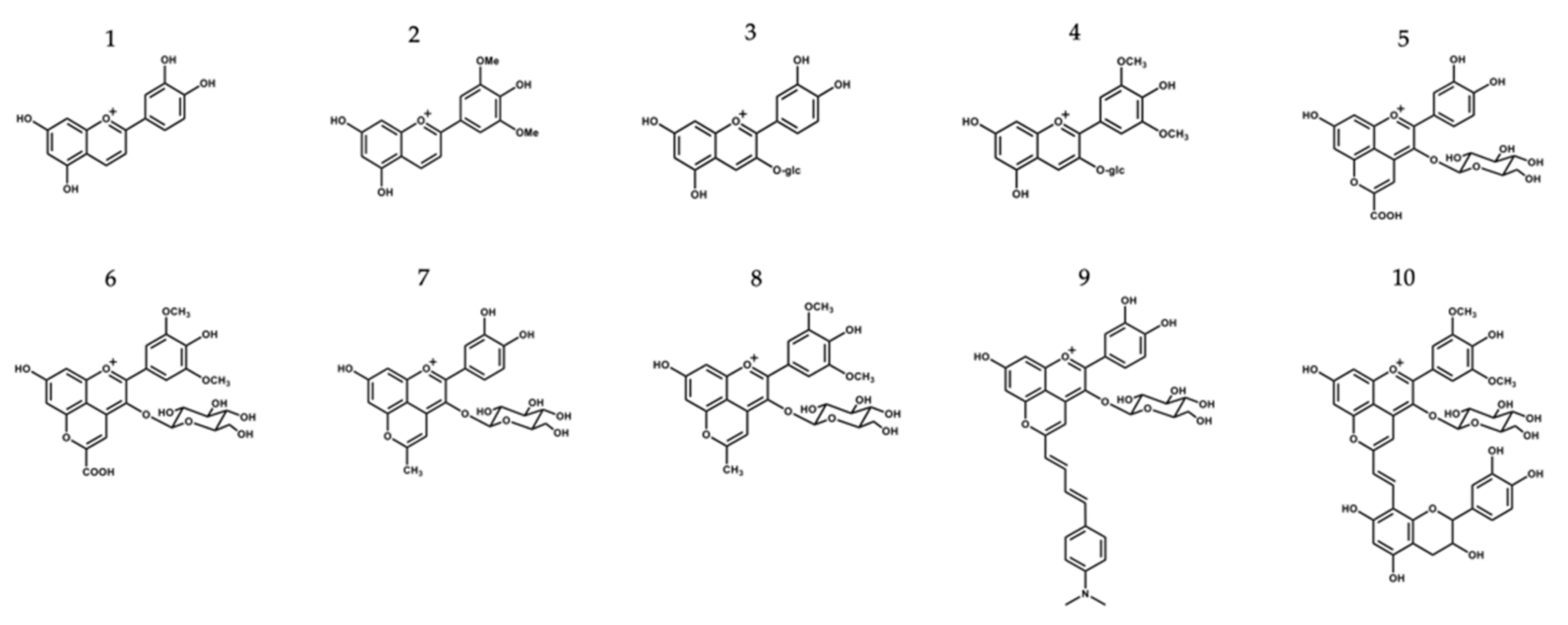
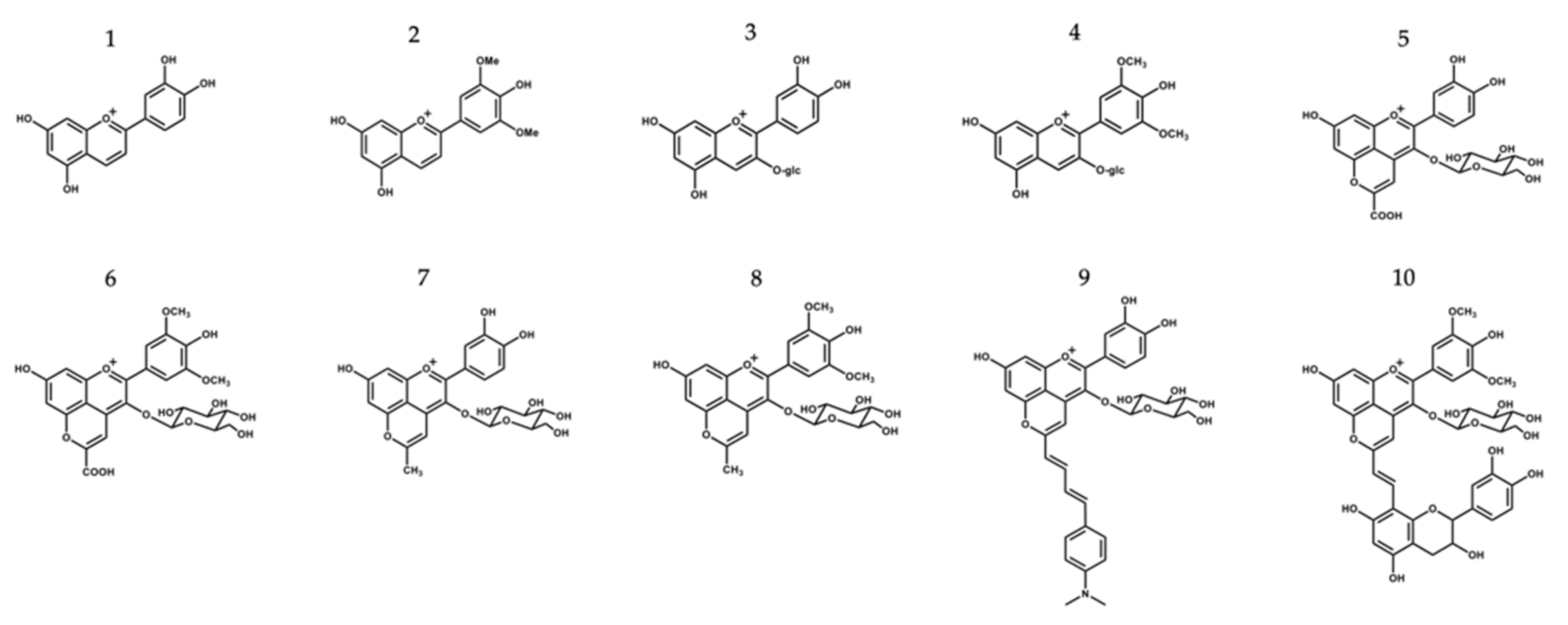
2.1. Isolation and Synthesis of the Different Compounds
2.2. Antimicrobial and Antibiofilm Assays
2.2.1. Bacterial Strains and Growth Conditions
2.2.2. Determination of Minimum Inhibitory and Minimum Bactericidal Concentrations
2.2.3. Biofilm Formation Inhibition Assay
2.3. Determination of Solar Protection Factor (SPF)
2.4. Cell Culture Assays
2.4.1. Cell Lines and Growth Conditions
2.4.2. Cytotoxicity Evaluation
2.4.3. ROS Formation Assay
2.4.4. Transport Assay
2.5. Enzymatic Inhibition Assays
2.5.1. Collagenase
2.5.2. Elastase
2.5.3. Hyaluronidase
2.6. Molecular Docking
2.7. Statistical Analysis
3. Results and Discussion
3.1. Antimicrobial and Antibiofilm Activities
3.1.1. Antibacterial Activity
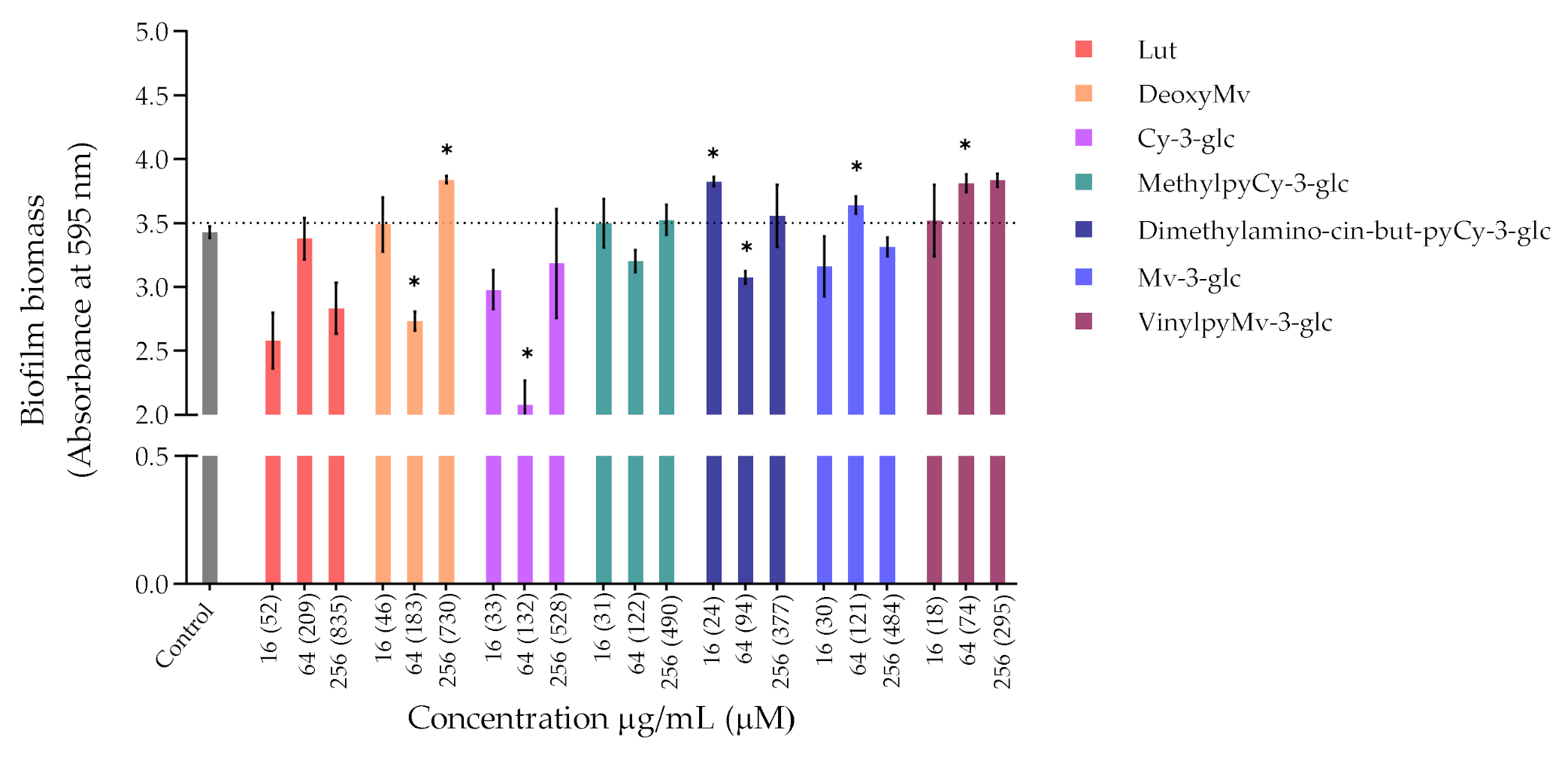
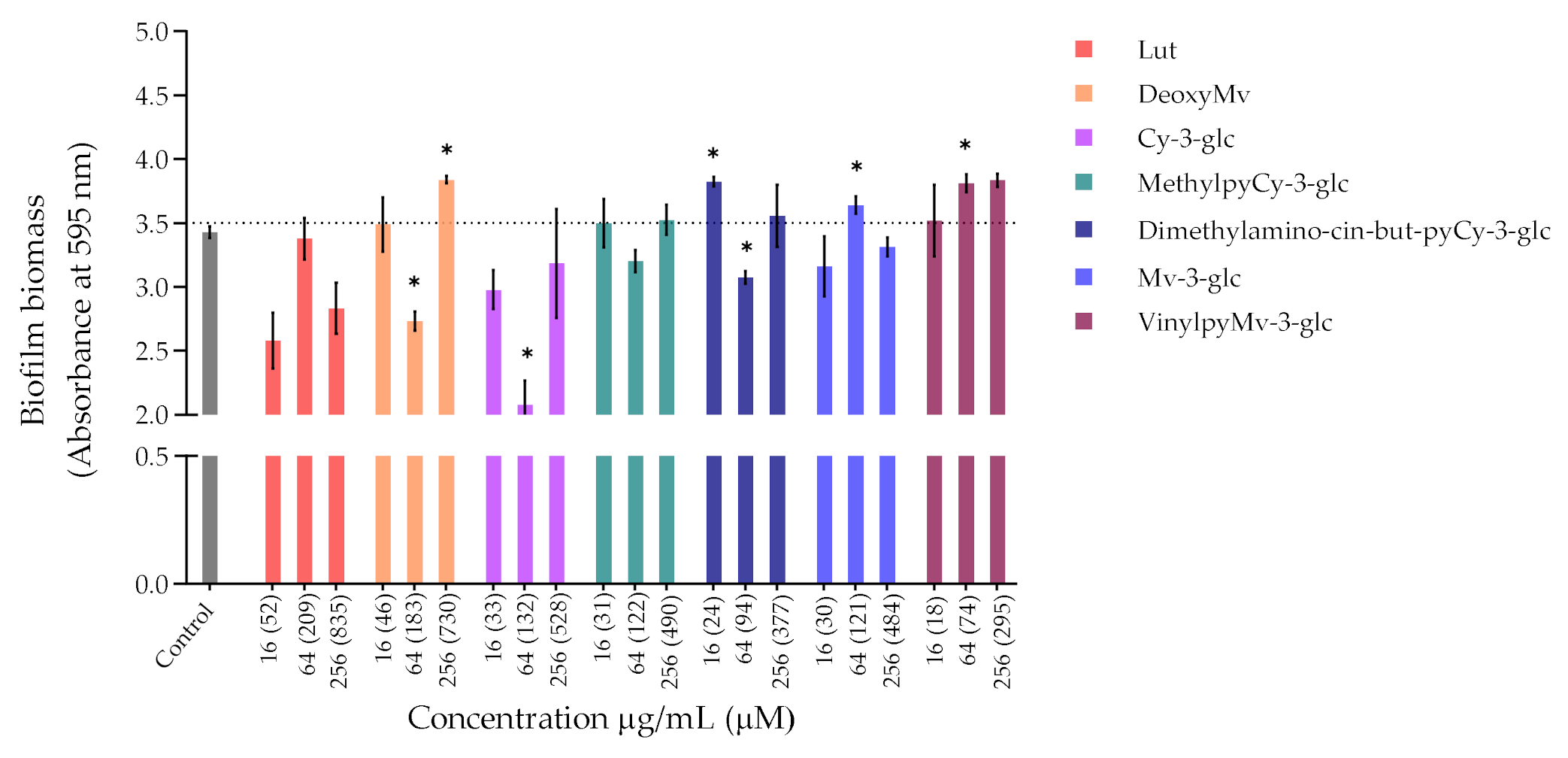
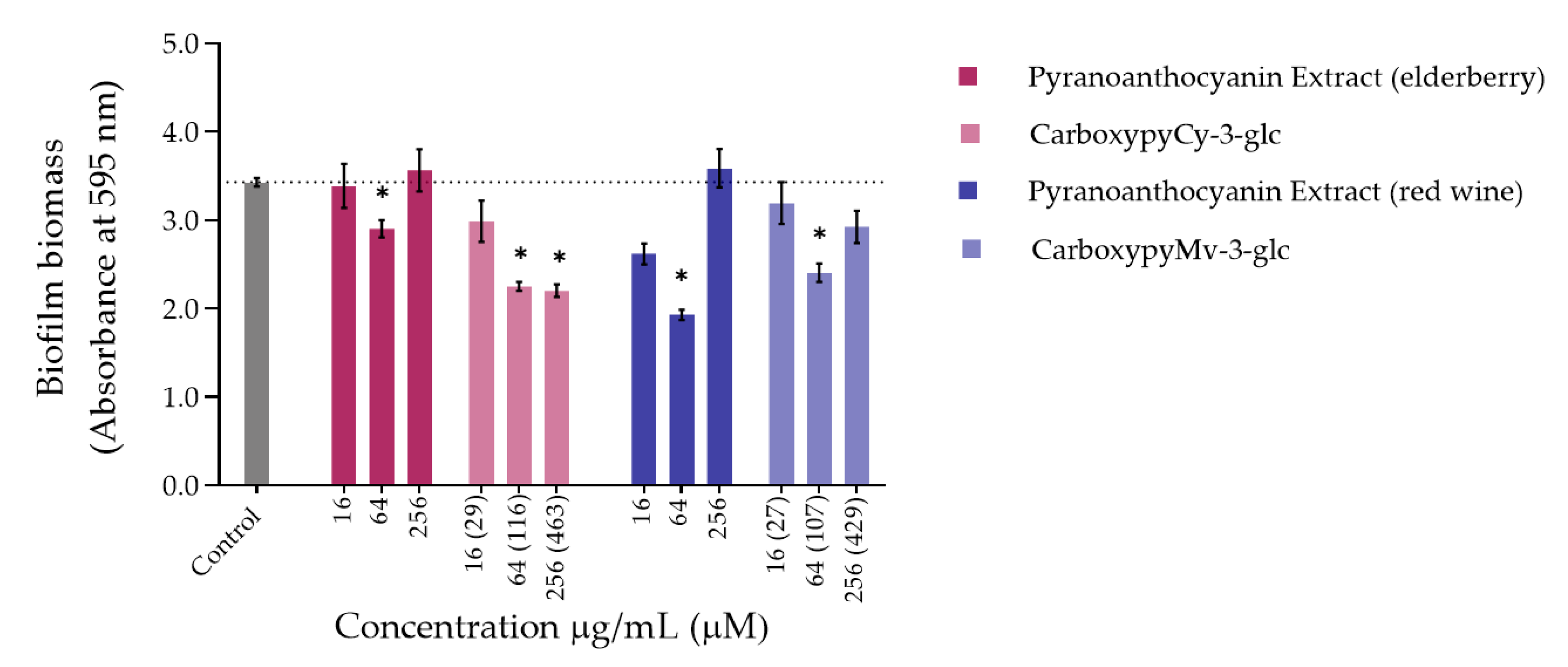
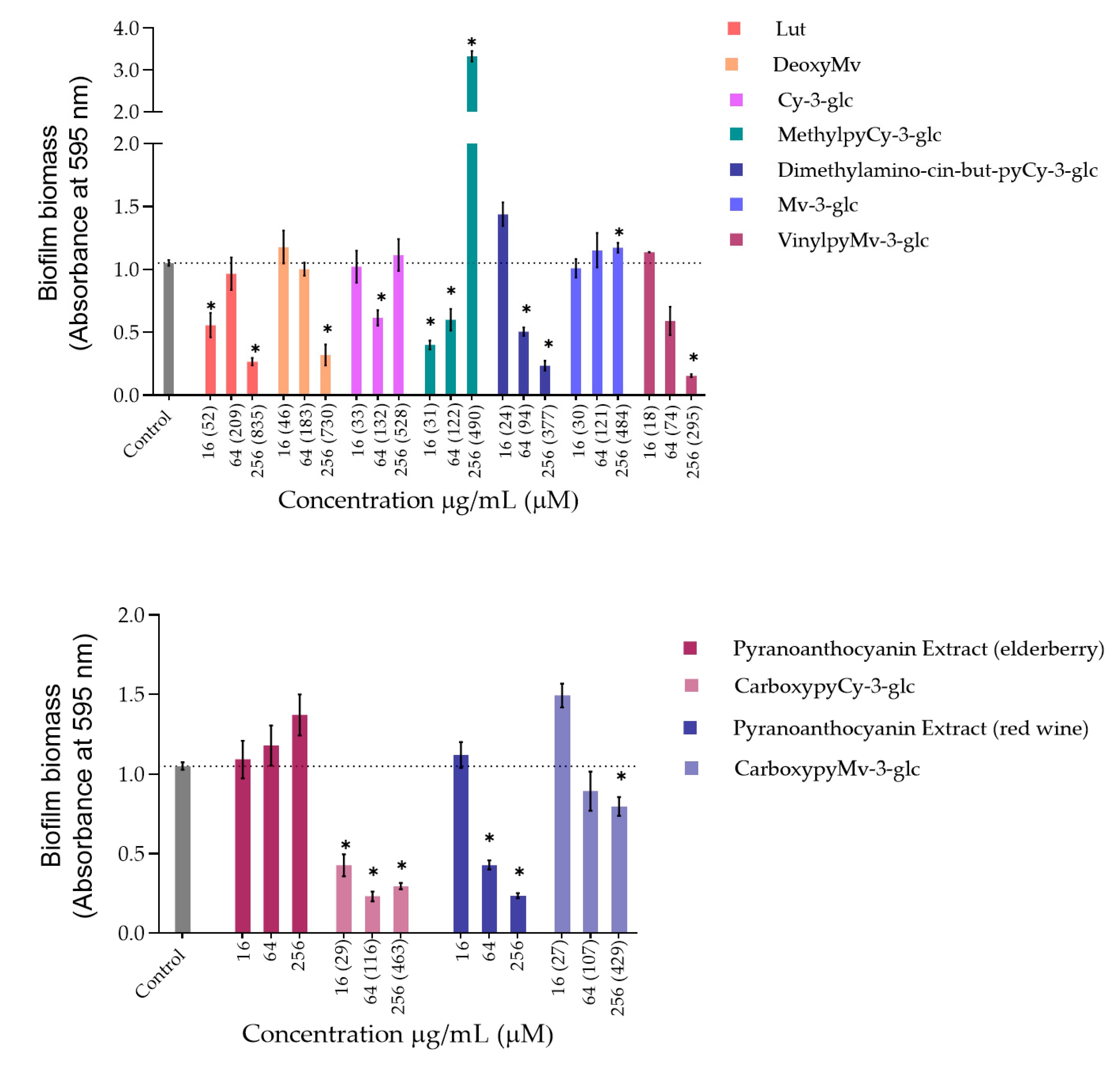
3.1.2. Inhibition of Biofilm Formation
3.2. Solar Protection Factor (SPF)
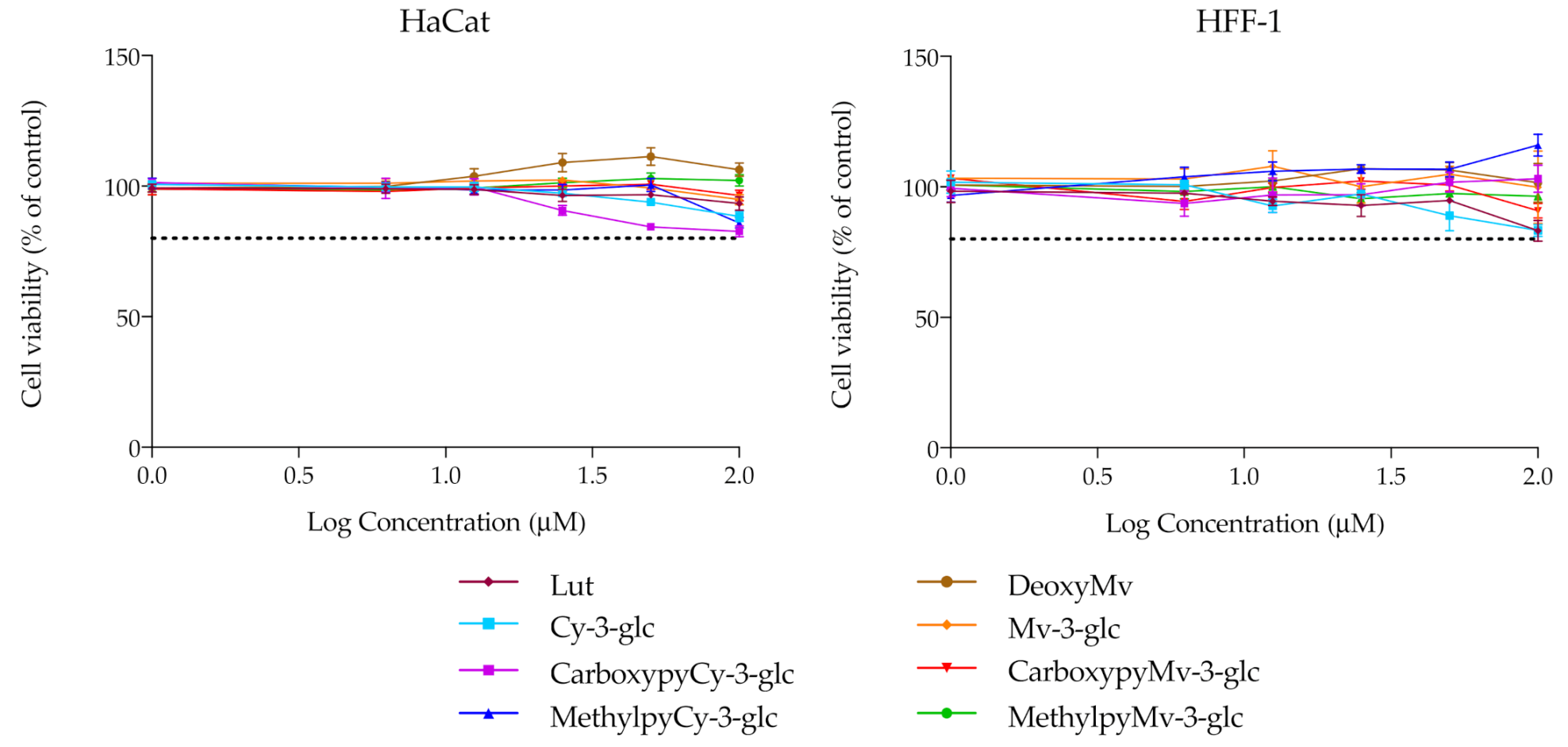
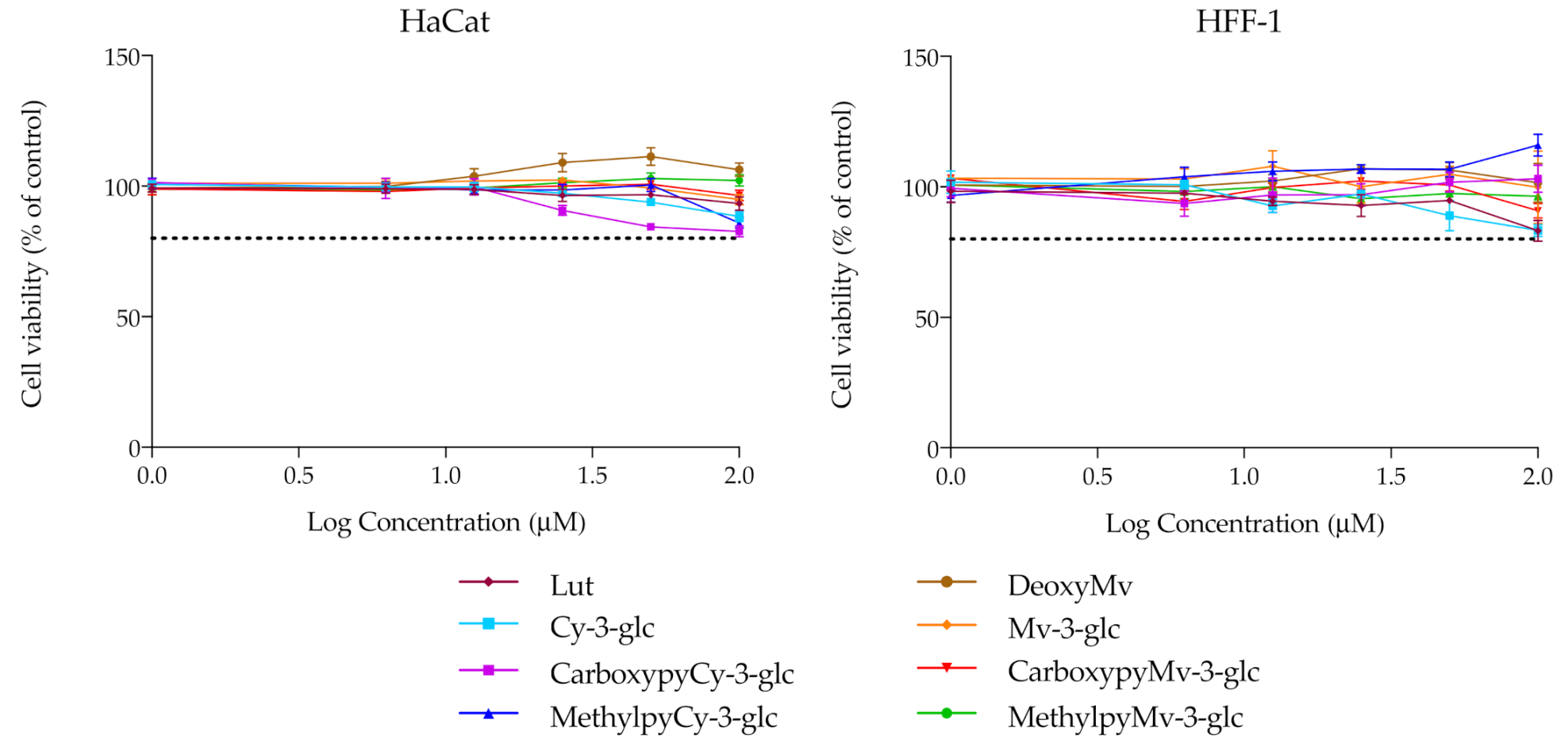
3.3. Cytotoxicity
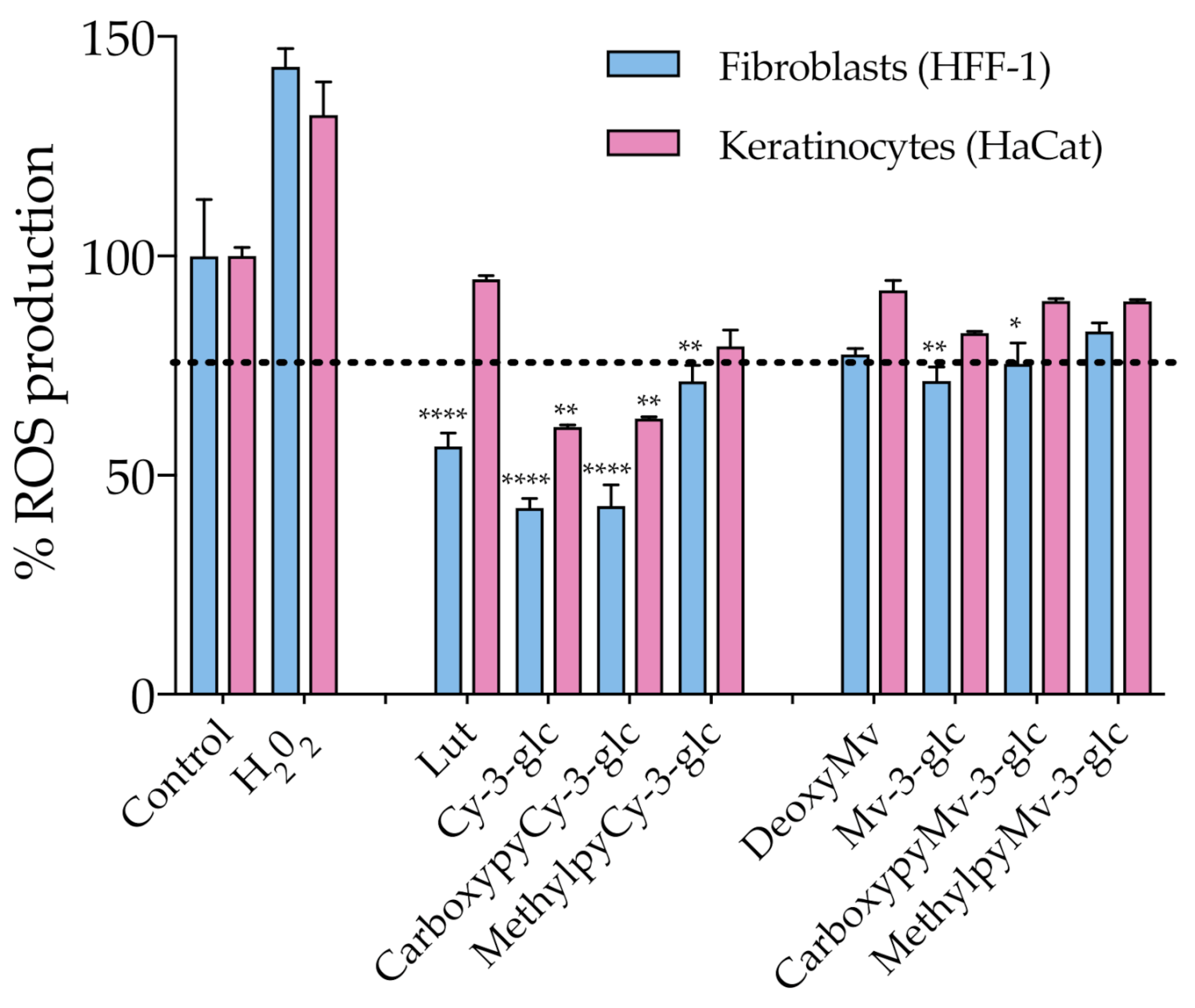
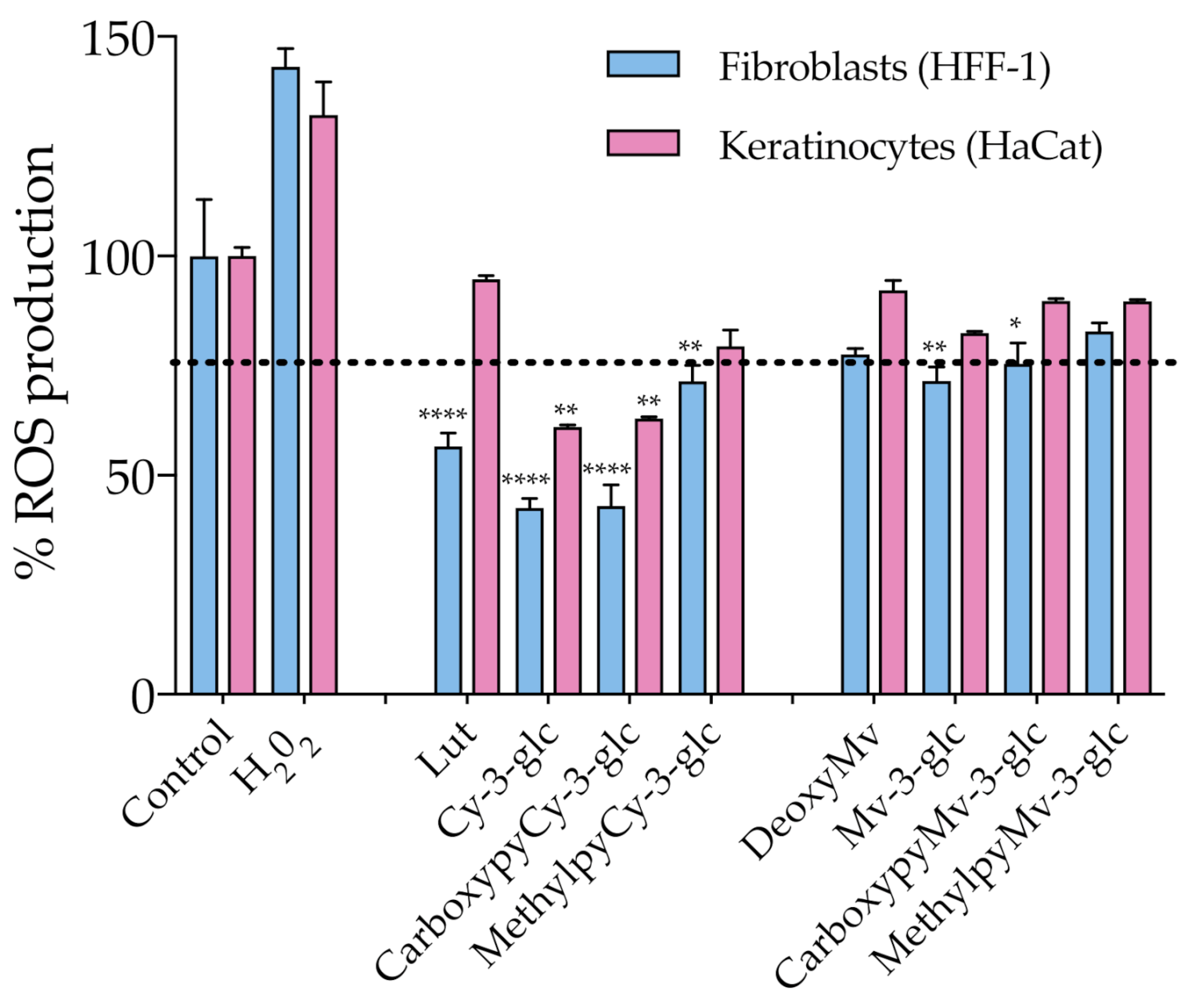
3.4. Antioxidant Affect
3.5. Inhibition of Skin Aging Related Enzymes Activity
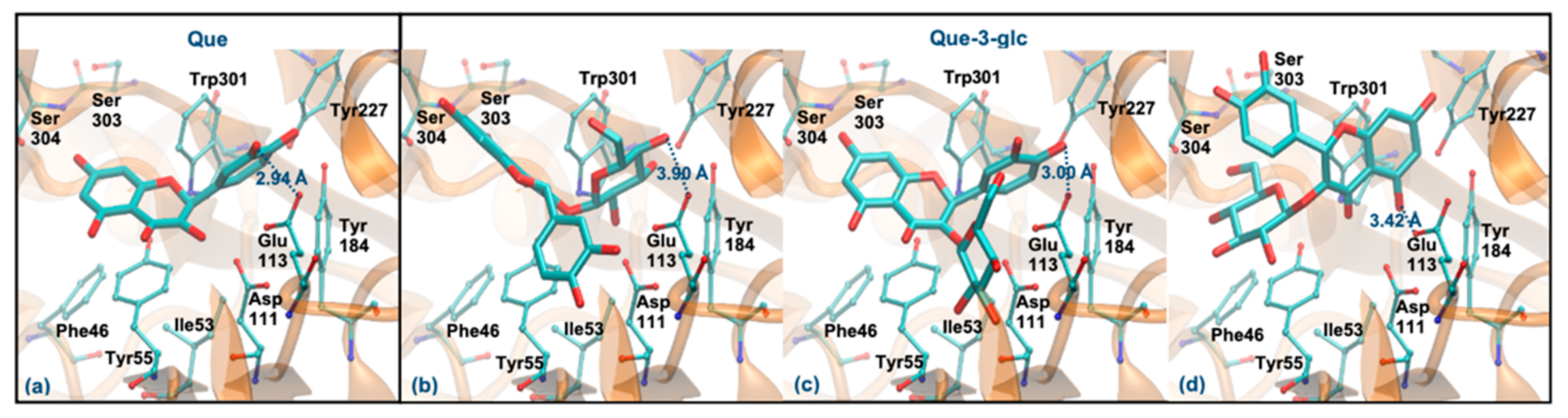
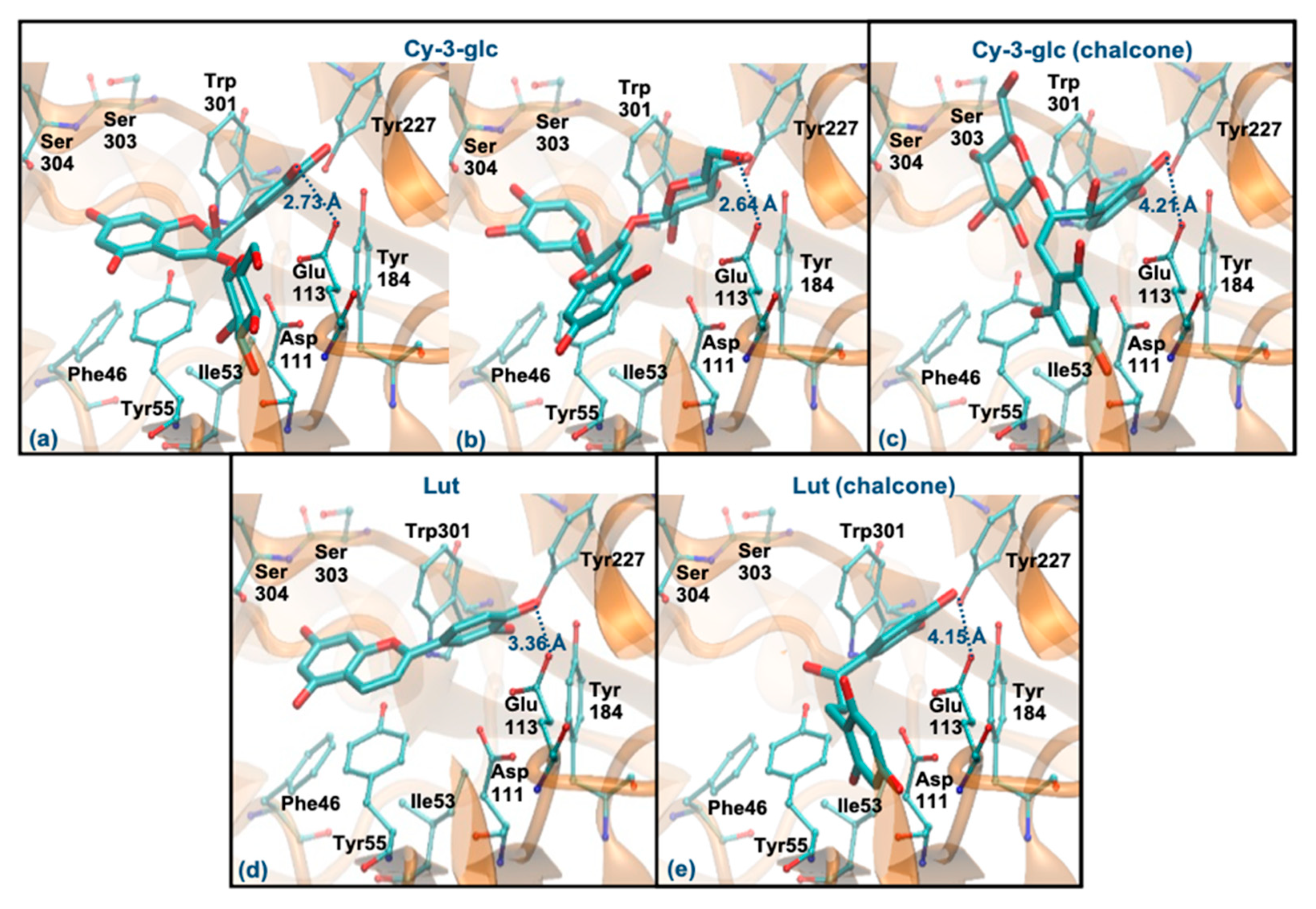
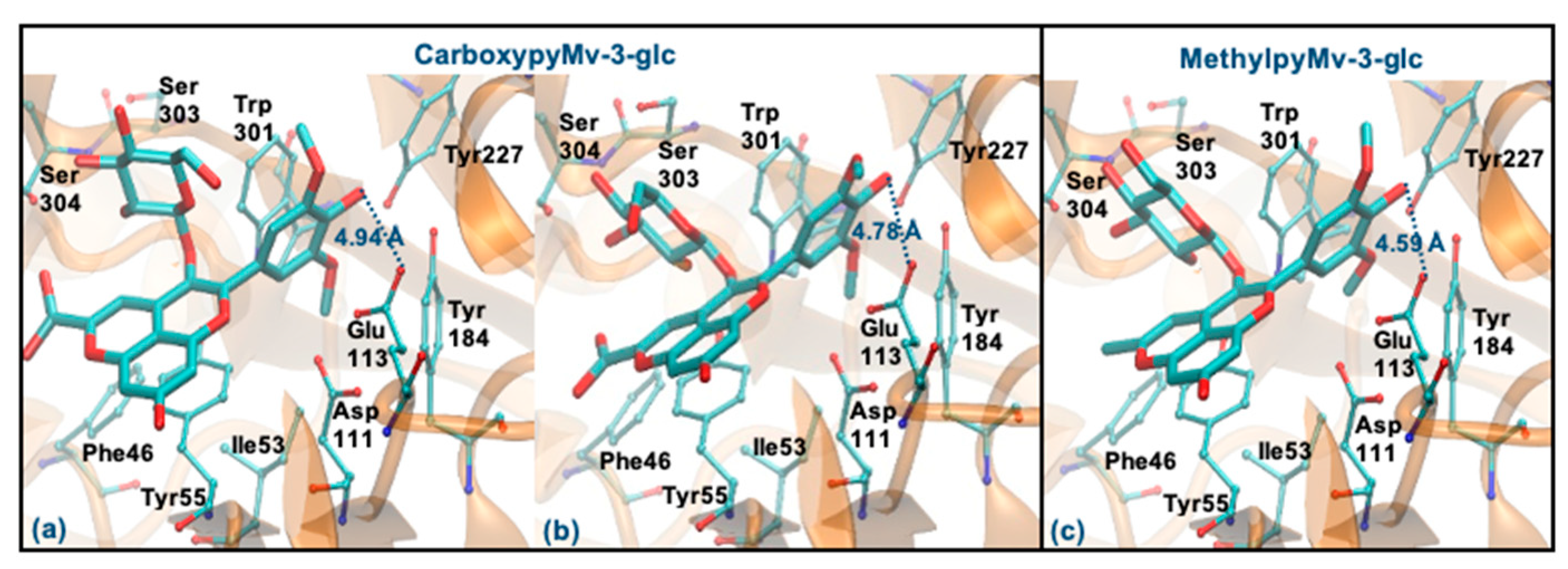
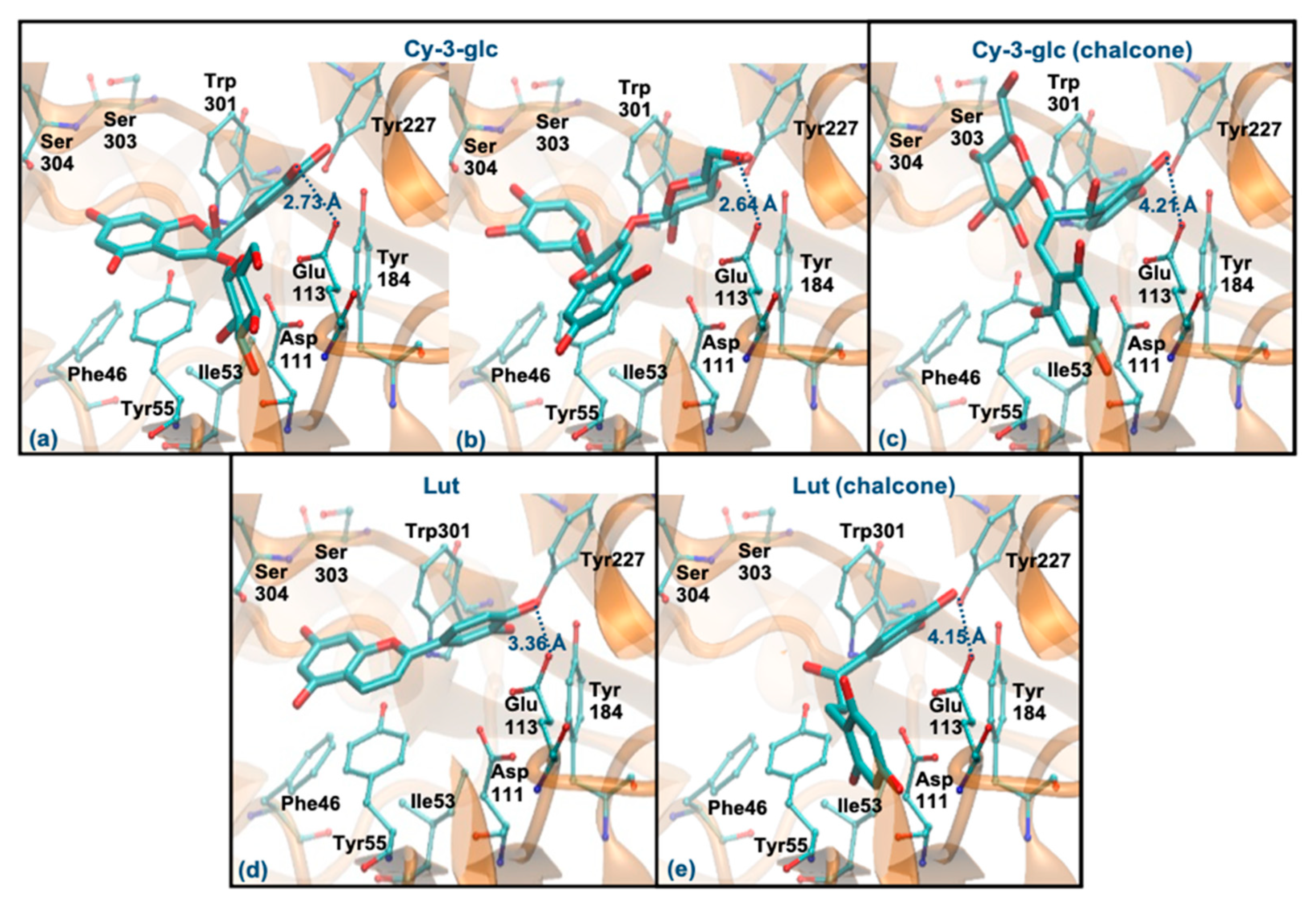
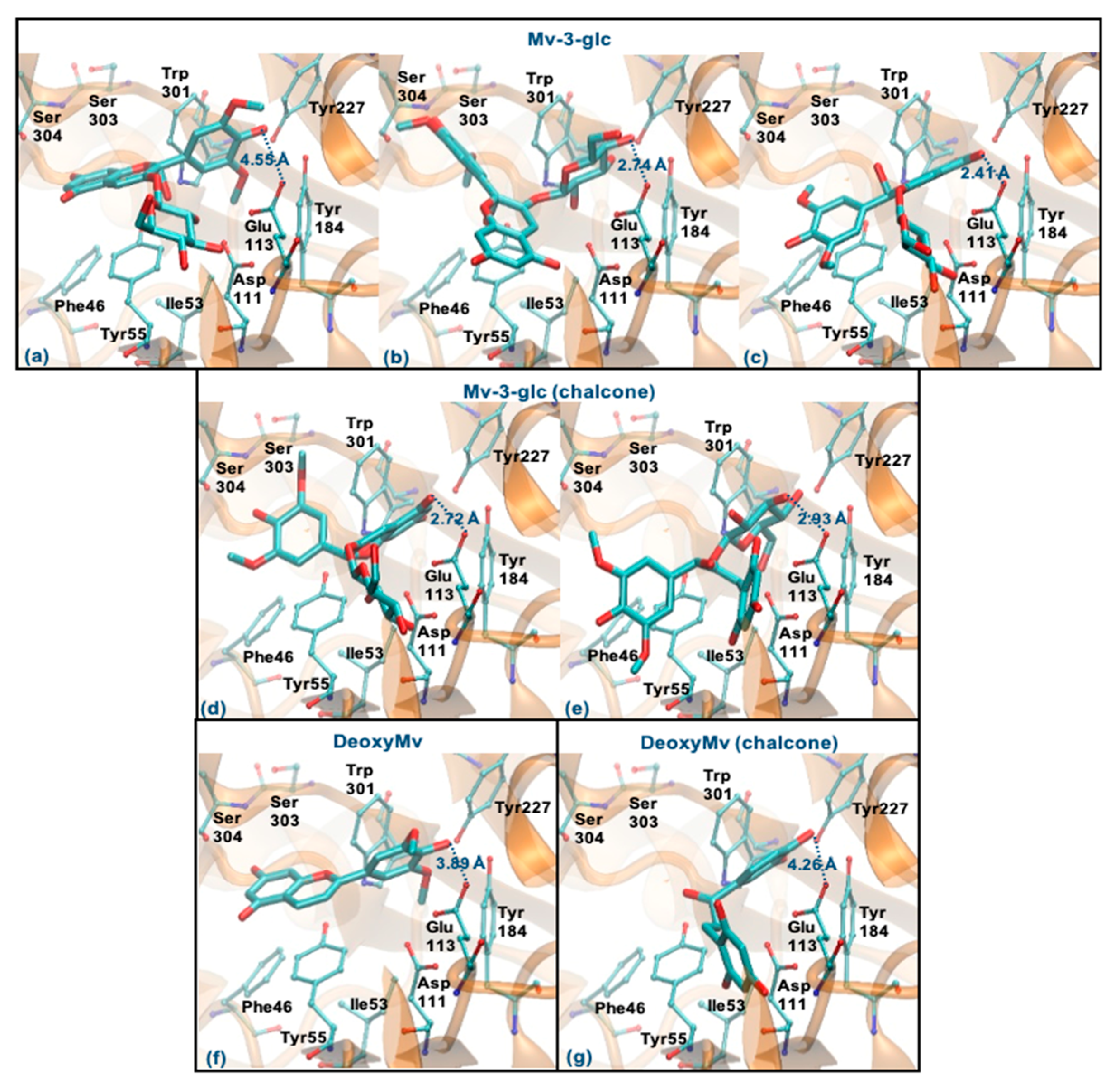
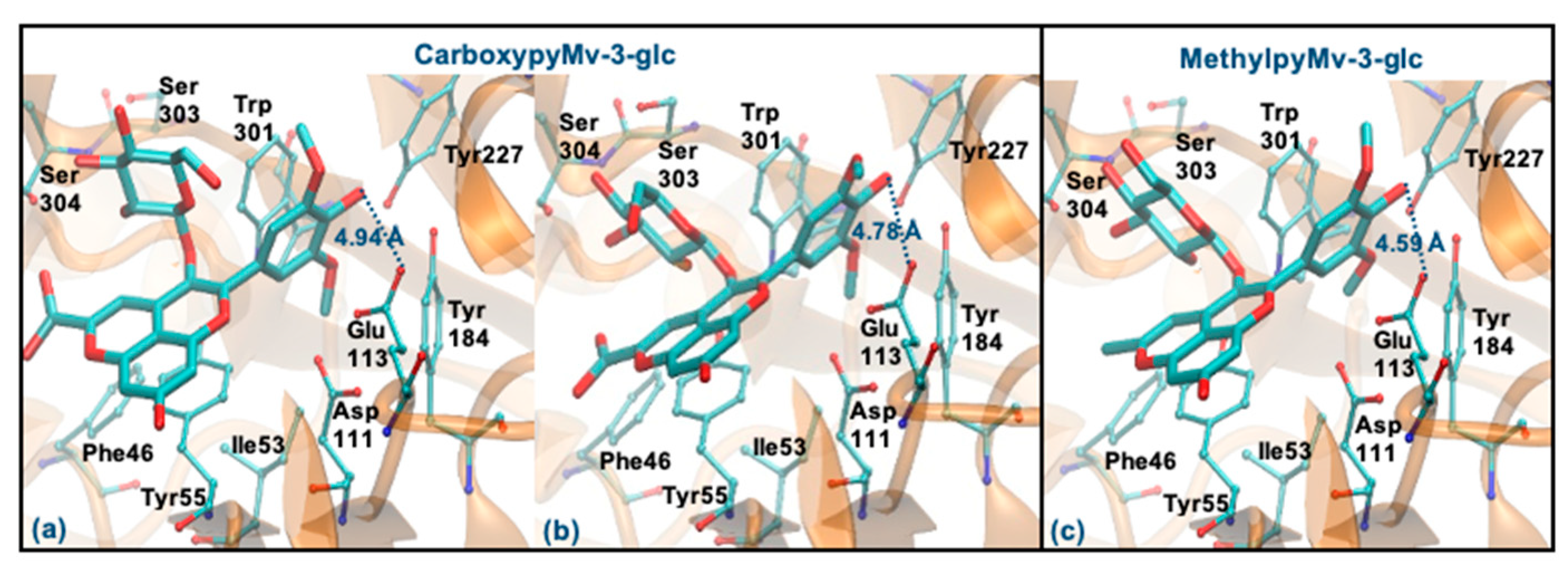
3.6. Molecular Docking Study with Hyaluronidase
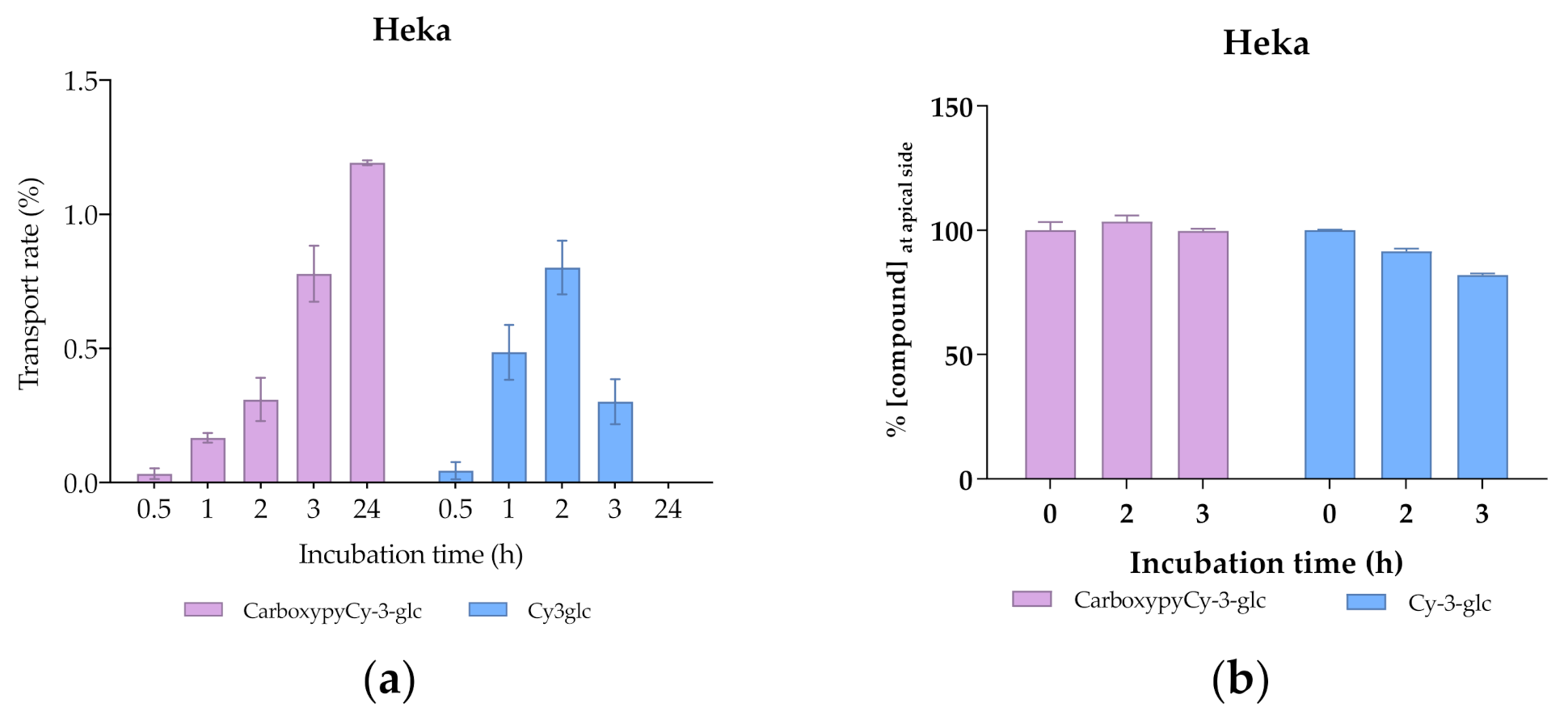
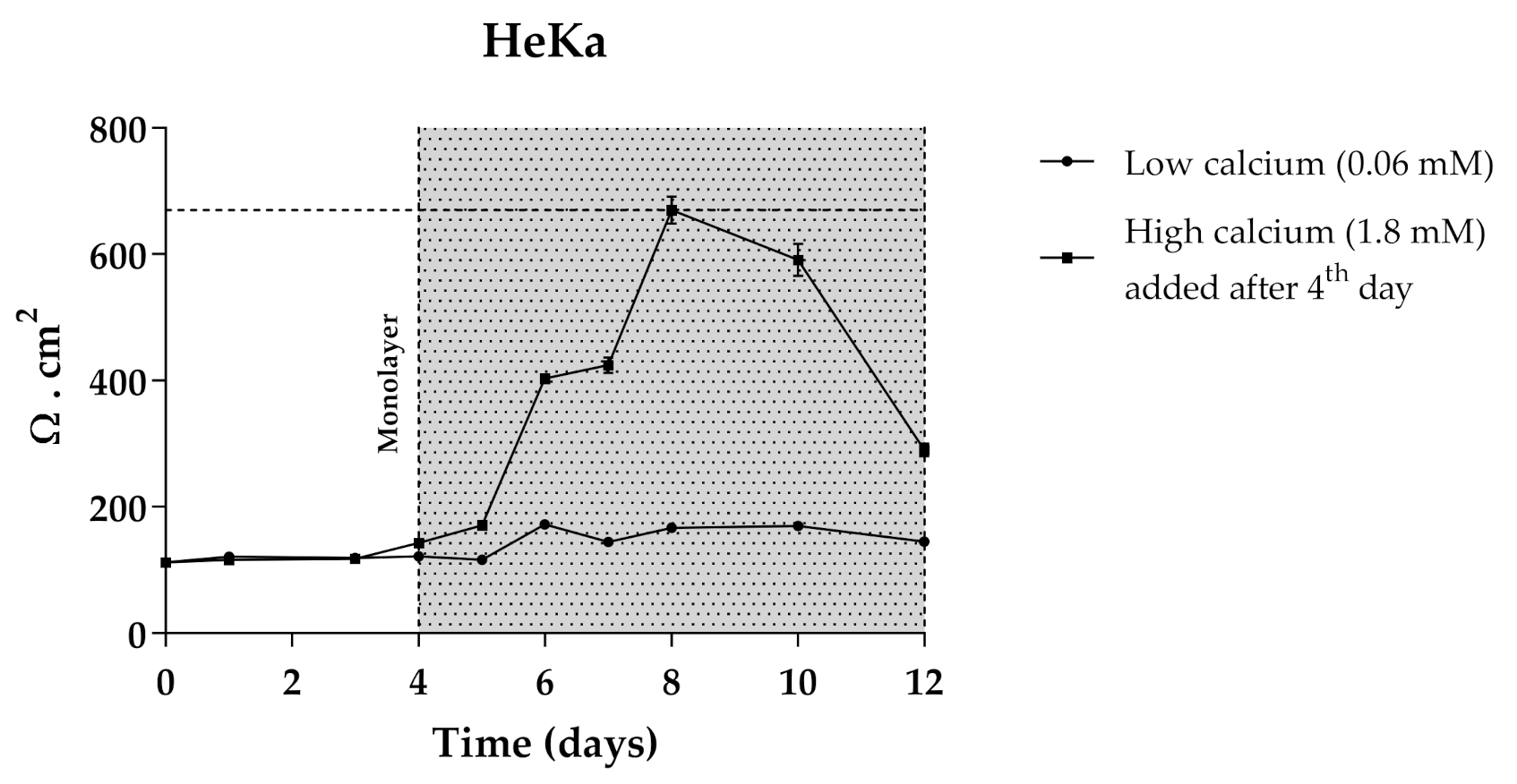
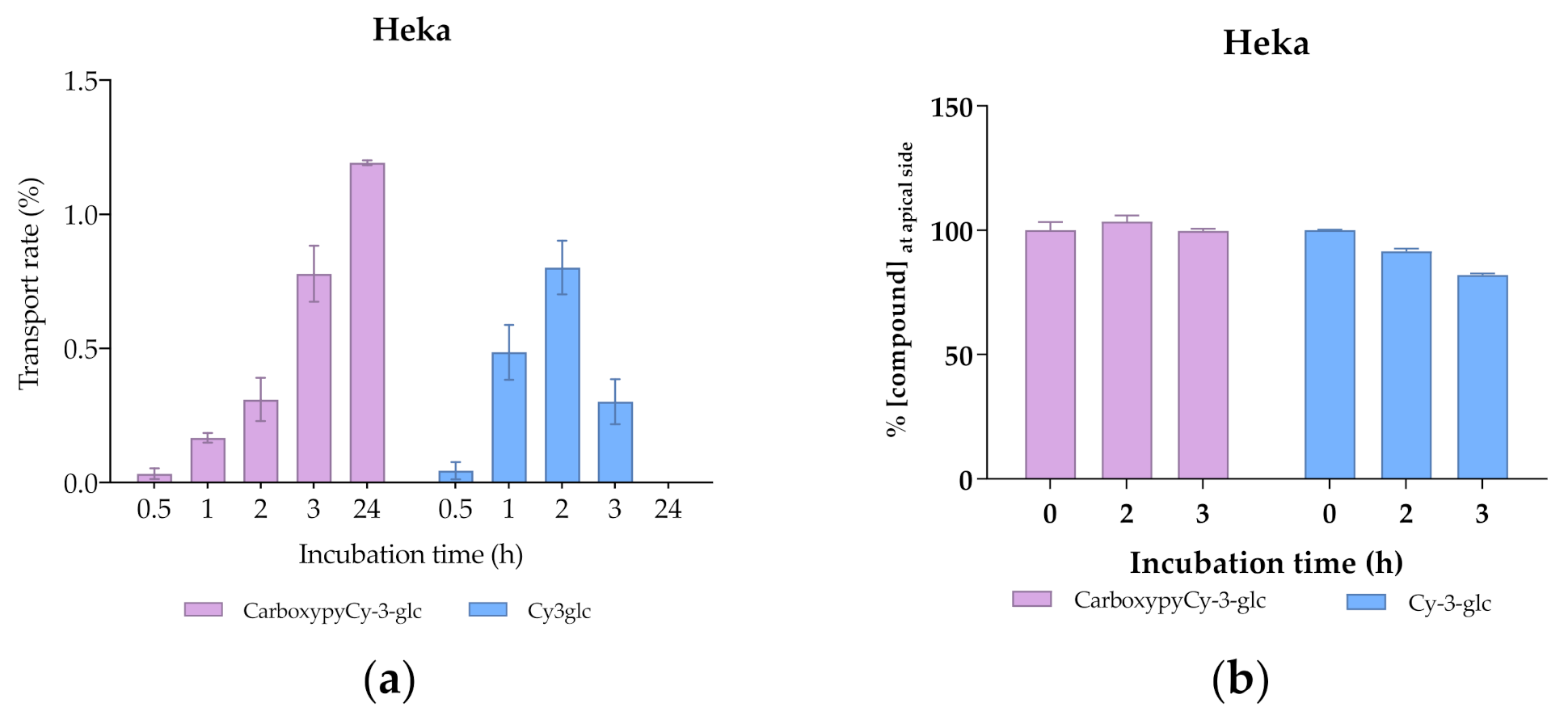
3.7. Transport Efficiency
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments

Conflicts of Interest
References
- Rittié, L.; Fisher, G.J. Natural and sun-induced aging of human skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orioli, D.; Dellambra, E. Epigenetic Regulation of Skin Cells in Natural Aging and Premature Aging Diseases. Cells 2018, 7, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.W.; Kwon, S.H.; Choi, J.Y.; Na, J.I.; Huh, C.H.; Choi, H.R.; Park, K.C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [Green Version]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef]
- Tu, Y.; Quan, T. Oxidative Stress and Human Skin Connective Tissue Aging. Cosmetics 2016, 3, 28. [Google Scholar] [CrossRef]
- Natarajan, V.T.; Ganju, P.; Ramkumar, A.; Grover, R.; Gokhale, R.S. Multifaceted pathways protect human skin from UV radiation. Nat. Chem. Biol. 2014, 10, 542–551. [Google Scholar] [CrossRef]
- Xu, Y.; Fisher, G.J. Ultraviolet (UV) light irradiation induced signal transduction in skin photoaging. J. Dermatol. Sci. Suppl. 2005, 1, S1–S8. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Hernandez, D.F.; Cervantes, E.L.; Luna-Vital, D.A.; Mojica, L. Food-derived bioactive compounds with anti-aging potential for nutricosmetic and cosmeceutical products. Crit. Rev. Food Sci. Nutr. 2020, 1–16. [Google Scholar] [CrossRef]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Li, K.; Zhang, M.; Chen, H.; Peng, J.; Jiang, F.; Shi, X.; Bai, Y.; Jian, M.; Jia, Y. Anthocyanins from black peanut skin protect against UV-B induced keratinocyte cell and skin oxidative damage through activating Nrf 2 signaling. Food Funct. 2019, 10, 6815–6828. [Google Scholar] [CrossRef]
- Bucci, P.; Prieto, M.J.; Milla, L.; Calienni, M.N.; Martinez, L.; Rivarola, V.; Alonso, S.; Montanari, J. Skin penetration and UV-damage prevention by nanoberries. J. Cosmet. Dermatol. 2018, 17, 889–899. [Google Scholar] [CrossRef]
- Peng, Z.; Hu, X.; Li, X.; Jiang, X.; Deng, L.; Hu, Y.; Bai, W. Protective effects of cyanidin-3-O-glucoside on UVB-induced chronic skin photodamage in mice via alleviating oxidative damage and anti-inflammation. Food Front. 2020, 1, 213–223. [Google Scholar] [CrossRef]
- Pal, H.C.; Chamcheu, J.C.; Adhami, V.M.; Wood, G.S.; Elmets, C.A.; Mukhtar, H.; Afaq, F. Topical application of delphinidin reduces psoriasiform lesions in the flaky skin mouse model by inducing epidermal differentiation and inhibiting inflammation. Br. J. Dermatol. 2015, 172, 354–364. [Google Scholar] [CrossRef]
- Oliveira, J.; Mateus, N.; de Freitas, V. Previous and recent advances in pyranoanthocyanins equilibria in aqueous solution. Dyes Pigment. 2014, 100, 190–200. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. 3-Deoxyanthocyanidin Colorant: Nature, Health, Synthesis, and Food Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1533–1549. [Google Scholar] [CrossRef] [Green Version]
- Al Bittar, S.; Mora, N.; Loonis, M.; Dangles, O. A simple synthesis of 3-deoxyanthocyanidins and their O-glucosides. Tetrahedron 2016, 72, 4294–4302. [Google Scholar] [CrossRef]
- Oliveira, J.; Fernandes, V.; Miranda, C.; Santos-Buelga, C.; Silva, A.; de Freitas, V.; Mateus, N. Color Properties of Four Cyanidin−Pyruvic Acid Adducts. J. Agric. Food Chem. 2006, 54, 6894–6903. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Unusual anthocyanin reaction with acetone leading to pyranoanthocyanin formation. Tetrahedron Lett. 2001, 42, 1371–1373. [Google Scholar] [CrossRef]
- Oliveira, J.A.P.; Fernandes, A.; Mateus, N.; de Freitas, V. Synthesis and structural characterization of amino-based pyranoanthocyanins with extended electronic delocalization. Synlett 2016, 27, 2459–2462. [Google Scholar] [CrossRef]
- Mateus, N.; Oliveira, J.; Santos-Buelga, C.; Silva, A.M.S.; de Freitas, V. NMR structure characterization of a new vinylpyranoanthocyanin–catechin pigment (a portisin). Tetrahedron Lett. 2004, 45, 3455–3457. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution of Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard, 10th ed.; CLSI: Wayne, PA, USA, 2015; Volume 35, pp. M-07–A-10. [Google Scholar]
- Bessa, L.J.; Eaton, P.; Dematei, A.; Placido, A.; Vale, N.; Gomes, P.; Delerue-Matos, C.; Sa Leite, J.R.; Gameiro, P. Synergistic and antibiofilm properties of ocellatin peptides against multidrug-resistant Pseudomonas aeruginosa. Future Microbiol. 2018, 13, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Marković-Housley, Z.; Miglierini, G.; Soldatova, L.; Rizkallah, P.J.; Müller, U.; Schirmer, T. Crystal Structure of Hyaluronidase, a Major Allergen of Bee Venom. Structure 2000, 8, 1025–1035. [Google Scholar] [CrossRef]
- Mohamed, E.M.; Hetta, M.H.; Rateb, M.E.; Selim, M.A.; AboulMagd, A.M.; Badria, F.A.; Abdelmohsen, U.R.; Alhadrami, H.A.; Hassan, H.M. Bioassay-Guided Isolation, Metabolic Profiling, and Docking Studies of Hyaluronidase Inhibitors from Ravenala madagascariensis. Molecules 2020, 25, 1714. [Google Scholar] [CrossRef] [Green Version]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An Advanced Semantic Chemical Editor, Visualization, and Analysis Platform. J. Cheminform. 2012, 4, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An Automated Pipeline for the Setup of Poisson-Boltzmann Electrostatics Calculations. Nucleic Acids Res. 2004, 32, 665–667. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the Natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cady, N.C.; McKean, K.A.; Behnke, J.; Kubec, R.; Mosier, A.P.; Kasper, S.H.; Burz, D.S.; Musah, R.A. Inhibition of biofilm formation, quorum sensing and infection in Pseudomonas aeruginosa by natural products-inspired organosulfur compounds. PLoS ONE 2012, 7, e38492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slobodníková, L.; Fialová, S.; Rendeková, K.; Kováč, J.; Mučaji, P. Antibiofilm Activity of Plant Polyphenols. Molecules 2016, 21, 1717. [Google Scholar] [CrossRef] [PubMed]
- Sitarek, P.; Merecz-Sadowska, A.; Kowalczyk, T.; Wieczfinska, J.; Zajdel, R.; Śliwiński, T. Potential Synergistic Action of Bioactive Compounds from Plant Extracts against Skin Infecting Microorganisms. Int. J. Mol. Sci. 2020, 21, 5105. [Google Scholar] [CrossRef]
- Cefali, L.C.; Franco, J.G.; Nicolini, G.F.; Ataide, J.A.; Mazzola, P.G. In vitro antioxidant activity and solar protection factor of blackberry and raspberry extracts in topical formulation. J. Cosmet. Dermatol. 2019, 18, 539–544. [Google Scholar] [CrossRef]
- Chan, C.F.; Lien, C.Y.; Lai, Y.C.; Huang, C.L.; Liao, W.C. Influence of purple sweet potato extracts on the UV absorption properties of a cosmetic cream. J. Cosmet. Sci. 2010, 61, 333–341. [Google Scholar]
- Suh, S.; Pham, C.; Smith, J.; Mesinkovska, N.A. The banned sunscreen ingredients and their impact on human health: A systematic review. Int. J. Dermatol. 2020, 59, 1033–1042. [Google Scholar] [CrossRef]
- Sabzevari, N.; Qiblawi, S.; Norton, S.A.; Fivenson, D. Sunscreens: UV filters to protect us: Part 1: Changing regulations and choices for optimal sun protection. Int. J. Women’s Dermatol. 2021, 7, 28–44. [Google Scholar] [CrossRef]
- de Oliveira, C.A.; Dario, M.F. Bioactive Cosmetics. In Handbook of Ecomaterials; Martínez, L.M.T., Kharissova, O.V., Kharisov, B.I., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–23. [Google Scholar] [CrossRef]
- Hubner, A.; Sobreira, F.; Vetore Neto, A.; Pinto, C.; Dario, M.F.; Díaz, I.E.C.; Lourenço, F.R.; Rosado, C.; Baby, A.R.; Bacchi, E.M. The Synergistic Behavior of Antioxidant Phenolic Compounds Obtained from Winemaking Waste’s Valorization, Increased the Efficacy of a Sunscreen System. Antioxidants 2019, 8, 530. [Google Scholar] [CrossRef] [Green Version]
- Évora, A.; de Freitas, V.; Mateus, N.; Fernandes, I. Effect of anthocyanins from red wine and blackberry on the integrity of a keratinocyte model using ECIS. Food Funct. 2017. submitted. [Google Scholar] [CrossRef]
- Lewis, D.A.; Krbanjevic, A.; Travers, J.B.; Spandau, D.F. Aging-Associated Nonmelanoma Skin Cancer: A Role for the Dermis. In Textbook of Aging Skin; Farage, M.A., Miller, K.W., Maibach, H.I., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 913–930. [Google Scholar] [CrossRef]
- Fallah, A.A.; Sarmast, E.; Jafari, T. Effect of dietary anthocyanins on biomarkers of oxidative stress and antioxidative capacity: A systematic review and meta-analysis of randomized controlled trials. J. Funct. Foods 2020, 68, 103912. [Google Scholar] [CrossRef]
- Xiang, Y.; Lai, F.; He, G.; Li, Y.; Yang, L.; Shen, W.; Huo, H.; Zhu, J.; Dai, H.; Zhang, Y. Alleviation of Rosup-induced oxidative stress in porcine granulosa cells by anthocyanins from red-fleshed apples. PLoS ONE 2017, 12, e0184033. [Google Scholar] [CrossRef]
- Arming, S.; Strobl, B.; Wechselberger, C.; Kreil, G. In vitro Mutagenesis of PH-20 Hyaluronidase from Human Sperm. Eur. J. Biochem. 1997, 247, 810–814. [Google Scholar] [CrossRef] [Green Version]
- Kuppusamy, U.R.; Khoo, H.E.; Das, N.P. Structure-Activity Studies of Flavonoids as Inhibitors of Hyaluronidase. Biochem. Pharmacol. 1990, 40, 397–401. [Google Scholar] [CrossRef]
- Fukunishi, Y.; Yamashita, Y.; Mashimo, T.; Nakamura, H. Prediction of Protein−compound Binding Energies from Known Activity Data: Docking-score-based Method and its Applications. Mol. Inform. 2018, 37, 1–11. [Google Scholar] [CrossRef]
- Ahmed, M.; Aldesouki, H.; Badria, F. Effect of Phenolic Compounds from the Rind of Punica granatum on the Activity of Three Metabolism-related Enzymes. Biotechnol. Appl. Biochem. 2019, 1, 1–13. [Google Scholar] [CrossRef]
- Oliveira, H.; Correia, P.; Bessa, L.J.; Guimarães, M.; Gameiro, P.; Freitas, V.D.; Mateus, N.; Cruz, L.; Fernandes, I. Cyanidin-3-Glucoside Lipophilic Conjugates for Topical Application: Tuning the Antimicrobial Activities with Fatty Acid Chain Length. Processes 2021, 9, 340. [Google Scholar] [CrossRef]
- Three-Dimensional In Vitro Skin and Skin Cancer Models Based on Human Fibroblast-Derived Matrix. Tissue Eng. Part C Methods 2015, 21, 958–970. [CrossRef]
- Wan, H.; Yuan, M.; Simpson, C.; Allen, K.; Gavins, F.N.E.; Ikram, M.S.; Basu, S.; Baksh, N.; O’Toole, E.A.; Hart, I.R. Stem/Progenitor Cell-Like Properties of Desmoglein 3dim Cells in Primary and Immortalized Keratinocyte Lines. Stem Cells 2007, 25, 1286–1297. [Google Scholar] [CrossRef]
- Zhang, J.; Giampieri, F.; Afrin, S.; Battino, M.; Zheng, X.; Reboredo-Rodriguez, P. Structure-stability relationship of anthocyanins under cell culture condition. Int. J. Food Sci. Nutr. 2019, 70, 285–293. [Google Scholar] [CrossRef]
- Shen, S.; Sampson, S.R.; Tennenbaum, T.; Wertheimer, E. Characterization of Glucose Transport System in Keratinocytes: Insulin and IGF-1 Differentially Affect Specific Transporters. J. Investig. Dermatol. 2000, 115, 949–954. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.; Roma-Rodrigues, C.; Santos, A.; Veigas, B.; Brás, N.; Faria, A.; Calhau, C.; de Freitas, V.; Baptista, P.V.; Mateus, N.; et al. GLUT1 and GLUT3 involvement in anthocyanin gastric transport- Nanobased targeted approach. Sci. Rep. 2019, 9, 789. [Google Scholar] [CrossRef]
- Westfall, A.; Sigurdson, G.T.; Rodriguez-Saona, L.E.; Giusti, M.M. Ex Vivo and In Vivo Assessment of the Penetration of Topically Applied Anthocyanins Utilizing ATR-FTIR/PLS Regression Models and HPLC-PDA-MS. Antioxidants 2020, 9, 486. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| λ (nm) | EE × I (Normalized) |
|---|---|
| 290 | 0.0150 |
| 295 | 0.0817 |
| 300 | 0.2874 |
| 305 | 0.3278 |
| 310 | 0.1864 |
| 315 | 0.0839 |
| 320 | 0.0180 |
| Total | 1 |
| Extract/Compound | P. aeruginosa ATCC 27853 | S. aureus ATCC 29213 | M. luteus ATCC 4698 | S. epidermidis ATCC 14990 | S. pyogenes ATCC 19615 |
|---|---|---|---|---|---|
| MIC—µg/mL (mM) | |||||
| Lut | >512 (>1.67) | >512 (>1.67) | 128 (0.42) * | 128 (0.42) * | >512 (>1.67) |
| DeoxyMv | >512 (>1.46) | >512 (>1.46) | >512 (>1.46) | >512 (>1.46) | >512 (>1.46) |
| Cy-3-glc | >512 (>1.06) | >512 (>1.06) | >512 (>1.06) | 512 (1.06) * | >512 (>1.06) |
| CarboxypyCy-3-glc | >512 (>0.93) | >512 (>0.93) | >512 (>0.93) | >512 (>0.93) | >512 (>0.93) |
| Elderberry carboxypyranoanthocyanin extract (Elderberry extract) | >512 | >512 | 512 * | >512 | >512 |
| MethylpyCy-3-glc | >512 (>0.98) | >512 (>0.98) | >512 (>0.98) | >512 (>0.98) | >512 (>0.98) |
| Dimethylamino-cin-but-pyCy-3-glc | >512 (>0.75) | >512 (>0.75) | 64 (0.09) * | >512 (>0.75) | 512 (0.75) * |
| Mv-3-glc | >512 (>0.97) | >512 (>0.97) | >512 (>0.97) | >512 (>0.97) | >512 (>0.97) |
| CarboxypyMv-3-glc | >512 (>0.86) | >512 (>0.86) | >512 (>0.86) | >512 (>0.86) | >512 (>0.86) |
| Red wine carboxypyranoanthocyanin extract (red wine extract) | >512 | >512 | 512 * | >512 | >512 |
| VinylpyMv-3-glc-catechin | >512 (>0.59) | >512 (>0.59) | >512 (>0.59) | >512 (>0.59) | >512 (>0.59) |
| Compound (0.2 mg/mL) | SPF | |
|---|---|---|
| - | + 0.1 mg Oxybenzone/mL | |
| Lut | 29.82 ± 0.02 | 71.89 ± 0.040 |
| DeoxyMv | 17.28 ± 0.03 | 55.25 ± 0.03 |
| Cy-3-glc | 22.38 ± 0.01 | 63.01 ± 0.02 |
| Blackberry anthocyanin extract | 20.10 ± 0.06 | 66.98 ± 0.03 |
| CarboxypyCy-3-glc | 20.71 ± 0.18 | n.d. |
| Elderberry carboxypyranoanthocyanin extract | 16.03 ± 0.01 | 64.36 ± 0.03 |
| MethylpyCy-3-glc | 18.53 ± 0.15 | 68.21 ± 0.08 |
| Dimethylamino-cin-but-pyCy-3-glc | 14.78 ± 0.02 | 59.69 ± 0.01 |
| Mv-3-glc | 20.96 ± 0.01 | n.d. |
| CarboxypyMv-3-glc | 13.92 ± 0.04 | 67.85 ± 0.04 |
| Red wine carboxypyranoanthocyanin extract | 21.60 ± 0.02 | 67.10 ± 0.02 |
| MethylpyMv-3-glc | 19.24 ± 0.01 | 56.85 ± 0.06 |
| VinylpyMv-3-glc-catechin | 8.35 ± 0.06 | n.d. |
| - | 41.01 ± 0.38 | |
| Oxybenzone | 69.03 ± 0.17 | |
| Hyaluronidase | Collagenase | Elastase | |
|---|---|---|---|
| Compound | % Inhibition (50 μM) | ||
| Lut | 40.1 ± 2.91 | 24.2 ± 2.95 | 27.1 ± 2.71 |
| Cy-3-glc | 31.7 ± 4.18 | 28.5 ± 2.69 | 13.4 ± 2.57 |
| CarboxypyCy-3-glc | 38.1 ± 3.56 | 40.4 ± 2.59 | 5.45 ± 1.81 |
| MethylpyCy-3-glc | 17.6 ± 1.67 | 7.33 ± 2.47 | 23.7 ± 1.87 |
| DeoxyMv | 21.8 ± 2.59 | n.a. | n.a. |
| Mv-3-glc | 40.8 ± 1.33 | n.a. | 5.01 ± 3.16 |
| CarboxypyMv-3-glc | 28.1 ± 2.67 | 40.5 ± 4.31 | 3.54 ± 5.51 |
| MethylpyMv-3-glc | 1.92 ± 0.467 | 8.04 ± 4.48 | 5.65 ± 5.37 |
| Quercetin | 77.6 ± 2.28 | n.d. | n.d. |
| EGCG | n.d. | 86.7 ± 5.31 | n.d. |
| MAAPVCK | n.d. | n.d. | 94.6 ± 0.759 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, P.; Araújo, P.; Ribeiro, C.; Oliveira, H.; Pereira, A.R.; Mateus, N.; de Freitas, V.; Brás, N.F.; Gameiro, P.; Coelho, P.; et al. Anthocyanin-Related Pigments: Natural Allies for Skin Health Maintenance and Protection. Antioxidants 2021, 10, 1038. https://doi.org/10.3390/antiox10071038
Correia P, Araújo P, Ribeiro C, Oliveira H, Pereira AR, Mateus N, de Freitas V, Brás NF, Gameiro P, Coelho P, et al. Anthocyanin-Related Pigments: Natural Allies for Skin Health Maintenance and Protection. Antioxidants. 2021; 10(7):1038. https://doi.org/10.3390/antiox10071038
Chicago/Turabian StyleCorreia, Patrícia, Paula Araújo, Carolina Ribeiro, Hélder Oliveira, Ana Rita Pereira, Nuno Mateus, Victor de Freitas, Natércia F. Brás, Paula Gameiro, Patrícia Coelho, and et al. 2021. "Anthocyanin-Related Pigments: Natural Allies for Skin Health Maintenance and Protection" Antioxidants 10, no. 7: 1038. https://doi.org/10.3390/antiox10071038
APA StyleCorreia, P., Araújo, P., Ribeiro, C., Oliveira, H., Pereira, A. R., Mateus, N., de Freitas, V., Brás, N. F., Gameiro, P., Coelho, P., Bessa, L. J., Oliveira, J., & Fernandes, I. (2021). Anthocyanin-Related Pigments: Natural Allies for Skin Health Maintenance and Protection. Antioxidants, 10(7), 1038. https://doi.org/10.3390/antiox10071038