
Antioxidant Properties of Ergosterol and Its Role in Yeast Resistance to Oxidation

 ,
,  , , , , ,
, , , , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Sterol and Fatty Acid Analysis of the Different Strains of Yeast
2.3. Assessment of Oxidation Effects on Yeast Viability and Plasma Membrane Integrity
2.4. Preparation of Liposomes from Natural Yeast Lipid Extracts
2.5. Assessment of Liposome Fluidity by Fluorescence Spectroscopy
2.6. Oxidation of Liposome Suspensions and Assessment of Lipid Peroxidation
2.7. Assessment Hydrogen Atom-Donating Ability of Sterols by the DPPH Method
2.8. Computational Analysis of Antioxidant Properties of Sterols
3. Results
3.1. Sterol and Fatty Acid Composition of the Different Yeast Strains
3.2. Wild-Type (WT) Yeasts Are More Resistant to Oxidative Treatment Than erg6Δ and erg2Δerg6Δ Strains
3.3. Sterols Protect Lipids from Oxidation in Liposomes
3.4. Ergosterol Displays Greater 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical-Scavenging Activity Than Zymosterol
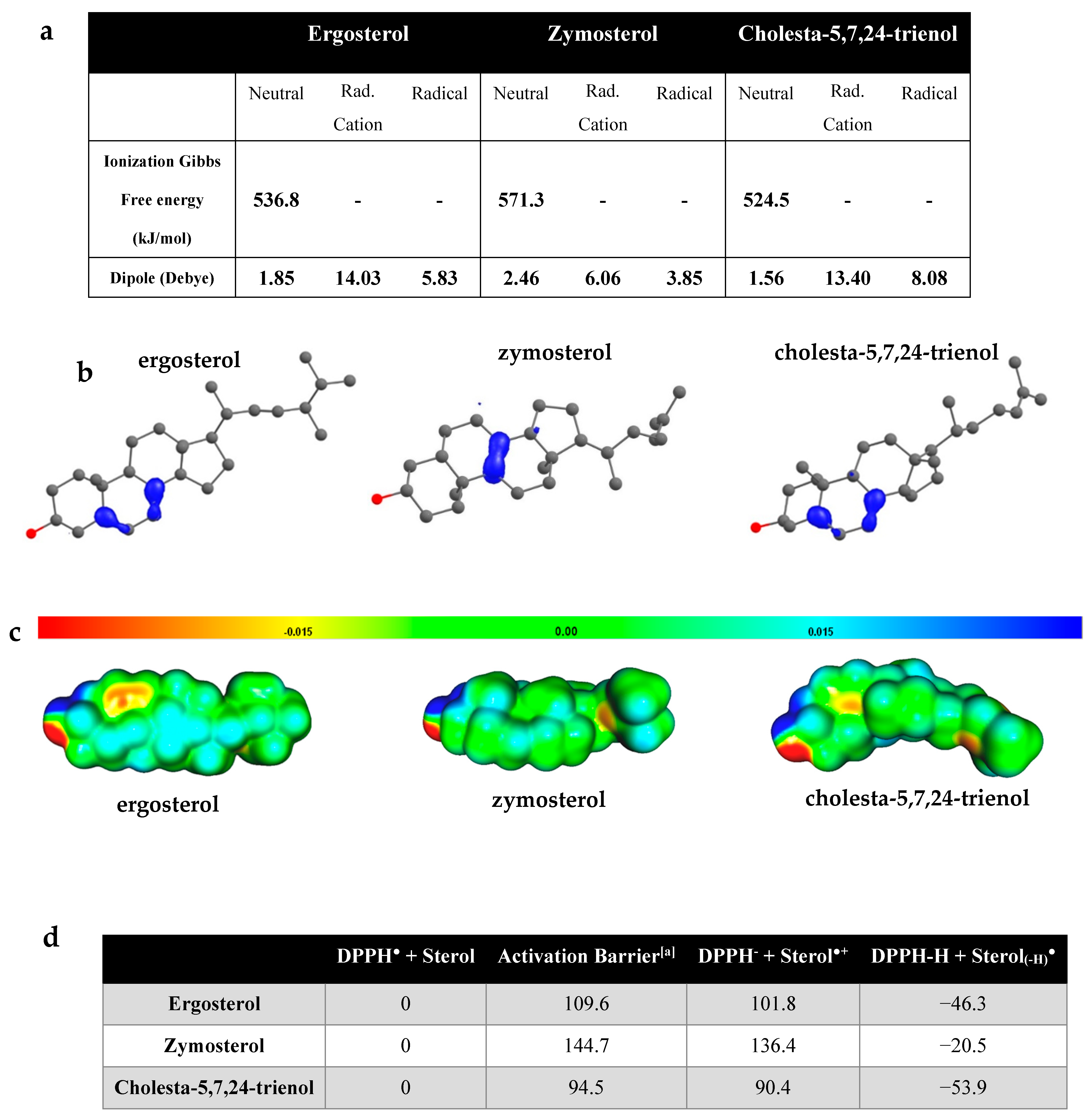
3.5. Computational Analysis of Antioxidant Properties of Sterols
- (1)
- Hydrogen Atom Transfer (HAT)
- (2)
- Electron Transfer (ET)
- (3)
- Electron Transfer followed by Proton Transfer (ET-PT)
- (4)
- Radical Adduct Formation (RAF)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
 ) and erg6Δ (
) and erg6Δ (  ) strains of S. cerevisiae are grown for 48 h at 25 °C in a yeast extract–peptone–dextrose (YPD) medium supplemented with various concentrations of t-BOOH (0, 0.1, 0.5, or 1 mM). Cells were spotted at different ten-fold dilutions (from 10−1 to 10−4) from the initial suspension at OD600 nm = 0.5. Data are presented as mean values ± standard deviation of three independent experiments.
) and erg6Δ ( ) strains of S. cerevisiae are grown for 48 h at 25 °C in a yeast extract–peptone–dextrose (YPD) medium supplemented with various concentrations of t-BOOH (0, 0.1, 0.5, or 1 mM). Cells were spotted at different ten-fold dilutions (from 10−1 to 10−4) from the initial suspension at OD600 nm = 0.5. Data are presented as mean values ± standard deviation of three independent experiments.
) strains of S. cerevisiae are grown for 48 h at 25 °C in a yeast extract–peptone–dextrose (YPD) medium supplemented with various concentrations of t-BOOH (0, 0.1, 0.5, or 1 mM). Cells were spotted at different ten-fold dilutions (from 10−1 to 10−4) from the initial suspension at OD600 nm = 0.5. Data are presented as mean values ± standard deviation of three independent experiments.
) and erg6Δ ( ) strains of S. cerevisiae are grown for 48 h at 25 °C in a yeast extract–peptone–dextrose (YPD) medium supplemented with various concentrations of t-BOOH (0, 0.1, 0.5, or 1 mM). Cells were spotted at different ten-fold dilutions (from 10−1 to 10−4) from the initial suspension at OD600 nm = 0.5. Data are presented as mean values ± standard deviation of three independent experiments.
Appendix C
Appendix C.1. Estimation of the Electron Transfer Activation Free Energy
Appendix C.2. Bond Dissociation Energies (kJ/mol) for Several H Atoms

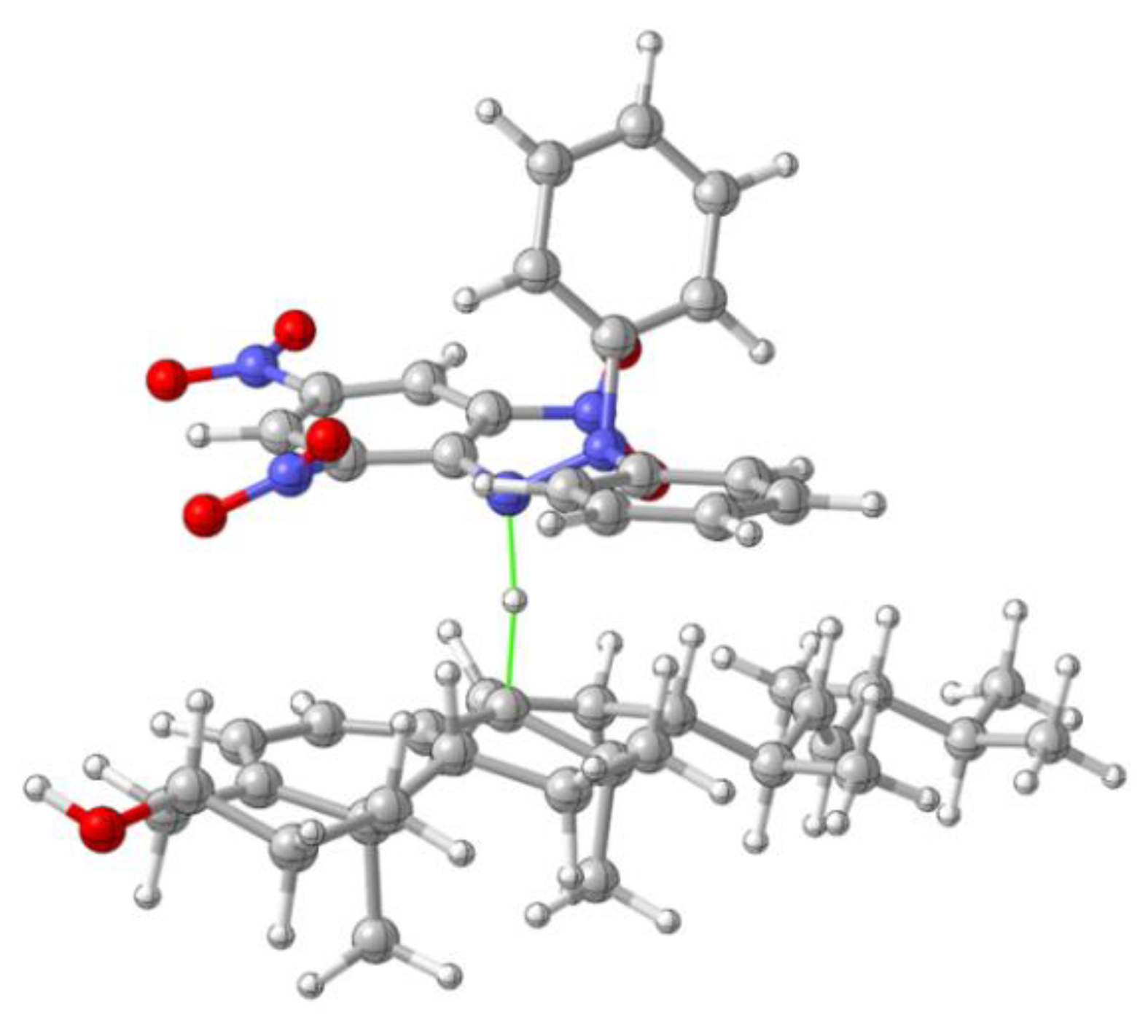
Appendix C.3. Transition State for the H Atom Tranfer from Ergosterol Radical Cation to DPPH−
References
- Nes, W.D. Biosynthesis of Cholesterol and Other Sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef]
- Tanret, C. Sur Un Nouveau Principle Immediate de l’ergot Deseigle Ergosterine. CR Seances Acad. Sci. 1889, 108, 98–100. [Google Scholar]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane Lipids: Where They Are and How They Behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Gray, K.C.; Palacios, D.S.; Dailey, I.; Endo, M.M.; Uno, B.E.; Wilcock, B.C.; Burke, M.D. Amphotericin Primarily Kills Yeast by Simply Binding Ergosterol. Proc. Natl. Acad. Sci. USA 2012, 109, 2234–2239. [Google Scholar] [CrossRef] [Green Version]
- Mercer, E.I. Inhibitors of Sterol Biosynthesis and Their Applications. Prog. Lipid Res. 1993, 32, 357–416. [Google Scholar] [CrossRef]
- Jordá, T.; Puig, S. Regulation of Ergosterol Biosynthesis in Saccharomyces Cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef]
- Parks, L.W.; Casey, W.M. Physiological Implications of Sterol Biosynthesis in Yeast. Annu. Rev. Microbiol. 1995, 49, 95–116. [Google Scholar] [CrossRef]
- Weete, J.D.; Abril, M.; Blackwell, M. Phylogenetic Distribution of Fungal Sterols. PLoS ONE 2010, 5, e10899. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Lemetais, G.; Ferreira, T.; Cayot, P.; Gervais, P.; Beney, L. Ergosterol Biosynthesis: A Fungal Pathway for Life on Land? Evol. Int. J. Org. Evol. 2012, 66, 2961–2968. [Google Scholar] [CrossRef]
- Dupont, S.; Rapoport, A.; Gervais, P.; Beney, L. Survival Kit of Saccharomyces Cerevisiae for Anhydrobiosis. Appl. Microbiol. Biotechnol. 2014, 98, 8821–8834. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Beney, L.; Ritt, J.-F.; Lherminier, J.; Gervais, P. Lateral Reorganization of Plasma Membrane Is Involved in the Yeast Resistance to Severe Dehydration. Biochim. Biophys. Acta BBA-Biomembr. 2010, 1798, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Leslie, S.B.; Teter, S.A.; Crowe, L.M.; Crowe, J.H. Trehalose Lowers Membrane Phase Transitions in Dry Yeast Cells. Biochim. Biophys. Acta BBA Biomembr. 1994, 1192, 7–13. [Google Scholar] [CrossRef]
- Rapoport, A.; Golovina, E.A.; Gervais, P.; Dupont, S.; Beney, L. Anhydrobiosis: Inside Yeast Cells. Biotechnol. Adv. 2019, 37, 51–67. [Google Scholar] [CrossRef]
- Garre, E.; Raginel, F.; Palacios, A.; Julien, A.; Matallana, E. Oxidative Stress Responses and Lipid Peroxidation Damage Are Induced during Dehydration in the Production of Dry Active Wine Yeasts. Int. J. Food Microbiol. 2010, 136, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jesus Pereira, E.; Panek, A.D.; Eleutherio, E.C.A. Protection against Oxidation during Dehydration of Yeast. Cell Stress Chaperones 2003, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Piette, J. New Trends in Photobiology: Biological Consequences Associated with DNA Oxidation Mediated by Singlet Oxygen. J. Photochem. Photobiol. B 1991, 11, 241–260. [Google Scholar] [CrossRef]
- Dupont, S.; Beney, L.; Ferreira, T.; Gervais, P. Nature of Sterols Affects Plasma Membrane Behavior and Yeast Survival during Dehydration. Biochim. Biophys. Acta BBA-Biomembr. 2011, 1808, 1520–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heleno, S.A.; Rudke, A.R.; Calhelha, R.C.; Carocho, M.; Barros, L.; Gonçalves, O.H.; Barreiro, M.F.; Ferreira, I.C. Development of Dairy Beverages Functionalized with Pure Ergosterol and Mycosterol Extracts: An Alternative to Phytosterol-Based Beverages. Food Funct. 2017, 8, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Hernandez, M.; Kramer, J.K.; Rinker, D.L.; Tsao, R. Ergosterol Profiles, Fatty Acid Composition, and Antioxidant Activities of Button Mushrooms as Affected by Tissue Part and Developmental Stage. J. Agric. Food Chem. 2010, 58, 11616–11625. [Google Scholar] [CrossRef] [PubMed]
- Munn, A.L.; Heese-Peck, A.; Stevenson, B.J.; Pichler, H.; Riezman, H. Specific Sterols Required for the Internalization Step of Endocytosis in Yeast. Mol. Biol. Cell 1999, 10, 3943–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa-Lopes, A.; Antunes, F.; Cyrne, L.; Marinho, H.S. Decreased Cellular Permeability to H2O2 Protects Saccharomyces Cerevisiae Cells in Stationary Phase against Oxidative Stress. FEBS Lett. 2004, 578, 152–156. [Google Scholar] [CrossRef] [Green Version]
- Biryukova, E.N.; Medentsev, A.G.; Arinbasarova, A.Y.; Akimenko, V.K. Tolerance of the Yeast Yarrowia Lipolytica to Oxidative Stress. Microbiology 2006, 75, 243–247. [Google Scholar] [CrossRef]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of Univalent Ions across the Lamellae of Swollen Phospholipids. J. Mol. Biol. 1965, 13, 238-IN27. [Google Scholar] [CrossRef]
- Xie, J.; Schaich, K.M. Re-Evaluation of the 2, 2-Diphenyl-1-Picrylhydrazyl Free Radical (DPPH) Assay for Antioxidant Activity. J. Agric. Food Chem. 2014, 62, 4251–4260. [Google Scholar] [CrossRef]
- Fadda, A.; Serra, M.; Molinu, M.G.; Azara, E.; Barberis, A.; Sanna, D. Reaction Time and DPPH Concentration Influence Antioxidant Activity and Kinetic Parameters of Bioactive Molecules and Plant Extracts in the Reaction with the DPPH Radical. J. Food Compos. Anal. 2014, 35, 112–119. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A Procedure to Measure the Antiradical Efficiency of Polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Boissy, G.; O’Donohue, M.; Gaudemer, O.; Perez, V.; Pernollet, J.-C.; Brunie, S. The 2.1 \AA Structure of an Elicitin-Ergosterol Complex: A Recent Addition to the Sterol Carrier Protein Family. Protein Sci. 1999, 8, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Tomasi, J.; Mennucci, B.; Cammi, R. Quantum Mechanical Continuum Solvation Models. Chem. Rev. 2005, 105, 2999–3094. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H. Gaussian 16 Revision B. 01; Gaussian Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Andrienko, G.A. Chemcraft. Graphical Software for Visualization of Quantum Chemistry Computations. 2010. Available online: http://www.chemcraftprog.com (accessed on 12 May 2021).
- Allouche, A.-R. Gabedit—A Graphical User Interface for Computational Chemistry Softwares. J. Comput. Chem. 2011, 32, 174–182. [Google Scholar] [CrossRef]
- Heese-Peck, A.; Pichler, H.; Zanolari, B.; Watanabe, R.; Daum, G.; Riezman, H. Multiple Functions of Sterols in Yeast Endocytosis. Mol. Biol. Cell 2002, 13, 2664–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummen, G.P.; van Liebergen, L.C.; den Kamp, J.A.O.; Post, J.A. C11-BODIPY581/591, an Oxidation-Sensitive Fluorescent Lipid Peroxidation Probe:(Micro) Spectroscopic Characterization and Validation of Methodology. Free Radic. Biol. Med. 2002, 33, 473–490. [Google Scholar] [CrossRef]
- Medina, M.E.; Iuga, C.; Trigos, Á. Mechanism and Kinetics of the Oxidative Damage to Ergosterol Induced by Peroxyl Radicals in Lipid Media: A Theoretical Quantum Chemistry Study. J. Phys. Org. Chem. 2016, 29, 196–203. [Google Scholar] [CrossRef]
- Abe, F.; Hiraki, T. Mechanistic Role of Ergosterol in Membrane Rigidity and Cycloheximide Resistance in Saccharomyces Cerevisiae. Biochim. Biophys. Acta BBA-Biomembr. 2009, 1788, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Ischer, F.; Parkinson, T.; Falconer, D.; Bille, J. Candida Albicans Mutations in the Ergosterol Biosynthetic Pathway and Resistance to Several Antifungal Agents. Antimicrob. Agents Chemother. 2003, 47, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Yu, B.P. Alterations in Mitochondrial Membrane Fluidity by Lipid Peroxidation Products. Free Radic. Biol. Med. 1994, 17, 411–418. [Google Scholar] [CrossRef]
- Itri, R.; Junqueira, H.C.; Mertins, O.; Baptista, M.S. Membrane Changes under Oxidative Stress: The Impact of Oxidized Lipids. Biophys. Rev. 2014, 6, 47–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, R.P.; Walter, M.F.; Mason, P.E. Effect of Oxidative Stress on Membrane Structure: Small-Angle X-Ray Diffraction Analysis. Free Radic. Biol. Med. 1997, 23, 419–425. [Google Scholar] [CrossRef]
- Mertins, O.; Bacellar, I.O.; Thalmann, F.; Marques, C.M.; Baptista, M.S.; Itri, R. Physical Damage on Giant Vesicles Membrane as a Result of Methylene Blue Photoirradiation. Biophys. J. 2014, 106, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Van der Paal, J.; Neyts, E.C.; Verlackt, C.C.; Bogaerts, A. Effect of Lipid Peroxidation on Membrane Permeability of Cancer and Normal Cells Subjected to Oxidative Stress. Chem. Sci. 2016, 7, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasowska, A.; Chmielewska, L.; Prescha, A.; Váchová, L.; Sigler, K. Viability and Formation of Conjugated Dienes in Plasma Membrane Lipids OfSaccharomyces Cerevisiae, Schizosaccharomyces Pombe, Rhodotorula Glutinis AndCandida Albicans Exposed to Hydrophilic, Amphiphilic and Hydrophobic pro-Oxidants. Folia Microbiol. 2002, 47, 145–151. [Google Scholar] [CrossRef]
- Megli, F.M.; Conte, E.; Ishikawa, T. Cholesterol Attenuates and Prevents Bilayer Damage and Breakdown in Lipoperoxidized Model Membranes. A Spin Labeling EPR Study. Biochim. Biophys. Acta BBA-Biomembr. 2011, 1808, 2267–2274. [Google Scholar] [CrossRef] [Green Version]
- Luu, W.; Sharpe, L.J.; Capell-Hattam, I.; Gelissen, I.C.; Brown, A.J. Oxysterols: Old Tale, New Twists. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Böcking, T.; Barrow, K.D.; Netting, A.G.; Chilcott, T.C.; Coster, H.G.; Höfer, M. Effects of Singlet Oxygen on Membrane Sterols in the Yeast Saccharomyces Cerevisiae. Eur. J. Biochem. 2000, 267, 1607–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casey, W.M.; Parks, L.W. A Role for Sterols in the Porphyrin Mediated Photosensitization of Yeast. Photochem. Photobiol. 1989, 50, 553–556. [Google Scholar] [CrossRef]
- Gordon, M.H.; Magos, P. The Effect of Sterols on the Oxidation of Edible Oils. Food Chem. 1983, 10, 141–147. [Google Scholar] [CrossRef]
- Wiseman, H. Vitamin D Is a Membrane Antioxidant Ability to Inhibit Iron-Dependent Lipid Peroxidation in Liposomes Compared to Cholesterol, Ergosterol and Tamoxifen and Relevance to Anticancer Action. FEBS Lett. 1993, 326, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.L. Another Cholesterol Hypothesis: Cholesterol as Antioxidant. Free Radic. Biol. Med. 1991, 11, 47–61. [Google Scholar] [CrossRef]
- Jamieson, D.J. Oxidative Stress Responses of the Yeast Saccharomyces Cerevisiae. Yeast 1998, 14, 1511–1527. [Google Scholar] [CrossRef]
- Gazdag, Z.; Máté, G.; Čertik, M.; Türmer, K.; Virág, E.; Pócsi, I.; Pesti, M. Tert-Butyl Hydroperoxide-Induced Differing Plasma Membrane and Oxidative Stress Processes in Yeast Strains BY4741 and Erg5Δ. J. Basic Microbiol. 2014, 54, S50–S62. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, N.E.; de Aguiar, I.; de Zawadzki, A.; Cardoso, D.R. Kinetics and Thermodynamics of 1-Hydroxyethyl Radical Reaction with Unsaturated Lipids and Prenylflavonoids. J. Phys. Chem. B 2014, 118, 14278–14287. [Google Scholar] [CrossRef]
- Shrivastava, S.; Chattopadhyay, A. Influence of Cholesterol and Ergosterol on Membrane Dynamics Using Different Fluorescent Reporter Probes. Biochem. Biophys. Res. Commun. 2007, 356, 705–710. [Google Scholar] [CrossRef]
- Khmelinskaia, A.; Marquês, J.M.T.; Bastos, A.E.P.; Antunes, C.A.C.; Bento-Oliveira, A.; Scolari, S.; Lobo, G.M.d.S.; Malhó, R.; Herrmann, A.; Marinho, H.S.; et al. Liquid-Ordered Phase Formation by Mammalian and Yeast Sterols: A Common Feature With Organizational Differences. Front. Cell Dev. Biol. 2020, 8, 337. [Google Scholar] [CrossRef]
- Brown, A.J.; Galea, A.M. Cholesterol as an Evolutionary Response to Living with Oxygen. Evol. Int. J. Org. Evol. 2010, 64, 2179–2183. [Google Scholar] [CrossRef]
- Galea, A.M.; Brown, A.J. Special Relationship between Sterols and Oxygen: Were Sterols an Adaptation to Aerobic Life? Free Radic. Biol. Med. 2009, 47, 880–889. [Google Scholar] [CrossRef]
- Rajendran, L.; Simons, K. Lipid Rafts and Membrane Dynamics. J. Cell Sci. 2005, 118, 1099–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossmann, G.; Opekarova, M.; Malinsky, J.; Weig-Meckl, I.; Tanner, W. Membrane Potential Governs Lateral Segregation of Plasma Membrane Proteins and Lipids in Yeast. EMBO J. 2007, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aresta-Branco, F.; Cordeiro, A.M.; Marinho, H.S.; Cyrne, L.; Antunes, F.; de Almeida, R.F. Gel Domains in the Plasma Membrane of Saccharomyces Cerevisiae Highly Ordered, Ergosterol-Free, and Sphingolipid-Enriched Lipid Rafts. J. Biol. Chem. 2011, 286, 5043–5054. [Google Scholar] [CrossRef] [Green Version]
- Marcus, R.A. Electron Transfer Reactions in Chemistry. Theory and Experiment. Rev. Mod. Phys. 1993, 65, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Nelsen, S.F.; Blackstock, S.C.; Kim, Y. Estimation of Inner Shell Marcus Terms for Amino Nitrogen Compounds by Molecular Orbital Calculations. J. Am. Chem. Soc. 1987, 109, 677–682. [Google Scholar] [CrossRef]
 ), RH448 WT (
), RH448 WT (  ), erg6Δ (
), erg6Δ (  ), and erg2Δerg6Δ (
), and erg2Δerg6Δ (  ) strains of S. cerevisiae. (a) Analysis of the sterol composition and the relative abundance. (b) Analysis of the fatty acid composition and the relative abundance. Vertical bars represent standard deviation.
), RH448 WT ( ), erg6Δ ( ), and erg2Δerg6Δ ( ) strains of S. cerevisiae. (a) Analysis of the sterol composition and the relative abundance. (b) Analysis of the fatty acid composition and the relative abundance. Vertical bars represent standard deviation.
) strains of S. cerevisiae. (a) Analysis of the sterol composition and the relative abundance. (b) Analysis of the fatty acid composition and the relative abundance. Vertical bars represent standard deviation.
), RH448 WT ( ), erg6Δ ( ), and erg2Δerg6Δ ( ) strains of S. cerevisiae. (a) Analysis of the sterol composition and the relative abundance. (b) Analysis of the fatty acid composition and the relative abundance. Vertical bars represent standard deviation.
 ), RH448 WT (
), RH448 WT (  ), erg6Δ (
), erg6Δ (  ), and erg2Δerg6Δ (
), and erg2Δerg6Δ (  ) strains of S. cerevisiae were washed, adjusted to a concentration corresponding to DO600 nm = 0.5, and placed in PBS containing t-BOOH (4 or 6 mM) for 1 or 2 h. (a) After treatment, the cells were washed and spotted at different ten-fold dilutions (from 10−1 to 10−4) to assess the cell viability. (b) Plasma membrane integrity was assessed by PI staining of cells and flow cytometry analysis. Data are presented as mean values ± standard deviation of three independent experiments. ANOVA was performed on R v3.6.1 software and if it was significant (p < 0.01), Tukey’s HSD (Honest Significant Difference) test was performed to observe significant differences among conditions. Letters a, b, c, d, and e represent significantly different groups.
), RH448 WT ( ), erg6Δ ( ), and erg2Δerg6Δ ( ) strains of S. cerevisiae were washed, adjusted to a concentration corresponding to DO600 nm = 0.5, and placed in PBS containing t-BOOH (4 or 6 mM) for 1 or 2 h. (a) After treatment, the cells were washed and spotted at different ten-fold dilutions (from 10−1 to 10−4) to assess the cell viability. (b) Plasma membrane integrity was assessed by PI staining of cells and flow cytometry analysis. Data are presented as mean values ± standard deviation of three independent experiments. ANOVA was performed on R v3.6.1 software and if it was significant (p < 0.01), Tukey’s HSD (Honest Significant Difference) test was performed to observe significant differences among conditions. Letters a, b, c, d, and e represent significantly different groups.
) strains of S. cerevisiae were washed, adjusted to a concentration corresponding to DO600 nm = 0.5, and placed in PBS containing t-BOOH (4 or 6 mM) for 1 or 2 h. (a) After treatment, the cells were washed and spotted at different ten-fold dilutions (from 10−1 to 10−4) to assess the cell viability. (b) Plasma membrane integrity was assessed by PI staining of cells and flow cytometry analysis. Data are presented as mean values ± standard deviation of three independent experiments. ANOVA was performed on R v3.6.1 software and if it was significant (p < 0.01), Tukey’s HSD (Honest Significant Difference) test was performed to observe significant differences among conditions. Letters a, b, c, d, and e represent significantly different groups.
), RH448 WT ( ), erg6Δ ( ), and erg2Δerg6Δ ( ) strains of S. cerevisiae were washed, adjusted to a concentration corresponding to DO600 nm = 0.5, and placed in PBS containing t-BOOH (4 or 6 mM) for 1 or 2 h. (a) After treatment, the cells were washed and spotted at different ten-fold dilutions (from 10−1 to 10−4) to assess the cell viability. (b) Plasma membrane integrity was assessed by PI staining of cells and flow cytometry analysis. Data are presented as mean values ± standard deviation of three independent experiments. ANOVA was performed on R v3.6.1 software and if it was significant (p < 0.01), Tukey’s HSD (Honest Significant Difference) test was performed to observe significant differences among conditions. Letters a, b, c, d, and e represent significantly different groups.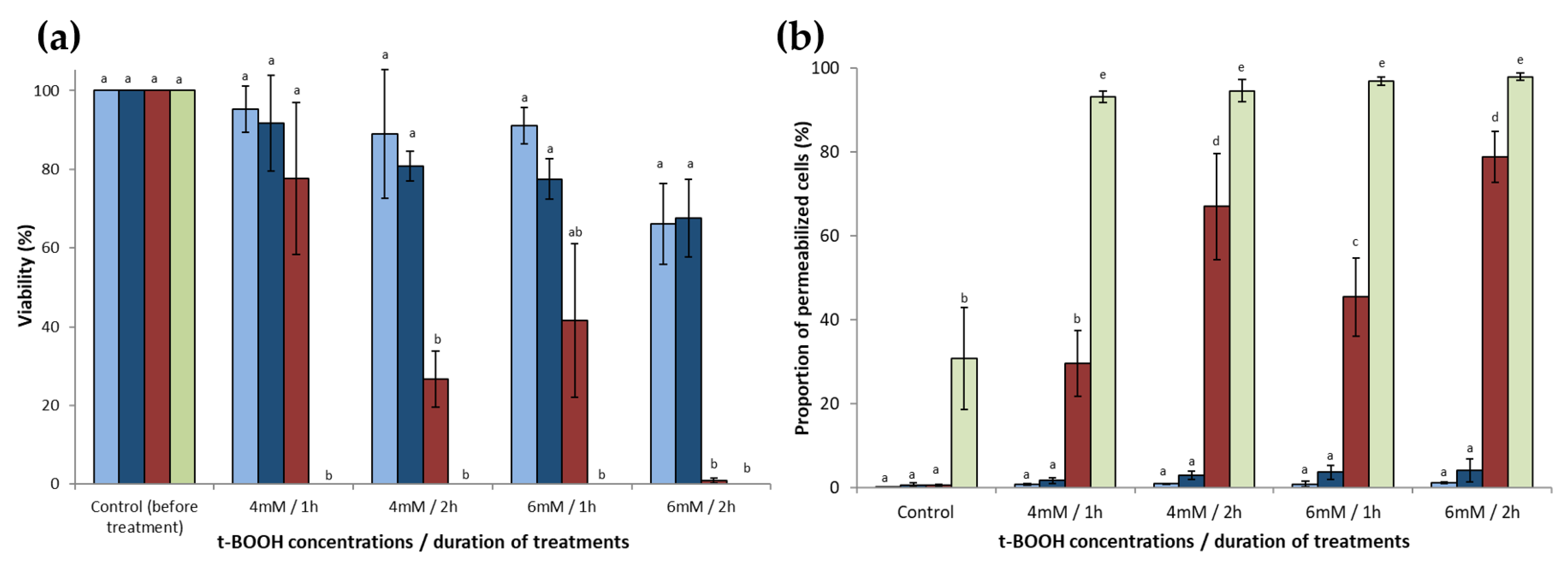
 ), with a zymosterol/phospholipid molar ratio of 1/3 (
), with a zymosterol/phospholipid molar ratio of 1/3 (  ), with a cholesta-5,7,24-trienol/phospholipid molar ratio of 1/3 (
), with a cholesta-5,7,24-trienol/phospholipid molar ratio of 1/3 (  ), or with an ergosterol/phospholipid molar ratio of 1/3 (
), or with an ergosterol/phospholipid molar ratio of 1/3 (  ). Control experiments were performed with lipososmes with a tocopherol/phospholipid molar ratio of 1/3 (
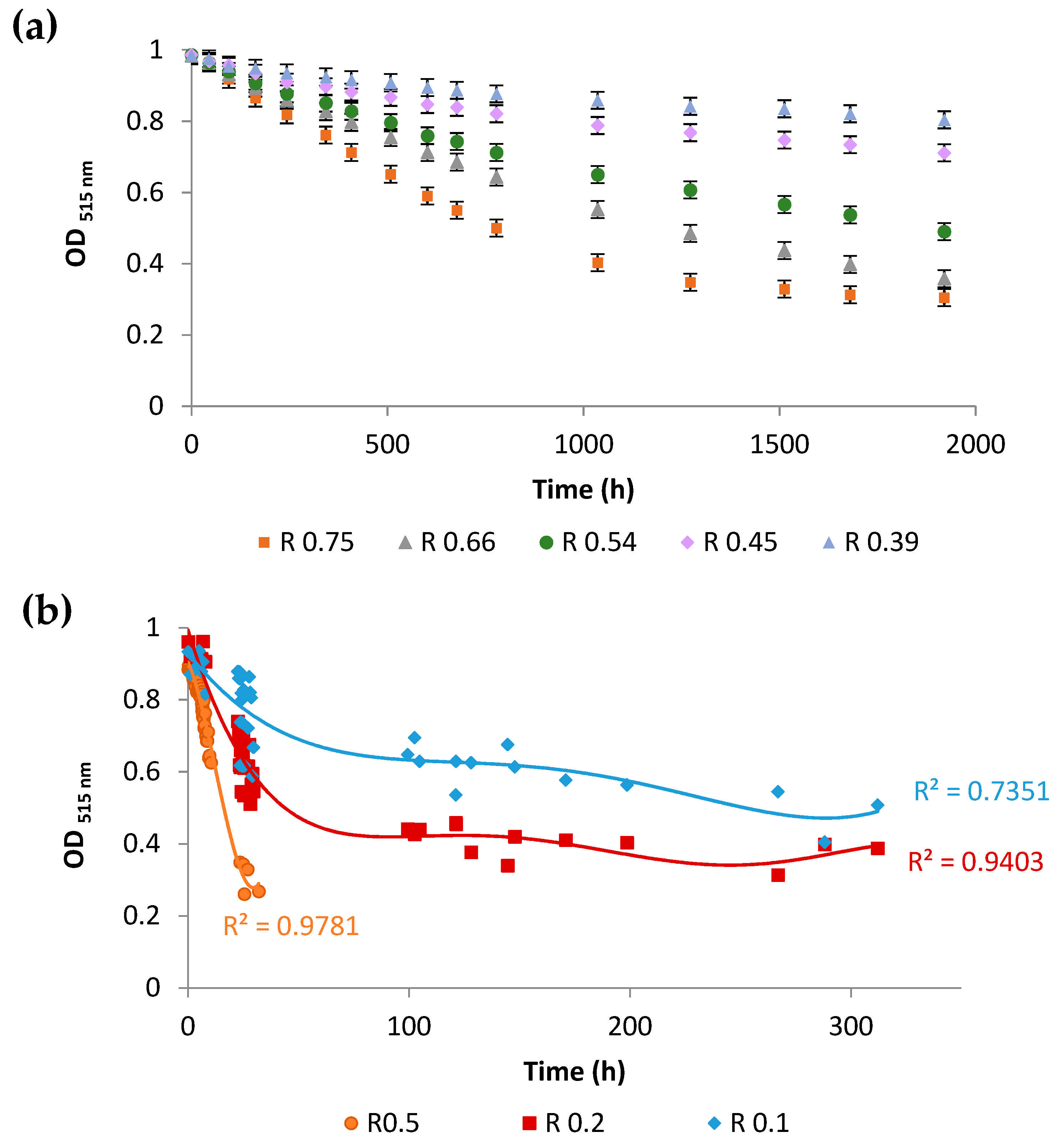
). Control experiments were performed with lipososmes with a tocopherol/phospholipid molar ratio of 1/3 (  ). (a) Phospholipid oxidation induced by cumene hydroperoxide and haemin was assessed by the oxidation kinetics of BP-C11 in liposomes with different sterol compositions. Oxidation was expressed as the intensity ratio (I520 nm)oxidized/(I590 nm)non-oxidized. Oxidation was started at t = 0 s. (b) Fluidity of liposomes was assessed by steady-state DPH fluorescence anisotropy measurement as a function of temperature in the range 4–48 °C. The results are presented with error bars corresponding to the standard deviation calculated from three repeated experiments.
), with a zymosterol/phospholipid molar ratio of 1/3 ( ), with a cholesta-5,7,24-trienol/phospholipid molar ratio of 1/3 ( ), or with an ergosterol/phospholipid molar ratio of 1/3 ( ). Control experiments were performed with lipososmes with a tocopherol/phospholipid molar ratio of 1/3 ( ). (a) Phospholipid oxidation induced by cumene hydroperoxide and haemin was assessed by the oxidation kinetics of BP-C11 in liposomes with different sterol compositions. Oxidation was expressed as the intensity ratio (I520 nm)oxidized/(I590 nm)non-oxidized. Oxidation was started at t = 0 s. (b) Fluidity of liposomes was assessed by steady-state DPH fluorescence anisotropy measurement as a function of temperature in the range 4–48 °C. The results are presented with error bars corresponding to the standard deviation calculated from three repeated experiments.
). (a) Phospholipid oxidation induced by cumene hydroperoxide and haemin was assessed by the oxidation kinetics of BP-C11 in liposomes with different sterol compositions. Oxidation was expressed as the intensity ratio (I520 nm)oxidized/(I590 nm)non-oxidized. Oxidation was started at t = 0 s. (b) Fluidity of liposomes was assessed by steady-state DPH fluorescence anisotropy measurement as a function of temperature in the range 4–48 °C. The results are presented with error bars corresponding to the standard deviation calculated from three repeated experiments.
), with a zymosterol/phospholipid molar ratio of 1/3 ( ), with a cholesta-5,7,24-trienol/phospholipid molar ratio of 1/3 ( ), or with an ergosterol/phospholipid molar ratio of 1/3 ( ). Control experiments were performed with lipososmes with a tocopherol/phospholipid molar ratio of 1/3 ( ). (a) Phospholipid oxidation induced by cumene hydroperoxide and haemin was assessed by the oxidation kinetics of BP-C11 in liposomes with different sterol compositions. Oxidation was expressed as the intensity ratio (I520 nm)oxidized/(I590 nm)non-oxidized. Oxidation was started at t = 0 s. (b) Fluidity of liposomes was assessed by steady-state DPH fluorescence anisotropy measurement as a function of temperature in the range 4–48 °C. The results are presented with error bars corresponding to the standard deviation calculated from three repeated experiments.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dupont, S.; Fleurat-Lessard, P.; Cruz, R.G.; Lafarge, C.; Grangeteau, C.; Yahou, F.; Gerbeau-Pissot, P.; Abrahão Júnior, O.; Gervais, P.; Simon-Plas, F.; et al. Antioxidant Properties of Ergosterol and Its Role in Yeast Resistance to Oxidation. Antioxidants 2021, 10, 1024. https://doi.org/10.3390/antiox10071024
Dupont S, Fleurat-Lessard P, Cruz RG, Lafarge C, Grangeteau C, Yahou F, Gerbeau-Pissot P, Abrahão Júnior O, Gervais P, Simon-Plas F, et al. Antioxidant Properties of Ergosterol and Its Role in Yeast Resistance to Oxidation. Antioxidants. 2021; 10(7):1024. https://doi.org/10.3390/antiox10071024
Chicago/Turabian StyleDupont, Sebastien, Paul Fleurat-Lessard, Richtier Gonçalves Cruz, Céline Lafarge, Cédric Grangeteau, Fairouz Yahou, Patricia Gerbeau-Pissot, Odonírio Abrahão Júnior, Patrick Gervais, Françoise Simon-Plas, and et al. 2021. "Antioxidant Properties of Ergosterol and Its Role in Yeast Resistance to Oxidation" Antioxidants 10, no. 7: 1024. https://doi.org/10.3390/antiox10071024
APA StyleDupont, S., Fleurat-Lessard, P., Cruz, R. G., Lafarge, C., Grangeteau, C., Yahou, F., Gerbeau-Pissot, P., Abrahão Júnior, O., Gervais, P., Simon-Plas, F., Cayot, P., & Beney, L. (2021). Antioxidant Properties of Ergosterol and Its Role in Yeast Resistance to Oxidation. Antioxidants, 10(7), 1024. https://doi.org/10.3390/antiox10071024